










-
Plants are persistently subjected to detrimental effects caused by environmental stresses such as drought, salinity, heavy metals, cold, and high temperature throughout their lifecycle. Moreover, the severity of these damages is further exacerbated by changing global climate conditions[1]. Among these abiotic stresses, soil salinization has a severely negative effect on plant growth, development and differentiation in the north of China[2]. During historical evolutionary processes, plants have evolved complex physiological and biochemical mechanisms to regulate adaptation and tolerance to abiotic stress. It is worth noting that the adaptive mechanisms are often coordinated by transcription factor (TF) proteins[3,4]. The overexpression of particular stress-responsive transcription factor genes consistently exerts a pronounced influence on plant stress tolerance[5].
MYB as one of the largest transcription factor families consists of two distinct regions: a differential regulatory region at the C-terminal and a conserved MYB DNA-binding domain at the N-terminal[6]. Based on the number of conserved structures, MYB genes were divided into four categories[7]. Firstly, MYB-related genes are an important category of the MYB family and members in this subfamily exert functions in a variety of biological processes, such as cell and organ morphology in plant growth, as well as responses to cold, drought, salt and phosphate starvation under adverse conditions[8,9]. Secondly, R2R3-MYB as the largest subfamily is extensively involved in all aspects of plant growth and development and serves as a key regulator of stress resistance[10,11]. For example, the AtMYB96 of Arabidopsis thaliana regulates the response to drought stress through the integration of the ABA and auxin signaling pathways[12]. Thirdly, it has been reported that 3R-MYB genes are associated with both cellular development and abiotic stress responses. Over-expression of OsMYB3R-2 in rice showed an improved cold-resistance phenotype[13,14]. Lastly, different from the above three subfamilies, members of 4R-MYB have not been well studied to date, and the potential roles of them are still unrevealed[13]. Overall, MYB TFs serve a significant role in regulating stress tolerance and plant development, thereby playing a crucial role in understanding the mechanisms that underlie plant stress resistance and genetic breeding[15].
Poplar, a fast-growing timber tree species, exhibits wide adaptability and rapid growth. However, in the northern regions of China it is consistently hampered by salt stress. Previous research has found that PagMYB73 (PagMYB73A homologous gene) of poplar is inducible by salt-stress[16], and its homologous gene MYB73 in Arabidopsis participates in lateral root growth and stress resistance by regulating auxin biosynthesis[17]. However, the mechanism of PagMYB73A in salt tolerance of poplar is still unclear. In this study, PagMYB73A overexpressing transgenic lines were cultivated using the fast growing poplar hybrid 84K (Populus alba × P. glandulosa) and its function in responding to salt stress was preliminarily revealed. This will provide the material and theoretical basis for improving the molecular mechanisms of salt tolerance of poplar.
-
PagMYB73A overexpression (OE) and wild-type (WT) plants were grown in a greenhouse at Shanxi Agricultural University (Shanxi, China) greenhouse with relative humidity 60%−70%, light 16 h/d, temperature 25 ± 2 °C[18]. The leaves of one-month-old poplar tissue culture seedlings were harvested and immediately frozen in liquid nitrogen for subsequent RNA extraction[19].
Gene cloning and generation of transgenic poplars
-
The pBI121-GFP vector was firstly single digested with XbaⅠ, then homologous recombination plasmid of PagMYB73A-OE was constructed using the specific primers[20, 21] and transformed into Escherichia coli (Supplemental Fig. S1). The positive clone was confirmed by PCR and DNA sequencing (Supplemental Table S1). The Agrobacterium-mediated transformation methods were employed to generate PagMYB73A overexpression lines[22] and the positive lines were subsequently identified using PCR and RT-qPCR (Supplemental Fig. S1). Among the transgenic lines, OE-1, OE-8 and OE-12 with the high, medium and low expression level of PagMYB73A were used for further analysis (Supplemental Fig. S1).
RNA extraction and RT-qPCR
-
After being subjected to salt treatment for 30 d, roots of tissue culture seedlings were collected and immediately frozen in liquid nitrogen. RNA extraction and cDNA synthesis were performed using TIANGEN 's RNAprep Pure Plant Kit and Fast Quant RT Kit, respectively. A real-time fluorescence quantitative PCR system (Agilent Mx3000P, California, USA) and SuperReal Pre Mix Kit (SYBR Green, Tiangen, Beijing, China) were used to detect the relative expression of the target gene using the 2−ΔΔCᴛ method[18, 23].
Phylogenetic and conserved domain analysis
-
The homologous proteins of PagMYB73A were searched using blastp on the NCBI website (
https://blast.ncbi.nlm.nih.gov/Blast.cgi , accession date: 5 March 2023). Multiple sequence alignment and phylogenetic tree construction were performed using ClustalX2.1 and MEGA5.1, respectively. The maximum likelihood method was employed for tree construction, with bootstrap detection of 1,000 replicates and p-distance modelling. The SMART database (https://smart.embl.de/ , accession date: 5 May 2023) was used to predict the conserved domains of PagMYB73A.Subcellular localization
-
A GFP fusion expression vector was constructed to analyze the subcellular localization of PagMYB73A. Following the gene cloning and vector constructing methods described above, pBI121-PagMYB73A-GFP plasmid was constructed successfully and transferred into Agrobacterium EHA105. Agrobacterium-mediated transient transformation methods were used to infect tobacco (Nicotiana tabacum) leaves which were used for subcellular localization analysis under the confocal laser scanning microscope (Olympus FV1000, Japan)[24].
Morphological characteristics
-
The apical buds of one-month-old overexpressing transgenic (OE-1, OE-8, OE-12) and wild-type (WT) tissue culture seedlings were subcultured for 30 d in 1/2 MS medium containing 0 mM (control) or 50 mM NaCl. The morphological indexes, including plant height, leaf number, adventitious root length, root dry weight, and root activity were determined[25]. The WinRHIZO system (Quebec Regent Instrument, Canada) was used for statistical analysis of root surface area, root length, root volume, and average root diameter, etc[26, 27]. Three plants of each line were used as three biological replicates in each experiment.
Physiological characteristics
-
Following a period of 30 d of salt stress, the physiological indexes of the 5th function leaves were measured. SOD, POD, CAT enzyme activities and malondialdehyde (MDA) content were determined by NBT photoreduction, guaiacol, ultraviolet spectrophotometry, and thiobarbituric acid (TBA) staining methods, respectively[27]. For stomatal analysis, the 2nd functional leaf was selected, lower epidermal strips were collected and used to measure stomatal characteristics using an Olympus BH-2 optical microscope (Japan)[28, 29].
Expression of adventitious root-related genes
-
The adventitious roots of both the WT and OE plants, which were cultivated on 1/2 MS medium supplemented with either 0 mM or 50 mM NaCl for a duration of one month, were obtained and employed as the materials for RNA extraction and subsequent RT-qPCR analysis. To detect the molecular function of PagMYB73A in relation to root growth, four genes (Potri.002G114800.1, Potri.004G188800.1, Potri.012G108500.1, Potri.002G196200.1)[30−33] positively involved in adventitious root growth and development were searched and their expression level was analyzed by RT-qPCR with specific primers (Supplemental Table S2).
Data analysis
-
IBM SPSS Statistics 26 (Version 26.0. Armonk, NY: IBM Corporation) was used to analyze the data using the one-way analysis of variance (ANOVA) with a significance level of p < 0.05. Origin 2018 64Bit (OriginLab, Corporation, Northampton, Massachusetts, USA) and Photoshop cc 2019 (Creative Cloud) were used for image production based on averages and standard errors.
-
The homologous sequences of the PagMYB73A protein were obtained by a blastp search on the NCBI website. As a result, we identified 10 proteins from various species that showed a high degree of homology. The evolutionary relationship among these proteins was analyzed by phylogenetic analysis. PagMYB73A of 84K poplar had the closest relationship with Populus trichocarpa (XP_002301201.1) (Fig. 1) and a relatively close distance with Salix brachista (KAB5568943.1). The MYB proteins in Ricinus communis (EEF52460.1), Hevea brasiliensis (XP_021647975.1), Jatropha curcas (XP_012071707.1), Momordica charantia (XP_022144294.1), Dimocarpus longan (UWU45087.1), Litchi chinensis (WAU48795.1), Pistacia vera (XP_031273492.1), Morella rubra (KAB1220389.1) were distantly related (Fig. 1a). Multiple sequence alignment showed that all of these above proteins contained the conserved R2R3-MYB domain and characteristic amino acids (Fig. 1b).

Figure 1.
Phylogenetic analysis and multiple sequence alignment of PagMYB73A and its homologous proteins. (a) Phylogenetic tree of MYB proteins constructed using the amino acid sequences. (b) Multiple sequence alignment of PagMYB73A and its homologues.
Subcellular localization of PagMYB73A
-
Based on results of ProtComp and WoLF PSORT, like most transcription factors, PagMYB73A was also localized in the nucleus (Supplemental Fig. S2). To further validate the prediction findings, we employed Agrobacterium-mediated transient transformation to express the PagMYB73A-GFP fusion protein in tobacco leaves. Intriguingly, the PagMYB73A-GFP fusion protein was detected throughout the cell, coinciding with the distribution pattern observed for the control 35S::GFP protein (Fig. 2). This finding provided evidence that PagMYB73A is situated within the nucleus, membrane, and cytoplasm[34].

Figure 2.
Subcellular localization of PagMYB73A, Bar = 20 μm. 35S::GFP, control protein. 35S::PagMYB73A-GFP, the positive fusion protein. GFP, GFP fluorescence. Chloroplast, chloroplast autofluorescence. Bright: bright fields. Merged: combination of GFP fluorescence, chloroplast autofluorescence, and bright field.
PagMYB73A enhances salt tolerance by promoting adventitious root elongation
-
In order to examine the regulatory mechanism of PagMYB73A in poplar trees subjected to salt stress, the growth indexes of OE and WT plants were quantitatively assessed and subsequently analyzed. Under normal growth conditions (CK), morphological measurements of the OE and WT poplar plants revealed that the OE plants displayed a greater number of leaves and were taller compared to the WT plants (Fig. 3a, b). This suggested that the overexpression of PagMYB73A promoted the longitudinal growth of poplar. Nevertheless, OE's plant height was considerably suppressed and smaller than WT when subjected to salt stress (Fig. 3b). However, under salt stress, OE showed higher adventitious root length and root dry weight than WT (Fig. 3b). Remarkably, both OE and WT exhibited an increase in root fresh weight and root activity in response to salt stress, with no significant difference between them. All of the above results suggested that PagMYB73A may suppress height growth to promote adventitious root elongation, so as to absorb more nutrients and water, when subjected to mild salt stress (50 mM).
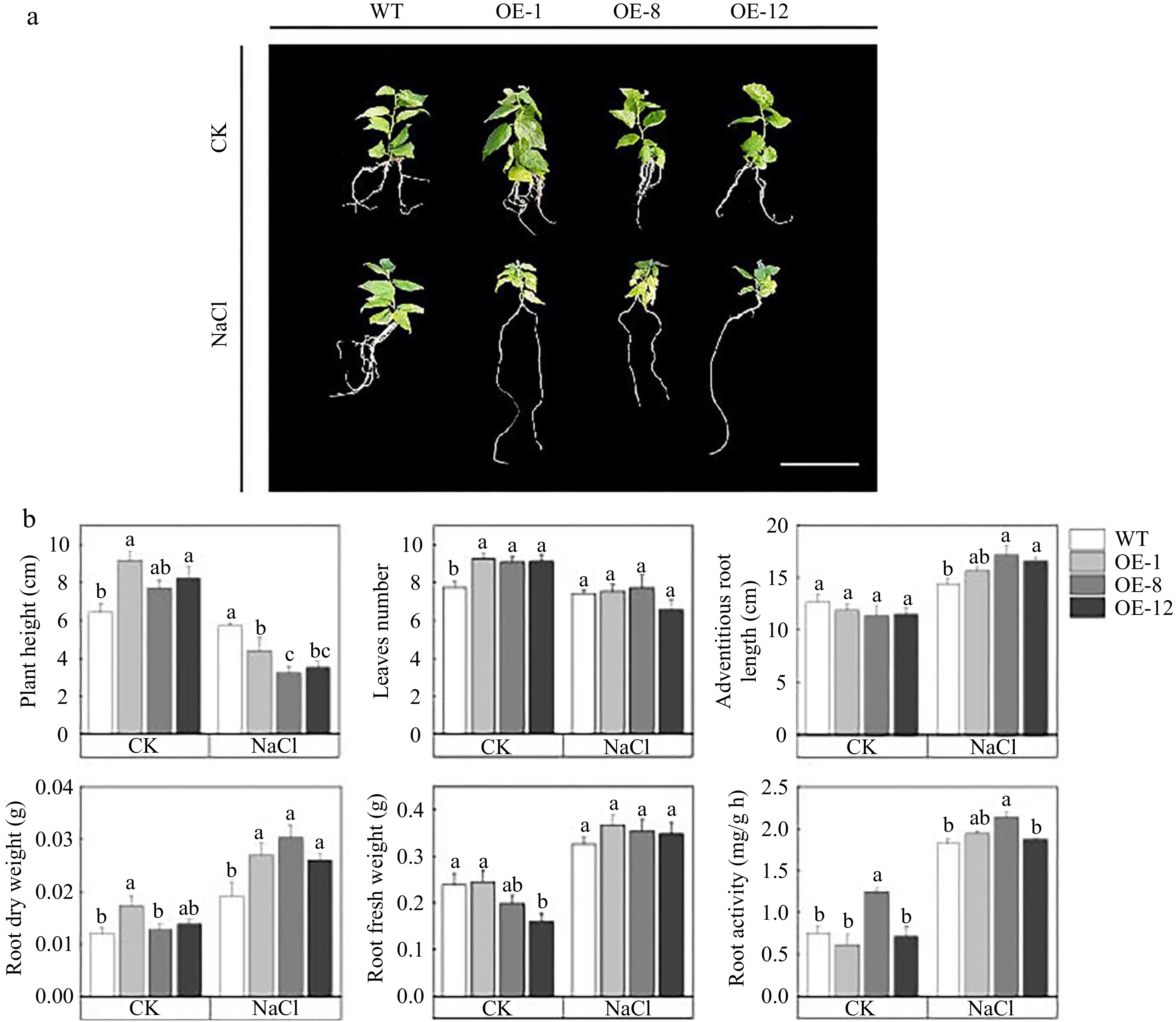
Figure 3.
PagMYB73A promoted adventitious root elongation under salt stress conditions. (a) The morphology of OE and WT, Bar = 2 cm. (b) Measurement of morphological parameters for OE and WT. The graph indicates the mean ± standard deviation of three plants per line. The different lowercase letters indicate significant differences among samples (p < 0.05).
PagMYB73A regulates expression of adventitious root related genes
-
To further substantiate the impact of PagMYB73A on adventitious root growth, a total of four genes involved in adventitious root growth and development were searched, and RT-qPCR was utilized to determine the expression of these four genes in response to salt stress. Our findings revealed that under normal conditions, the expression of these adventitious root-related genes was higher in the WT than in OE. However, when exposed to salt stress, the expression levels of these genes were significantly induced in OE and much higher than in WT plants (Fig. 4a−d). This result suggested that PagMYB73A may promote the growth of adventitious roots by regulating the expression of these genes under salt stress conditions.
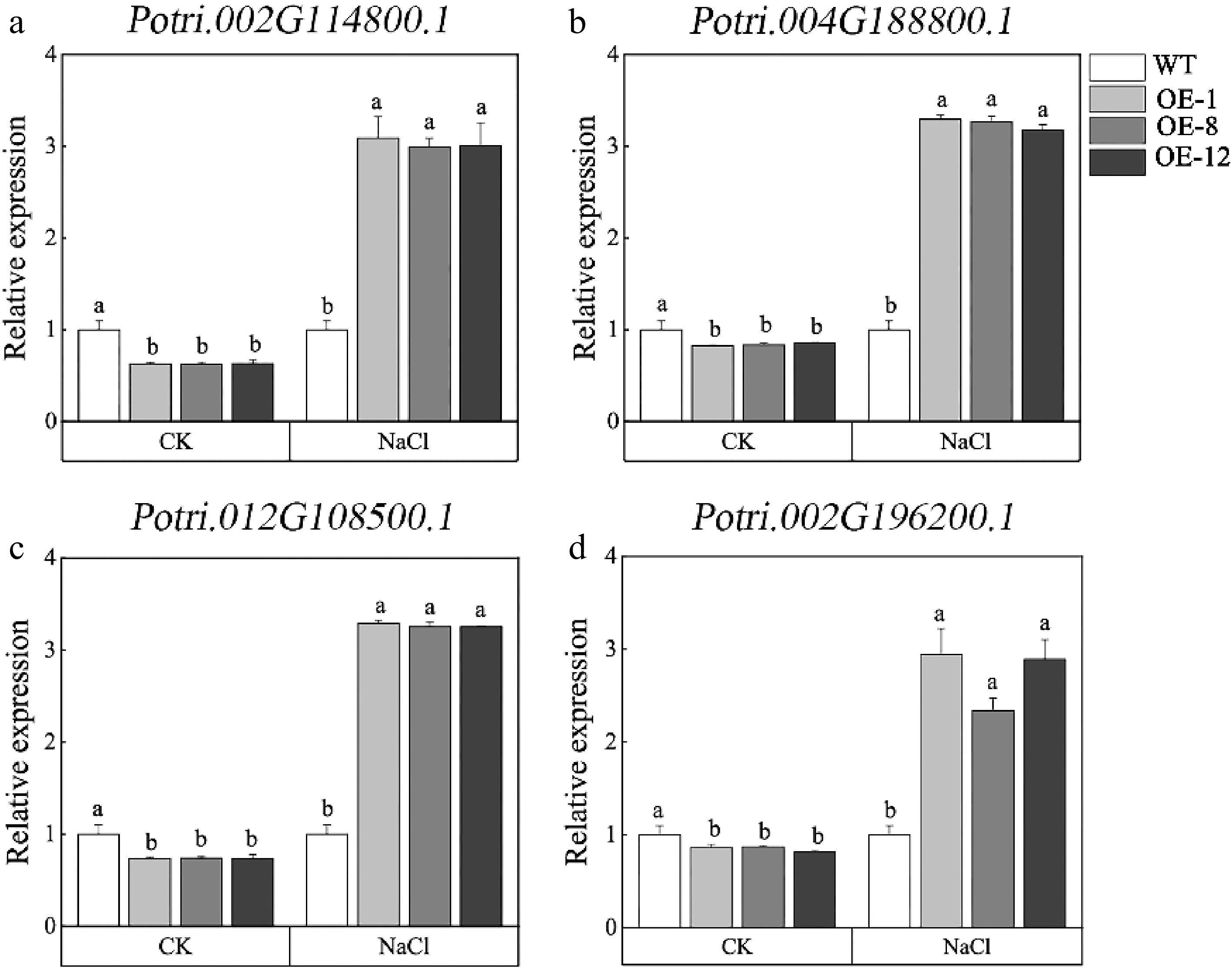
Figure 4.
Expression level of genes involved in adventitious root growth and development. The graph indicates the mean ± standard deviation of three repeats. The different lowercase letters indicate significant differences among samples (p < 0.05).
PagMYB73A positively regulates root growth but inhibits fine roots under salt stress
-
Under both normal and salt stress conditions, the total root area, length, and volume of OE were greater than those of WT (Fig. 5a−c). However, the average root diameter of the OE was lower than WT after treating with salt stress (Fig. 5d). For fine roots (diameter < 1.0 mm), we found that the length and surface area of OE were smaller than WT under salt stress conditions (Fig. 5e, f). Above results indicated that although PagMYB73A positively promoted root growth, it inhibited fine root growth of poplar under salt stress.

Figure 5.
PagMYB73A positively regulated roots growth. (a) Total root surface area, (b) total root length, (c) total root volume, (d) average root diameter, (e), (f) surface area and length of fine roots with diameter < 1.0 mm, respectively. The graph indicates the mean ± standard deviation of three plants per line. The different lowercase letters indicate significant differences among samples (p < 0.05).
PagMYB73A maintains stomata density under salt stress
-
Stomata play a crucial role in the adaptation of plants to abiotic stress, and their density is influenced by various environmental factors[35]. Surprisingly, under both normal and salt stress conditions, no significant difference in stomatal width and density was observed between OE and WT plants (Fig. 6a−c). The density of WT decreased significantly after salt treatment, whereas OE showed no difference (Fig. 6b). These findings suggest that PagMYB73A could maintain stomatal density even under salt stress. Moreover, both OE and WT showed an increase in stomatal width under salt stress (50 mM NaCl) (Fig. 6c).

Figure 6.
PagMYB73A maintains stomata density. (a) Stomata morphology of the 2nd mature leaf, Bar = 20 μm. (b), (c) Statistical analysis of stomatal density and width. The graph indicates the mean ± standard deviation of three plants per line. The different lowercase letters indicate significant differences among samples (p < 0.05).
PagMYB73A reduces cellular membrane damage under salt stress
-
MDA is always used to evaluate the state of membrane lipid peroxidation. Under control conditions, the MDA content of WT and OE showed no significant difference. However, after salt stress, MDA content of OE lines was 22.8%, 17.1%, and 8.5% lower than WT, respectively (Fig. 7a). In addition, plants maintain a dynamic equilibrium of reactive oxygen species (ROS) under control conditions. Plant antioxidant enzyme systems, including superoxide dismutase (SOD), peroxidase (POD), and catalase (CAT) are known to be associated with stress tolerance as they can eliminate ROS. Surprisingly, under normal and salt stress conditions, we found no significant differences in SOD, POD and CAT activities between OE and WT plants (Fig. 7b−d). These findings imply that PagMYB73A regulated MDA pathways rather than ROS pathways to increase poplar's tolerance to salt.
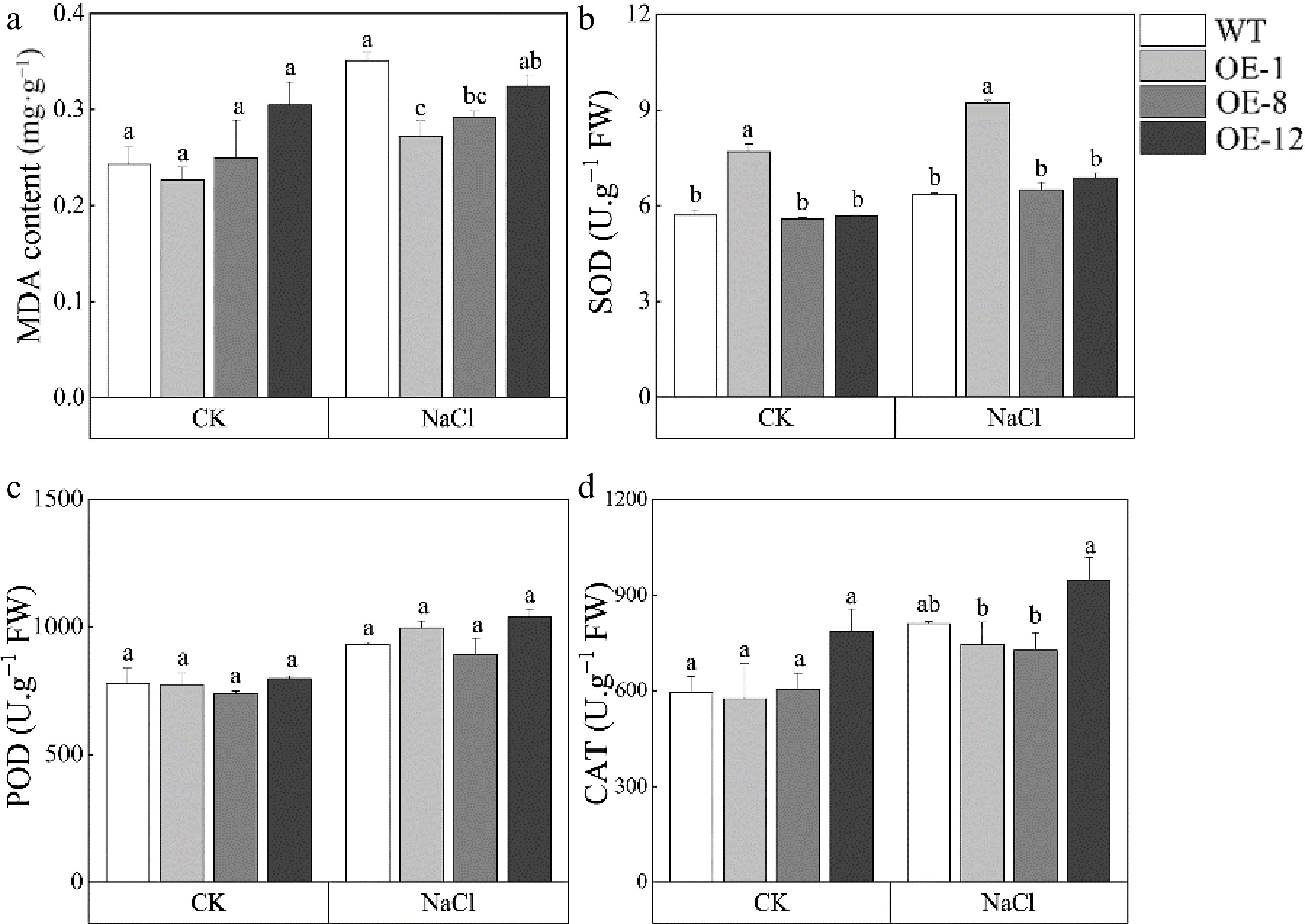
Figure 7.
MDA content and antioxidant enzyme activity of PagMYB73A overexpression transgenic poplar. The graph indicates the mean ± standard deviation of three repeats. The different lowercase letters indicate significant differences among samples (p < 0.05).
-
MYB transcription factors, as one of the key regulators, play vital roles in accommodating growth, development and abiotic stress endurance progresses during the long history of plant evolution[36]. One notable example is AtMYB20, which increases the salt stress tolerance of the AtMYB20-overexpressed transgenic plants by negatively regulating protein phosphatases[37]. Similarly, overexpression of ZmMYB3R improved salt tolerance in transgenic plants with higher survival rates and antioxidant enzyme activities[38]. Additionally, MYB transcription factors were also implicated in plant root growth and development[39]. MYB36 alleviated salt stress by stimulating adventitious root elongation through the induction of adventitious root related genes, which were involved in the differentiation and formation of the Casparian strip[40−42]. In Arabidopsis, MYB73, a homolog of PagMYB73A, interacts with ARF to regulate plant growth, hormone signaling, and lateral root growth[43]. Furthermore, MYB TFs can promote plant development by participating in auxin and ABA hormone synthesis[44]. An example is OsMYB91, which enhances salt tolerance in rice by participating in the ABA-dependent signal transduction pathway[45]. However, the mechanism of how PagMYB73A regulates poplar stress resistance dependent/independent on hormone biosynthesis needs further study.
The primary channels via which gases are exchanged between plants and their environment are the stomata[46]. Under salt conditions, the stomata width of both OE and WT was increased (Fig. 6c, d), which was consistent with the finding that extracellular mild Ca2+ (0.1 mmol·L−1 CaCl2) could promote NaCl-induced stomatal opening, while high concentrations of extracellular Ca2+ effectively inhibited stomatal opening[47]. Of course, the mechanism behind this remains to be revealed. The adjustment of stomatal density is a strategy for plants to adapt to environment stress. Low stomatal density can reduce transpiration and thus favour salt stress tolerance[48]. Overexpression of PdEPFL6 or MtCAS31 improves plant drought tolerance by reducing water loss rate and stomatal density[49, 50]. In this study, salt stress decreased the stomatal densities of both WT and OE, but OE's decrease was not significant (Fig. 6b). This may indicate that OE with greater salt tolerance could maintain stomatal density even under mild salt stress conditions.
Plants with antioxidant enzymes can scavenge reactive oxygen species (ROS) or stop their formation to shield cells from oxidative damage[51]. It is well known that the activity of antioxidant enzymes always increases when plants are exposed to harsh environments[52]. Under salt stress conditions, barley has a marked increase in antioxidant enzyme activity in the root system[53]. In the meantime, transcription factors like ERF38 can boost tobacco's ability to withstand salt by enhancing the activity of antioxidant enzymes[54]. The three main antioxidant defense enzymes, POD, SOD, and CAT, have a close relationship with plants' ability to withstand salt stress[53, 55]. In this study, salt stress raised the activities of SOD, POD, and CAT of both OE and WT, though not significantly (Fig. 7b−d). The results may be explained by the low level of salt stress and even a minor increase of antioxidant defense enzyme activity is also an effective adaptive strategy for plants to cope with adversity[56, 57]. In addition, MDA is a product of plant cell membrane lipid peroxidation, which is an essential sign and always used as the indicator to reflect the damage of the cell membrane[58]. For example, reducing membrane lipid peroxidation (MDA) by the overexpression of PagERF072 improves salt tolerance in transgenic poplar[59]. Under drought stress, transgenic lines overexpressing OsMYB48-1 displayed reduced oxidative damage with low MDA content[60]. Similarly, overexpressing PagMYB73A decreased the MDA content of poplar which may promote the tolerance of OE to salt stress.
-
In summary, PagMYB73A promoted adventitious root growth at the expense of plant height in favour of poplar endurance to salt stress (Fig. 8). The low MDA content reduced the cell membrane damage and contributed to the salt tolerance of the OE. Meanwhile, the ability to maintain stomatal density under salt stress conditions further strengthened the adaptability of OE to salt stress. Collectively, these findings provide novel insights into the fundamental role of PagMYB73A in enhancing poplar's endurance of salt stress.

Figure 8.
Function model of PagMYB73A enhancing salt tolerance of 84K poplar. When subjected to salt stress, PagMYB73A promoted adventitious root growth at the expense of plant height and reduced the MDA content of the poplar as well as maintained stomatal density.
-
The authors confirm contribution to the paper as follows: conceived the idea and designed the research: Wang S, Han Y; modified the manuscript: Wang S, Gailing O; conducted the guidance of conclusion analysis: Han Y, Gailing O; conducted experiments and wrote the manuscript: Jin X, Hu J, Zhou L; provide guidance for experimental operation: Zhao K, Du S. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analysed during the current study are available in online repositories. The names of the repository/repositories and accession number(s) can be found at:
https://submit.ncbi.nlm.nih.gov/ , PRJNA760939. This work was supported by the Natural Science Foundation of Shanxi Province (202303021211091, 20210302123425, 202103021223150) and the Biobreeding Project of Shanxi Agricultural University (YZGC140).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primer sequences used in this study.
- Supplemental Table S2 Primers used for RT-qPCR.
- Supplemental Fig. S1 (a) Escherichia coli colony PCR test. M: DL2000 DNA labeling, 1−3: Three colonies were successfully ligated to the expression vector. The illustration shows that the sample is from the same experiment and that the gel / imprinting is processed in parallel. (b) Agrobacterium colony PCR detection. M: DL2000 DNA labeling, 1−8: 2 were successfully transformed into Agrobacterium. The illustration shows that the sample is from the same experiment and that the gel / imprinting is processed in parallel. (c) PCR detection of transgenic poplar. M: DL2000 DNA labeling, P: Recombinant plasmid containing PagMYB73A, H2O: Blank control, WT: Non-transgenic plant genomic DNA, 1−5: Different lines of poplar transformed with PagMYB73A-OE gene. The illustration shows that the sample is from the same experiment and that the gel / imprinting is processed in parallel. (d) RT-qPCR detection of transgenic plants. WT: Non-transgenic plants, OE-1 , OE-8 , OE-9 , OE-10 , OE-12 : PagMYB73A transgenic plants.
- Supplemental Fig. S2 (a) Subcellular localization of PagMYB73A in ProtComp. (b) Subcellular localization of PagMYB73A in WOLF PSORT.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Jin X, Zhao K, Hu J, Gailing O, Zhou L, et al. 2024. PagMYB73A enhances poplar salt tolerance by facilitating adventitious roots elongation and stomata density. Forestry Research 4: e003 doi: 10.48130/forres-0023-0032 

PagMYB73A enhances poplar salt tolerance by facilitating adventitious roots elongation and stomata density
- Received: 29 September 2023
- Revised: 13 December 2023
- Accepted: 21 December 2023
- Published online: 24 January 2024
Abstract: MYB transcription factors (TF) are recognized as significant regulators in facilitating plant adaptation to various biotic and abiotic stresses. In order to elucidate the involvement of MYB TFs in the growth, development, and salt tolerance of poplar, PagMYB73A was cloned from 84K poplar (Populus alba × P. glandulosa) and utilized as the target gene for generating PagMYB73A overexpressing transgenic lines (OE-1 , OE-8 , OE-12). Compared to the wild type (WT) plants under normal conditions, the OE lines exhibited enhanced plant height and increased leaf number. While under NaCl treatment, the OE lines displayed elongated adventitious roots but reduced plant height compared to WT. At the transcriptional level, the expression of four genes associated with adventitious root growth and development was examined, revealing higher expression levels in WT compared to OE under normal conditions, but the opposite pattern was observed under salt stress. Physiological index analysis demonstrated a significant decrease in stomatal density of WT under salt stress, whereas the reduction was not significant in OE lines. Furthermore, OE lines exhibited decreased content of malondialdehyde (MDA), a marker of cellular membrane damage, compared to WT under salt stress. In summary, this study preliminarily revealed the regulatory function of PagMYB73A in poplar growth and salt stress resistance, highlighting its active role in the adaptation of poplar to salt stress.
-
Key words:
- Salt stress /
- Poplar /
- Gene function /
- Genetic transformation /
- PagMYB73A