









-
In eukaryotes, chromosome DNA with a long linear length needs to be packaged into the nucleus, which presents a topological problem. To this end, the nucleosome is crucially needed in packaging and condensing the genome as the basic structural unit of chromatin[1]. The canonical nucleosome is an octamer composed of two copies of each of the four canonical histones (H3, H4, H2A and H2B), surrounded by 147 bp DNA. The dense structure of chromatin is conducive to the storage of genetic information and stable inheritance, but it also causes obstacles for DNA elements to be exposed in a regulated manner during gene transcription, DNA replication and DNA repair. In this case, nucleosome is further considered as a highly dynamic unit through which the genome can be epigenetically regulated, such as DNA methylation, histone modification, non-coding RNA and ATP-dependent chromatin remodeling[2]. ATP-dependent chromatin remodeling complexes participate in various regulatory processes to regulate gene expression, transcription, replication, and recombination, such as loosening DNA entanglement by disrupting the interaction between DNA and histones, loosening DNA tangles, slipping nucleosome DNA, expelling histone dimers or octamers, and exchanging histone dimers[3].
SWI/SNF (switch defective/sucrose nonfermentable) complexes, a class of the most exhaustively studied ATP-dependent chromatin remodelers so far, can regulate the accessibility of transcription factors or other DNA-binding proteins to DNA by altering the interaction between DNA and histone octamers. Although the identification and characterization of chromatin remodeling complexes in plants lag behind yeast and animals, recent studies have revealed whether and how chromatin remodelers in Arabidopsis function in multi-subunit complexes[4,5,6,7]. Otherwise, conserved and plant-specific subunits of chromatin remodeling complexes have been identified and characterized which provided a basis for further study of the molecular mechanisms of chromatin remodeling complexes in plants[4]. Due to the importance of these remodeling complexes, the absence of core accessory subunits usually leads to severe developmental defects[8]. As the component of the minimal functional core of the conserved SWI/SNF complex, SWI3 subunit discovered in Arabidopsis (four AtSWI3 proteins) may exert non-redundant regulatory functions by participating in the assembly of other SWI/SNF complexes, affecting the nutritional and reproductive stages of embryogenesis and plant development, which shows remarkable functional diversification[5]. Even though plants have been at the forefront of understanding the biological role of chromatin remodeling factors in growth, development and stress response through forward genetics[6], there are few related studies on fruit trees. In recent years, 11 AcSWC subunits (SWI2/SNF2-Related 1 chromatin remodeling complex) in pineapple has been identified and AcSWC6 was characterized as involved in the stress response in plants[7,9]. Four SlSWI3s were identified and characterized in tomato, in which SlSWI3B could function on gene regulating in reproductive development, and SlSWI3C may participate in vegetative growth[10]. Nevertheless, there is no relevant research in citrus to date which suggests the identification and mechanical analysis of SWI/SNF complexes needed to be deeply explored.
Citrus is one of the most widely cultivated fruit crops with important economic and nutritional value around the world. Sugars and organic acids are considered to be significant contributors to flavor quality among the major compounds in citrus fruits, and the sugar/acid ratio is recognized as the major determinant of sweetness and ripeness[11]. In this study, we identified four SWI3 family genes in Citrus sinensis. All members of the CsSWI3 gene family were comprehensively analyzed, including physicochemical properties, phylogenetic relationships, chromosome distribution, subcellular localization, conserved motifs and domains, gene structure, promoter cis-regulatory elements, expression profiles in different development periods of fruit and other tissues in 'Bingtang' orange, and correlation between their expression and accumulation of sugar and organic acids in citrus fruits. In general, our works provide a basis for further study on chromatin remodeling dominated by SWI/SNF complex and its core accessory subunits in citrus.
-
The 'Bingtang' sweet orange (Citrus sinensis) materials include fruit, stem, leaf and flower used in gene expression analysis were obtained from Quzhou, Zhejiang Province, China. Four developmental stages were selected: 105, 135, 165 and 195 DAFB (days after full bloom) in 2021. Fruits with uniform size and appearance were selected in each sample point as biological replicates. Fruit flesh, stems, leaves and flowers were frozen in liquid nitrogen and stored at −80 °C for further analysis.
Identification of SWI3 family in C. sinensis genome
-
Four AtSWI3 protein sequences in Arabidopsis thaliana[5] were used to retrieve the SWI3 family members in sweet orange based on its genome database (
http://citrus.hzau.edu.cn/download.php ) through the Blast function in TBtools software. Conserved domains of the putative SWI3 family members were analyzed on Pfam database (http://pfam.xfam.org ) and visualized by TBtools software[12,13]. The members containing SWIRM domain (Pfam: PF04433), SWIRM-associated region 1 domain (Pfam: PF16495) and SANT domain (Pfam: PF00249) were finally identified as SWI3 family protein sequences. Analysis of physiological and biochemical properties including molecular weight (MW), theoretical isoelectric point (pI), grand average of hydropathicity (GRAVY) and instability index (II) was achieved by uploading CsSWI3 protein sequences to the ExPASy database (www.expasy.org/tools/protparam.html/ )[14].Phylogenetic analysis and chromosomal location of SWI3 family
-
Phylogenetic analysis of SWI3 family proteins in sweet orange, Arabidopsis, rice and tomato was performed using MEGA11 software[15]. Clustal W was used to perform multiple sequence alignment and the phylogenetic tree was constructed based on the maximum likelihood method with 1,000 bootstrap tests[16]. Moreover, iTOL (
https://itoleditor.letunic.com/ ) was used to ornament the final phylogenetic tree[17]. The genome annotation file of sweet orange in gff format downloaded on its genome database (http://citrus.hzau.edu.cn/download.php ) and visualized by TBtools software was used to analyze the chromosomal localization of CsSWI3s.Gene structure, conserved motif and domain analysis of CsSWI3s
-
Gene structure information of SWI3s in sweet orange and Arabidopsis were involved in their genome annotation files respectively. MEME Suite 5.5.2 tool (
https://meme-suite.org/meme/tools/meme ) was used to analyze the conserved motif of CsSWI3s based on their protein sequences[18]. In addition, conserved domain analysis was achieved by Pfam database. All the analysis results above were illustrated by TBtools.Subcellular localization of CsSWI3 genes
-
The CDS (coding sequence) of CsSWI3 genes without stop codon were constructed in the pCAMBIA1300-35S-eGFP vector and electroporated into Agrobacterium tumefaciens GV3101[19]. The cultures of Agrobacterium in appropriate amount were suspended in infiltration buffer (10 mM MES, 10 mM MgCl2 and 150mM AS) with an OD600 of 0.75 and was transiently expressed in transgenic N. benthamiana (expressed H2B-RFP as a nuclear marker) leaves by a needleless syringe. After 2 d of growth in the greenhouse with a light/dark cycle of 16:8 h at 24 °C, the injection area was imaged with a confocal laser scanning microscope (Nikon 578 A1-SHS, Tokyo, Japan).
Promoter cis-regulatory elements analysis of CsSWI3s
-
The Plant CARE database (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) was used to analyze the promoter cis-regulatory elements of CsSWI3s based on the 2kb upstream sequences extracted from their genomic nucleotide sequences with Phytozome 13 database (https://phytozome-next.jgi.doe.gov )[20,21]. Furthermore, the analysis results were visualized by TBtools.RNA extraction and cDNA synthesis
-
Total RNA was extracted with RNAiso Plus (Takara, Beijing, China) kit following the manufacturer's protocol. Subsequently, gDNA wiper (Vazyme, Nanjing, China) was used to remove genomic DNA from 1ug of extracted total RNA, and the first strand cDNA was synthesized using HiScript® II qRT SuperMix (Vazyme).
Expression analysis of CsSWI3s
-
Primer sequences for RT-qPCR were designed by the Primer-Blast tool in NCBI website (
https://www.ncbi.nlm.nih.gov/tools/primer-blast/ ). The concentration of cDNA template was adjusted according to the CT value of Actin gene (Ciclev10025866m). The RT-qPCR reaction mixture (20 μl) included 10 μl ChamQ Universal SYBR qPCR Master Mix, 2 μl diluted cDNA, 0.4 μl of each gene-specific primer (10 μM) and 7.2 μl DEPC-treated water. The reaction was performed using Ssofast Eva Green Supermix Kit and CFX96 instrument (Bio-RAD) with following program: initiated at 95 °C for 5 min, followed by 50 cycles of 95 °C for 10 s, 60 °C for 10 s and 75 °C for 15 s. Melting curves with the citrus Actin gene (XM_006464503) as internal control were used to examine each gene at the end of each run[22]. The 2−ΔCᴛ method was used to analyze the gene expression levels. Primers used for RT-qPCR are listed in Supplemental Table S1.Statistical analysis
-
Data analysis was performed using Microsoft Excel and Graph Pad Prism 9. All experiments included at least three biological replicates in this research and the error bar represented standard error (SE). The least significant difference (LSD) between different developmental stages was calculated (p < 0.05). Figures were drawn using Graph Pad Prism 9 and Microsoft PowerPoint.
-
A total of four CsSWI3 genes were identified from the C. sinensis genome by homologous sequence alignment with the AtSWI3 genes in Arabidopsis. All of the CsSWI3 proteins contain SWIRM domain (Pfam: PF04433), SWIRM-associated region 1 domain (Pfam: PF16495) and SANT domain (Pfam: PF00249) which have been reported in Arabidopsis before[6]. Bioinformatic analysis showed that the CsSWI3 proteins contained 466 to 1038 amino acids with their molecular weights (MW) ranging from 52.00 to 112.70 kDa, theoretical isoelectric point (pI) from 4.77 to 5.49 and the instability index (II) from 35.42 to 50.95, and all of them were hydrophilic proteins. (Supplemental Table S2)
Phylogenetic analysis and chromosomal location of SWI3 family
-
A phylogenetic tree has been constructed in order to elucidate the evolutionary relationships between CsSWI3s and the same family members in other species reported to date, including four AtSWI3s in Arabidopsis thaliana, six OsSWI3s in Oryza sativa (rice) and four SlSWI3s in Solanum lycopersicum (tomato). The 16 SWI3 proteins were classified into four subfamilies (SWI3A to SWI3D) predicated on previous studies in Arabidopsis[5], which is also responsible for the naming of CsSWI3s (Fig. 1). Each subfamily contains one CsSWI3 member (CsSWI3A, CsSWI3B, CsSWI3C, CsSWI3D) respectively.

Figure 1.
Phylogenetic relationship of SWI3 family. Branches in purple, red, blue and yellow represent the subfamily SWI3A to SWI3D respectively. Different colored marks next to the labels depict different species: red pentagram, sweet orange; purple circle, Aridopsis; blue square, rice and green triangle, tomato.
The results of chromosomal localization analysis showed that four CsSWI3s distributed on only three chromosomes of the C. sinensis genome: CsSWI3A on chr7, CsSWI3B and CsSWI3C on chr3, CsSWI3D on chr8 (Fig. 2).
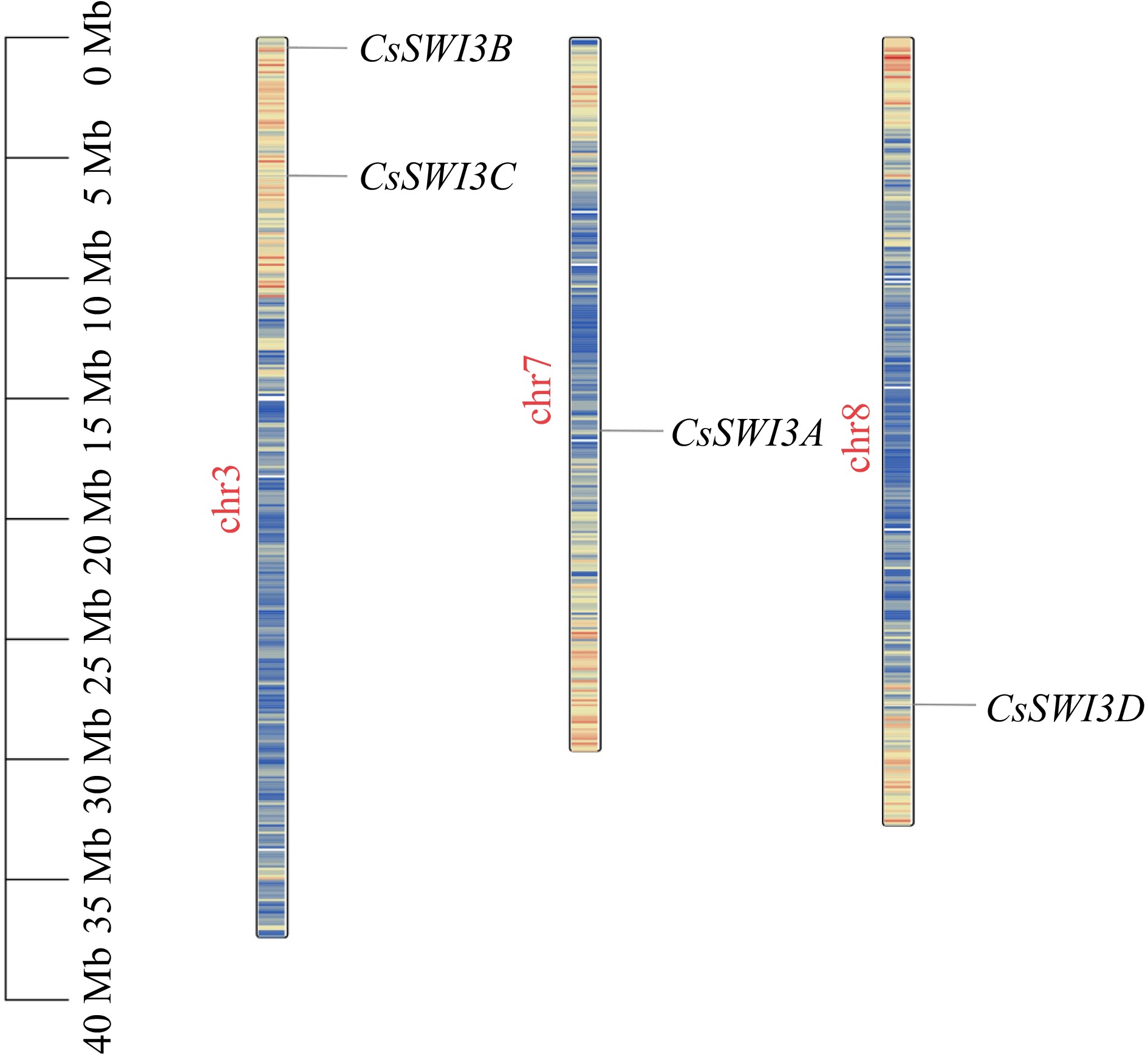
Figure 2.
Chromosomal distribution of CsSWI3s. Color of the chromosome represents the gene density.
Gene structure, conserved motif and domain analysis of CsSWI3s
-
Gene structure analysis shed lights on the exons and introns framework of AtSWI3s and CsSWI3s and may suffice for the further understanding of evolutionary relationships of CsSWI3 family. The number of exons within CsSWI3s ranges from 5 to 8 (Fig. 3). Meanwhile, CsSWI3s in the same cluster with AtSWI3s have the same quantity of exons with the exception of CsSWI3B, which further confirms the results of the phylogenetic analysis.
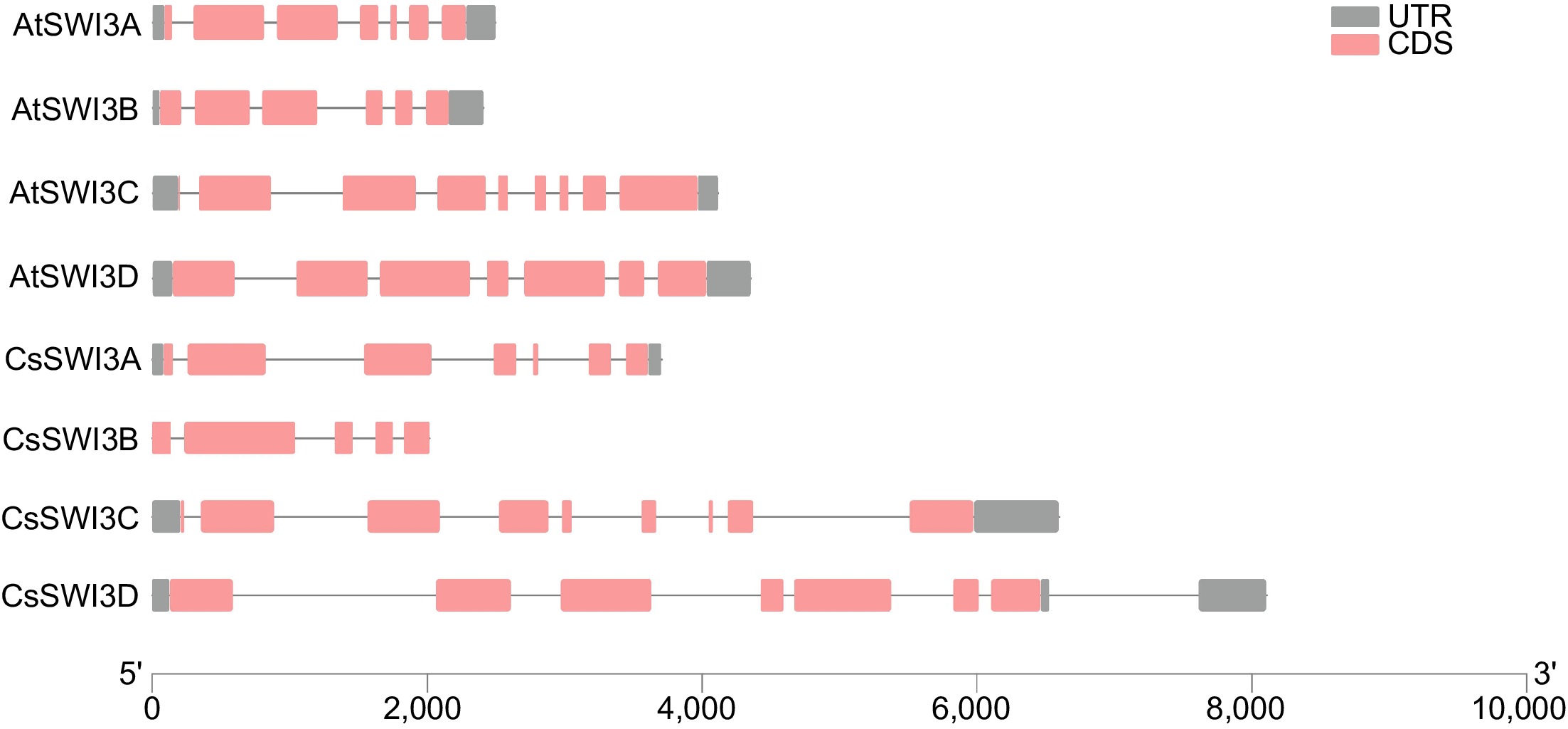
Figure 3.
Gene structure of CsSWI3s and AtSWI3s. Red and gray blocks represent exons and untranslated regions (UTRs) respectively, black lines represent introns.
A total of ten motifs were identified (Fig. 4b) and their logos predicted by MEME database are displayed in Fig. 4d. Except for SlSWI3B, all of the SWI3 family proteins contain SWIRM domain (Pfam: PF04433), SWIRM-associated region 1 domain (Pfam: PF16495) and SANT domain (Pfam: PF00249), besides that, only the members in cluster SWI3D contain a Zinc finger-ZZ type domain (Fig. 4c). Accordingly, proteins with higher homology have more analogous conserved motifs and domains, presumably performing similar functions.

Figure 4.
Conserved motif and domain analysis of SWI3 proteins. (a) Phylogenetic tree of 18 SWI3 proteins. (b) Conserved motifs distribution of SWI3 proteins. Blocks in different colors refer to different motifs. (c) Conserved domains distribution of SWI3 proteins. Different domains are presented in different colored blocks. (d) Ten conserved motifs with the consensus sequences predicted by MEME.
Subcellular localization of CsSWI3 genes
-
A subcellular location assay of CsSWI3 was performed by expressing in Nicotiana benthamiana leaves to investigate the cellular location. Strong green fluorescence of CsSWI3s was observed in the cell nucleus, meanwhile, the control fluorescence signals were widely observed in nucleus, cytoplasms and members, which indicated that four CsSWI3 proteins were all located in the nucleus (Fig. 5).
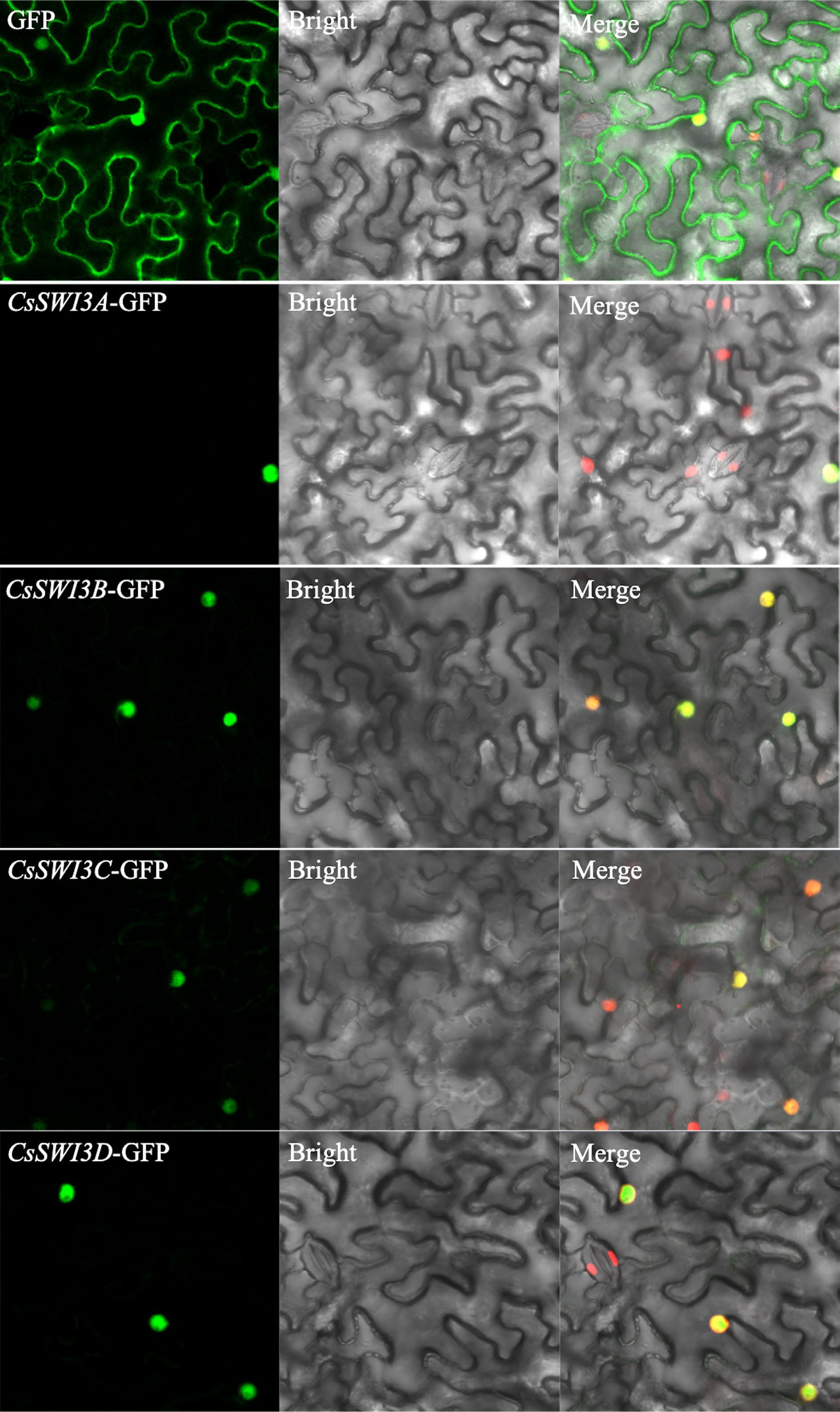
Figure 5.
Subcellular localization of CsSWI3s. Bar = 20 μm.
Promoter cis-regulatory elements analysis of CsSWI3s
-
A total of 84 promoter cis-regulatory elements within the first 2 kb fragment upstream of CsSWI3s were predicted and divided into three categories based on their functions: phytohormone response, plant growth and development and stress response, which containing 18, 8 and 58 elements respectively. With the largest amount, stress response elements including 14 for anaerobic induction, four for drought-inducibility, one for defense and stress responsiveness, 37 for light responsiveness and two for low-temperature responsiveness, indicating that CsSWI3s presumed to play a significant role in the stress resistance of sweet orange. Otherwise, phytohormone response elements (including one for ABA responsiveness, one for auxin responsiveness, four for gibberellin responsiveness, and 12 for MeJA responsiveness) and plant growth and development elements (including two for circadian control, two for cell cycle regulation, two for differentiation of the palisade mesophyll cells and two for meristem expression) predicted on CsSWI3 promoters indicated that phytohormone could regulate CsSWI3 genes expression to some extent and CsSWI3s played a minor role during plant growth and development (Fig. 6). Exact information of promoter cis-regulatory elements on CsSWI3s is displayed in Supplemental Table S3.
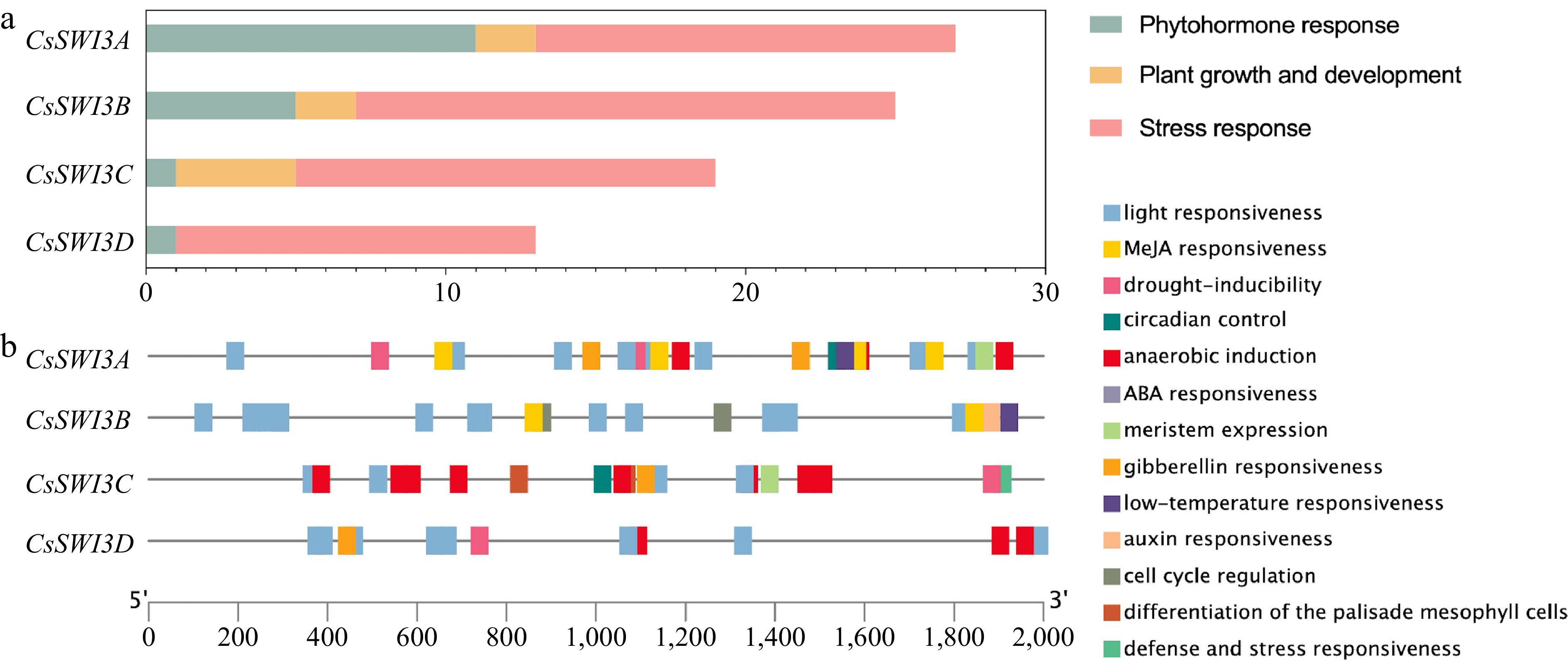
Figure 6.
Promoter cis-regulatory elements analysis of CsSWI3s. (a) The number of three types of cis-regulatory elements on CsSWI3 genes promoter. (b) The cis-regulatory elements within the first 2000 bp fragment upstream of CsSWI3s. Blocks in different colors refer to different elements.
Expression analysis of CsSWI3s in different development periods of fruits and different tissues in 'Bingtang' sweet orange
-
Soluble sugars, including fructose, glucose and sucrose, gradually accumulated during fruit development periods, with a peak of 31.55 mg/g, 31.40 mg/g, 34.42 mg/g at 195DAFB. As the main organic acids in 'Bingtang' sweet orange, citric acid content increased during the fruit early developmental stages (peaked at 135 DAFB) and decreased afterwards till the maturity stage (195 DAFB), while malic acid had slight changes during development (Fig. 7).
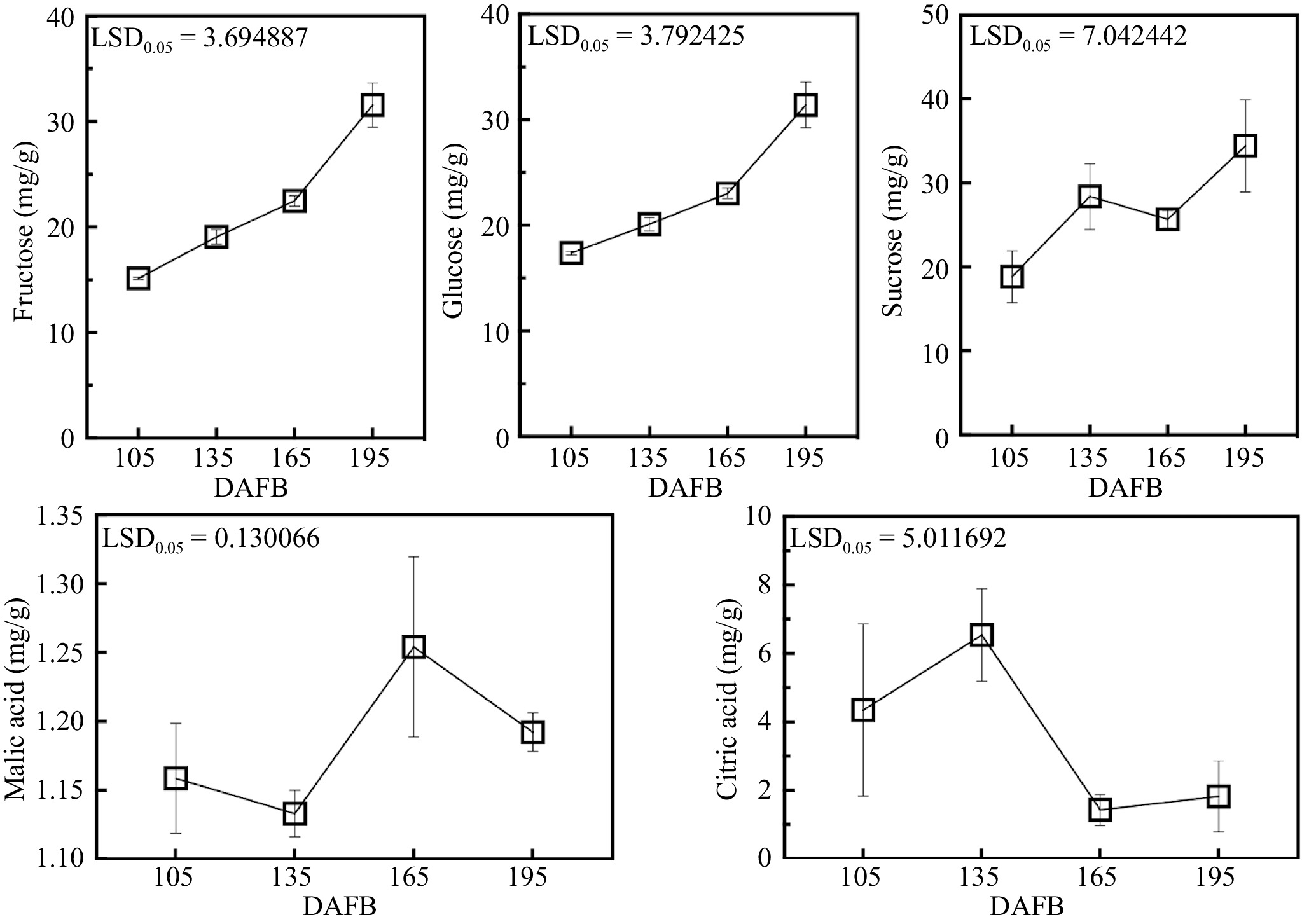
Figure 7.
Sugar (fructose, glucose and sucrose) and organic acid (malic acid and citric acid) content in fruit flesh during different develop periods of 'Bingtang' sweet orange. Error bars represented SE (n = 3), LSD represents the least significant difference between different developmental stages.
In order to further investigate the role of CsSWI3s during the development of 'Bingtang' sweet orange, we analyzed the expression patterns of CsSWI3s in the fruit flesh during different development periods and three other tissues (stem, leaf and flower) (Figs 8, 9). As shown in Fig. 8, all CsSWI3 genes expressed in the four development periods in 'Bingtang' sweet orange fruit and CsSWI3B displayed the highest expression level.
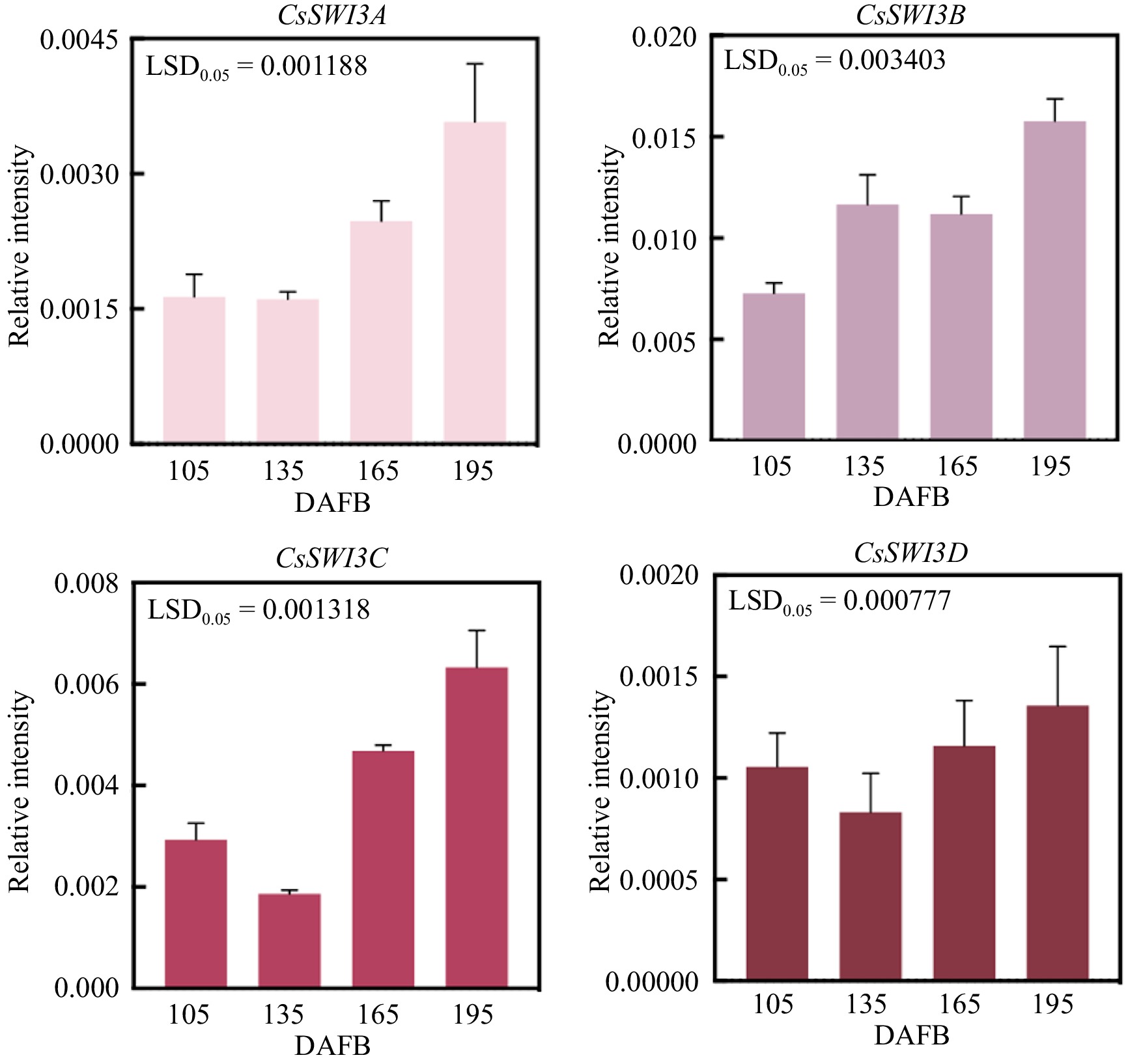
Figure 8.
Gene expression of CsSWI3s at 105, 135, 165 and 195 DAFB in 'Bingtang' sweet orange fruit. Error bars represent SE (n = 3), LSD represents the least significant difference between different developmental stages.
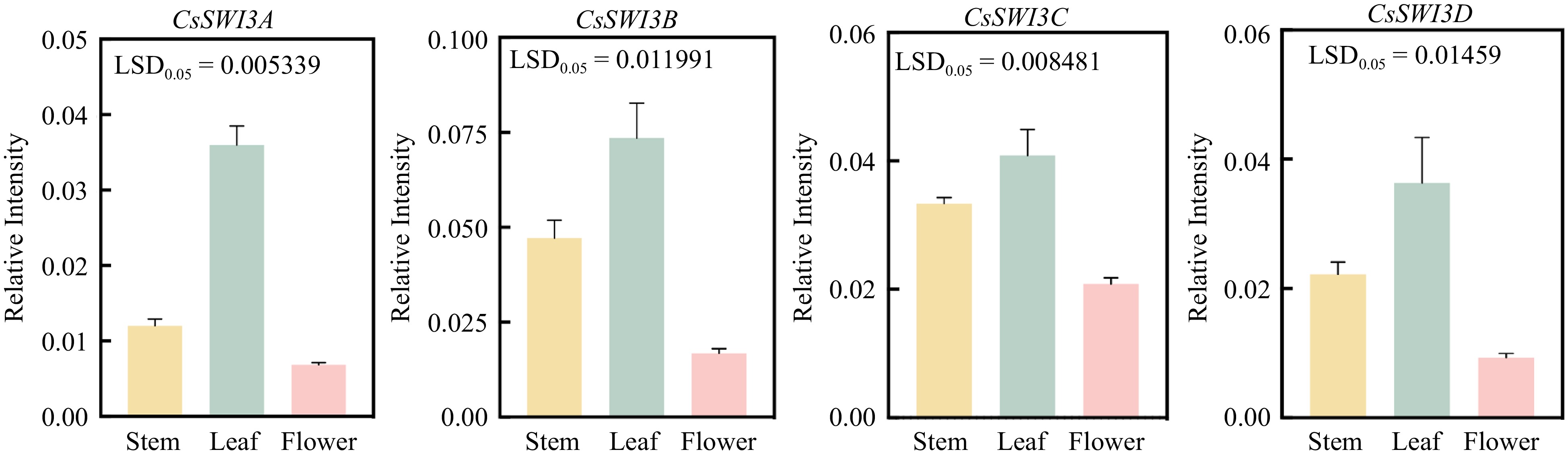
Figure 9.
Gene expression patterns of CsSWI3s in different tissues of 'Bingtang' sweet orange. Error bars represent SE (n = 3), LSD represents the least significant difference between different tissues.
This fact apart, we found that CsSWI3s showed different expression patterns in different tissues of sweet orange. The highest expression level of all the CsSWI3s was found in the leaf, and CsSWI3B still displayed the highest level among all of them (Fig. 9).
In order to study the relationship between CsSWI3 genes and sugar as well as organic acids. The linear regression analysis between CsSWI3 gene expression and content of sugar and organic acids were conducted, the results showed that the expression levels of CsSWI3A, CsSWI3B and CsSWI3C were positively correlated with glucose, fructose and sucrose contents, in which the expression level of CsSWI3B showed strongly positive correlation with fructose content (R2 = 0.7163, p < 0.01), sucrose content (R2 = 0.6849, p < 0.01) and glucose content (R2 = 0.6759, p < 0.01), while a lower positive correlation was observed between the expression level of CsSWI3A and fructose content (R2 = 0.509, p < 0.01), sucrose content (R2 = 0.5075, p < 0.01) and glucose content (R2 = 0.4844, p < 0.05); the expression level of CsSWI3C and fructose content (R2 = 0.5530, p < 0.01), glucose content (R2 = 0.5393, p < 0.05) and sucrose content (R2 = 0.3365, p < 0.05) (Fig. 10). However, there was no correlation between organic acid content and the expression levels of CsSWI3s, as well as sugar content and CsSWI3D expression level.

Figure 10.
Correlation analysis between CsSWI3s gene expression and content of sugar and organic acids based on the linear regression method.
-
SWI3 subunit is one of the core accessory subunits of SWI/SNF complex, which is responsible for the normal development of plants under various growth conditions upon altering the interaction between DNA and histone octamers and thus regulating the accessibility of TFs or other DNA-binding proteins to DNA[23,24]. In Arabidopsis, AtSWI3s could play regulatory roles by participating in the assembly of other SWI/SNF complexes, affecting the nutritional and reproductive stages of embryogenesis and plant development, showing remarkable functional diversity[5]. However, the study on chromosome remodeling in fruit trees, especially in citrus, is still in its preliminary stage. In this research, we performed a comprehensive analysis on the identification and characterization of SWI3 subunits in Citrus sinensis.
Four CsSWI3 subunits (CsSWI3A, CsSWI3B, CsSWI3C and CsSWI3D) with three conserved domains (SWIRM, SWIRM-associated region 1 and SANT domain) have been identified based on genome database in C. sinensis. Through the phylogenetic analysis of SWI3 proteins in four species, CsSWI3 had a similar figure to those in Arabidopsis (4 AtSWI3s), rice (6 OsSWI3s) and tomato (4 SlSWI3s). All these members were divided into four clusters, SWI3A to SWI3D. Upon the gene structure analysis, we found that CsSWI3s had a similar structure to AtSWI3s. Analysis of conserved motifs and domains exhibited that the members in the same cluster had a strong similarity, which indicated that SWI3 genes seem to be functionally analogous in the same cluster. Only the members in cluster SWI3D contain a Zinc finger-ZZ type domain, indicating that this subfamily may have particular functions. It was found in Arabidopsis that plants carrying the atswi3d mutation showed severe dwarfing, alterations in the number and development of floral organs, and complete male and female sterility[6]. Consistent with the roles as the subunit of chromatin remodeling complexes, all of the CsSWI3 members were localized in the cell nucleus, which was similar to AtSWI3s in Arabidopsis[4]. Promoter cis-regulatory elements (CREs) containing binding sites for TFs or other regulatory molecules can control plant development and physiology process by regulating gene expression[25]. Cis-regulatory elements identified in 2 kb upstream promoters of CsSWI3s shown in Fig. 6 shed light on their significant stress resistance function, participation in growth and development (except CsSWI3D) in citrus coupled with the response to phytohormone.
In order to explore the temporal and spatial specific expression profile of CsSWI3s in citrus, we analyzed the expression pattern of CsSWI3s in different development periods of fruits and in different tissues (stem, leaf and flowers) of 'Bingtang' sweet orange through qRT-PCR experiments. Gene expression of the CsSWI3s showed an upward trend during fruit development and obvious tissue specificity. The highest expression level in leaves was consistent with AtSWI3B in Arabidopsis, which presumed the potential function of leaf development in CsSWI3B and needed further functional verification[26]. In addition, to study the potential roles of the CsSWI3s in sugar/acid metabolism, the contents of sugar and organic acids, which were the vital contributors to the palatability of citrus fruits, in different developmental stages of 'Bingtang' sweet orange fruits were determined, and their correlation with CsSWI3s gene expression was ulteriorly analyzed upon the linear regression method. As for organic acids, there was no significant correlation between their contents and CsSWI3s expression levels. Besides, intriguingly, CsSWI3A, CsSWI3B and CsSWI3C were all found to be positively correlated with the content of the three sugars (fructose, glucose and sucrose), and CsSWI3B showed the most significant positive correlation, indicating that it may be involved in the regulation of sugar accumulation in citrus fruits, despite further functional verification needed to be performed.
-
In the present study, a total of four CsSWI3 family genes belonging to four subfamilies have been identified based on C. sinensis genome database. Each of CsSWI3 protein contains the same three conserved domains as Arabidopsis, furthermore, subcellular localization in the nucleus is consistent with their function as the core accessory subunits of SWI/SNF chromatin remodeling complex. Meanwhile, CsSWI3s were found widely involved in plant response to various stress and hormones, and regulation of growth and development through the promoter cis-regulatory element analysis. This fact apart, CsSWI3s were expressed ubiquitously in various citrus tissues and differently during the different development stages in fruit. In addition, it showed positive correlation between the expression level of CsSWI3A, CsSWI3B and CsSWI3C and sugar contents, especially for CsSWI3B. Our results may provide comprehensive information for the CsSWI3 gene family in citrus and establish a foundation for the functional identification of CsSWI3 genes in the regulation of sugar accumulation during citrus fruit development, and provides a basis for the further study of chromatin remodeling complex in citrus and other fruit trees.
-
The authors confirm contribution to the paper as follows: study conception and design: Li S, Li Y; data collection: Li Y, Lu D; technical assistance: Wang D; analysis and interpretation of results: Li Y, Xu M; draft manuscript preparation: Li Y, Liu S, Li S. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed in this study are included in this published article and supplementary information files.
This work was supported by grants from the National Natural Science Foundation of China (31801591).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primers used in this paper.
- Supplemental Table S2 The biophysical properties of CsSWI3 proteins.
- Supplemental Table S3 The cis-regulatory elements on CsSWI3' spromoter.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li Y, Xu M, Lu D, Wang D, Liu S, et al. 2024. SWI3 subunits of SWI/SNF complexes in Sweet Orange (Citrus sinensis): genome-wide identification and expression analysis of CsSWI3 family genes. Fruit Research 4: e002 doi: 10.48130/frures-0023-0038 

SWI3 subunits of SWI/SNF complexes in Sweet Orange (Citrus sinensis): genome-wide identification and expression analysis of CsSWI3 family genes
- Received: 20 September 2023
- Revised: 10 October 2023
- Accepted: 23 October 2023
- Published online: 02 January 2024
Abstract: SWI3 proteins as the core accessory subunits of SWI/SNF chromatin remodeling complexes (CRCs) could jointly take part in the genome epigenetic regulation upon disrupting the interaction between DNA and histones, ulteriorly regulating the accessibility of DNA-binding proteins or TFs to DNA. Research on chromatin remodeling complexes in plants lags behind yeast and animals, however, the last decade has witnessed an intensive effort to enhance our understanding of identification, characterization and molecular mechanisms of CRCs in Arabidopsis which provided the information for further studies in other plant species. So far, genome-wide identification of SWI3 family in citrus has not been reported. Here, four CsSWI3 genes based on Citrus sinensis genome were identified and clustered into four subfamilies. According to conserved domains and motifs analysis, we found that each CsSWI3 protein contained three conserved domains and the members in the same subfamily showed strong similarity with those in Arabidopsis. All of the CsSWI3 members were localized in the cell nucleus, which was consistent with the role as the subunit of CRCs. Analysis of promoter cis-regulatory elements indicated that CsSWI3 genes may be involved in stress response, phytohormone response and growth and development of citrus. Meanwhile, they were expressed extensively in citrus tissues and disparate development stages in fruit. We found that the expression level of CsSWI3A, CsSWI3B and CsSWI3C are positively correlated with sugar content during fruit development, especially for CsSWI3B. This study provides comprehensive information for the CsSWI3 gene family and sets a basis for its function identification in citrus.
-
Key words:
- Fruit quality /
- Gene family /
- Epigenetic regulation