










-
The UV-B light in sunlight (280−315 nm) plays a crucial role in the growth, development, and adaptation of plants. UV-B has been regarded as a potential stressor for organisms, as mild UV-B induces plant adaptive responses, while severe UV-B causes plant metabolic disturbances, such as causing damage to DNA, protein, photosynthetic machinery, photosynthetic processes, and arresting the cell cycle[1]. UV-B signals cause UV RESISTANCE LOCUS 8 (UVR8) homodimers to split into monomers, which then interact with CONSTITUTIVELY PHOTOMORPHOGENIC 1 (COP1), and the E3 ubiquitin of COP1 is regulated by SUPPRESSOR of PHYA (SPAs)[2]. COP1/SPA regulates the activity of photomorphogenesis transcription factors (TFs), such as ELONGATED HYPOCOTYL 5 (HY5), HY5-HOMOLOG (HYH), and some B-box TFs[3]. HY5, a bZIP TF, is a positive regulator of photomorphogenesis. Under all light conditions, hy5 mutants of Arabidopsis display elongated hypocotyl[4]. Besides, HY5 is also involved in regulating stress responses. There is evidence that nearly one-third of Arabidopsis gene promoters are bound to HY5[5]. HY5 increases plants' survivability and promotes anthocyanin biosynthesis under UV-B stress by negatively regulating DECREASE WAX BIOSYNTHESIS (DEWAX) expression[6]. HY5 binds to the promoter of ABA-INSENSITIVE 5 (ABI5) with high affinity, which can be significantly enhanced by ABA[7].
As major environmental factors, water deficiencies limit the growth, development, establishment, and productivity of forest stands. Lack of water causes osmotic stresses, affects plants growth and development, even leads to death. It is thought that TFs that recognize and bind to cis-acting elements of stress-responsive gene promoters may regulate drought tolerance. The basic leucine zipper (bZIP) is one of the largest and most conserved transcription factors in plants. The bZIP mainly contains two functional regions, a strictly conservative basic N-terminal DNA-binding region (N- × 7-R/K) with 18 amino acids and a leucine zipper dimerization domain. This aspect of the research suggests that bZIP TFs participate in various plant processes such as seed maturation, growth, light signaling, secondary metabolites biosynthesis, and stress response.
Abiotic stresses, such as UV-B and drought, can trigger both abscisic acid (ABA)-dependent and ABA-independent transcriptional responses in plants[8]. ABA is recognized as an essential hormone in plant development and stress responses. Endogenous ABA level is accumulated in response to water stress, inducing the expression of stress-related genes, such as dehydration-responsive genes. The exogenous ABA is added to model dehydration reactions, which induces the expression level change of various dehydration-responsive genes. Diversiform regulatory mechanisms of plant tolerance to abiotic mediated by bZIP TFs are related to the ABA signal. ABRE binding protein 9 (ABP9) binds to ABA-response element ABRE2 and enhances osmotic and oxidative stress resistances in maize[9]. Furthermore, OsbZIP23, a key regulator in ABA signaling, enhances drought tolerance and decreases the sensitivity of ABA in rice, positively regulating the expression of OsPP2C49[10].
Betula platyphylla (birch) is a deciduous tree species with ecological and economic importance in the Northern Hemisphere, which is widely used in lumber, furniture, buildings, landscape trees, papermaking, medication, and cosmetics. Birch is regarded as a pioneer tree and is often found in open habitats (ridges, rocks, and deforested areas), resulting in increased susceptibility to UV-B and drought stress. On this basis, the present study proposes to explore the potential functions of BpHY5 from birch, particularly for tolerance to UV-B and drought stresses. The research on the characteristics of BpHY5 will be beneficial to genetic improvement, sustainable afforestation, and wood production of birch.
-
The experiment was conducted at Northeast Forestry University, China (45.7203° N, 126.6346° E). The birch seeds were sterilized and cultured in Woody Plant Medium (WPM). After 1.5 months of growth, the seedlings were transplanted into the culture soil in the controlled environment under long-day (16 h light/8 h dark) conditions at 25 ± 2 °C and 60% relative humidity.
Gene cloning and qRT-PCR analysis
-
Specific primer pairs were designed based on the sequence of transcription factor HY5 in birch (KJ466369) from NCBI (Supplemental Table S1). The full-length coding sequence of BpHY5 was amplified by PCR using cDNA from wild-type birch as a template. Total RNA extraction and qRT-PCR were performed as previously described by Gao et al.[11]. The heatmaps were developed by TBtools.
Bioinformatics analyses of BpHY5
-
The homologous HY5 proteins of Arabidopsis thaliana, walnut (Juglans regia), poplar (Populus trichocarpa), jujube (Ziziphus jujuba), Cannabis sativa, melon (Cucumis melo), cork oak (Quercus suber), peach (Prunus persica), castor bean (Ricinus communis), Jatropha curcas, Eucalyptus grandis, grape (Vitis riparia), and cotton (Gossypium hirsutum) were obtained by BLAST searches of the NCBI database. Phylogenetic trees were created using the Jones-Taylor-Thornton (JTT) model and neighbor-joining (NJ) method through MEGA-X. The conserved functional domains of HY5 were defined using TBtools. The conserved motif analysis was done by MEME and TBtools. SWISS-MODEL was used to predict tertiary structure. The promoter sequence of BpHY5 was defined by the genome database of Betula platyphylla (
https://phytozome-next.jgi.doe.gov/info/Bplatyphylla_v1_1 ) and BioEdit software. The promoter was predicted and analyzed by PlantCARE.Vector construction and subcellular location of BpHY5
-
The recombinant vector pROKII-BpHY5-GFP was introduced into Agrobacterium tumefaciens strain GV3101 cells to transform birch and Arabidopsis. The recombinant plasmid (pEarleyGate 103-35S::BpHY5-GFP) and empty vector (pEarleyGate 103-35S::GFP) were introduced into GV3101, which were transformed into onion epidermal cells, respectively. After 24 h of incubation, confocal microscopy LSM800 (Zeiss, Oberkochen, Germany) was used to observe GFP and DAPI.
Transient transformation and stress treatment of birch seedlings
-
To analyze the time-specific expression of BpHY5, the samples of different periods (0, 3, 6, 9, 12, 15, 18, 21 h) from one-month-old birch in soil were collected. One-month-old birch in soil was treated with 0.6 W/m2 UV-B to analyze the expression pattern of BpHY5 under different durations (0, 0.5, 3, and 6 h). The birch seedlings of 1.5-month-old were subjected to a 3, 6, or 12-h treatment with 200 mM mannitol and 10 μM ABA in medium to analyze the expression pattern under different abiotic stresses.
The method for transient transformation of birch seedlings was carried out according to Wang et al.[12]. The transgenic seedlings were subjected to a 3-h treatment with 200 mM mannitol and 10 μM ABA to determine physiological parameters and analyze the gene expression. To ensure accuracy, all samples were harvested in at least three biological replicates.
Transformation and stress treatment of Arabidopsis
-
The mutants hy5 (CS71, hy5-1) were from AraShare. The wild type used in the study was the Columbia line (Col). Transformation of Arabidopsis was performed using the floral dip method. The Homozygous T3 strain was obtained by self-crossing to be used in the experiment and the transgenic line exhibiting the highest level of BpHY5 expression across independent lines and different genetic backgrounds was selected for further analysis. The seedlings of Arabidopsis (Col, hy5, BpHY5/hy5, and BpHY5/Col) in the medium were used to measure the hypocotyl length under darkness or visible light for 7 d.
For the UV-B tolerance assay, seed (Col, hy5, and BpHY5/Col) germination and greening rates were measured under varying UV-B levels (0.8 and 1.6 W/m2) from day 0 to day 6. The fresh weight and root length of 14-day-old seedlings were measured. Seedlings were transferred to soil at 7 d and grown for 7 d. Two-week-old plants were subjected to 0.8 and 1.6 W/m2 UV-B treatment for 10 d for measuring fresh weight, chlorophyll fluorescence, malondialdehyde (MDA), H2O2, superoxide anion content relative conductivity, and component content of stems.
For drought and ABA tolerance assay, seeds (Col and BpHY5/Col) were planted in a medium containing mannitol (100, 200 mM) and ABA (0.5, 1 μM) to measure germination and greening rates from day 0 to day 6.5. The 16-day-old seedlings were harvested to measure fresh weight, root length, MDA, superoxide anion content, and relative conductivity. The whole 16-day-old plants (Col and BpHY5/Col) were harvested to analyze the expression of tolerance-related genes. After being transferred to soil for 20 d, the 27-day-old plants were subjected to drought stress without water for 7 d and re-watered for 3 d.
Chlorophyll fluorescence and staining
-
The chlorophyll fluorescence of Arabidopsis under UV-B was measured by kinetic chlorophyll fluorescence imaging systems (Heinz Walz GmbH, Free State of Bavaria, Germany). Fv/Fm = (Fm−Fo)/Fm. The leaves of materials were stained with DAB (1 mg/mL) and Evans blue (0.5%) overnight, and NBT (1 mg/mL) for 4 to 6 h, respectively.
Physiological, biochemical measurements and cell wall component analysis
-
The relative conductivity was carried out as per the method described by Li et al.[13]. Water loss was analyzed with reference to the method described by Chen et al.[14]. The MDA, hydrogen peroxide, superoxide anion, lignin, cellulose, and hemicellulose content were quantified using a detection kit (Grace Biotechnology, Suzhou, China).
Statistical analysis
-
Data presented as mean ± SD was analyzed using IBM SPSS 22 for statistical significance at p < 0.05. All data were analyzed in triplicate.
-
The full-length ORF of BpHY5 (KJ466369) is 504 bp, encoding a putative protein of 167 amino acids. The phylogenetic analysis of BpHY5 and HY5s which were identified from the other thirteen plant species was performed (Fig. 1a). Amino acid sequence alignment indicated that BpHY5 contained bZIP_HY5-like conserved domain, that was similar to HY5s from other thirteen species, sharing 82.25%, 76.92%, and 73.81% identities with PtHY5, GhHY5, and AtHY5 (Fig. 1b−d). The predicting 3D structure of BpHY5 was shown in Fig. 1e. Analysis of cis-acting elements showed the promoter of BpHY5 contained numerous stress-responsive elements (such as ABRE, ARE), light-responsive elements and hormone-responsive elements (such as abscisic acid) (Fig. 1f). Further, BpHY5-GFP signal distribution was strictly restricted to the nucleus, supporting the prediction that BpHY5 acted as a TF in the nucleus (Fig. 1g). The results indicated that BpHY5 belonged to bZIP family and could be involved in regulating stress responses.
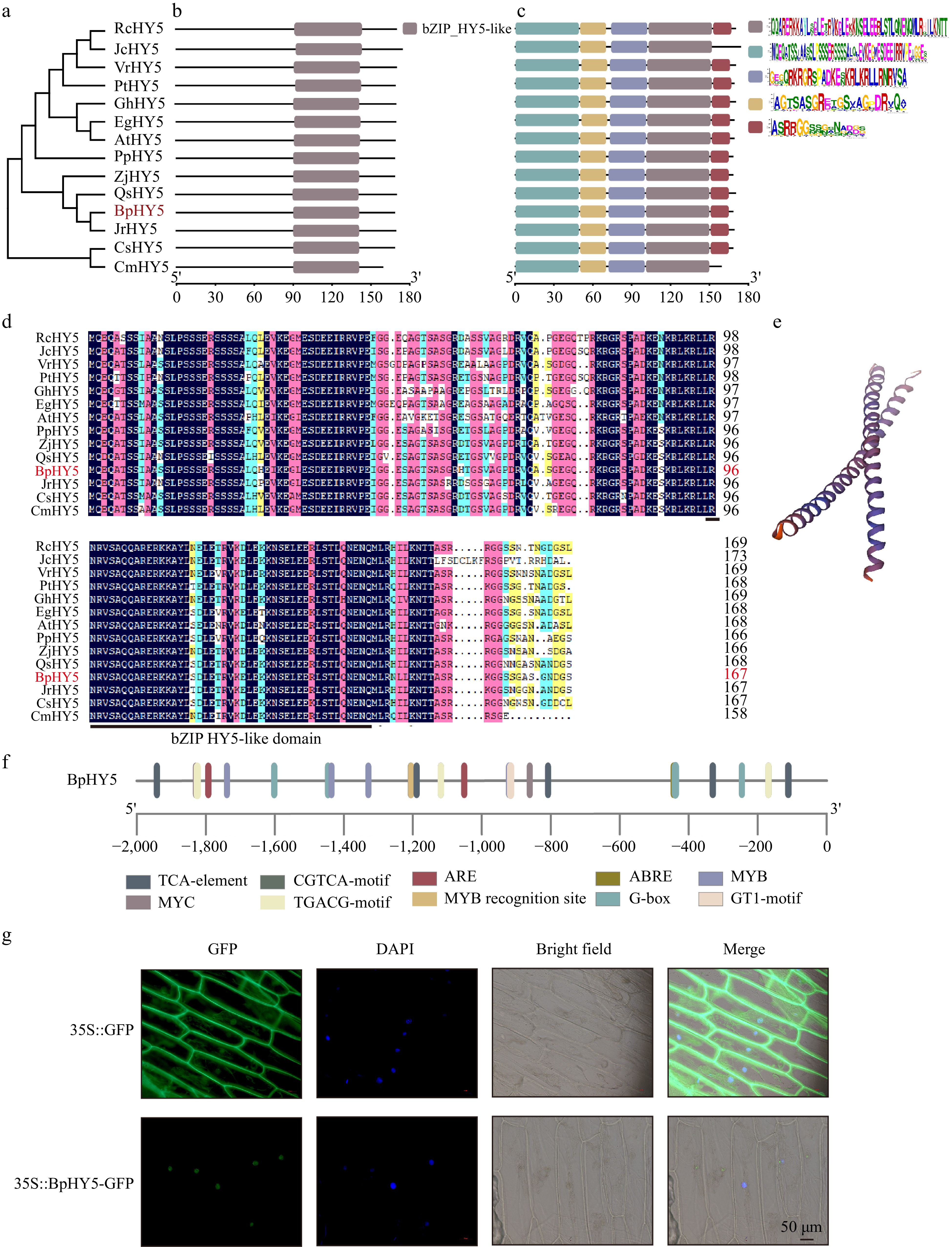
Figure 1.
Phylogenetic, sequence analysis, and subcellular localization of BpHY5. (a) Phylogenetic tree of HY5. Predicted conserved (b) domain and (c) motifs of HY5 proteins, with five colored boxes representing five motifs. (d) Multiple sequence alignment. (e) The predicting 3D structure of BpHY5. (f) Cis-acting element analysis of the promoter sequence of BpHY5. (g) Subcellular localization of 35S::GFP and 35S::BpHY5-GFP. Images represent GFP, DAPI, bright field, and their merged, respectively. The scale bar represents 50 μm.
Expression profiles of BpHY5
-
To study the potential roles of BpHY5, we employed qRT-PCR to assess the expression levels of BpHY5 in various treatments. It showed that the expression reached a peak at 3 h under 0.3 W/m2 UV-B (Fig. 2a). The expression levels of BpHY5 in roots induced by mannitol peaked at 12 h (2.10-fold), while the expression levels in stems and leaves peaked at 3 h (1.42-fold and 4.05-fold) (Fig. 2c). The upregulation of BpHY5 induced by ABA occurred at 12 h in roots (3.05-fold) and 3 h in leaves (4.57-fold) (Fig. 2d). These results indicated that the expression levels of BpHY5 were induced by UV-B, Mannitol, and ABA, suggesting BpHY5 was involved in responses to abiotic stresses, which was consistent with the analysis of the promoter cis-acting elements.
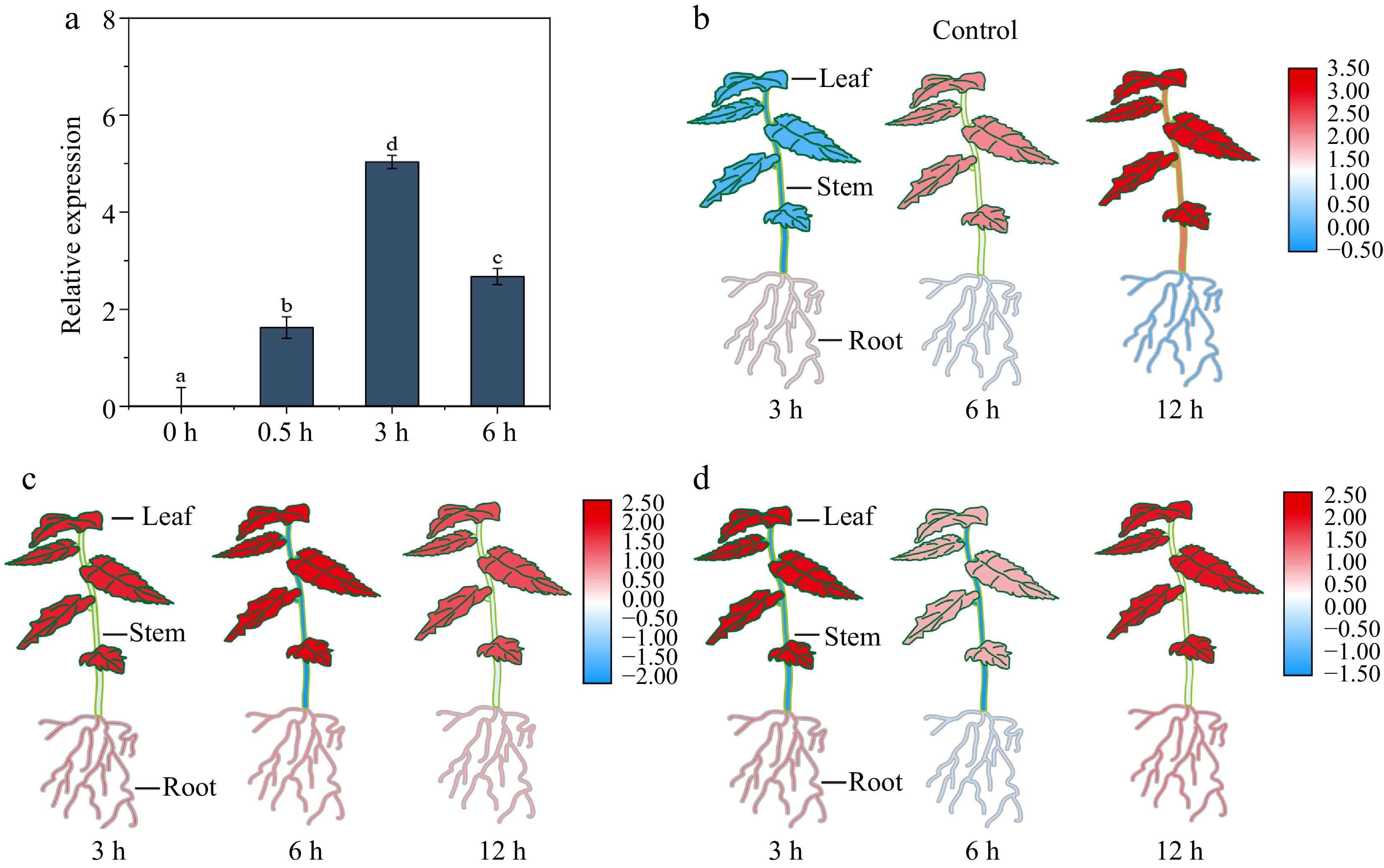
Figure 2.
The expression profiles of BpHY5. (a) The expression level of BpHY5 after 0, 0.5, 3, and 6 h treatment of 0.3 W/m2 UV-B. Heatmap of BpHY5 expression under (b) control, (c) Mannitol, and (d) ABA treatments. Significance was determined by Student's t-test (p < 0.05).
Furthermore, the expression level of BpHY5 at different time points in one day (16 h light/8 h dark) showed that BpHY5 was induced by light (Supplemental Fig. S1a). The hy5 displayed drastically elongated hypocotyls and BpHY5/Col displayed shortened hypocotyls in both light and darkness, whereas the hypocotyls length of BpHY5/hy5 was similar to Col, which indicated BpHY5 restored hy5 mutant phenotype and had similar functions to AtHY5 (Supplemental Fig. S1b, c).
Overexpression of BpHY5 increases the UV-B tolerance
-
To determine the effect of BpHY5 on UV-B tolerance in transgenic Arabidopsis, the growth of plants exposed to UV-B was compared to normal growth. Although there was no discernible difference between Col, hy5, and BpHY5/Col under control, the germination rate of Col was lower than BpHY5/Col under 0.8 W/m2 UV-B and the greening rate of BpHY5/Col was higher than Col under both 0.8 and 1.6 W/m2 UV-B (Fig. 3a−g). The fresh weight of BpHY5/Col was significantly higher, which was 1.23-fold and 1.92-fold of Col under 0.8 and 1.6 W/m2 UV-B, whereas the fresh weight of Col was significantly higher than hy5 (Fig. 3h−j). The root length of BpHY5/Col was increased 1.41-fold and 1.65-fold of Col under 0.8 and 1.6 W/m2 UV-B, respectively, whereas that of hy5 was decreased by 72.82% and 76.79% compared to Col (Fig. 3k).
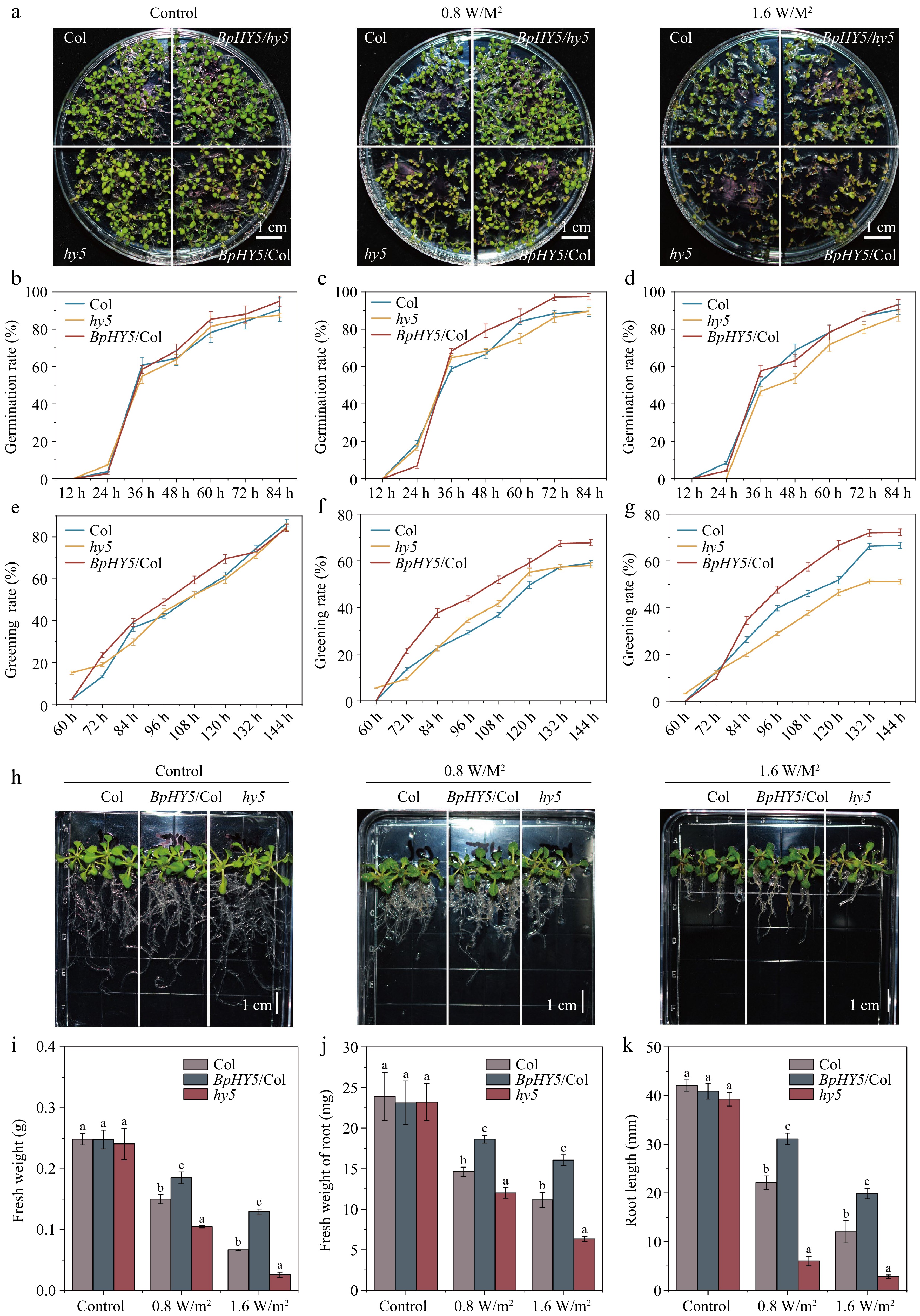
Figure 3.
Comparison of germination, greening rate, root length, and fresh weight between BpHY5/Col, hy5, and Col under UV-B treatment. (a) Physical phenotypes of Col, hy5, BpHY5/hy5, and BpHY5/Col lines under 0.8 and 1.6 W/m2 UV-B treatment after 6 d. The germination rate of Col, BpHY5/Col, and hy5 under (b) control, (c) 0.8, and (d) 1.6 W/m2 UV-B. The greening rate of Col, BpHY5/Col, and hy5 under (e) control, (f) 0.8, and (g) 1.6 W/m2 UV-B. (h) The phenotypes of Col, BpHY5/Col, and hy5 seedlings exposed to UV-B at 14 d. The statistics of (i) fresh weight, (j) fresh weight of root, and (k) root length of Col, BpHY5/Col, and hy5 under 0.8 and 1.6 W/m2 UV-B at 14 d. The scale bar represents 1 cm. Statistical significance (p < 0.05) between Col, BpHY5/Col, and hy5 is indicated by a, b, and c.
After 10 d of UV-B treatment, the size of BpHY5/Col was more developmental than Col and the hy5 began to wilt and turned yellow and brown (Fig. 4a). The fresh weight of BpHY5/Col was 1.31-fold and 1.52-fold of Col under 0.8 and 1.6 W/m2 UV-B, respectively (Supplemental Fig. S2c). Under 0.8 and 1.6 W/m2 UV-B, Fo, and Fm of BpHY5/Col were significantly greater than those of Col, while Fo of hy5 was lower (Fig. 4b−e). Furthermore, compared to Col, Fv/Fm of BpHY5/Col was higher under both control (1.03-fold of Col) and UV-B (1.02-fold and 1.03-fold of Col) conditions, indicating that overexpression of BpHY5 promoted photosynthetic potential and increased the plant tolerance to UV-B (Fig. 4f). In conclusion, these results suggested that BpHY5 positively regulated the plant resistance to UV-B.
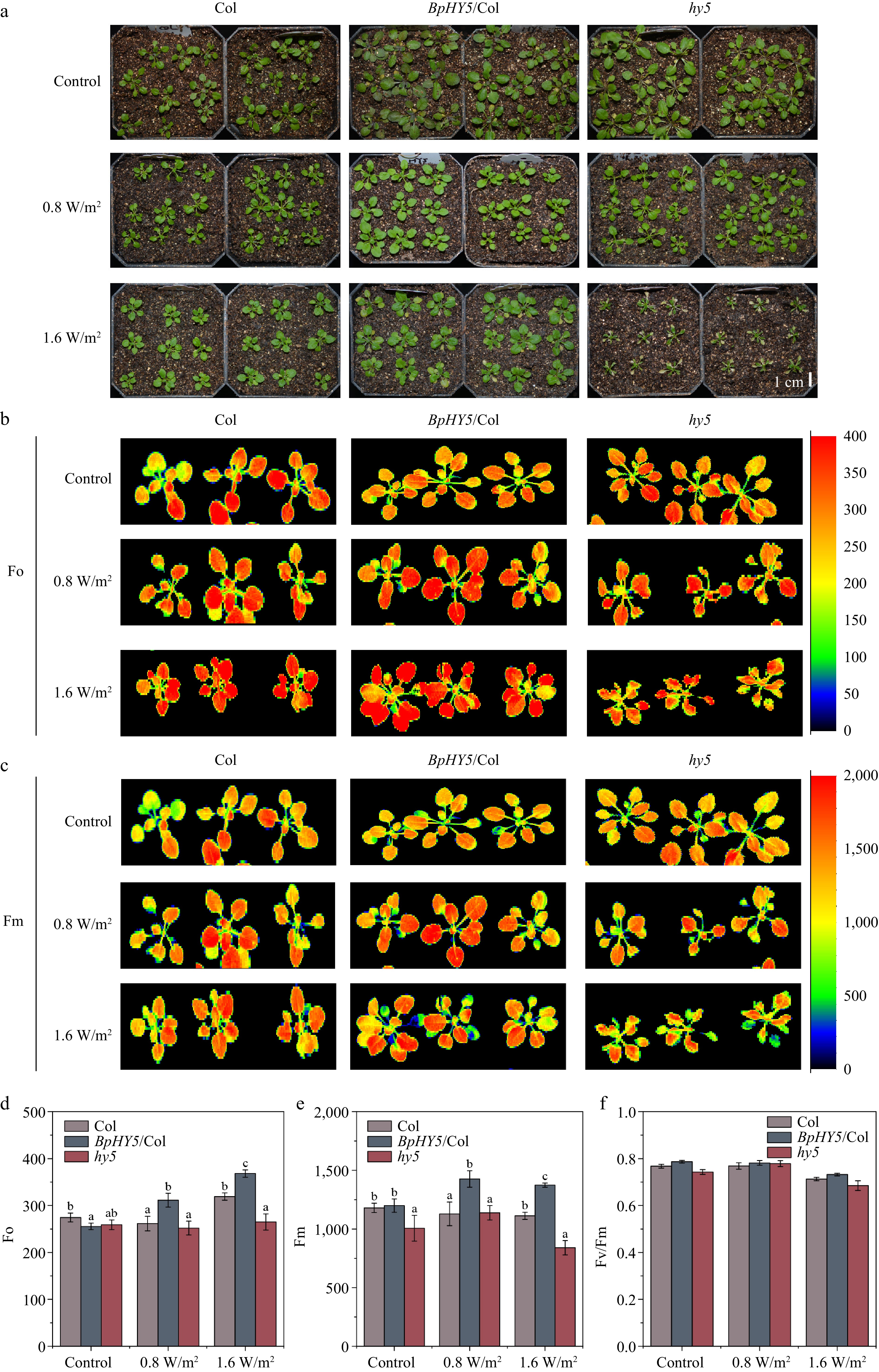
Figure 4.
Transgenic BpHY5 Arabidopsis showed higher UV-B tolerance. (a) Physical phenotypes of Col, BpHY5/Col, and hy5 seedlings in soil under UV-B treatment. The chlorophyll fluorescence of Col, BpHY5/Col, and hy5 seedlings after UV-B treatment for 10 d. Fo is (b) minimal fluorescence, and Fm is (c) maximal fluorescence. The statistics of (d) Fo, (e) Fm, and (f) Fv/Fm of Col, BpHY5/Col, and hy5 under 0.8 and 1.6 W/m2 UV-B treatment after 10 d. The scale bar represents 1 cm. Significance was determined by Student's t-test (p < 0.05).
BpHY5 improves the ROS scavenging ability and affects the cell wall components of stems under UV-B stress
-
After 10 d treatment with UV-B, BpHY5/Col exhibited less damage compared to hy5 and Col. The BpHY5/Col had lower brown coloration or blue spots than Col and hy5 (Fig. 5a). The MDA, H2O2, superoxide anion content, and relative conductivity of all Arabidopsis lines increased after UV-B treatment (Fig. 5b−e). As expected, compared to Col, BpHY5/Col had lower MDA content (0.83-fold, 0.62-fold), lower H2O2 content (0.51-fold, 0.56-fold), lower superoxide anion content (0.78-fold, 0.62-fold), and lower relative conductivity (0.83-fold, 0.79-fold). These findings demonstrated overexpressing BpHY5 enhanced tolerance to UV-B radiation by inhibiting ROS accumulation.
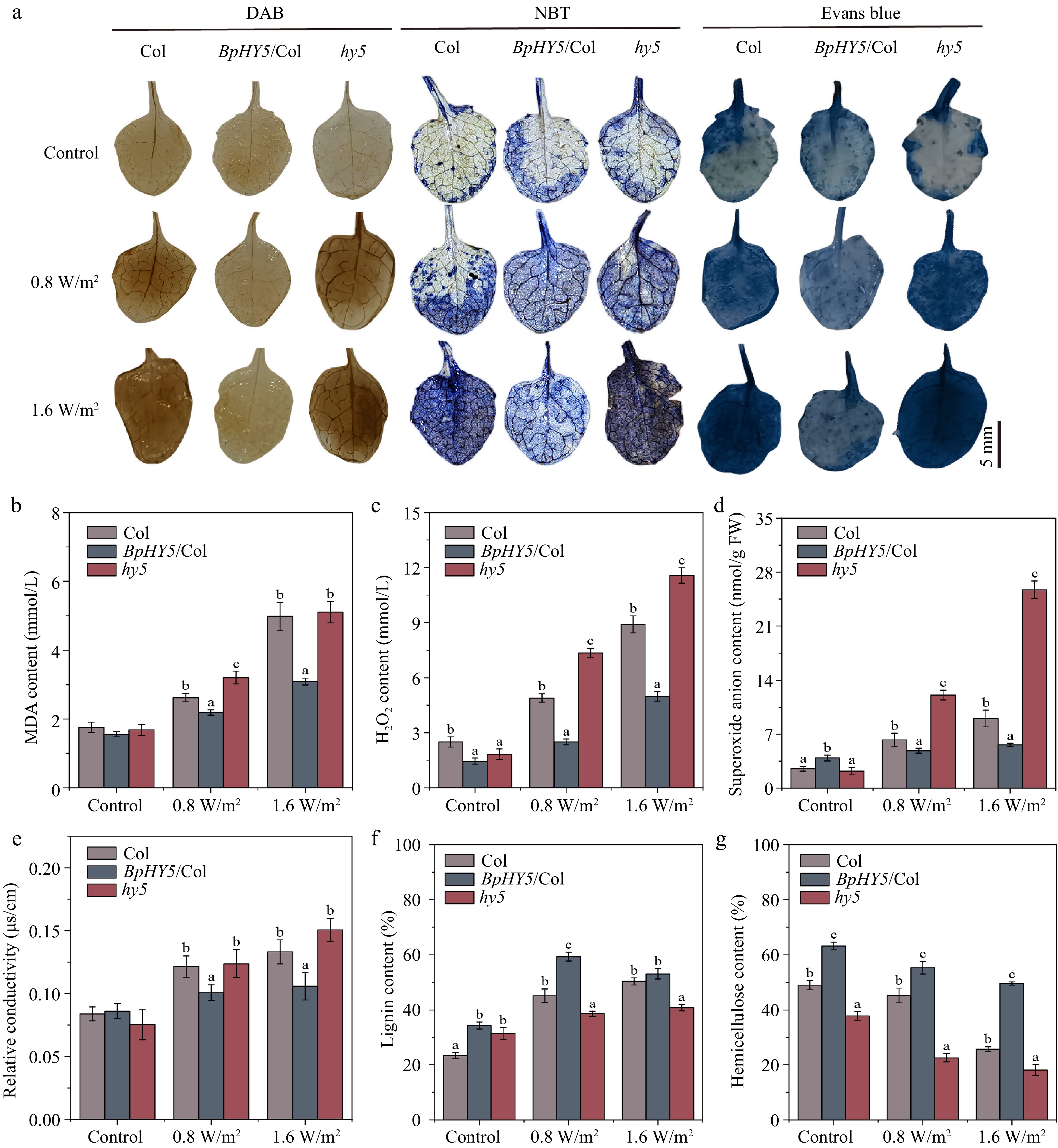
Figure 5.
BpHY5 affected the ROS accumulation and component content of stems under UV-B. (a) Determination of ROS content using DAB, NBT, and Evans blue staining. Scale bar represents 5 mm. The statistics of (b) MDA content, (c) hydrogen peroxide content, (d) superoxide anion content, and (e) relative conductivity. The statistics of component content of stems of Col, BpHY5/Col, and hy5 after 10 d of treatment of 0.8 and 1.6 W/m2 UV-B, including (f) lignin content and (g) hemicellulose content. Significance was determined by Student's t-test (p < 0.05).
Interestingly, under UV-B condition, the lignin content of BpHY5/Col was higher than that of Col (1.31-fold, 1.05-fold), whereas the lignin content of hy5 was lower than that of Col (0.85-fold, 0.81-fold) (Fig. 5f). Similarly, the hemicellulose content in BpHY5/Col was approximately 22.26% and 92.77% higher than that in Col, whereas the hemicellulose content in hy5 was 50.00% and 29.52% lower than that in Col (Fig. 5g). It is worth noting that under the control condition, the cellulose content of BpHY5/Col was increased by 18.53% compared to Col. However, the cellulose content of BpHY5/Col was only 9.49% higher than that of Col after 0.8 W/m2 UV-B treatment, whereas it was 11.44% lower than that of Col after 1.6 W/m2 UV-B treatment (Supplemental Fig. S2d). These results demonstrated that BpHY5 promoted the accumulation of lignin and hemicellulose.
BpHY5 is involved in the response to drought stress and ABA
-
The experiment exposed the seeds of BpHY5/Col and Col to different concentrations of mannitol and ABA to investigate the role of BpHY5 in drought response. The exogenous ABA was added to model dehydration reactions. BpHY5/Col had significantly higher germination and greening rates than Col when treated with mannitol and ABA (Fig. 6a−c). After 16 d, the fresh weight of BpHY5/Col increased 1.83-fold and 2.64-fold of Col, and the root length of BpHY5/Col was 46.50% and 57.43% higher than Col under 200 mM and 300 mM mannitol, respectively. The fresh weight of BpHY5/Col roots was 1.29 times and 1.34 times as large as Col, and the root length of BpHY5/Col increased 1.17-fold and 1.31-fold of Col after 0.5 μM and 1 μM ABA treatment. In addition, the drought stress caused curly and wilted leaves in both BpHY5/Col and Col, but Col exhibited more damage than BpHY5/Col. After 3 d of rewatering, the withered leaves of BpHY5/Col recovered, whereas most of the leaves of Col did not recover and died (Fig. 6i). These results indicated that overexpression of BpHY5 resulted in increased drought tolerance and reduced ABA sensitivity.
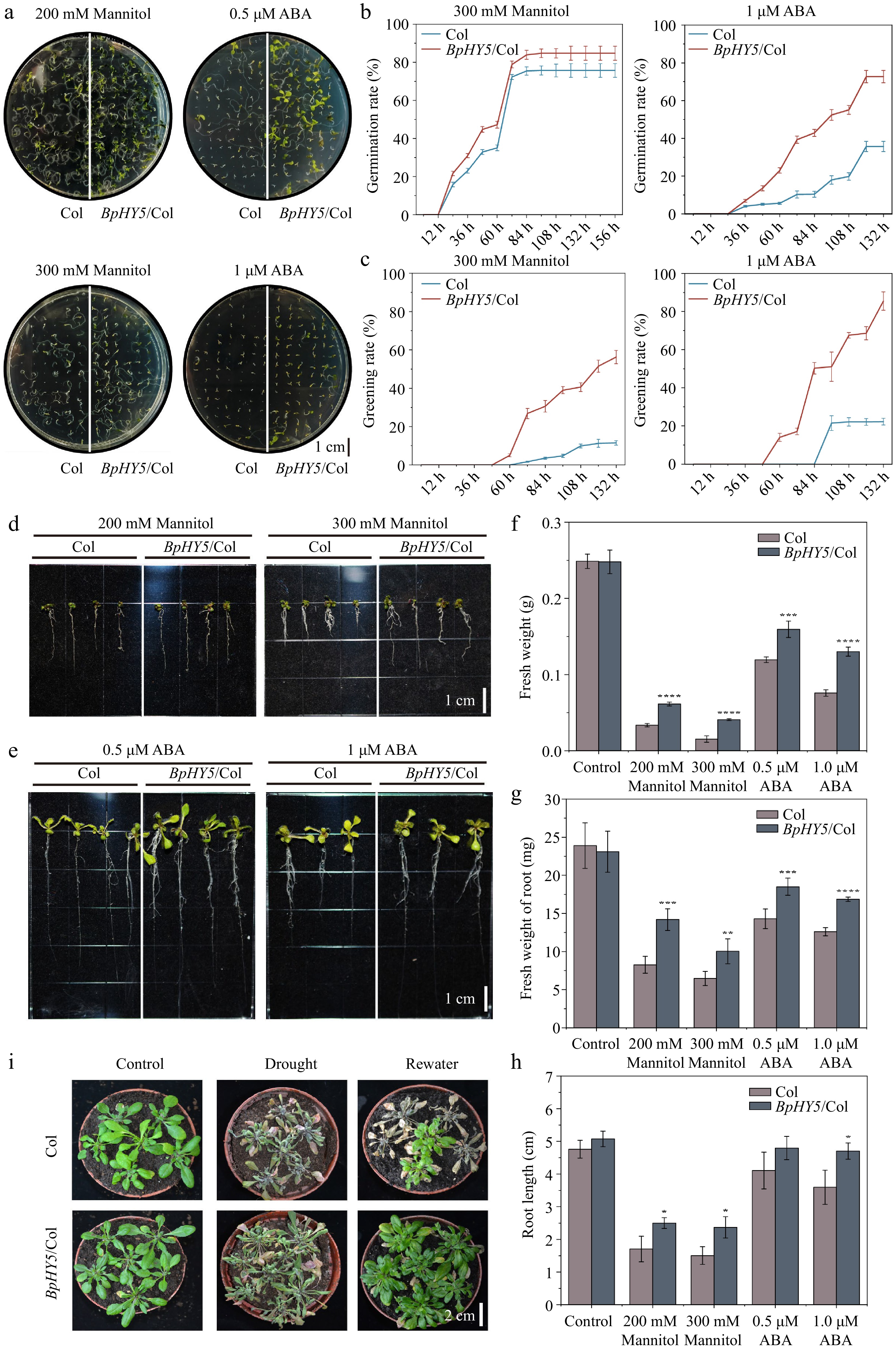
Figure 6.
BpHY5 decreased the sensitivity to drought and ABA in Arabidopsis. (a) Phenotypes of seed germination rate of Col and BpHY5/Col under 200, 300 mM mannitol and 0.5, 1.0 μM ABA at 7 d. Comparison of (b) germination rate and (c) greening rate of Col and BpHY5/Col under 300 mM mannitol and 1.0 μM ABA conditions. The root length phenotypes of Col and BpHY5/Col under (d) 200, 300 mM mannitol and (e) 0.5, 1.0 μM ABA treatments after 16 d. The statistics of the (f) fresh weight, (g) fresh weight of root, and (h) root length of Col and BpHY5/Col under mannitol (200, 300 mM) and ABA (0.5, 1.0 μM) treatment after 16 d. (i) Physical phenotypes of Col and BpHY5/Col subjected to drought stress without water for 7 d and re-watered for 3 d. Asterisks indicate levels of significance (t-test; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001). Scale bars: (a) = 1 cm; (d) = 1 cm; (e) = 1 cm; (i) = 2 cm.
Overexpression of BpHY5 improves drought and ABA resistance in Arabidopsis and birch
-
Plants exposed to mannitol and ABA were stained deeper by DAB and NBT than those under control conditions, suggesting that mannitol and ABA caused damage to plants (Fig. 7a). What stands out was that Col showed greater brown coloration and blue spots compared to BpHY5/Col under different stress conditions, demonstrating that the increased expression of BpHY5 resulted in improved activity of scavenging ROS. Water loss rate, MDA, superoxide anion content, relative conductivity, and H2O2 are important evaluation indicators for plant damage. The water loss rate of BpHY5/Col was lower than that of Col during the period of dehydration (Fig. 7b). The MDA content, superoxide anion content, and relative conductivity of BpHY5/Col were all significantly lower than those of Col under mannitol and ABA treatments (Fig. 7c).
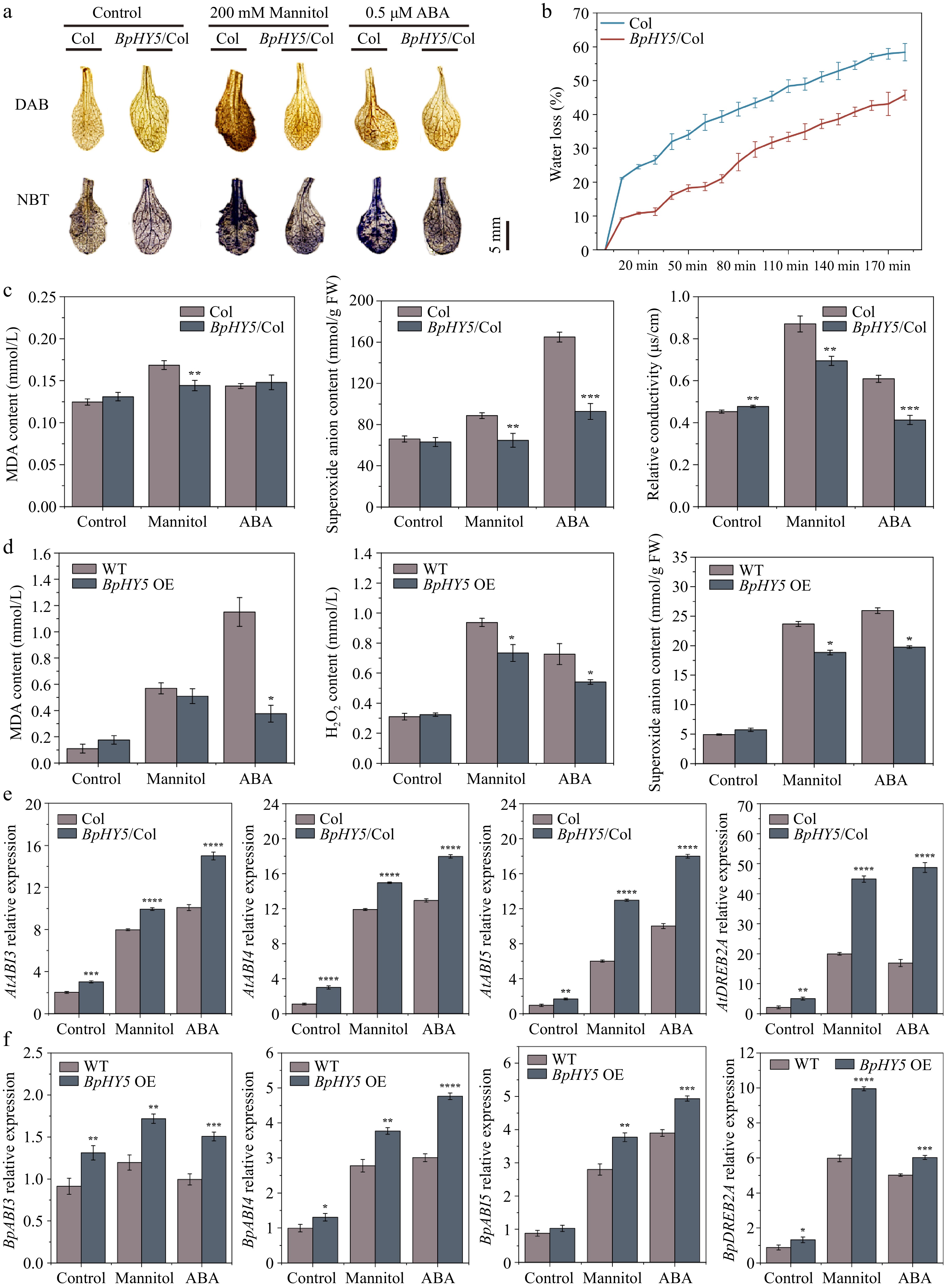
Figure 7.
Effect of BpHY5 overexpression on biochemical indicators and gene expression in Arabidopsis and birch under mannitol and ABA conditions. (a) DAB staining and NBT staining. Scale bar represents 5 mm. (b) Water loss rate of 16-day-old Col and BpHY5/Col seedlings. (c) The statistics of MDA content, superoxide anion content, and relative conductivity of 16-day-old Col and BpHY5/Col seedlings under mannitol and ABA conditions. (d) The statistics of MDA content, hydrogen peroxide content, and superoxide anion content of WT and transient transgenic birch under mannitol and ABA conditions. (e) The expression levels of AtABI3/4/5 and AtDEEB2A in Col and BpHY5/Col under mannitol and ABA conditions. (f) The expression levels of BpABI3/4/5 and BpDEEB2A in birch under mannitol and ABA conditions. Asterisks indicate levels of significance between Col and BpHY5/Col or WT and BpHY5 OE (t-test; *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
To further investigate the impact of BpHY5 on drought and ABA resistance in birch, birch seedlings were transiently transformed and treated with mannitol and ABA. For accurately determining the degree of damage, MDA, H2O2, and superoxide anion content were measured (Fig. 7d). There was a 10.53% and 67.36% decrease in MDA content of BpHY5/Col under mannitol and ABA treatments compared to Col, respectively. The H2O2 content of BpHY5/Col was 0.78-fold and 0.74-fold of Col under different stresses. Likewise, compared to Col, the superoxide anion content of BpHY5/Col decreased by 20.41% and 23.84% under different treatments. These results also demonstrated that BpHY5 participated in the plant responses to drought and ABA with inhibited membrane damage, improved ROS scavenging ability, and enhanced tolerance to drought and ABA.
Overexpression of BpHY5 affects the expression of stress-related genes
-
The expression of stress-related genes was evaluated to explore whether they could be induced by BpHY5 (Supplemental Fig. S2g, h). The expression levels of some B-box protein (BBX) transcription factors related to UV-B response (such as BBX4, BBX11, BBX21, BBX22) were up-regulated in transgenic Arabidopsis and birch. The expression of ABA biosynthesis-related genes was up-regulated in BpHY5 OE, for example, the expression levels of BpNCED3 and BpNCED9 in BpHY5 OE were 15.08-fold and 18.41-fold of WT. Some genes responded to abiotic stresses and ABA, such as MYB59, MYB74, ERD10, and RD20, were up-regulated in BpHY5/Col and BpHY5 OE. The transgenic plants showed significantly higher expression of ABA signaling pathway genes, including AtABI3/4/5, BpABI3/4/5, AtDREB2A, and BpDREB2A (Fig. 7e, f). Surprisingly, compared to control, the expression of BpDREB2A in birch overexpressing BpHY5 increased 7.54-fold and 4.57-fold under mannitol and ABA treatments. Furthermore, the expression of ROS signaling-related genes (such as ERF4, NDB2, and APX2) was up-regulated in transgenic Arabidopsis and birch. In particular, the expression level of BpERF4 in BpHY5 OE was 15.56-fold of WT. In summary, these results showed that overexpression of BpHY5 improved the expression of ABA signaling, ROS-responsive, and stress-responsive genes, promoted ABA signal transduction and enhanced the tolerance to UV-B and drought stress (Fig. 8).
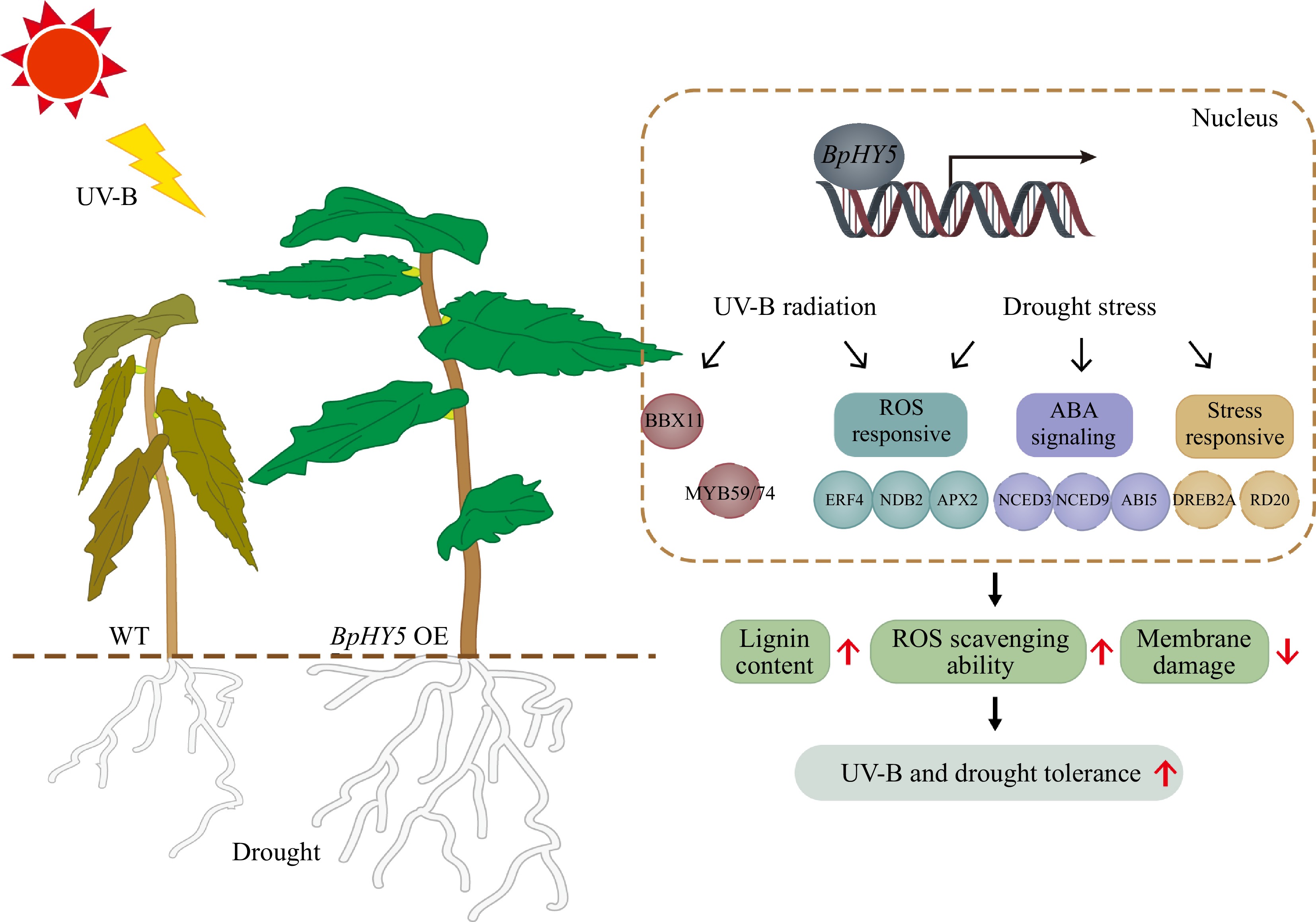
Figure 8.
Schematic model of UV-B and drought stress response of BpHY5. Overexpression of BpHY5 promotes the expression of BBX TFs, ABA-related genes (such as NCED3/6/9 and ABI3/4/5), increases the expression of stress-response genes (such as DREB2A, MYB59/74, ERD10 and RD20) and ROS-response genes (such as ERF4, NDB2 and APX2) to enhance the plant tolerance to UV-B and drought stresses with higher lignin content, greater ROS scavenging ability and lower membrane damage. It has been demonstrated that HY5 binds the promoter of TFs in the solid line through previous studies; however, the TFs represented by dotted lines have not been explicitly demonstrated.
-
HY5 is a member of bZIP TFs, which is involved in seed maturation, plant growth and development, light signal transduction, biosynthesis of plant secondary metabolites, and stress responses. In the present study, we cloned BpHY5 in birch and explored the functions of BpHY5 in response to UV-B radiation and drought stress.
BpHY5 enhances plant UV-B tolerance which may be related to the higher lignin content
-
UV-B radiation acts as a cue for photomorphogenesis as well as a stress factor, depending on the dose[15]. Arabidopsis has two pathways for responding to UV-B radiation: a non-specific pathway (high level of UV-B causing DNA damage and ROS production) and a specific pathway (mediated by low level of radiation)[16]. The BBX11-HY5 feedback loop regulates tolerance under UV-B by promoting the accumulation of antioxidants and inducing the expression of photoprotection (ELIP, CHI, CHS) and DNA repair genes (UVR2, UVR3)[17]. In Arabidopsis, the promoter of DEWAX is bound directly and negatively regulated by HY5 to decrease plant sensitivity to UV-B[6]. The study has identified BpUVR8 as a positive regulator in the photomorphogenesis mediated by UV-B in birch, positively regulating BpHY5[13]. According to our study, the expression level of BpHY5 was induced by UV-B (Fig. 2a). Both 0.8 and 1.6 W/m2 UV-B decreased the germination and greening rate of seeds, reduced the root elongation, fresh weight, and photosynthetic potential, and increased the ROS activity leading to the damage of cell membrane, suggesting that high level of UV-B inhibited the plant growth and development (Figs 3−5). The germination rate and greening rate of BpHY5/Col was higher than Col under 0.8 W/m2 UV-B. The fresh weight and root length of BpHY5/Col was significantly higher than Col under the UV-B condition. The leaves and plant size of BpHY5/Col were much more developmental than Col under VU-B, whereas that of hy5 was smaller than Col. Photosynthetic efficiency is affected by stress conditions. Fv/Fm is a parameter that reflects the health and growth by measuring plants' potential maximum photosynthetic capacity[18]. Fv/Fm of BpHY5/Col was higher under UV-B than Col, which indicated that overexpression of BpHY5 promoted photosynthetic potential. The MDA, H2O2, superoxide anion content, and relative conductivity of plants overexpressed BpHY5 was lower than Col, suggesting that BpHY5 regulated the tolerance to UV-B by increasing the ROS scavenging activity and decreasing the damage degree of the cell membrane.
The plant cell wall is a dynamic structure that provides mechanical support and participates in stress responses by regulating cell wall biosynthesis[19]. Previous studies have shown that various abiotic stresses, such as drought and waterlogging, can significantly affect the cell wall composition, and structural rearrangement and change the lignin content[20]. UV-B radiation stimulates the phenylpropanoid pathway and the cotyledons of Cucumis sativus L. accumulate more lignin[21]. It is thought that lignin deposition in the epidermis of quinoa acts as a protective mechanism against UV-B rays[22]. The transgenic rice with increased lignin content shows higher UV-B resistance[23]. ChS2, a type of Camellia sinensis L., with higher resistance against UV-B radiation, shows a lignin-like layer on the callus surface. Exposure to UV-B radiation results in a notable increase in lignin accumulation, indicating that lignin plays a crucial role in cellular protection against UV-B[24]. To further explore the mechanism of BpHY5 in regulating UV-B resistance, cell wall components were analyzed. In the present study, the lignin content of BpHY5/Col was higher than Col under control and UV-B conditions and the lignin content of hy5 was lower than Col under UV-B stress, which was in line with previous results (Fig. 5f).
BpHY5 functions as a stress-related transcription factor by regulating gene expression
-
Since the expression of over 3,000 genes can be directly or indirectly regulated by HY5, HY5 responds to various stresses and hormonal signals[25]. In Arabidopsis, the CRYPTOCHROME2 (CRY2)-COP1-HY5-BBX7/8 module regulates cold acclimation mediated by blue light[26]. The coordinated action of HY5 and MYB15 positively regulates cold tolerance by controlling the expression of CBFs in tomatoes[27]. HY5 and HISTONE DEACETYLASE 9 (HDA9) are degraded by high temperature, which releases the binding to the promoter of HSfA2 to promote gene expression and reduce salt tolerance[28]. However, previous research on responding to stresses to date has tended to focus more on roles in cold and salt tolerance than its effects on UV-B radiation and drought resistance.
In the present study, the BpHY5 protein was found to be located within the nucleus, suggesting that BpHY5 acted as a TF (Fig. 1). And BpHY5 might have regulated plant tolerance to abiotic stresses by controlling the expression of downstream genes. NtHY5 affects the lignification of interfascicular and vascular tissues[29]. MYB59 and MYB74 with HY5 binding sites are positively regulated by HY5 and associated with secondary cell wall synthesis[5,30]. In the present study, the expression levels of MYB59 and MYB74 were up-regulated in BpHY5/Col, which suggested that the strengthening of UV-B resistance mediated by BpHY5 may be connected to the increased lignin content and improved expression of genes associated with cell wall components biosynthesis.
Furthermore, we also confirm the expression levels of BBX genes. HY5 positively regulates the expression of BBX4/11/21/22, while negatively regulates BBX30/31 at the expression levels[31]. In Arabidopsis, BBX11 promotes the accumulation of antioxidants and induces the expression of DNA repair genes to protect plants from high levels of UV-B. HY5 binds to the promoter of BBX11 and overexpressing BBX11 has been found to alleviate the high sensitivity to UV-B radiation observed in hy5 to a certain extent[17]. Besides, BBX20/21/22 have been identified as rate-limiting cofactors of HY5[32]. In the current research, the expression of AtBBX11, BpBBX4, BpBBX21, and BpBBX22 were up-regulated in transgenic Arabidopsis and birch, which was consistent with previous research results (Fig. 7). These results identified that BpHY5 was a positive transcription factor of plant tolerance to UV-B stress by regulating related-genes expression, such as MYB59, MYB74, and BBX11.
BpHY5 improves plant tolerance to drought which may be related to ABA biosynthesis and signal transduction
-
In tomatoes, HY5 plays a key role in green light-induced drought responses[33]. In the present study, analysis of cis-acting elements showed the promoter of BpHY5 contained stress response elements and hormone response elements (such as abscisic acid), in which ABRE was an ABA-response element and always involved in ABA signal transduction (Fig. 1f). To explore the gene function of BpHY5 in drought stress, transgenic plants were treated by mannitol and ABA (exogenous ABA was added to model dehydration reactions). The germination and greening rates, fresh weight, and root length of BpHY5/Col were significantly higher than Col under mannitol and ABA-added conditions. The plants of BpHY5/Col recovered after 3 d of rewatering, whereas most of Col did not recover and died (Fig. 6). Furthermore, the MDA, superoxide anion, H2O2 content and relative conductivity of BpHY5 OE were decreased compared to wild type under drought treatment (Fig. 7). These results suggested that overexpression of BpHY5 enhanced the plant tolerance to drought and decreased sensitivity to ABA by increasing ROS scavenging activity and decreasing membrane damage.
According to the results of qRT-PCR, it was noted that there were some differentially expressed genes associated with ABA signaling. ABA plays a vital role in the adaptation of plants to environmental stresses, especially young seedlings. NCED is regarded as the rate-limiting step of ABA biosynthesis[34]. NCED3 promotes ABA accumulation and cooperates with NCED5 to protect against drought stress[35,36]. Previous research has shown that AtNCED6 and AtNCED9 are necessary for ABA biosynthesis during seed development[37]. Analysis of HY5 genomic binding sites indicated that AtNCED3 and AtNCED9 are genes positively regulated by AtHY5[5]. As expected, our data showed that AtNCED6, AtNCED9, BpNCED3, and BpNCED9 were up-regulated in overexpression lines (Fig. 7). Especially, the expression of BpNCED3 and BpNCED9 in birch overexpressed was 15.08-fold and 18.41-fold of WT, respectively.
ABI3/4/5 have functions that overlap in ABA signaling transduction. ABI5 is the crucial positive regulator in ABA signal. HvABI5 regulates the drought response in an ABA-dependent manner[38]. Transgenic cotton expressing AtABI5 exhibited higher resistance to drought and increased photosynthesis with greater root system and leaf area[39]. HY5 and ABI5 physically interact in response to ABA signaling in Arabidopsis, and HY5 positively regulates ABA signaling[40]. The studies proved that HY5 binds to the promoter of ABI5 to initiate the expression of ABI5 and ABI5-targeting genes[7,41]. The present findings indicate that BpHY5 overexpression resulted in a significant increase in AtABI3/4/5, and BpABI3/4/5 expression levels than WT plants under control, drought, and ABA conditions, and the expression of AtABI5 (7.71-fold) and BpABI5 (3.70-fold) was induced by drought (Fig. 7). These findings indicated the enhancement of drought resistance mediated by BpHY5 may be connected to the expression increase of genes associated to ABA, including NCED3, NCED9, and ABI5.
DREB2A and DREB2B are hypothesized to be the main TFs activated by drought stress. Overexpression of DREB2A significantly improves drought resistance in Arabidopsis and enhances drought, heat, and salinity tolerance in Pennisetum glaucum[42,43]. RD20 is an ABA-responsive gene and participates in drought tolerance mechanisms by regulating stomatal aperture and plant growth[44]. The analysis results showed that DREB2A and RD20 have the binding sites of HY5, and they are predicted to be the target genes of HY5[5]. However, there is no direct evidence of an interaction at present. Furthermore, previous studies document that HY5 binds to the promoter of ROS-responsive genes[45]. ETHYLENE-RESPONSIVE TRANSCRIPTION FACTOR4 (ERF4), NAD(P)H dehydrogenase B2 (NDB2), Ascorbateperoxidase2 (APX2), belonging to ROS signaling pathways, are confirmed to be the key genes targeted by HY5[45]. The results of Sweetman et al. also suggest HY5 mediated the transcription of NDB2[46]. The expression level of APX2 is downregulated in the hy5 mutant, which further proves that APX2 is the downstream target gene of HY5[47]. Our results showed that the expression of the genes mentioned was up-regulated in transgenic Arabidopsis and birch. Especially, the expression level of ERF4 in transgenic birch was 15.56-fold of WT (Fig. 7). The above results suggested that BpHY5 enhanced drought tolerance by increasing transcript expression of stress- and ROS-responsive genes, including DREB2A, RD20, ERF4, NDB2, and APX2. However, we are also aware of several limitations of this study and the molecular mechanisms of BpHY5 in UV-B and drought responses need to be further studied.
-
In summary, BpHY5 is a transcription factor of the bZIP family, which was highly induced by UV-B, drought, and ABA in the nucleus. Overexpression of BpHY5 increased UV-B and drought tolerance and decreased ABA sensitivity, with longer roots, stronger ROS scavenging ability, lower damage degree, and higher stress-related gene expression levels. Overall, the present study indicated that BpHY5 plays an important role in the plant stress endurance process, which suggests BpHY5 is a promising candidate gene for birch breeding and provides a basis for further investigation of BpHY5 functions in plant tolerance.
-
The authors confirm contribution to the paper as follows: study conception and design: Xin Y, Zeng F; draft manuscript preparation and analysis of results: Gao S; interpretation of results: Chen X; preforming the experiments and data collection: Lin M, Yin Y, Li X; manuscript modification: Zhan Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was financially supported by the National Natural Science Foundation of China (31870588), the Fundamental Research Funds for the Central Universities (2572021DT06), the Innovation Project of State Key Laboratory of Tree Genetics and Breeding (Northeast Forestry University) (2022A03), and the Heilongjiang Touyan Innovation Team Program (Tree Genetics and Breeding Innovation Team).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primer sequences.
- Supplemental Fig. S1 (a) The expression level of BpHY5 at 0, 3, 6, 9, 12, 15, 18 and 21 h in one day. (b) Phenotypes of seedlings of Col, hy5, BpHY5/hy5, and BpHY5/Col under visible light or darkness for 7 d. Scale bar represents 1 cm. (c) Hypocotyl length of Col, hy5, BpHY5/hy5, and BpHY5/Col under visible light or darkness for 7 d. Data are represented as the mean ± SE of three biological replicates; statistical significance (p < 0.05) is indicated by a, b, c, d, e, f.
- Supplemental Fig. S2 (a) Expression levels of BpHY5 in different transgenic lines of Arabidopsis. Statistical significance (p < 0.05) is indicated by a, b, c, d, e. (b) The relative expression level of BpHY5 in transient transgenic birch and wild type. Data were represented as the mean ± SE of three biological replicates; asterisks indicate levels of significance (t-test; ****p < 0.0001). (c) The statistics of fresh weight of Col, BpHY5/Col, and hy5 after 10 days of treatment of 0.8 W/m2 and 1.6 W/m2 UV-B. (d) The statistics of cellulose content of stems of Col, BpHY5/Col, and hy5 after 10 days of treatment of 0.8 W/m2 and 1.6 W/m2 UV-B. Data are represented as the mean ± SE of three biological replicates; statistical significance (p < 0.05) between Col, BpHY5/Col, and hy5 is indicated by a, b, c. (e) Phenotypes of seed germination rate of Col and BpHY5/Col under control condition at 7 d. (f) Phenotypes of root length of Col and BpHY5/Col under control condition at 16 d. (g, h) Heatmap of stress-related genes expression of control, transgenic Arabidopsis (g), and transient transgenic birch (h).
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gao S, Chen X, Lin M, Yin Y, Li X, et al. 2024. A birch ELONGATED HYPOCOTYL 5 gene enhances UV-B and drought tolerance. Forestry Research 4: e022 doi: 10.48130/forres-0024-0019 

A birch ELONGATED HYPOCOTYL 5 gene enhances UV-B and drought tolerance
- Received: 25 November 2023
- Revised: 14 April 2024
- Accepted: 21 May 2024
- Published online: 19 June 2024
Abstract: UV-B radiation and drought majorly restrict plant growth, particularly in summer. ELONGATED HYPOCOTYL 5 (HY5), a bZIP transcription factor (TF), has a beneficial impact on photomorphogenesis. However, the sequence of HY5 from Betula platyphylla (BpHY5) has not been identified and the gene functions remain unclarified. We cloned the sequence of BpHY5, which was targeted to the nucleus. The hypocotyl phenotypes of heterologous expression in Arabidopsis thaliana and reverse mutation showed that BpHY5 is homologous to AtHY5. The expression of BpHY5 was increased in response to UV-B radiation, drought conditions, and the presence of abscisic acid (ABA). The overexpression of BpHY5 resulted in increased tolerance to UV-B radiation and drought and decreased ABA sensitivity with higher germination and greening rate, more developmental root system, stronger reactive oxygen species scavenging ability, and lower damage degree. The lignin content under UV-B condition of BpHY5/Col was higher than that of Col. Furthermore, overexpressing BpHY5 up-regulated the expression of genes related to tolerance (NCED3/9, ABI5, DREB2A, RD20, ERF4, NDB2, and APX2). In brief, the study suggests that BpHY5 from birch serves as a beneficial modulator of plant responses to UV-B radiation and drought stress.
-
Key words:
- ABA /
- Betula platyphylla /
- BpHY5 /
- bZIP /
- Drought stress /
- UV-B