







-
Apple (Malus × domestica Borkh.) ranks as a top fruit in both production and consumption globally. In 2022, the global production of apples reached 95.8 million tons, with China leading the output at 49.60%, followed by the US (5.02%) and Poland (4.62%)[1]. Globally, over 7,500 apple varieties have been identified, many tracing their lineage to widely adaptable genotypes such as 'Gala' and 'Golden Delicious'. In China, Shanxi province is recognized as one of China's key apple-growing zones. This province has consistently contributed 10% of China's total apple production[2]. Apples are a nutrient-dense fruit, abundant in health-enhancing bioactive compounds. Key constituents such as sugars, organic acids, and phenolic compounds play a pivotal role in shaping both the sensory attributes and the nutritional value of apples, significantly influencing their overall quality[3,4] .
Metabolomics, which involves the comprehensive quantification and identification of metabolites within a biological framework, has become a critical technology for deciphering the biochemical signature of organisms. Applying nuclear magnetic resonance (NMR) metabolomics integrated with multivariate analytical techniques in the analysis of apples opens a unique avenue to evaluate apple quality, identify specific metabolites, and distinguish apples from various cultivars and geographical origins[5]. NMR metabolomics study has shown that apples from Japan ('Fuji', 'Orin', and 'Jonagold') had higher concentrations of L-rhamnitol compared to apples from New Zealand ('Fuji', 'Jazz', and 'Envy'). In addition, levels of sucrose, glucose, xylose, quinic acid, and aspartic acid were significantly different between the apple cultivars from Japan and New Zealand[5]. Another study has shown amino acids leucine, isoleucine, and valine exhibited higher concentrations in 'Rubens' apples compared to 'Golden Delicious' and 'Braeburn' cultivars, while 'Golden Delicious' apples showed the highest levels of acetaldehyde as revealed by NMR[6]. Our previous research studying apples harvested from China including Shanxi province has shown cultivars 'Qinguan', 'Zhongqiuwang', and 'Nagafu 2' were more intense in ascorbic acid than the other 13 cultivars as revealed by gas chromatography coupled with flame ionization detection (GC-FID), which indicated the potential difference in their metabolite profile and nutritional values[7].
Like any other fruit, apples exhibit a wide range of metabolites mainly depending on their genetic background. The compositional variation caused by genetic diversity is not only crucial for flavor and texture but also for nutritional value and shelf life, important for both consumers and the food industry. It has been postulated that the genetic background e.g. cultivar plays a more significant role in influencing compositional variations compared to other factors such as growth location and storage conditions[8−11]. However, altitude is another critical factor. The metabolic processes in plants are known to be altitude-sensitive, primarily due to changes in atmospheric pressure, temperature, and sunlight intensity. By studying apples grown at different altitudes, it is possible to gain new insights into how these environmental pressures shape the metabolite composition in apple fruits. Research has shown that 'Golden Delicious' apples grown at a lower altitude of 600 m exhibit larger sizes and have higher concentrations of titratable acids and soluble solids compared to those grown at a higher altitude of 1000 m[12]. Similarly, a study has shown that the contents of sugars and vitamins were different in 'Golden Delicious' apples from different altitudes in South Tyrol, Italy[13].
In this study, fruits from 22 apple cultivars were collected across five major apple-growing cities in Shanxi province, China. Although apple production is significant worldwide, detailed data on the biochemical profile of apples from specific regions with large apple production (Shanxi province) remains scarce. Understanding the compositional parameters of various apple varieties and the factors affecting them is crucial for assessing fruit quality and offers valuable insights for breeding, cultivation, and processing practices. The primary goal was to systematically profile the metabolite profiles of these prominent apple resources and to compare the compositions across these cultivars and to investigate the impact of growth locations on the composition and quality of apple fruits.
-
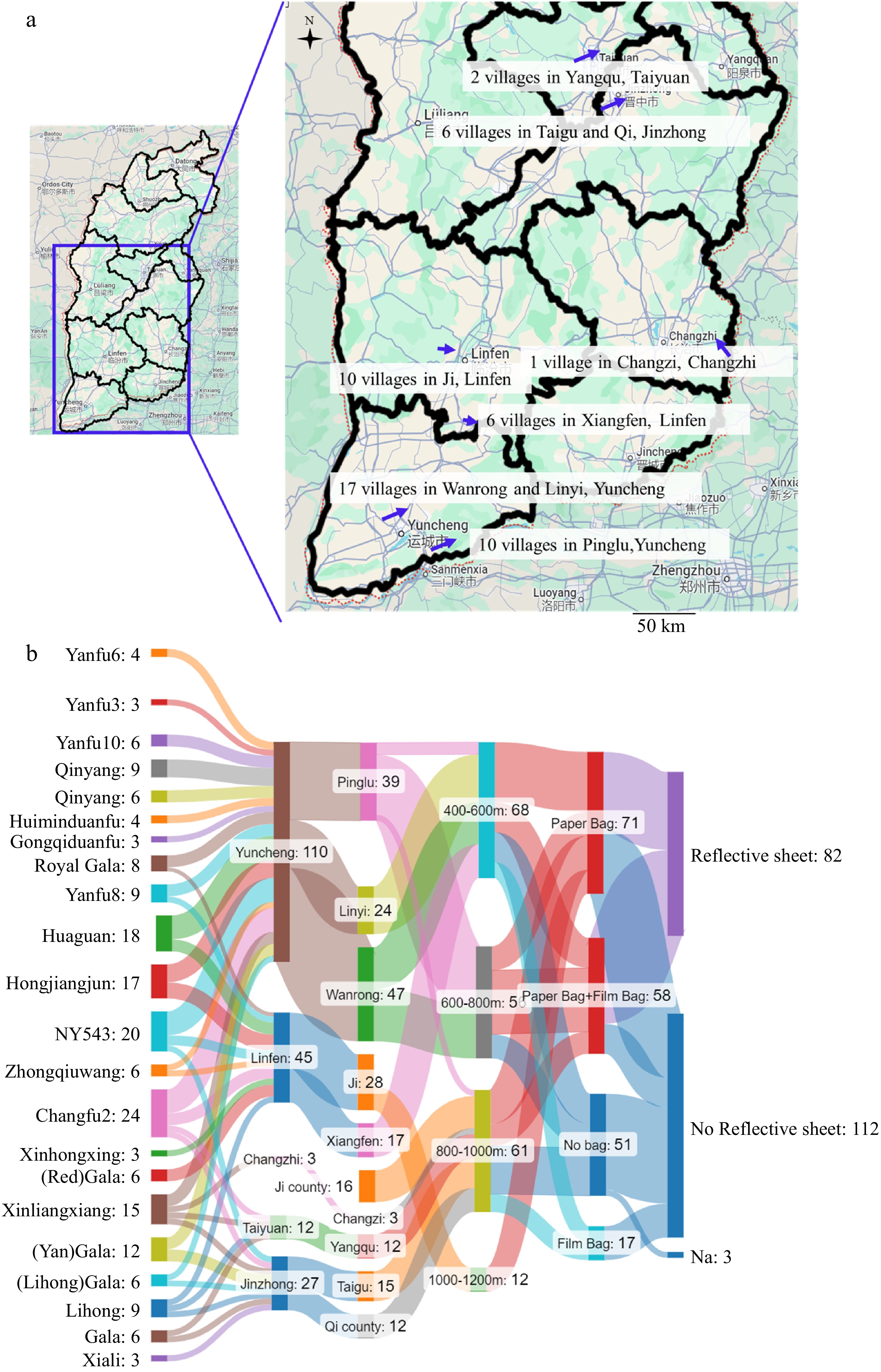
Fresh fruits from 22 different apple cultivars were harvested from nine counties across five cities in the middle and southern parts of Shanxi province, China. In total, 197 apple samples were collected. The most represented cultivars were 'Changfu2' with 24 samples, 'NY543' with 20 samples, and 'Huaguan' with 18 samples. The collection sites varied in altitude, ranging from 0−1,200 m. Moreover, samples were harvested from August to October 2018, Supplementary Table S1 provides comprehensive details about the studied samples including cultivars, bagging method, reflecting sheet usage, altitude, and growth sites. The fruit samples were collected at peak ripeness during the standard commercial harvest period for each cultivar. Approximately 20 fruits per cultivar were selected from a larger batch sourced from multiple trees. These fruits were then pooled and randomly divided into three groups, each serving as a biological replicate (Supplementary Table S1).
The apples were sliced using an apple slicer, and the pieces were immediately frozen with dry ice. They were then ground into a fine powder using a Knife Mill Grindomix GM 300 (Retsch Corp., Haan, Germany) and stored at −30 °C.
Sample preparation for NMR analysis
-
Four hundred mg apple powders were thawed and homogenized for 1 min after adding 800 μL of ice-cold potassium phosphate buffer (90 mM, pH 6.0), maintaining a pulp-to-buffer ratio of 1:2 (w/v). Subsequently, the samples were ultrasonicated for 30 min in a water bath maintained at 4 °C. Following ultrasonication, the samples were centrifuged at 15,000 × g for 15 min at 4 °C. An aliquot of 600 μL of the supernatant was then filtered using a centrifugal filter unit (modified nylon, 0.45 μm, VWR) by centrifugation at 15,000 × g for 5 min. Subsequently, 495 μL of the filtrate was mixed with 55 μL of Chenomx standard solution (containing 5 mM deuterated DSS in D2O) and vortexed for 15 s. The pH of the resulting mixture was adjusted to 6.4 using 1 M NaOH. An aliquot of 520 μL of the final solution was transferred to a 5 mm NMR tube for spectral acquisition. The NMR spectra were acquired using the noesypr1d pulse sequence with a spectral width of 14 ppm, 32 K data points, a relaxation delay of 5 s, and 256 scans at 298 K.
Metabolite identification
-
Metabolite identification was carried out using 1D NOESY NMR chemical shifts referenced from literature, Chenomx NMR Suite 7.5 software (Chenomx Inc., Edmonton, Alberta, Canada), and the Human Metabolome Database (HMDB,
www.hmdb.ca ). The identifications were subsequently validated through two-dimensional (2D) NMR techniques including 1H−13C heteronuclear single-quantum correlation spectroscopy (HSQC), and 1H−1H correlation spectroscopy (COSY), identification of condensed polyphenols (6.81−6.97 ppm, non-monomeric phenolic compounds)[6] and L-rhamnitol (1.25−1.27 ppm) were assisted by the literature[6,7]. A representative spectrum of apple samples labeled with identified 36 metabolites is shown in Supplementary Fig. S1. The chemical shifts and peak multiplicities of the metabolites are detailed in Supplementary Table S2.Statically analysis
-
The 1D1H NMR spectra of apple samples were baseline- and phase-corrected as well as quantified with the Chenomx NMR software suite (Professional edition, version 8.3, Chenomx, Edmonton, AB, Canada). The quantitation was performed in both binned method (0.02 ppm) and metabolite fitting (Chenomx library). The quantification standard used was 5 mM deuterated DSS at the chemical shift at 0 ppm in the spectra. Data are Pareto-scaled before multivariate data analysis using SIMCA-P+ (V12.0, Umetrics AB, UmeÅ, Sweden). The relative quantification of condensed polyphenols (6.81−6.97 ppm) and L-rhamnitol (1.25−1.27 ppm) that were not identified in the Chenomx library was expressed as intensities. The concentration of metabolites identified in the Chenomx library were expressed as mM. Statistical analysis for each parameter was performed using ANOVA with IBM SPSS Statistics 26.
-
Figure 1a shows the geographic distribution of apple samples collected from different regions in Shanxi province. Samples were collected from 197 apple samples across 22 different cultivars. These samples were collected from nine counties spread across five cities in the middle and southern parts of Shanxi province. Figure 1b shows the sample information on cultivars, altitude, bagging treatment, and reflective sheet usage during the ripening process. The most represented cultivars were cultivars 'Changfu2' (24), 'NY543' (20), and 'Huaguan' (18). Figure 1b also shows the altitude at which these apples were grown, categorized into different altitude ranges: 0−200 m, 200−400 m, 400−600 m, 600−800 m, 800−1,000 m, and above 1,000 m. The samples from counties Ji, Taigu, Qi, Changzi, and Yangqu were at higher altitudes than the rest of the counties. More than half of the apple samples were from Yuncheng City, specifically from Pinglu, Linyi, and Wangrong counties. The combination of geographic distribution, cultivars/varieties, and cultivation practices illustrated in these charts form complex interactions that contribute to apple quality in Shanxi province.
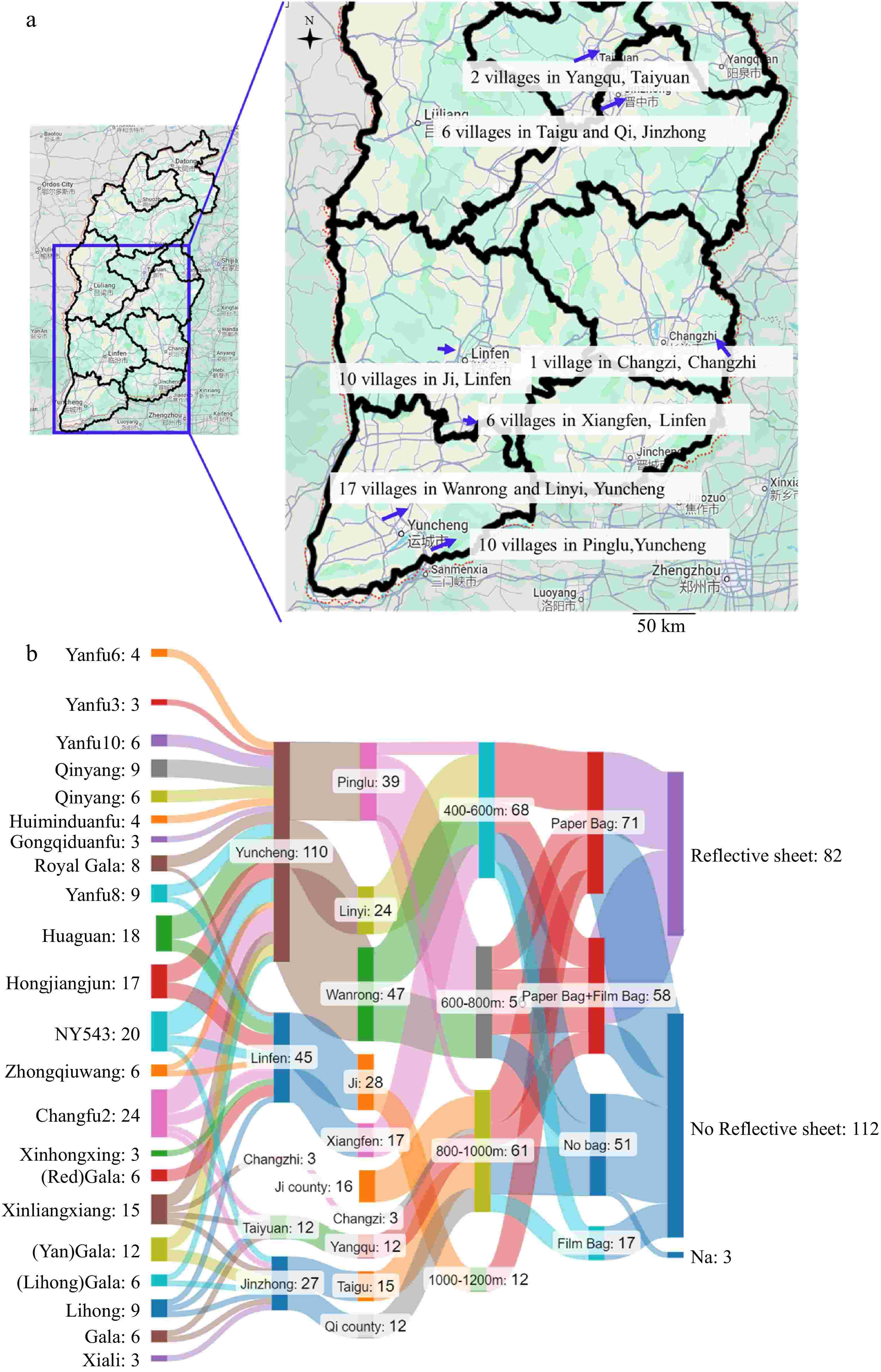
Figure 1.
(a) Geological locations of sampling in Shanxi province, China. (b) Flow diagram demonstrating different sample grouping categories. Na indicates no information.
Multivariate and univariate analysis of 1H NMR metabolic profile of apple samples
-
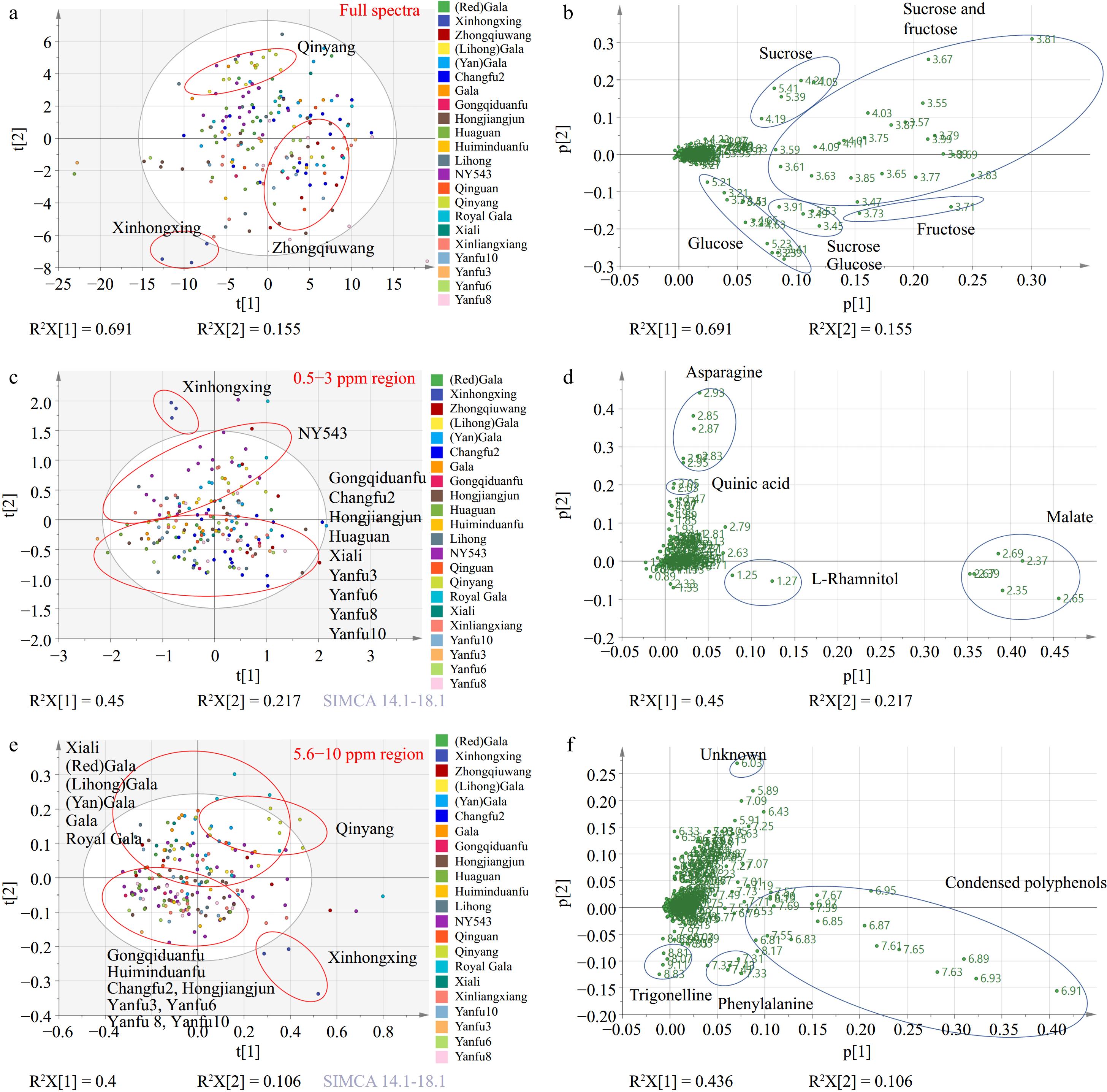
PCA analysis was generated using binned data from full 1H NMR spectra, based on different category strategies: cultivars, location at the county level, bagging method, altitude, and reflective sheet usage (Fig. 2a, Supplementary Fig. S2a−d). Sample separation was not observed in the PCA score plots grouped by the county, bagging method, altitude, or reflective sheet condition.
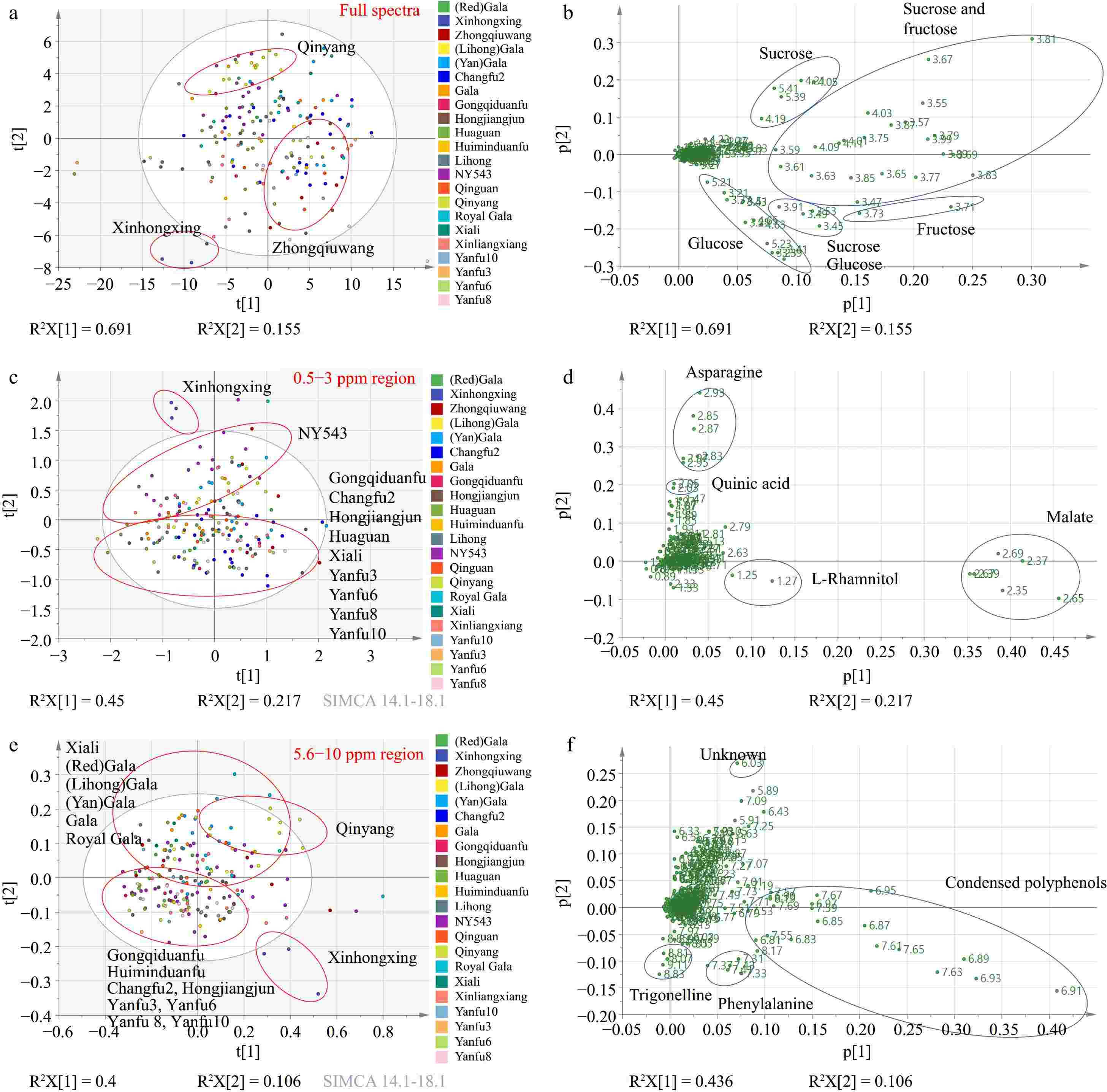
Figure 2.
(a) PCA score plot and (b) corresponding loading plot generated from full spectra of 1H NMR metabolomics. (c) PCA score plot and (d) corresponding loading plot derived from 0.5−3 ppm region. (e) PCA score plot and (f) corresponding loading plot created from 5.6−10 ppm region.
PCA score plots and loading plots of PCA analysis based on cultivar revealed distinct separations among the cultivars 'Qinyang', 'Zhongqiuwang', and 'Xinhongxing' (Fig. 2a & b). This separation is primarily driven by differences in glucose, fructose, and sucrose concentrations (Fig. 2b). Among the studied cultivars, 'Xinhongxing' exhibited the highest glucose levels but the lowest levels of sucrose and fructose (Supplementary Fig. S3a−c). In contrast, 'Zhongqiuwang' had the highest fructose concentration, while 'Qinyang' had the lowest glucose levels (Supplementary Fig. S3a & c).
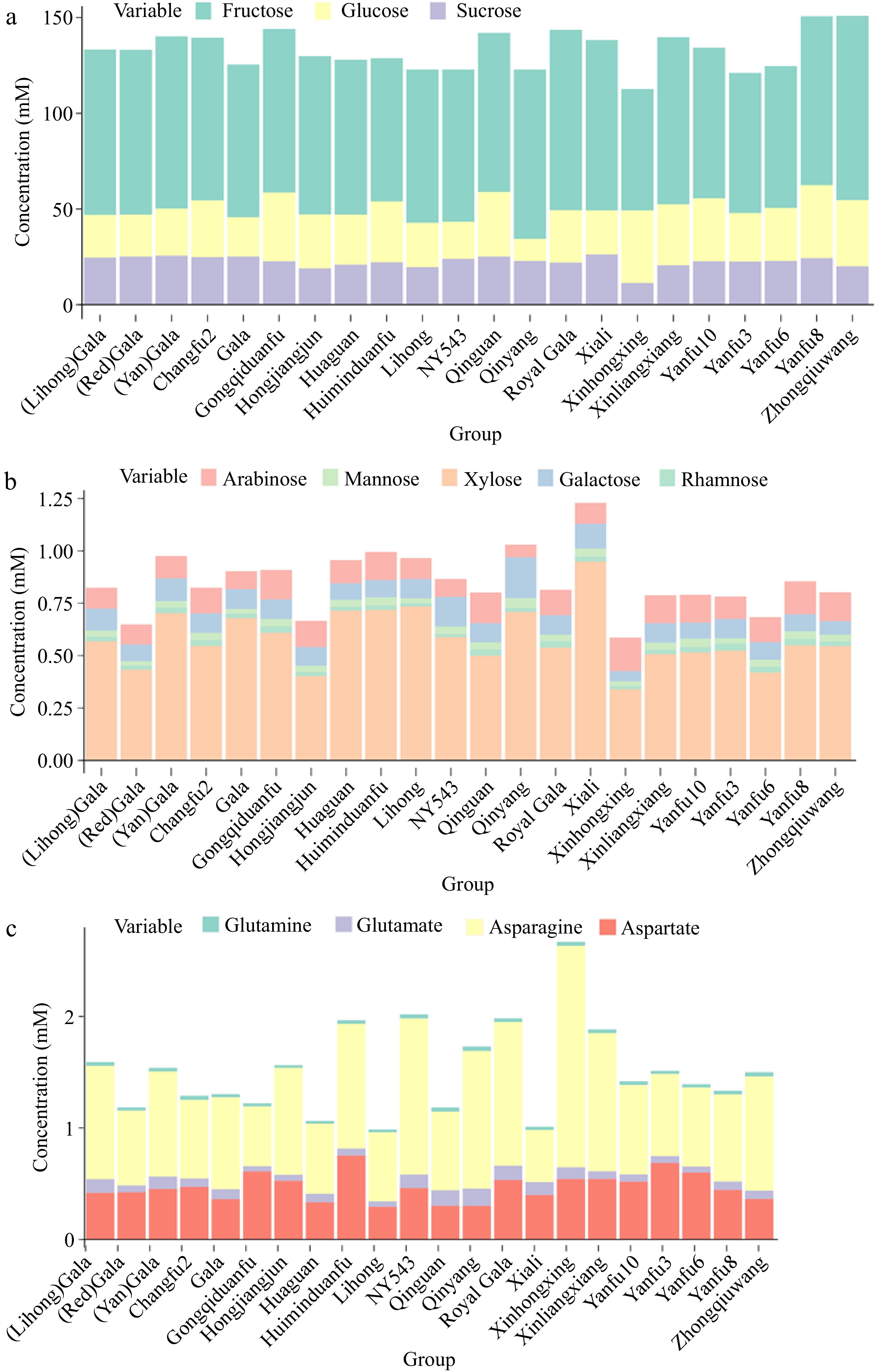
Soluble sugars, which include sucrose, fructose, and glucose, are critical contributors to the sweetness of apples and greatly influence consumer acceptance[14]. Fructose, glucose, and sucrose differ markedly in sweetness, with fructose being nearly 1.8 times sweeter than sucrose and glucose achieving only 60% of sucrose's sweetness[15]. Thus, the taste of fruit is shaped by the composition and concentration of soluble sugars. The total sugar of different cultivars is shown in Supplementary Fig. S4. 'Xinhongxing' might be the least sweet due to its lowest total sugar and fructose levels among the cultivars studied. On the other hand, 'Zhongqiuwang' and 'Yanfu 8' from all locations, with their higher total sugar and fructose levels, are likely to be the sweetest cultivars (Supplementary Fig. S4 & Fig. 3a). More detailed sugar profiles with less concentration are shown in Fig. 3b.
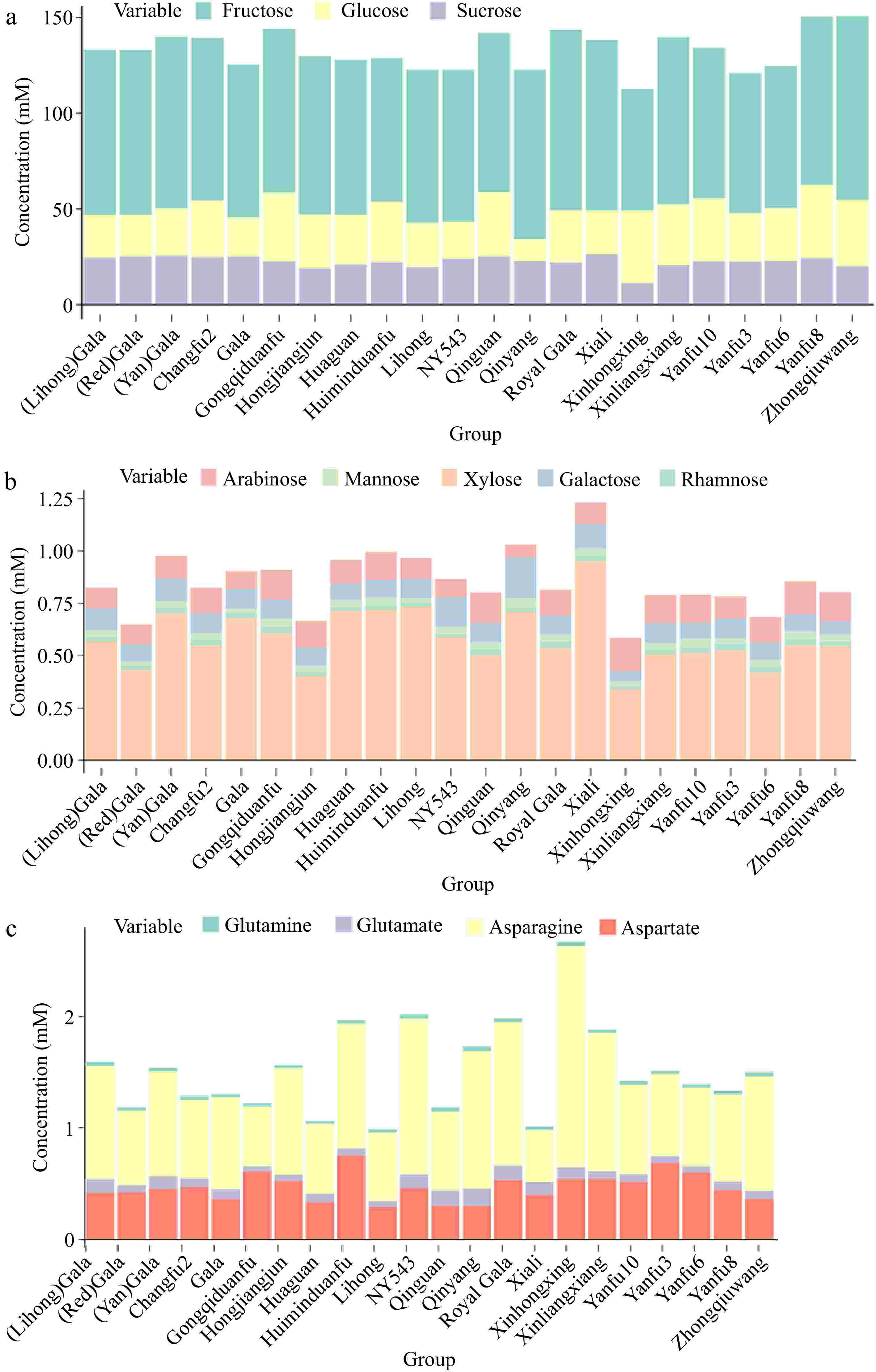
Figure 3.
Stacking plot of (a) fructose, glucose, and sucrose, (b) other sugars, and (c) initial amino acids associated with primary ammonium assimilation in apple cultivars.
However, the rising intake of sugars has been identified as a contributing factor to the obesity epidemic. While fructose was once suggested as a suitable sweetener for individuals with diabetes, it is now recognized that fructose, unlike glucose, strongly promotes lipogenesis, thereby playing a significant role in metabolic dysfunction. One of the apple studies suggested that apples with lower levels of fructose should be recommended[16], although apples with high sweetness are more favorable to consumers. This presents a challenge in balancing consumer preferences and health considerations. To eliminate the impact of the quantitative predominance of the sugars, we excluded the sugar region of the NMR spectra to focus on and better reveal the differences in the amino acid (0.5−3 ppm) and aromatic (5.6−10 ppm) regions.
Multivariate and univariate analysis of 1H NMR metabolic profile of amino acid in apples
-
In the amino acid region (0.5−3 ppm) analysis (Fig. 2c & d), the cultivars were differentiated primarily by the second PCA component, especially 'Xinhongxing' and 'NY543' were separated from the cultivars positioned in the lower part of the PCA plot. The key metabolites driving the separation included asparagine, quinic acid, and L-rhamnitol.
Notably, 'Xinhongxing' and 'NY543' exhibited the highest concentrations of asparagine and quinic acid (Supplementary Fig. S3d−e), with the quinic acid level in 'Xinhongxing' being twice as high as that in 'NY543', although the latter was the cultivar with the second-highest concentration. A 1H NMR study conducted on ten traditional apple cultivars from Italy's Piedmont region identified quinic acid as one of the distinguishing metabolites of apple varieties[17]. The 'Carla' cultivar exhibiting the highest quinic acid concentration had levels 1.6 times greater than that of the 'Grigia di Torriana' cultivar with the second highest concentration[17]. This significant variation in quinic acid content highlights the potential for using the metabolite to differentiate between apple varieties. In apples, quinic acid is synthesized mainly through the shikimate pathway, which is a fundamental biosynthetic route in plants for producing aromatic compounds[18]. Shikimate is converted into quinic acid by the enzyme shikimate dehydrogenase. Once synthesized, quinic acid can serve as a building block for more complex compounds. It often forms esters with other acids, such as caffeic acid, resulting in compounds like chlorogenic acid, which is prevalent in many fruits, including apples[19]. The results indicate that 'Xinhongxing' and 'NY543' might have an active shikimate pathway, potentially leading to higher synthesis of aromatic compounds, particularly in 'Xinhongxing'. Nitrogen metabolism is a fundamental physiological process in plants. Compounds such as glutamate, glutamine, aspartate, and asparagine are the primary products of ammonium assimilation, serving as precursors for the synthesis of other amino acids and nitrogenous compounds within plant transport systems. Among these, asparagine stands out due to its high nitrogen-to-carbon ratio (2:4) and remarkable stability[20], making it a vital carrier for nitrogen transport and storage in plants. This was also confirmed by a strong positive correlation between the content of total nitrogen and asparagine[21]. The stacking plot of glutamate, glutamine, aspartate, and asparagine reveals that the total concentration of these four amino acids is highest in the 'Xinhongxing' cultivar, followed by 'Huiminduanfu', 'NY543', 'Royal Gala', and 'Xinliangxiang' from all locations (Fig. 3c). The highest concentration of asparagine and total levels of glutamate, glutamine, aspartate, and asparagine in 'Xinhongxing' implies efficient nitrogen storage and transport, indicating the strongest ammonium assimilation, which could contribute to the overall vigor and productivity of this cultivar. This trait is advantageous for maintaining nitrogen balance, especially under varying environmental conditions, such as fluctuating nitrogen supply.
Furthermore, 'Xinhongxing' had the lowest concentration of L-rhamnitol (Supplementary Fig. S3f). L-rhamnitol has been recognized as a dietary biomarker for apple consumption and a key metabolite for distinguishing apple varieties based on their geographical origin[5]. A previous study has shown that apples from Japan ('Fuji', 'Orin', and 'Jonagold') contained more L-rhamnitol compared to apples from New Zealand ('Fuji', 'Jazz', and 'Envy')[5]. In the current study, 'Xinhongxing' had the lowest concentration of L-rhamnitol among the analyzed cultivars. 'Yanfu3' and 'Gongqiduanfu' were the two cultivars with the highest level of L-rhamnitol (Supplementary Fig. S3f). To further explore L-rhamnitol as a potential metabolite for geographical discrimination of apple varieties, box plots grouped by location at the city and county levels were made. Regionally, apples from Yuncheng (Linyi, Pinglu, and Wangrong) and Linfen (Xiangfen and Ji) contained more L-rhamnitol than Changzhi, Jinzhong, and Taiyuan (Supplementary Fig. S5). Additional studies are required to clarify the metabolic pathways and genetic mechanisms underlying L-rhamnitol production in apples. Studies involving controlled growth conditions, gene expression analysis, and enzyme activity assays could shed light on how different factors contribute to the observed variations.
Multivariate and univariate analysis of 1H NMR metabolic profile of aromatic region in apple samples
-
In the aromatic region (5.6−10 ppm) analysis (Fig. 2e & f), the second PCA component effectively distinguished the cultivars, particularly 'Qinyang' and 'Xinhongxing'. Key metabolites such as condensed polyphenols, trigonelline, phenylalanine, and an unknown compound, contributed to the separation. 'Xinhongxing' was characterized by the highest concentrations of phenylalanine and condensed polyphenols (Supplementary Fig. S3g, S3h). Additionally, cultivars positioned in the first and second quadrants of the PCA plots exhibited higher levels of an unknown compound (6.03 ppm) and lower levels of trigonelline compared to those in the third and fourth quadrants (Fig. 2e & f, Supplementary Fig. S3i, S3j).
The content of condensed polyphenols can be determined by factors such as the content of total phenolic compounds, polyphenol oxidase activity, and exposure to air when processing[6], indicating 'Xinhongxing' might have more total phenolic compounds and polyphenol oxidase activity. The cultivar 'Xinhongxing', exhibited the highest concentration of phenylalanine, having levels more than 20 times higher than those of the cultivar with the second-highest concentration. Similar to quinic acid, phenylalanine in apples is also primarily synthesized through the shikimate pathway, which is a major biochemical pathway in plants for producing aromatic amino acids including phenylalanine[22]. Phenylalanine is the initial step in the synthesis of phenols and anthocyanins, which converts phenylalanine into cinnamic acid, a precursor for many secondary metabolites including flavonoids and anthocyanins, by phenylalanine ammonia-lyase[23]. The highest levels of phenylalanine and condensed polyphenols in the 'Xinhongxing' suggest again an active shikimate pathway and robust secondary metabolite synthesis, which are crucial for various plant functions and for potential health benefits to humans.
Effect of altitude on malate levels in apple cultivars
-
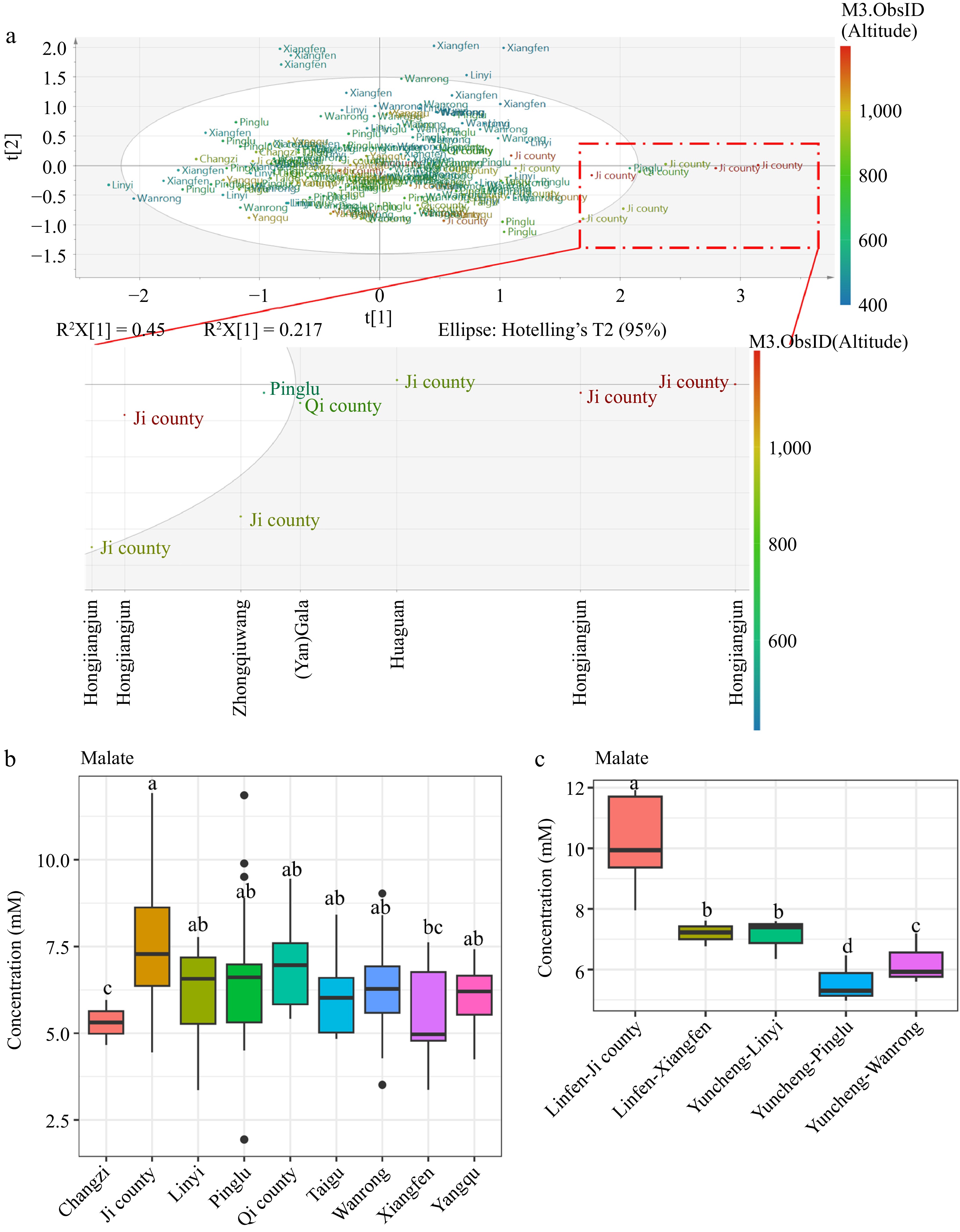
Malic acid is believed to offer various health benefits, including its role in regulating urinary pH, relieving dry mouth symptoms, supporting cardiovascular health, and inhibiting α-glucosidase activity[24−26]. Malate as shown in Fig. 2d contributed to the sample separation by the first component, however, there were no clear cultivar groups separated by the first component, indicating other variables other than cultivar contributing to the sample separation. The PCA score plot for the amino acid region (0.5−3 ppm) was annotated with county information (Fig. 4a), and further analysis showed that apple samples (particularly of cultivar 'Hongjiangjun') from Ji county, which has the highest altitude range of 936 to 1,200 m among the counties growing this cultivar, exhibited elevated malate levels (Fig. 4b, c, Supplementary Fig. S6). However, apples from Changzi, represented by only three samples from the 'Xinliangxiang' cultivar at an altitude of 911 m, showed lower malate levels compared to 'Xinliangxiang' apples from Taigu (altitude 936 m), Pinglu (altitude 555 m), Yangqu (altitude 1,000 m), and Wangrong (altitude 596 m) counties (Supplementary Fig. S6). The contradictory results indicate that altitude-induced malate production might be cultivar-specific. Malate dehydrogenase (MDH), an essential enzyme, facilitates the reversible conversion of oxaloacetate to malate and plays a critical role in controlling malate accumulation. Previously, a study using the apple cultivar 'Gala' showed that MDH transcript expression could be induced by cold (typically at higher altitudes) and salt stresses[27]. Overexpression of MDH in apple plants improves their tolerance to these stresses, potentially by altering biological processes related to the citrate cycle, hormone signal transduction, and oxidation-reduction[27]. However, a study showed that the peel tissues of 'Fuji' apple from high-altitude had substantially lower malate content than those grown at low altitudes during apple ripening[28]. In this study, we found that altitude had a strong impact on the level of malate in fruits of the cultivar 'Hongjiangjun', and 'Hongjiangjun' apples from Ji county might be a more health-promoting cultivar from the perspective of the beneficial effects of malate. For a comprehensive understanding of how altitude might specifically affect malate content in apples, more systematic research would be needed, particularly studies that directly measure malate levels in the same cultivars grown at a range of altitudes with similar soil conditions and standardized practices of cultivation.
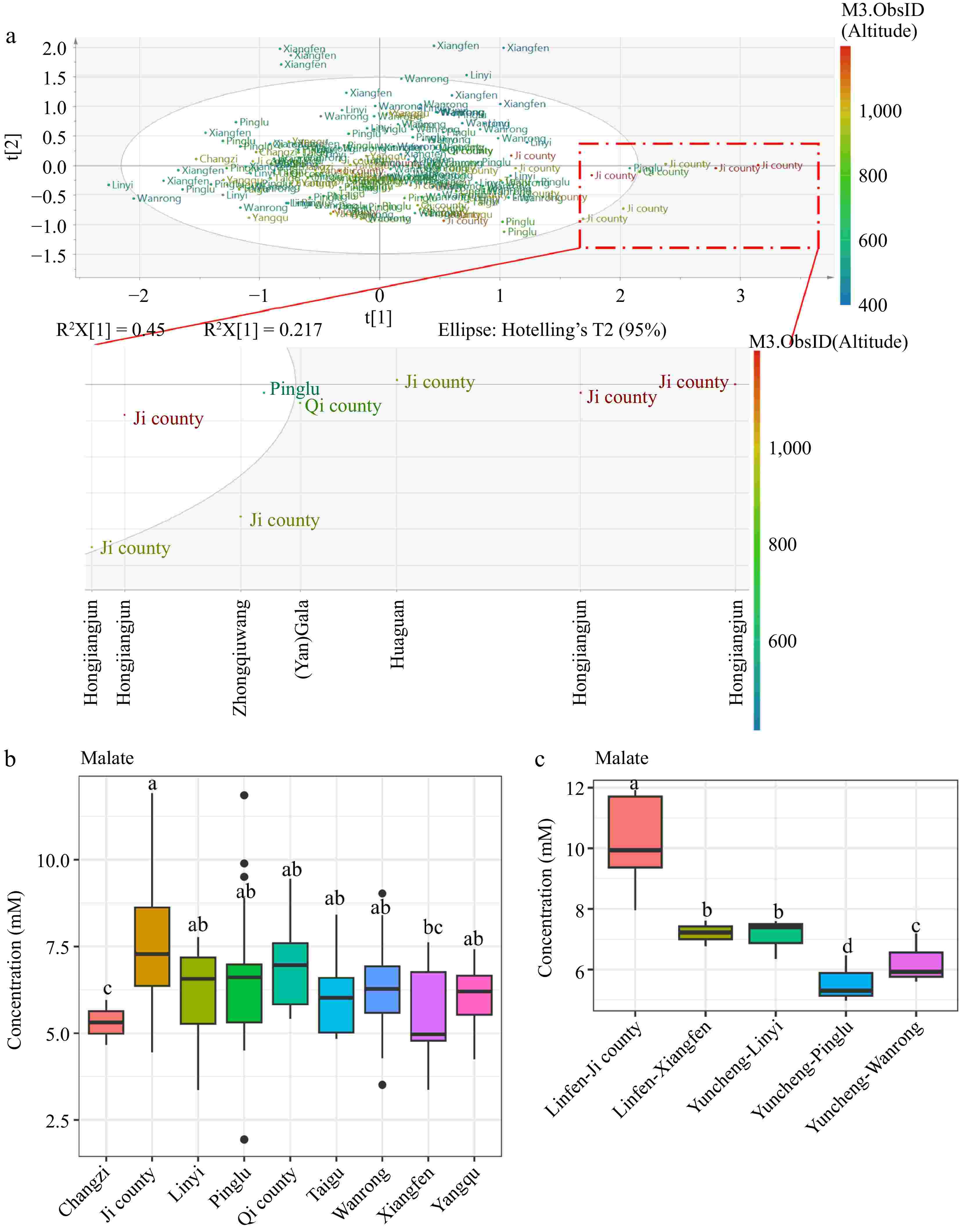
Figure 4.
PCA score plot generated from full spectra and labeled with (a) county information. (b) Concentration of malate in different counties. (c) The concentration of malate in 'Hongjiangjun' cultivars cultivated in different counties. Different letters above the bars indicate statistically significant differences at p < 0.05 (one-way ANOVA).
K means clustering heatmaps based on growing location and cultivar
-
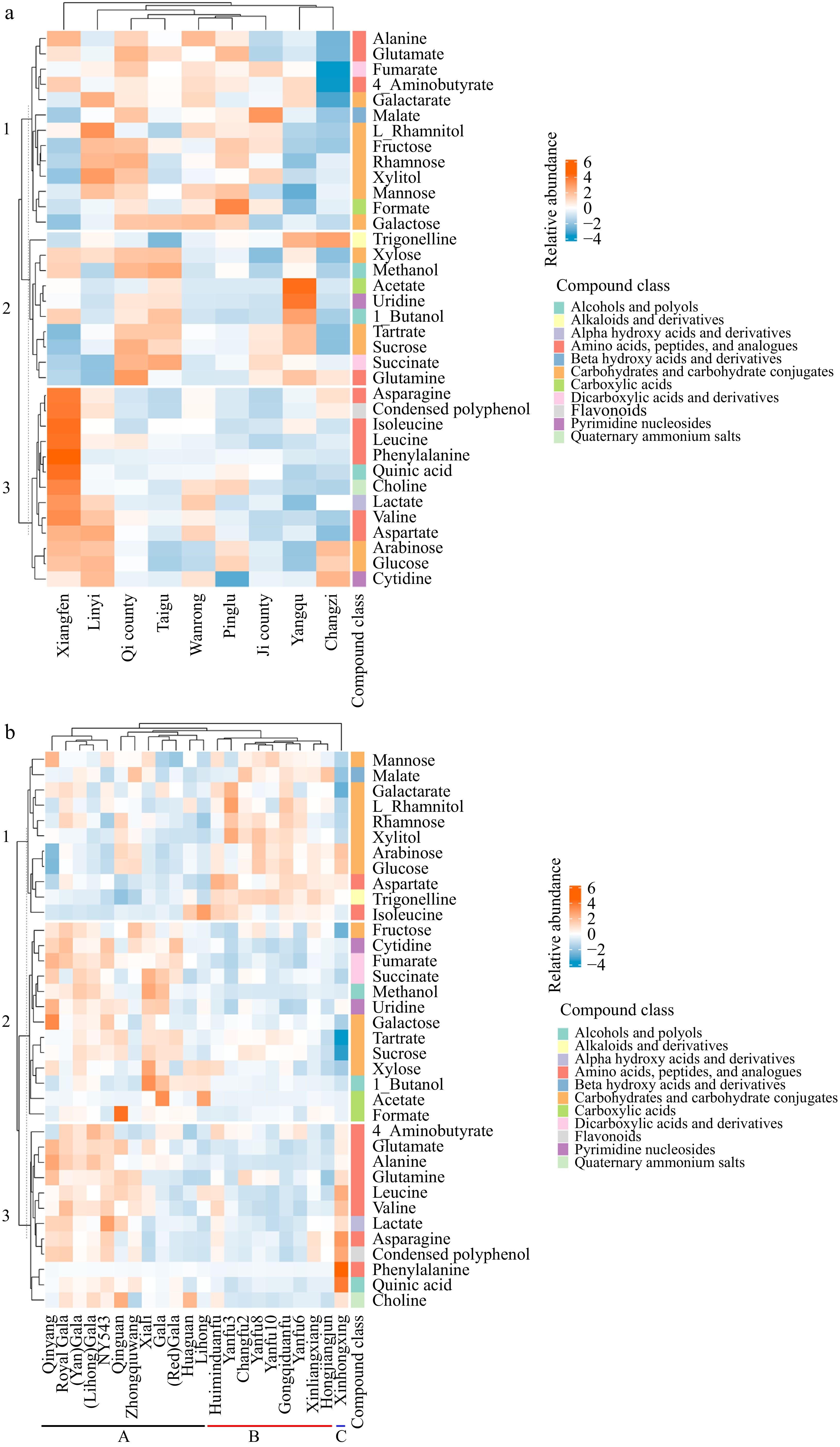
K means clustering heatmaps were generated (Fig. 5) from profiling metabolite data based on growth location (Fig. 5a) and cultivar (Fig. 5b) to illustrate the impact of these two factors on the detailed metabolomics profile of apples. Therefore, in Fig. 5a, the heatmap data of each location are the average values of all the cultivars grown in the location, showing the overall impact of growth location on the composition of apples, regardless of the cultivar. Figure 5b shows the difference among the cultivars independent of the growth locations; here the heatmap values of each cultivar represent the averages from all growth locations. Compounds identified were categorized according to their chemical subclass in the Human Metabolome Database (HMDB). Among these, uridine and cytidine, lacking specific subclass information, were grouped under the broader class of pyrimidine nucleosides. Additionally, condensed polyphenol was manually classified under the flavonoid category.
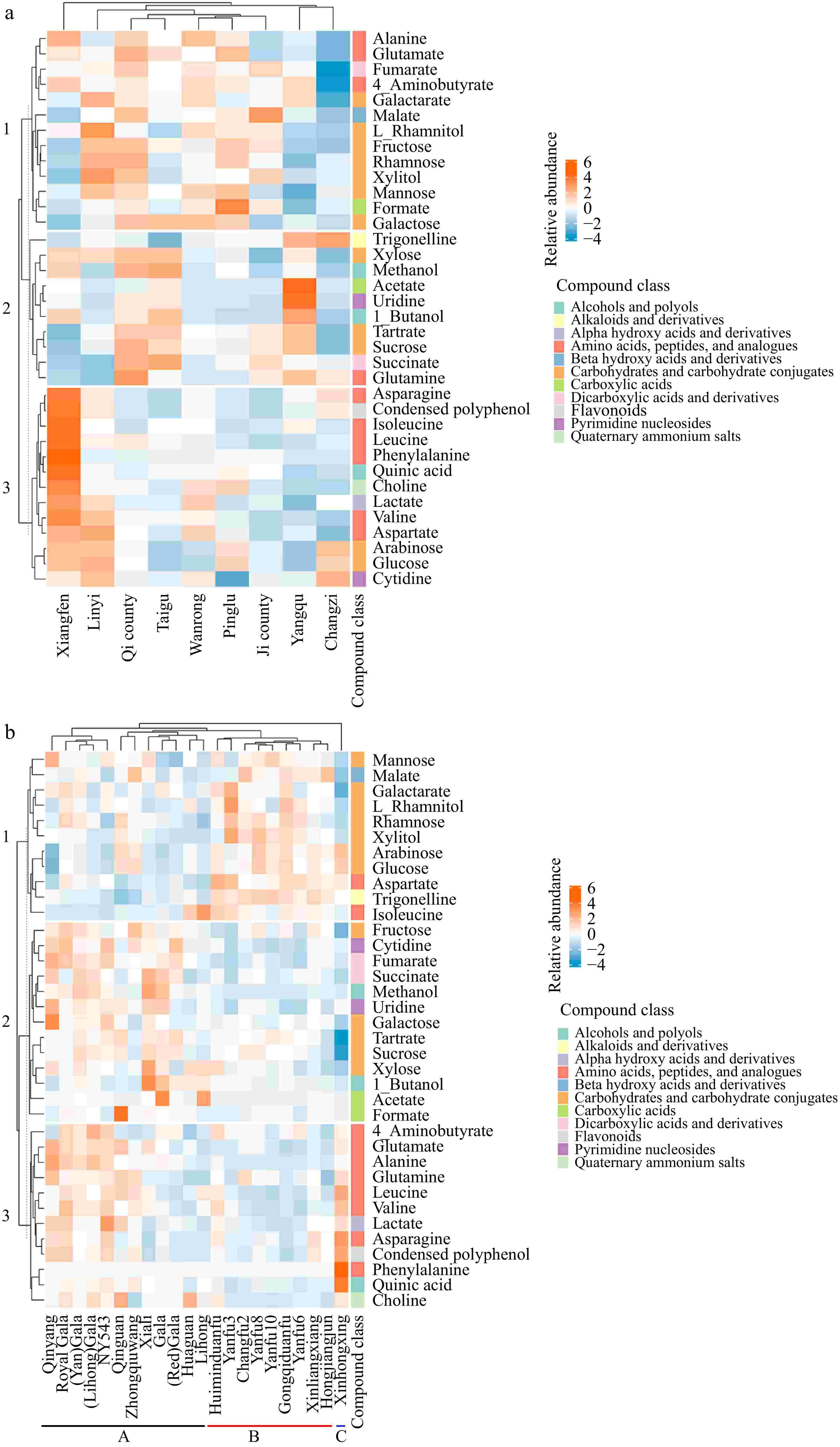
Figure 5.
K means clustering heatmaps generated from profiling metabolite data based on (a) growth location, and (b) cultivar.
Apples from Xiangfen were rich in amino acids (asparagine, isoleucine, leucine, phenylalanine, and valine), quinic acid, choline, and condensed polyphenols, as depicted in cluster 3 of Fig. 5a. In contrast, apples from Linyi were characterized by higher levels of carbohydrates and carbohydrate conjugates, with Linyi specifically showing elevated levels of L-rhamnitol. Additionally, apples from Pinglu had a higher formate content, while apples from Yangqu were notable for increased levels of acetate and uridine.
Figure 5b shows that the cultivars were divided into three clusters: Cluster A includes 'Qingyang', 'NY543', 'Zhongqiuwang', 'Qinguan', 'Xiali', 'Lihong', 'Huaguan', and cultivars related to 'Gala'; Cluster B comprises 'Huiminduanfu', 'Xinliangxiang', 'Hongjiangjun', 'Changfu2', 'Gongqiduanfu', and cultivars related to 'Yanfu'; Cluster C consists solely of the cultivar 'Xinhongxing'. Cultivars in cluster A exhibited higher levels of metabolites, particularly in metabolite clusters 2 and 3, which included amino acids, peptides, and analogues; carboxylic acids; pyrimidine nucleosides; alcohols, and polyols; carbohydrates and carbohydrate conjugates (fructose, galactose, tartrate, and sucrose). Cultivars in cluster B were enriched in carbohydrates and carbohydrate conjugates (L-rhamnitol, rhamnose, xylitol, arabinose, glucose, and mannose), trigonelline, and aspartate, in metabolite cluster 1. In cluster C, the cultivar 'Xinhongxing' was distinguished by higher levels of quinic acid, phenylalanine, and condensed polyphenols. Polyphenols are well-known for their role in plant defense (against pests and pathogens) and environmental stress tolerance (e.g., UV radiation, drought)[29]. The high levels of polyphenols in apples from 'Xiangfen' and 'Xinhongxing' suggest that these regions/cultivars might have an evolutionary advantage in harsh growing conditions.
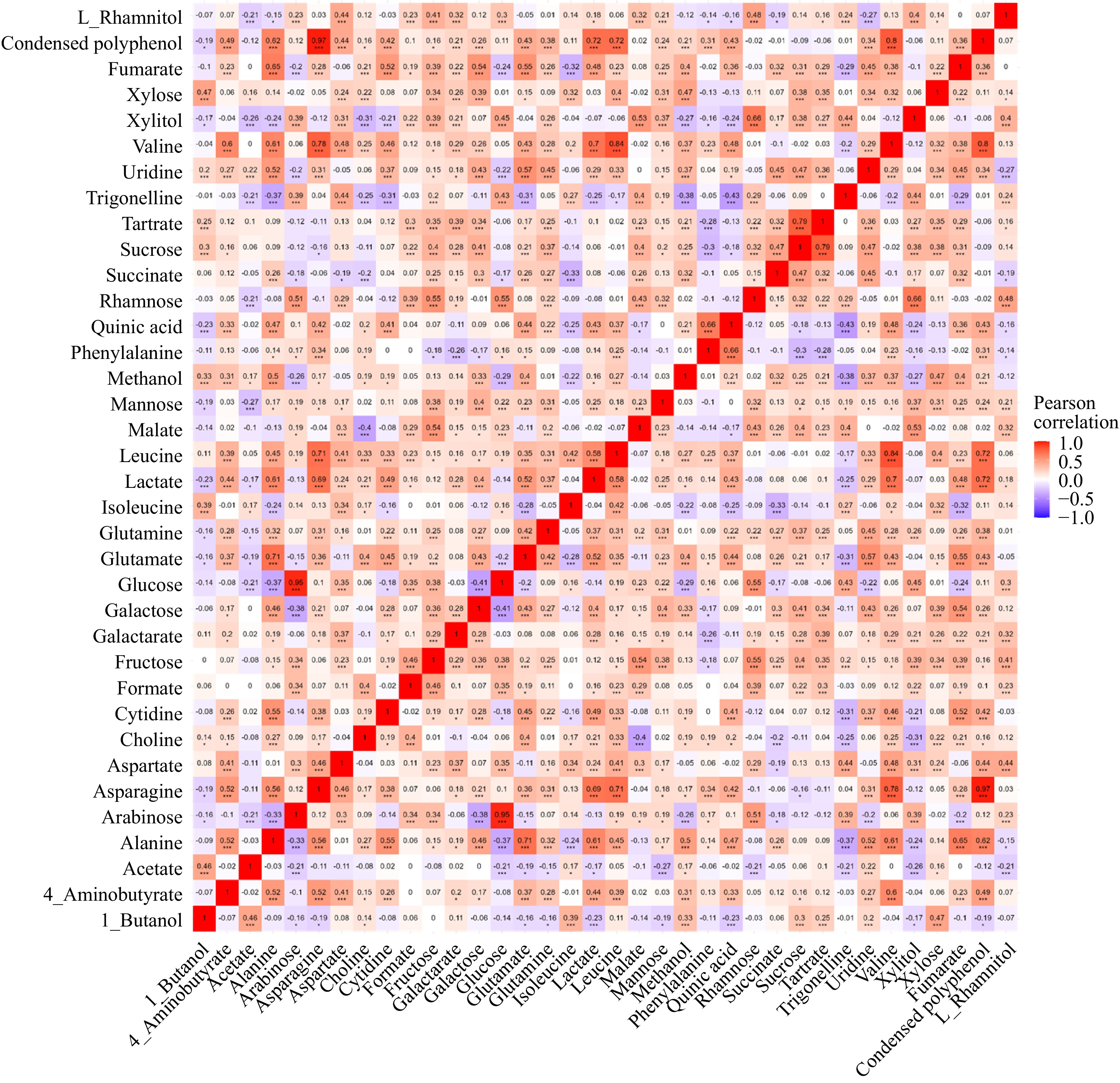
Correlation analysis
-
A correlation analysis of identified metabolites was performed (Fig. 6). In the current study, condensed polyphenol and asparagine exhibited a significant positive correlation (R = 0.97, p < 0.001). Asparagine facilitates nitrogen transport, ensuring resources for the phenylpropanoid pathway, which drives polyphenol production. High asparagine levels signal sufficient nitrogen reserves, allowing plants to prioritize secondary metabolite production for defense and stress adaptation[30]. The correlation between these has not been reported previously. However, a study on the chemical profile of apples cultivated in Norway showed that higher levels of polyphenols tend to accompany a lower level of asparagine[21], reflecting different mechanisms of nitrogen allocation and secondary metabolite synthesis in response to environmental and genetic factors. Quinic acid was positively correlated with phenylalanine (R = 0.66, p < 0.001). Quinic acid is a pivotal intermediate in the shikimate pathway, which is essential for the biosynthesis of aromatic amino acids, including phenylalanine, tyrosine, and tryptophan, highlighting their biochemical interaction[31].
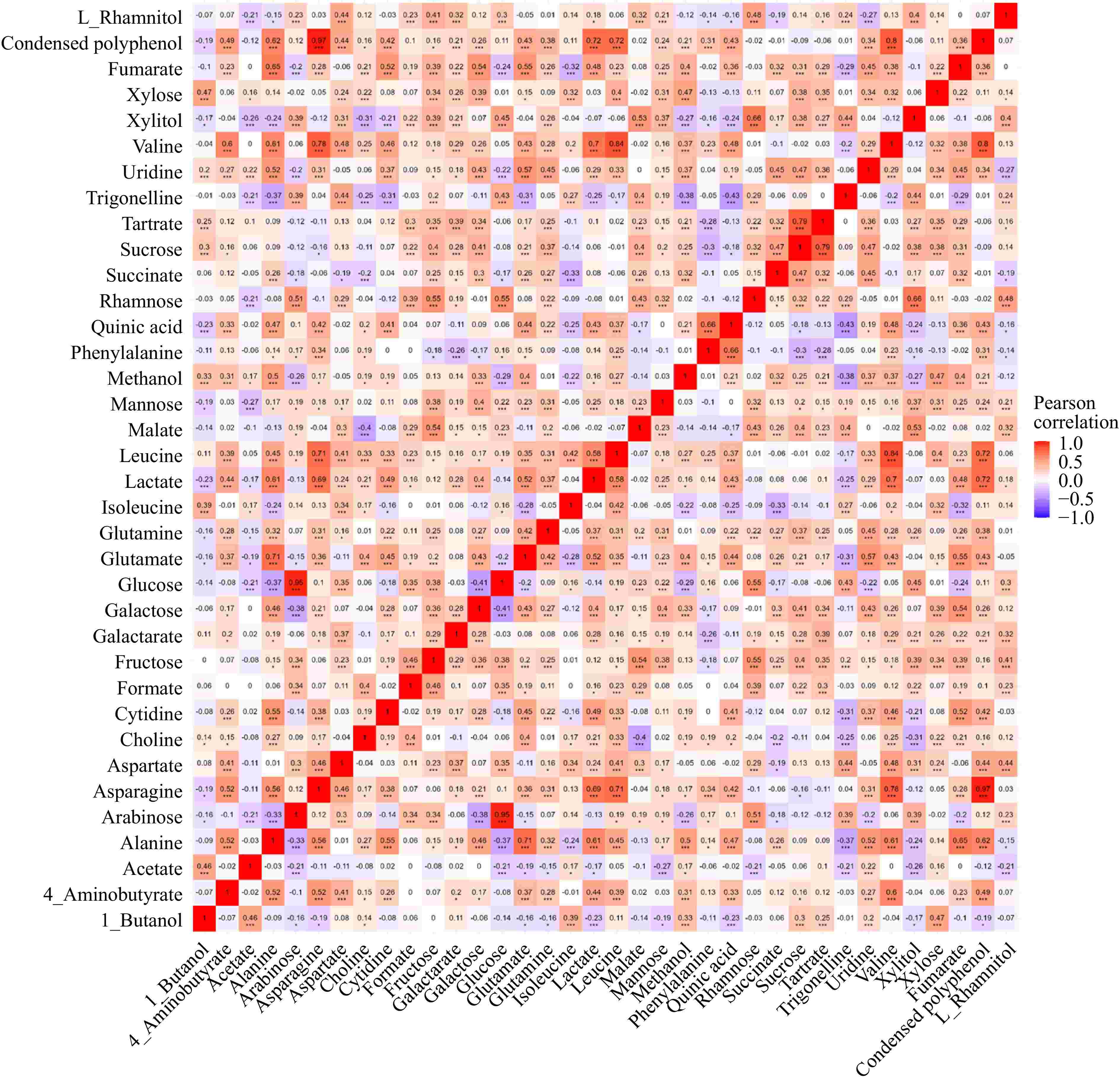
Figure 6.
Correlation plot of identified metabolites in apples.
Valine, alanine, and leucine are branched-chain amino acids (BCAAs) synthesized through a well-coordinated pathway. Their metabolism is tightly regulated to maintain amino acid balance and support vital physiological processes such as protein synthesis and energy production. Asparagine is pivotal in nitrogen transport and storage within plants[30], ensuring efficient nitrogen assimilation and distribution. This process is important for plant growth and development, especially under varying environmental factors. The positive correlation between BCAAs and asparagine observed in our study might reflect the essential role of asparagine in managing nitrogen storage and distribution and its impact on broader metabolic networks.
Tartrate and sucrose were positively correlated (R = 0.79, p < 0.001). Such correlation was not reported in apples, however, a study on the ripening of grapes showed the simultaneous dissimilation of tartrate and the metabolism of sucrose during the ripening process, suggesting a potential interplay between these two compounds[32]. This correlation indicates that the metabolic pathways of sucrose and tartrate might be interconnected.
-
This study provides a comprehensive metabolic profiling of 22 apple cultivars collected from five major apple-growing regions in Shanxi province, China. The systematic profiling of these apple resources revealed critical differences in the levels of sugars, amino acids, and other metabolites, shedding light on the unique characteristics of some cultivars. Our analysis demonstrated that the cultivar plays a more critical role than other factors, such as bagging and reflective sheet conditions, in determining the metabolic composition of apples. The cultivar 'Xinhongxing' displayed the highest glucose levels but the lowest levels of sucrose and fructose, indicating distinct sugar metabolism patterns compared to other cultivars. 'Xinhongxing' and 'NY543' exhibited particularly high concentrations of asparagine and quinic acid. 'Xinhongxing' was also characterized by the highest concentrations of phenylalanine and condensed polyphenols. Additionally, 'Hongjiangjun' from higher altitude regions tended to exhibit elevated malate levels, suggesting potential effects of altitude on the accumulation of this metabolite in apple metabolism. Although the apple treatment information was collected (e.g., bagging method or reflective sheet), the small sample size for each cultivar at each location subjected to specific treatments makes it challenging to discern whether the location, bagging method, or reflective sheet impacted the observed metabolites.
In conclusion, the metabolite profiling of apple cultivars from Shanxi province revealed significant variations in their biochemical compositions, influenced by both genetic and environmental factors. This work reinforces the importance of considering both cultivar and geographical factors in apple production and offers valuable insights into the nutritional value, sensory quality, and potential health benefits of different apple cultivars. Identifying cultivars with higher concentrations of beneficial compounds can guide the development of new varieties that meet consumer preferences for taste and health benefits.
The research was funded by Shanxi Agricultural University, the Finland-China Food and Health Network, and Ningbo University (Grant No. ZX2024000249), the authors thank Professor Yumei Zhang at Peking University for the assistance during the shipping of the samples.
-
The authors confirm their contribution to the paper as follows: study conception and design: Qin S, Yang B, Li J, Qiao X; experimental processing: Chen K, Pariyani R, Li Y; data collection: Pariyani R, Chen K, Li Y; analysis and interpretation of results: Chen K, Pariyani R; draft manuscript preparation: Chen K. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
The authors declare that they have no conflict of interest. Baoru Yang is the Editorial Board member of Food Innovation and Advances who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
- Supplementary Table S1 1H chemical shift assignments of the metabolites observed in the NMR spectra of apple samples using one dimensional (1D) NOESY, two-dimensional COSY, HSQC spectra and comparing standard NMR spectrum of individual metabolites taken from human metabolites database (HMDB).
- Supplementary Fig. S1 1H NMR spectrum of apples: Overview of the spectra (a); Spectrum from 0.5-3 ppm region (b); Spectrum from 3-5.6 ppm region (c); Spectrum from 5.6-10 ppm region (d).
- Supplementary Fig. S2 PCA score plots generated using binned data from full 1H NMR spectra, based on different category strategies: location at the county level (a), bagging method (b), altitude (c), and reflective sheet condition (d).
- Supplementary Table S2
- Supplementary Fig. S3 Quantification of apple metabolites.
- Supplementary Fig. S4 Stacking plot of total sugars.
- Supplementary Fig. S5 Quantification of L-rhamnitol in different locations at the city (a) and county level (b).
- Supplementary Fig. S6 Concentration of malate in different cultivars and locations.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen K, Pariyani R, Li Y, Li J, Qiao X, et al. 2025. Metabolomics profiling of apples of different cultivars and growth locations. Food Innovation and Advances 4(1): 116−126 doi: 10.48130/fia-0025-0012 

Figure 1.
(a) Geological locations of sampling in Shanxi province, China. (b) Flow diagram demonstrating different sample grouping categories. Na indicates no information.