









-
The loquat, scientifically known as Eriobotrya japonica Lindl., is a perennial fruit tree that originates from China. It is classified under the family Rosaceae, specifically the subfamily Maloideae[1,2]. The fruit serves as an excellent reservoir of essential nutrients such as vitamin A, vitamin B6, potassium, magnesium, and dietary fibre[3]. It is characterised by its vibrant orange colour and possesses a delightful, subtle sweetness[4]. The loquat fruit, with its delicate and luscious flesh, along with its thin skin, has a limited shelf life and cannot be stored for a prolonged duration[5]. In addition to its utilisation as a fruit, it serves as an exceptional honey source. The flowers of this plant, especially white ones, possess a strong attraction for honey bees[6]. The cultivation of loquat is primarily concentrated in the south-central region of China, as well as in Japan, Korea, India, Pakistan, and Korea[7]. China, being the foremost global producer and exporter of loquat, cultivates this fruit on an extensive area spanning over 130 thousand hectares. In China, the annual production of loquats amounts to 650 thousand tonnes[8]. The presence of reduced sugar levels and elevated fruit acidity has had a notable detrimental impact on the overall fruit quality and market value of loquat crops in commercial cultivation[9].
Fruits undergo a fascinating process of biochemical and nutritional development, which ultimately contributes to their distinct and desirable quality[10−12]. The development of fruits is frequently accompanied by the accumulation and breakdown of sugars and organic acids, which plays a crucial role in the establishment of fruit flavour. This particular process promotes the synthesis of sugar and organic acid metabolites[12,13]. The ultimate sugar-acid concentration in mature fruits is influenced by the interplay of sugar-acid synthesis, degradation, and vacuolar retention[14,15]. Soluble sugars, such as glucose, sucrose, and fructose, are highly prevalent in certain fruits, while sucrose is commonly found in various fruits[15−17]. Fructose is the predominant soluble sugar present in loquats, as supported by scientific literature[18,19].
Methyl jasmonate (MeJA) is a natural compound that plays an important role in growth, development and response to various stresses. Foliar applications of MeJA have been shown to regulate antioxidant systems against oxidative stress induced by biotic and abiotic stimuli[20] and change expression of genes responsible for fruit ripening, pollen production, leaf buds, shoots and root hairs growth[21]. Due to its volatile nature, it has the ability to diffuse in biological membranes and negatively regulate the production of endogenous jasmonic acid, and stabilizes the membrane lipids. Preservation of membrane lipids leads to blocking the effects of ethylene, preventing water loss, reducing respiration and many other processes related to senescence, like softening[22]. MeJA also plays a vital role in improving plant productivity and quality of fruits. Strawberry plants treated with MeJA exhibited significant decrease in vegetative growth traits and increase in total soluble solids in fruits[23].
Here, in this study, we investigated the effect of different concentrations of MeJA on sugar metabolism-related genes in loquat fruit. Initially, loquat plants were foliar sprayed with 0 (control), 5, 10, and 15 mM MeJA, and changes in fruit weight, size, and sugar profile were recorded. The samples from best performing treatment (10 mM MeJA) and control (0 mM MeJA) were used to measure the expressions of sugar metabolism-related genes using RT-qPCR technique.
-
The loquat trees (Cv. Zaozhong 6) employed in this study were selected from an orchard situated in the subtropical region of Fujian province, specifically in Fuqing (latitude 25°41'38.9"N, longitude 119°10'56.7"E). These trees varied in height, ranging from 4 to 5.5 m, with canopy diameters measuring between 4 and 5 m. The spacing between trees in the orchard was approximately 6 m × 6 m.
To ensure the trees' well-being and productivity, a systematic approach was applied, involving pruning and thinning methods. Additionally, NPK fertilizer (15:15:15) was consistently applied at a rate of 5 kg per tree per season over the past three growing seasons. The trees were assigned to different treatments using a randomized complete block design (RCBD), with four replications (blocks) for each treatment. Each individual tree served as a replicate for each treatment.
Conventional agricultural practices, including drip irrigation, careful management of mineral nutrition, effective weed control, and the prevention and management of insect/pest infestations and diseases, were employed to ensure the overall health and optimal growth of the loquat trees.
In this study, we have undertaken an investigation to assess the impact of varying concentrations of MeJA (≥ 95%, CAS: 39924-52-2, Solarbio Life Sciences, Beijing, China) on the expression of genes related to sugar metabolism in loquat fruit. In the initial phase, the loquat plants underwent foliar application of different concentrations of MeJA including 0 (control), 5, 10, and 15 mM. Subsequently, we observed the changes in the fruit weight, size, and sugar composition (Supplemental Table S1). The samples obtained from the treatment that exhibited the highest performance (10 mM MeJA) and the control group (0 mM MeJA) were utilized to assess the expression levels of genes associated with sugar metabolism through the use of the RT-qPCR technique. The foliar application was carried out in triplicate after the full bloom stage with two week intervals. Loquat trees were foliar sprayed with MeJA early in the morning using 5 L electronic sprayer operated at a constant speed. Loquat fruits were sampled from the sun-exposed tree canopy[1,2], at about 1.5–2.5 m height, at five different developmental stages of loquat i.e., fruitlet, immature green, mature green, breaking, and full ripe at 65, 80, 95 and 110 d after full bloom, respectively.
Fruit weight and size
-
The fresh weight, length (measured from the highest vertical point), and diameter (measured from the widest horizontal point) of the fruit were determined by averaging measurements from five separate batches, each consisting of 10 loquats from the same treatment group. Fruit weight was assessed using a digital weighing balance (MJ-W176P, Panasonic, Japan), while length and diameter were measured using digital Vernier calipers (DR-MV0100NG, Ningbo Dongrun Imp. & Exp. Co., Ltd., China). The fruit shape index was calculated for each fruit by determining the length-to-diameter ratio[24].
Total soluble sugars, titratable acids, and sugar-acid ratio
-
Total titratable acids were determined with NaOH-based titrimetric method[25] and expressed as percent citric acid. The soluble sugars of fruit tissues were extracted with boiling ethanol/water and determined by the method of Sharma et al.[26], using an ultraviolet spectrophotometer (UV-5100B, Shanghai Metash Instruments Co., Ltd, China). The concentration of soluble sugars was expressed as milligram per gram (mg·g−1). The sugar-acid ratio was calculated by dividing soluble sugars to total titratable acids within the same sample.
Fructose, glucose, and sucrose determination through HPLC
-
The fruit samples, with their pulp stored at a temperature of −80 °C, underwent initial processing by means of crushing and pulverization using liquid nitrogen. Subsequently, a 2 g sample of finely ground powder was blended with 10 mL of 95% methanol, incorporating a suitable quantity of polyvinylpyrrolidone. The mixture was subjected to ultrasonication at a temperature of 40 °C for a duration of 30 min, after which it was centrifuged at a speed of 1,000 rpm for a period of 10 min. The supernatant fluid obtained was carefully transferred into a distilling flask with a capacity of 20 mL. The aforementioned procedure was replicated utilizing the agricultural residues in conjunction with 8 mL of ultrapure water. Following the procedure, the transparent liquid was effectively passed through a 0.22 µm syringe filter, specifically from ANPEL (China).
HPLC analysis was performed utilizing the Waters 2695 autosampler system. For the separation of soluble sugars, a column called Ellistat Supersil NH2 (4.6 mm × 250 mm, 5 µm particle size) manufactured by Waters Inc, (Zellik, Belgium) was employed. The column was operated at a temperature of 40 °C. The composition of the mobile phase comprised an 82% acetonitrile and 18% ultrapure water solution. The flow rate was adjusted to 1.2 mL·min−1, while the injection volume was determined to be 20 µL. Ultimately, the determination of the concentration of each distinct soluble sugar was derived by utilizing the calibration curve specific to the corresponding standard. The measurements for soluble sugars were conducted in triplicate, and the outcomes were expressed in milligrams per gram (mg·g−1).
RNA extraction and real-time quantitative PCR
-
The extraction of total RNA from loquat fruit pulp was carried out using a Total RNA kit provided by TianGen Biotech (Beijing, China). The assessment of RNA quantity and quality was conducted utilizing the NanoDrop N-1000 spectrophotometer, developed by NanoDrop technologies (Wilmington, DE, USA). The Prime Script RT Reagent Kit with a gDNA Eraser, (TaKaRa, Dalian, China), was employed for the synthesis of first-strand cDNA. This process utilized 1 µg of total RNA as the starting material. The real-time qPCR analysis was conducted utilizing a high-performance real-time PCR instrument, specifically the LightCycler® 96 (Roche Applied Science, Penzberg, Germany). The qRT-PCR primers were meticulously designed using the Primer-blast tool, and their details can be found in Supplemental Table S2.
The reaction mixture comprised of 10 μL of 2×RealStar Green Fast Mixture (GenStar, Bejing, China), 1 µL of cDNA, 0.25 µM of each primer, and water was incorporated to achieve a total volume of 20 µL. The actin protein, specifically the EVM0004523.1 variant, was employed as an internal control in this study[27−29]. To determine the relative gene expression, the 2−ΔCᴛ method was utilized[30]. The validation of the 2−ΔCᴛ method was conducted through an analysis of ΔCᴛ variation at various template concentrations[31−33]. Each sample was subjected to analysis using three independent biological replicates.
Statistical analysis
-
The experimental design employed was a randomized complete block design (RCBD) consisting of four blocks. Within each block, there was one replication of each treatment. The data collected were subjected to analysis of variance (ANOVA) using the statistical software 'Statistix 8.1' (
www.statistix.com ). To compare the means of replicated data from each treatment, Student's t-test was applied. For the purpose of Principal Component Analysis (PCA) involving treatments and tested variables, Pearson's method (n) was utilized. This analysis was conducted using 'XLSTAT ver. 2019' (www.xlstat.com/en ). The clustering of variables associated with treatments was determined based on their highest squared cosine values corresponding to PCA factors, namely F1, F2, or F3. -
Methyl jasmonate significantly influenced the fruit weight and size of loquats fruits during their growth and development (Fig. 1). At fruitlet stage, the pericarp of loquat was green, the fruit was small and oval, with obvious villus on the surface, and there was no significant difference among MeJA treatments. At immature green stage, the fruit began to develop, the pericarp was dark green, and the epidermal villi were not obvious. At mature green stage, the fruit expanded rapidly, the skin surface of loquat changed from dark green to light green, the skin surface villus further degraded, the skin began to thin, and there was no significant difference in fruit weight and fruit shape index. At breaking stage, the fruit entered the color transformation stage, the fruit size and fruit shape index were basically consistent with the maturity stage (as shown in Fig. 1a−d), and the pericarp changed from light green to yellow-green (Fig. 1e, Supplemental Fig. S1). At the full ripe stage, the fruit weight of 10 mM MeJA-treated fruits was significantly 26.94% increased (p ≤ 0.05) compared to those of the control (0 mM MeJA). Fruit diameter was significantly (p ≤ 0.01) 11.40% increased under the influence of 10 mM MeJA, whereas length remained unchanged, caused 9.73% decline in fruit shape index at full ripe stage.
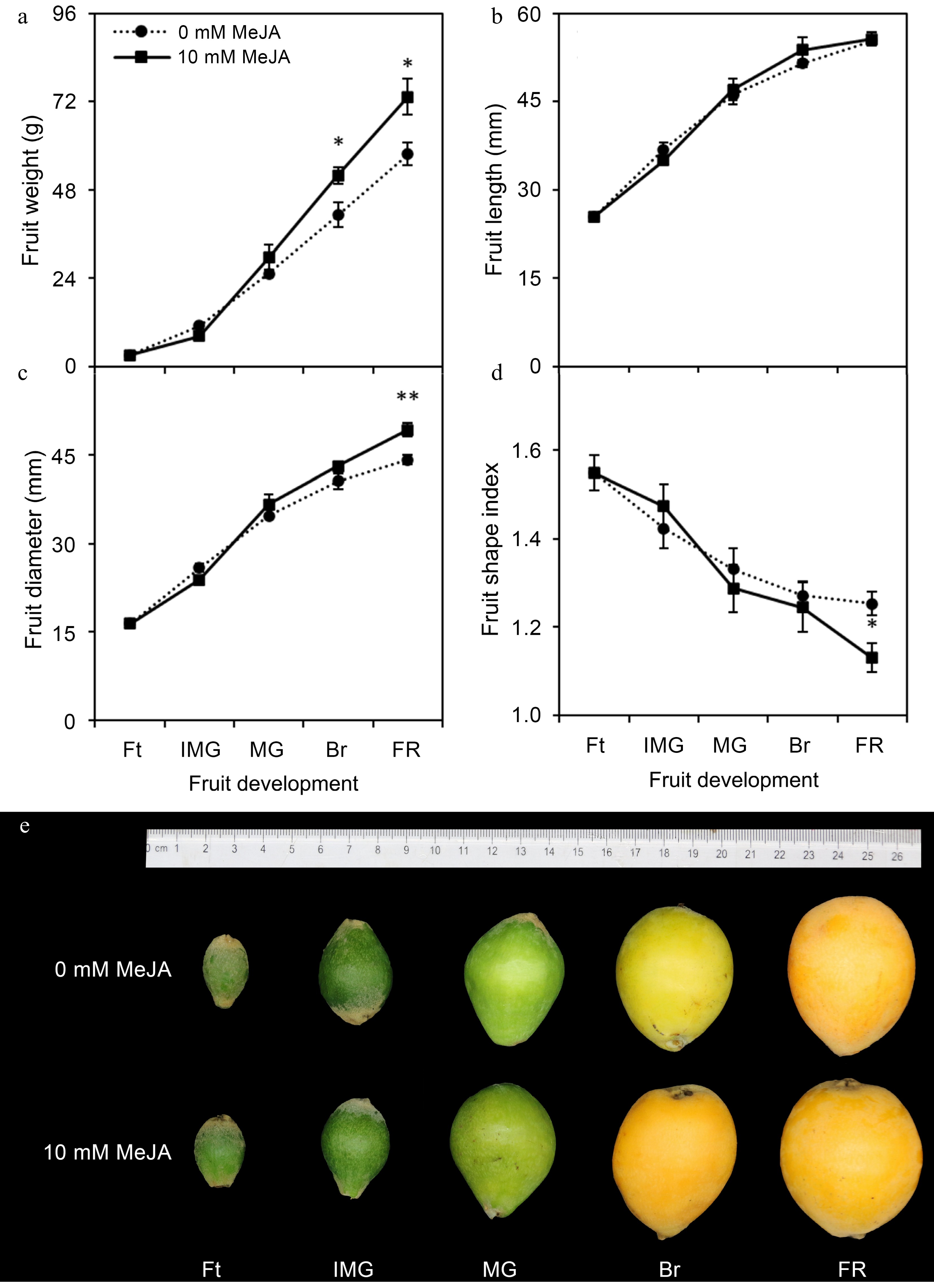
Figure 1.
Influence of foliar applied methyl jasmonate on fruit weight and size of loquat at different developmental stages. Treatment means were compared using Student's t-test, where *, **, and *** denote significance level (α) of 0.05, 0.01, and 0.001, respectively. Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
Sugar and acid profile
-
The content of soluble sugars was gradually increased with fruit development. However, MeJA treatment significantly (p ≤ 0.01) increased soluble sugars (up to 22.50%) in loquat at full ripe stage as compared with control (0 mM MeJA) (Fig. 2a). In case of titratable acids (TA), fruits treated with MeJA showed significant (p ≤ 0.01) decline at mature green and full ripe stage. However, maximum TA (1.79%) was recorded in the fruits obtained from 0 mM MeJA-treated plants, which was gradually decreased with fruit development (Fig. 2b). Figure 2c indicates a significant difference of sugar-acid ratio between MeJA treatments after mature green stage. At full ripe stage, 10 mM MeJA-treated fruits showed sugar-acid ratio of 110.78 which was 2.4-fold higher than that of control.
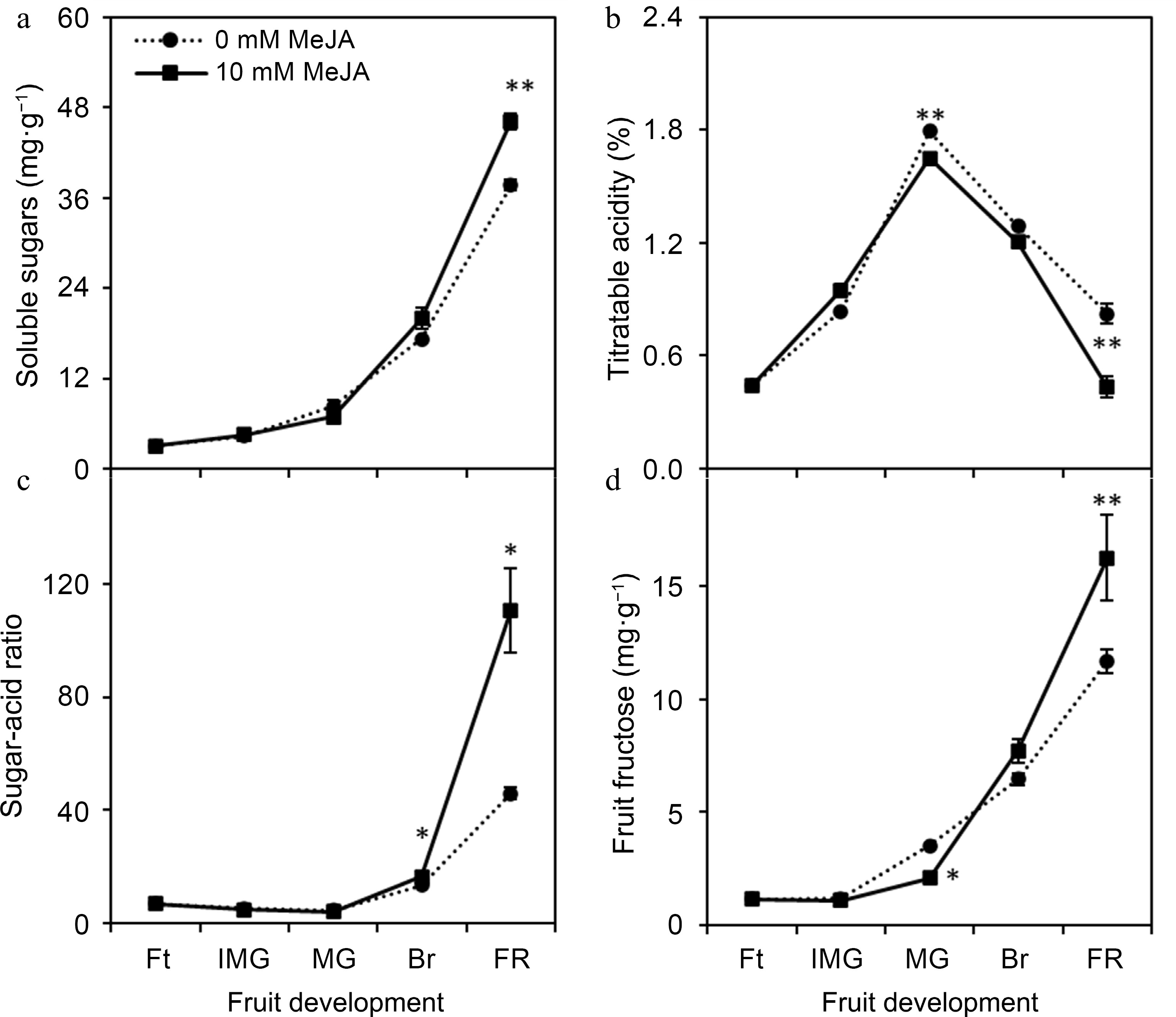
Figure 2.
Effect of foliar applied methyl jasmonate on (a) fruit soluble sugars, (b) titratable acidity, (c) sugar-acid ratio and (d) fruit fructose content of loquat at different developmental stages. Sugar determination involved the use of fresh weight in the assessment process. Treatment means were compared using Student's t-test, where *, **, and *** denote significance level (α) of 0.05, 0.01, and 0.001, respectively. Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
To better understand the sugar dynamics of loquat, three kinds of soluble sugars i.e., fructose, sucrose, and glucose were measured using HPLC. However, fructose was the only sugar detected in abundance at all developmental stages of loquat. Fruit fructose gradually increased with fruit development and was found significantly 38.96% increased in 10 mM MeJA-treated fruit as compared with control at full ripe stage (Fig. 2d).
Key genes regulating soluble sugars metabolism
-
Among seven fructokinase genes[19], six genes were expressed in loquat fruits at different developmental stages (Fig. 3). The EjFK1 and EjFK2 were significantly (p ≤ 0.01) up-regulated under the influence of applied MeJA at immature green stage. At mature green stage, EjFK4, EjFK6 and EjFK7 showed down-regulation in fruits obtained from 10 mM MeJA-treated plants comparing with those of 0 mM MeJA-treated plants. EjFK5 was significantly influenced by MeJA treatment at immature green stage. Interestingly, under the influence of 10 mM MeJA, EjFK7 showed 1.54-fold downregulation at mature green stage, was found up-regulated by 1.42-fold at full ripe stage.
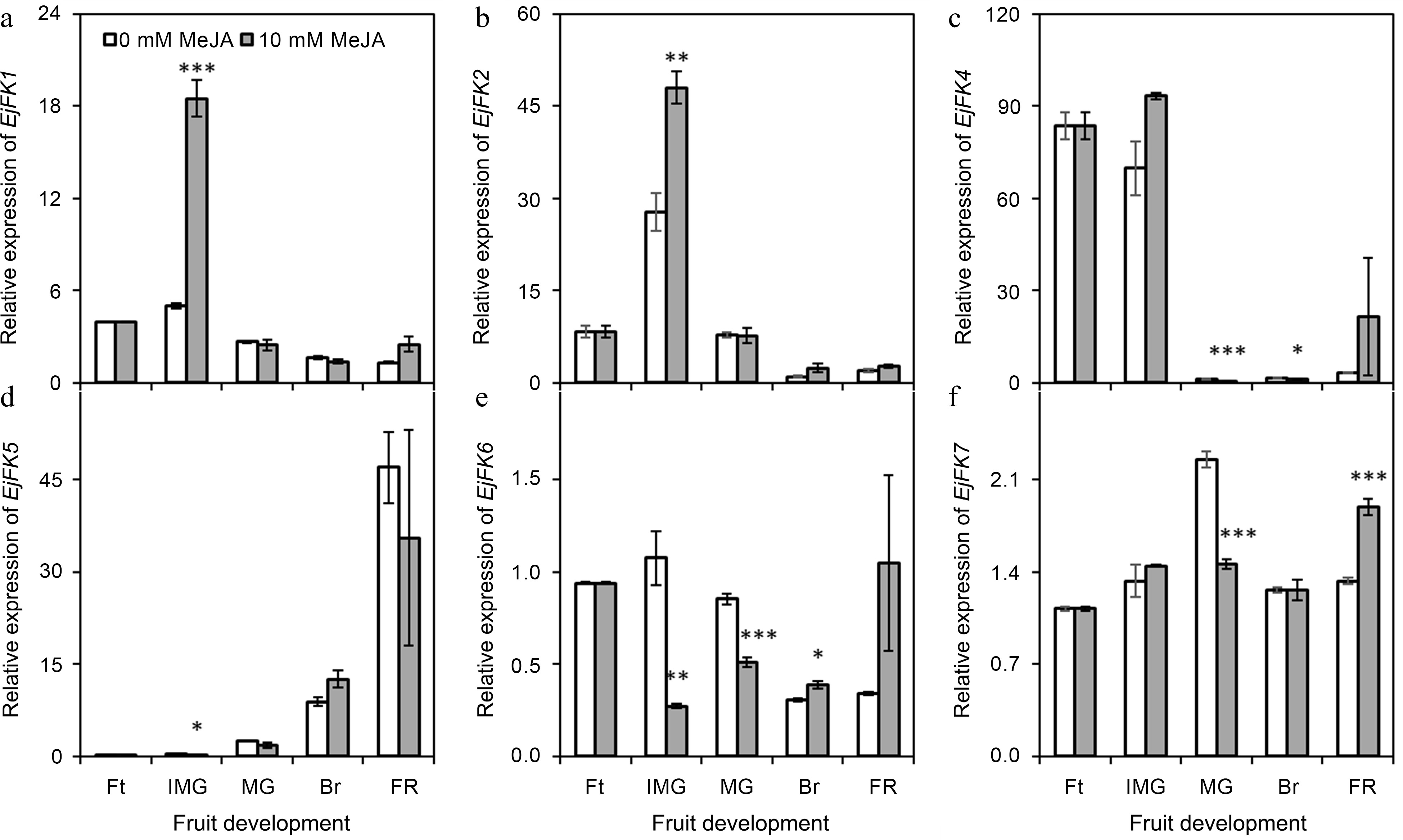
Figure 3.
Effect of foliar applied methyl jasmonate on expression patterns of EjFK genes in loquat fruits at different developmental stages. Treatment means were compared using Student's t-test, where *, **, and *** denote significance level (α) of 0.05, 0.01, and 0.001, respectively. Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
The expression of EjHK1 was found to be significantly (p ≤ 0.01) reduced upon the application of MeJA during the immature green and mature green stages. However, it was observed to be up-regulated (p ≤ 0.001) during the breaking and full ripe stages. Furthermore, the EjHK1 was maximally expressed at mature green stage in loquats obtained from 0 mM MeJA-treated plants (Fig. 4a). In a similar pattern, EjHK2 was also found to be significantly (p ≤ 0.01) downregulated at immature and mature green stages (Fig. 4b). Likewise, the expression of EjHK3 was also decreased after the application of 0 mM MeJA at immature green, mature green and breaking stage. Interestingly, its expression was found comparably increased at full ripe stage under the same treatment (Fig. 4c).
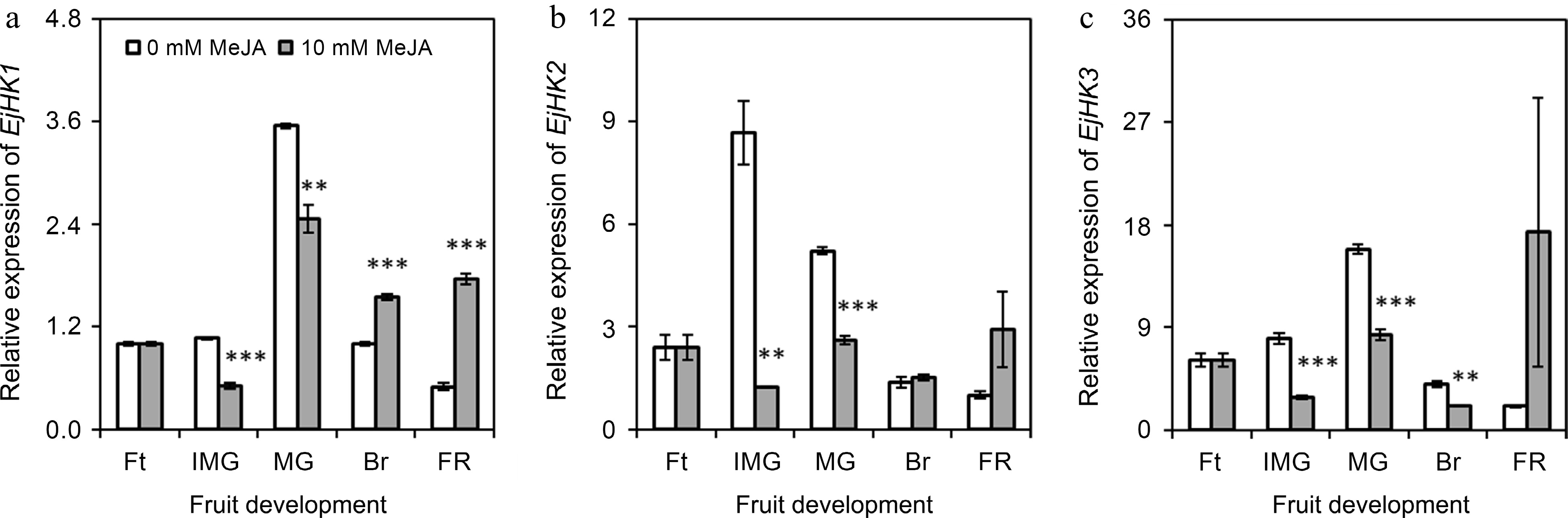
Figure 4.
Effect of foliar applied methyl jasmonate on expression patterns of EjHK genes in loquat fruits at different developmental stages. Treatment means were compared using Student's t-test, where *, **, and *** denote significance level (α) of 0.05, 0.01, and 0.001, respectively. Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
The expression of EjSPP1 was found to be significantly (p ≤ 0.05) reduced upon the application of MeJA during the mature green stage of loquat (Fig. 5a). In the control group, the expression of EjSPP2 was gradually increased with fruit development till breaking stage, then decreased at full ripe stage. Its expression was found to be significantly (p ≤ 0.01) reduced upon the application of MeJA during the immature green and mature green stages. However, it was observed to be up-regulated (p ≤ 0.001) in loquat fruits during the breaking stage (Fig. 5b). Interestingly, the expression of EjSPP3 suddenly increased by 4.23-fold in fruits obtained from 10 mM MeJA-treated plants as compared with control at immature green stage. Post the immature green stage, its expression gradually decreased in MeJA-treated samples. However, significant differences between MeJA treatments were observed at the immature green (p ≤ 0.001), mature green (p ≤ 0.001), and full ripe stages (p ≤ 0.01) (Fig. 5c).
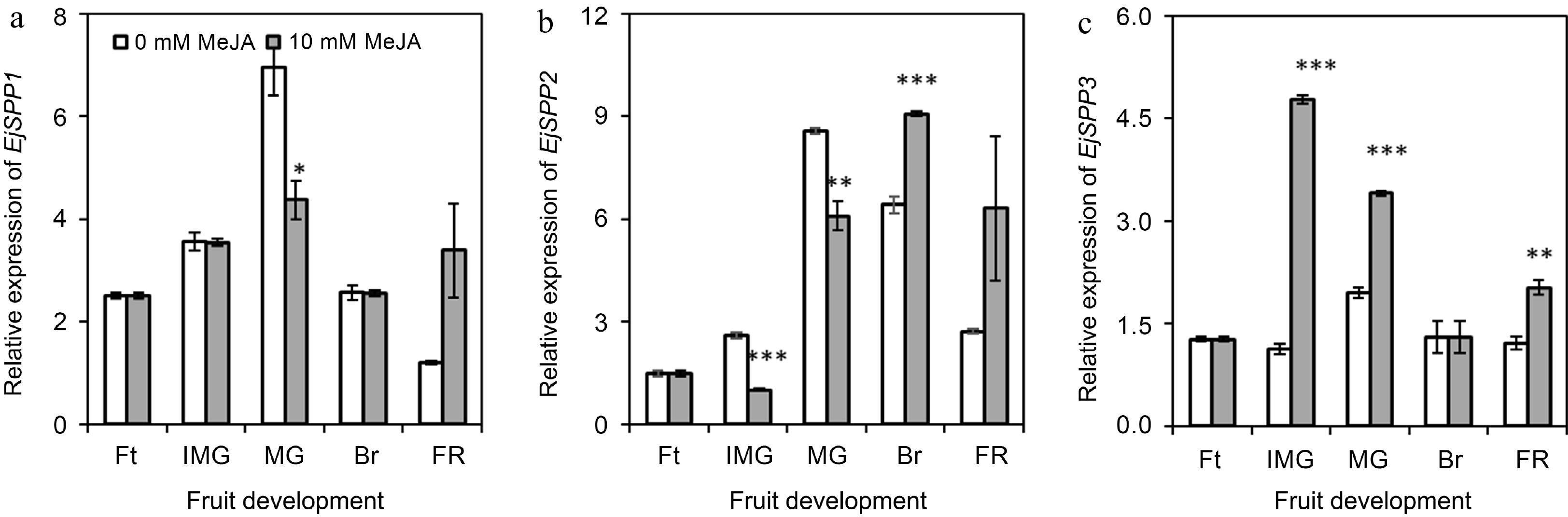
Figure 5.
Effect of foliar applied methyl jasmonate on expression patterns of EjSPP genes in loquat fruits at different developmental stages. Treatment means were compared using Student's t-test, where *, **, and *** denote significance level (α) of 0.05, 0.01, and 0.001, respectively. Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
The expression of EjSPS1 increased dramatically with fruit development, regardless of MeJA applied. Moreover, it was found to be significantly (p ≤ 0.05) reduced upon the application of MeJA during the mature green stage and breaking stage of loquat comparing with control (Fig. 6a). The expression of EjSPS2 increased at immature green stage, then gradually decreased with fruit development. Its expression remained unchanged under the influence of MeJA treatment except at immature green stage, where it was found to be significantly (p ≤ 0.001) increased (Fig. 6b). Likewise, the expression of EjSPS3 gradually increased with fruit development. Its expression remained unchanged under the influence of MeJA treatment except at immature green stage, where it was found to be significantly (p ≤ 0.001) increased (Fig. 6c). The genetic expression of EjSPS4 was also up-regulated after MeJA application at immature green (p ≤ 0.001) and breaking stage (p ≤ 0.01) (Fig. 6d).
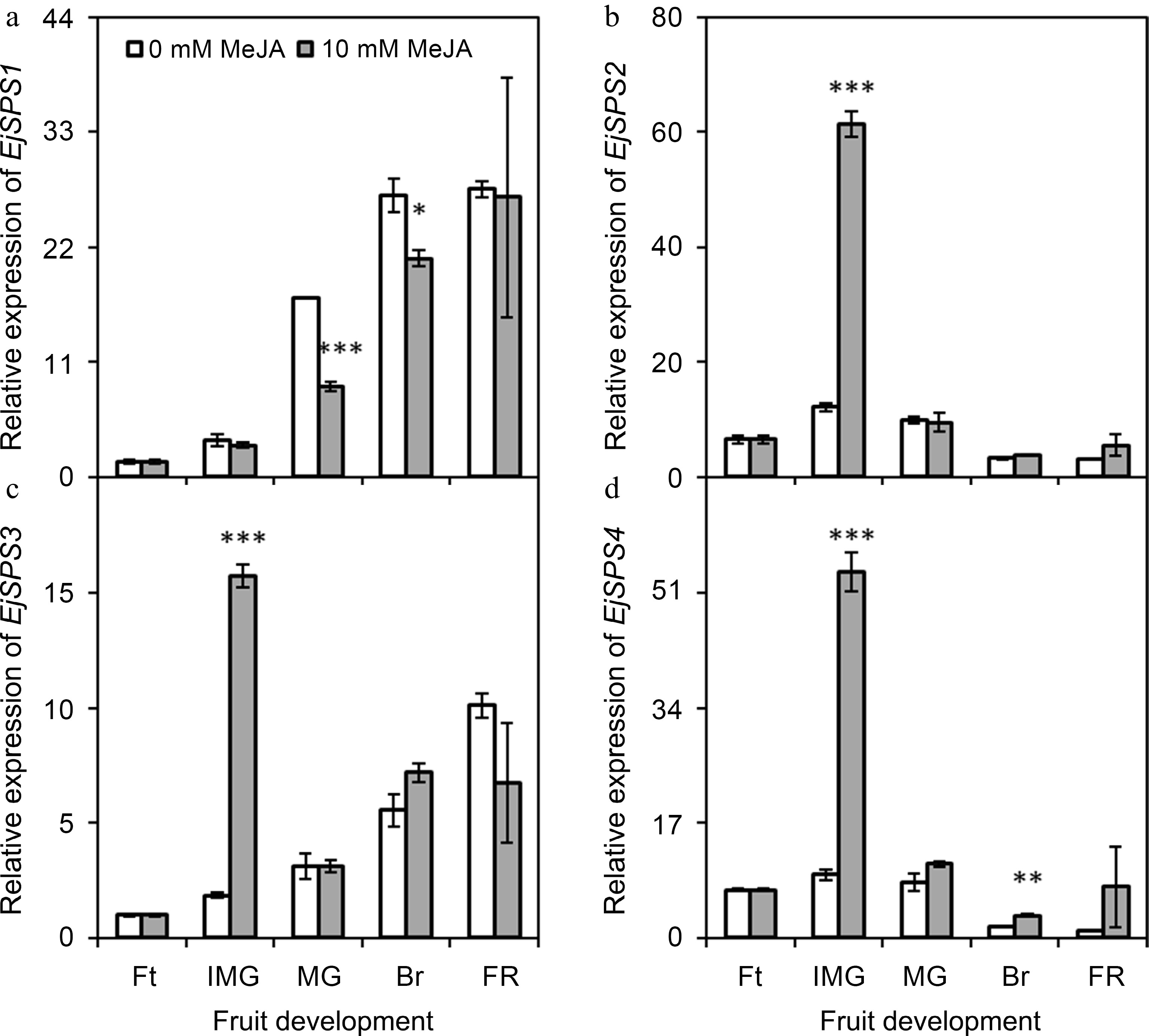
Figure 6.
Effect of foliar applied methyl jasmonate on expression patterns of EjSPS genes in loquat fruits at different developmental stages. Treatment means were compared using Student's t-test, where *, **, and *** denote significance level (α) of 0.05, 0.01, and 0.001, respectively. Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
The expression of EjSS1 increased at immature green stage, then suddenly decreased at mature green stage. It was found to be significantly (p ≤ 0.001) reduced upon the application of MeJA during the immature green, mature green and breaking stage of loquat comparing with control. The maximal reduction (about 47 times) was observed at immature green stage (Fig. 7a). The expressions of EjSS3, EjSS5 and EjSS6 were maximally observed at immature green stage (Fig. 7b−d). Moreover, EjSS3 and EjSS6 exhibited downregulation and EjSS5 showed significant up-regulation (p ≤ 0.05) under the influence of MeJA treatment at immature green stage.
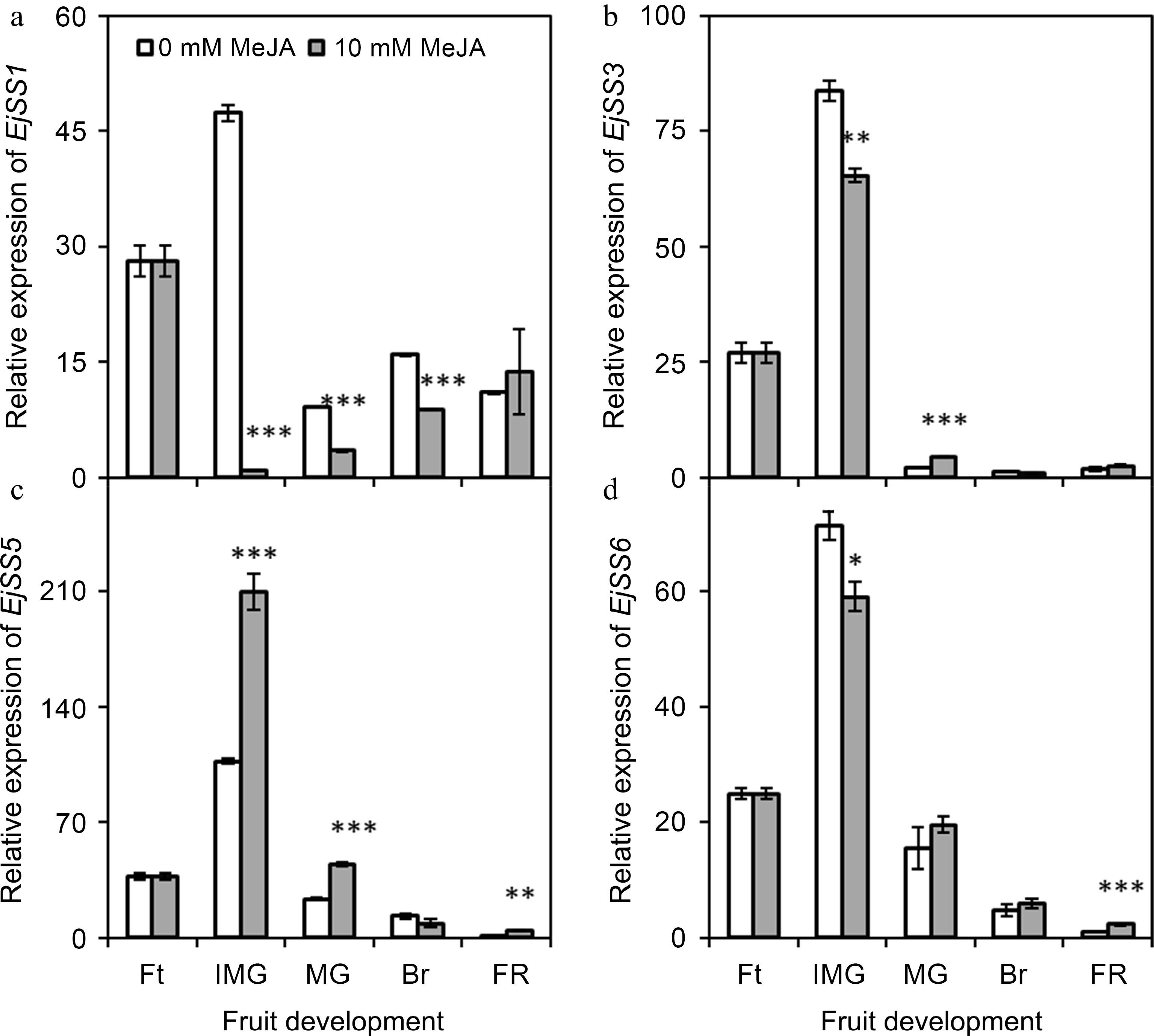
Figure 7.
Effect of foliar applied methyl jasmonate on expression patterns of EjSS genes in loquat fruits at different developmental stages. Treatment means were compared using Student's t-test, where *, **, and *** denote significance level (α) of 0.05, 0.01, and 0.001, respectively. Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
Principal Component Analysis (PCA)
-
PCA was carried out to demonstrate the relationship of applied MeJA treatments and studied parameters, including sugar profile of loquat and expression of its biosynthesis-related genes (Fig. 8). PCA coordinates were plotted against three PCA factors. PCA factor 1 (F1) having an eigenvalue of 12.62 and variability of 45.072, contained highest squared cosines of fruit weight, length, diameter, soluble sugars, fructose, EjSPP2, EjFK5, and EjSPS1, with MeJA treatments (including control) at breaking and full ripe stage. Another cluster of same factor (F1) was plotted in the opposite-side-quadrant containing fruit shape index, EjFK1, EjFK2, EjFK4, EjSS3, EjSS5, and EjSS6 with fruitlet, immature green stage, indicating their positive association with each other. PCA factor 2 (F2) having eigenvalue of 5.62 and variability of 20.096 showed clustering of some genes i.e., EjSPP3, EjSPS2, EjSPS3, EjSPS4, EjHK2, EjFK6, and EjSS1. Likewise, third factor of PCA (F3) had an eigenvalue of 4.69 and variability of 16.784. This factor showed clustering of titratable acidity, EjSPP1, EjFK7, EjHK1, and EjHK3 with mature green stage, regardless of applied MeJA treatments. Thus, PCA helped to delineate the expression of different variables at different growth stages of loquat.
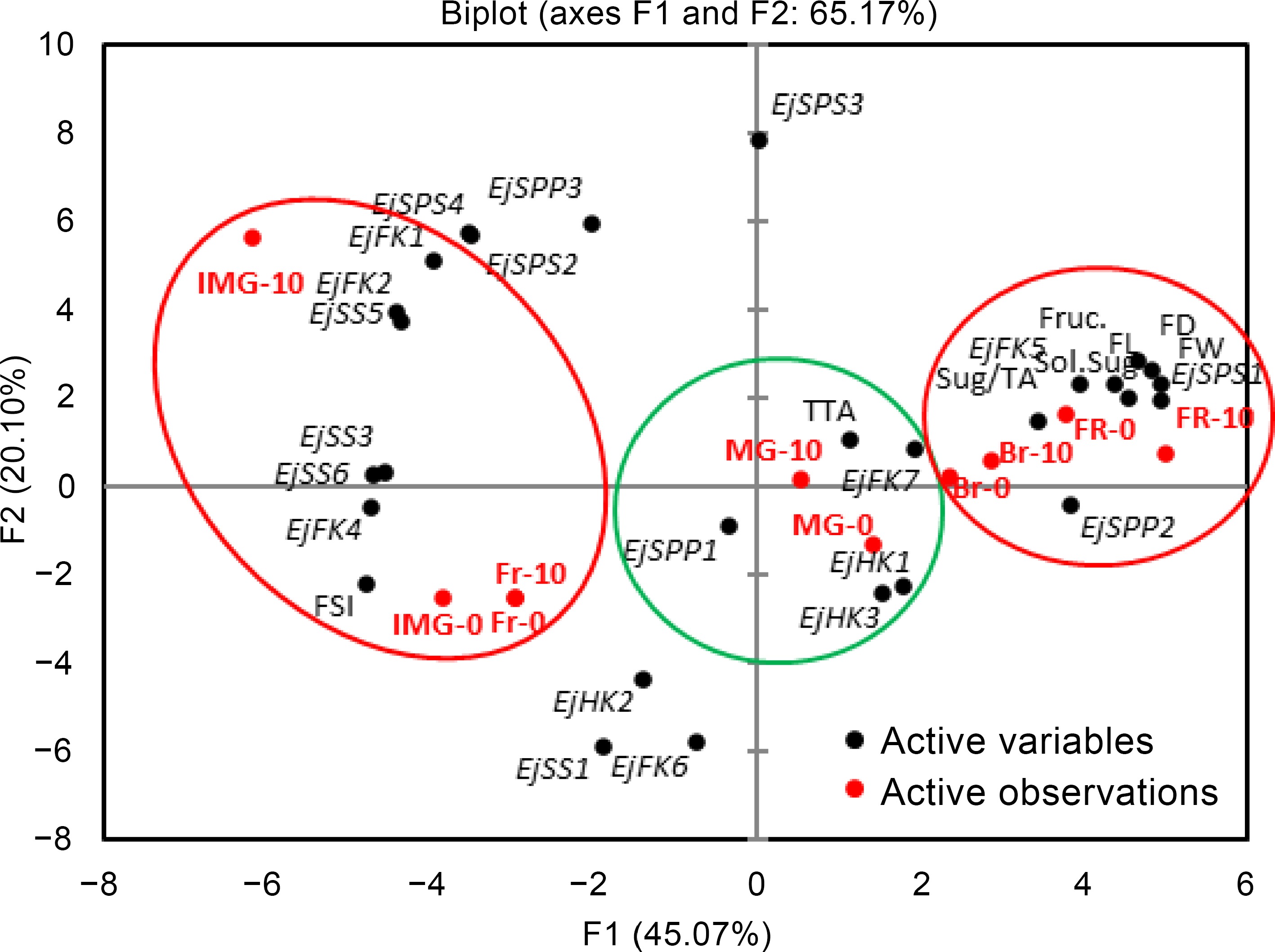
Figure 8.
Principal component analysis among MeJA treatments (treatment variables) and studied parameters (observation variables). Clustering of variables has been done for PCA F1 (red) and F3 (green) according to their highest squared cosines. Fr – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe, FW – fruit weight, FL – fruit length, FD – fruit diameter, FSI – fruit shape index, Sol.Sug – soluble sugars, TTA – total titratable acidity, Sug/TA – sugar-acid ratio, Fruc. – fruit fructose.
-
Soluble sugars and organic acids, being the fundamental constituents of fruit flavour, exert a substantial influence on the overall quality of the fruit[34]. Sucrose, glucose, and fructose are the primary sugars identified in loquat fruits[35,36]. Our study focused on a specific species of loquat 'Zaozhong 6', and our analytical methods did not detect the presence of sucrose at any of the developmental stages. While it is unusual to observe the complete absence of both glucose and sucrose, especially given the importance of these sugars in plant metabolism, we believe this phenomenon could be attributed to the unique biochemical characteristics of the loquat species under investigation. Sucrose absence is expected due to the known absence of this sugar in the wild species of loquat studied[37].
Loquat cultivars exhibit variations in their sugar content. The majority of cultivars exhibit elevated sucrose levels, succeeded by fructose and glucose, whereas it is worth noting that wild species lack sucrose[38]. Based on our observations, the highest fructose concentration (16.25 mg·g−1) was observed in loquat fruits at full ripe stage. The same findings have been reported by Ali et al.[19] in 'Jiefangzhong' loquat. However, being an acidic cultivar, Jiefangzhong's sugar content were maximally recorded as 23 mg·g−1, which is almost half of our studied cultivar i.e., Zaozhong 6. Also, the highest fruit fructose contents and reduced fruit citric acid and malic acid contents were found in raspberry cultivars treated with MeJA[39].
In the present study, soluble sugars continued to accumulate during fruit development process, however, slowly accumulated in the early stage of fruit development (fruitlet – mature green stage). In the later stage of fruit development (breaking – full ripe stage), the accumulation was rapid. At full ripe stage, soluble sugars were recorded comparatively higher in the fruit obtained from 10 mM MeJA-treated plants. One possible explanation could be that when loquat fruit was exposed to 10 mM MeJA treatment, it initiated the maturity stage prematurely, leading to an increase in its sugar content. The results are consistent with the findings of Dong et al.[40], that sprayed methyl jasmonate on 'Huangguogan' citrus.
In several fruits, an increase in the soluble sugars and a concomitant reduction of the titratable acidity are observed during fleshy fruit ripening, which determine final fruit flavor and acceptance of fruit[41−47]. Changes in the sugar-acid ratio by preharvest field MeJA applications have been reported in cultivars of blackberries[48], red raspberry[39], and Chilean strawberry[49]. Here, we found higher values of sugar-acid ratio in 10 mM MeJA-treated fully ripe loquat fruits compared with control. Based on the findings of Zuñiga et al.[50], it is evident that a single application of MeJA is insufficient to effectively modify the sugar-acid ratio. Our research aligns with this notion, as we have observed that a minimum of three consecutive applications of MeJA, starting from the flowering or early fruit developmental (fruitlet) stages, is necessary to enhance this particular parameter in commercial loquat cultivation.
During the metabolic processes of soluble sugars in plant cells, sucrose undergoes degradation, resulting in the formation of UDG-glucose and fructose. This breakdown is facilitated by the enzyme sucrose synthase (SS). Alternatively, sucrose can also be hydrolyzed into glucose and fructose through the action of invertases[51]. Fructokinases (FKs) and hexokinases (HKs) exhibit remarkable substrate specificity and affinity when it comes to phosphorylating free fructose[16,52]. Considering this, it is believed that the process of fructose phosphorylation by FKs and HKs is necessary for the breakdown of sucrose and the overall metabolism of sugars. These processes play a crucial role in the appropriate growth and development of vascular tissue[53,54]. Sucrose phosphate synthase (SPS) and sucrose synthase (SS) are the primary enzymes responsible for the biosynthesis of sucrose. While SS is responsible for the conversion of sucrose into UDP-glucose and fructose[55], SPS facilitates the synthesis of 6-phosphate sucrose through the reaction between UDP-glucose and 6-phosphate fructose. The cytosol and plasma membrane house the majority of the SS proteins, with a smaller number also present in the vacuole, cell wall, or mitochondria[56]. Arabidopsis thaliana and Malus domestica exhibit the presence of four and six SPS genes, respectively[57,58]. The variability in the number of self-incompatibility (SS) genes is observed to be significant across different plant species. In Arabidopsis, carrot (Daucus carota), soybean (Glycine max), and tobacco (Nicotiana tabacum), the number of SS genes is 6, 8, 12, and 14, respectively[59,60]. According to the available literature, it has been documented that the Chinese pear variety under investigation exhibits a total of 30 SS genes[61].
Under the effect of MeJA treatments, the most significant genes involved in sugar metabolism and accumulation were examined for their patterns of expression. Although sucrose concentration was not detectable, the SPS genes showed rather high levels of expression. A negative link between sucrose build-up and SPS activity was shown by the elevated transcript level. This mismatch may be caused by the fact that sucrose, which is the principal soluble sugar in tomato and watermelon[62−64], accumulates more readily than fructose in the loquat[65]. Quantitative analysis of key genes of sugar metabolism in loquat fruits showed that EjFK1,2,4,6, EjHK2-3, EjSPP1-2, EjSPS1-4, and EjSS1,3 were significantly influenced by MeJA treatments during early growth stages. While, EjFK7, EjHK1, EjSPP3, and EjSS5,6 showed significant differences compared with control during late maturity stages. The application of MeJA through leaf spraying appears to have had a notable impact on the expression of crucial genes involved in the sugar metabolism of loquat fruits.
-
The present investigation not only elucidated the changes in sugar composition throughout various growth phases of 'Zaozhong 6' loquat fruit but also provided insights into the impact of MeJA on modulating these dynamics. Based on the findings, it can be inferred that the application of 10 mM MeJA exhibits promising potential as a treatment to optimize sugar composition, thereby leading to an enhancement in the overall fruit quality of loquat. MeJA treatment resulted in notable changes in fruit morphology, with a significant increase in fruit weight (26.94%) and diameter (11.40%) at the full ripe stage. Moreover, MeJA application led to a substantial enhancement (up to 22.50%) in soluble sugars at the full ripe stage, contributing to a higher sugar-acid ratio (110.78). The study also elucidated the intricate regulation of key genes involved in soluble sugars metabolism, such as fructokinase (FK), hexokinase (HK), sucrose-phosphate synthase (SPS), and sucrose synthase (SS), revealing stage-dependent expression patterns influenced by MeJA treatment. Moreover, the findings underscore the importance of the sugar-acid ratio in assessing the quality of fruits and highlight the necessity for accurate implementation of MeJA application strategies (10 mM) in the cultivation of loquat. The insights provided greatly contribute to our comprehension of fruit quality improvement and the intricate molecular mechanisms that regulate sugar metabolism in loquats.
-
The authors confirm contribution to the paper as follows: study conceptualization: Ma S, Ali MM, Chen F; data curation: Ali MM, Gull S; funding acquisition: Hou Y, Chen F; methodology: Ma S, Ali MM, Shi M, Wang X, Li L, Peng C; supervision: Hou Y, Che F; writing – original draft: Ma S, Ali MM, Gull S; writing – review and editing: Shi M, Wang X, Li L, Peng C, Hou Y, Chen F. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This research was funded by 'The Key Project of Leading Counties in Agricultural Science and Technology Modernization, Fujian Province', 'Key Laboratory of Fujian Higher Education Institutions (Putian University)' and 'Ministerial and Provincial Joint Innovation Centre for Safety Production of Cross-Strait Crops, Fujian Agriculture and Forestry University, Fuzhou, China'.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Songfeng Ma, Muhammad Moaaz Ali
- Supplemental Table S1 Effect of foliar applied methyl jasmonate on fruit weight, size, soluble sugars, titratable acidity, sugar-acid ratio and fruit fructose content of loquat at different developmental stages.
- Supplemental Table S2 Primer sequences for qRT-PCR.
- Supplemental Fig. S1 Loquat fruits at different developmental stages under the influence of foliar-applied methyl jasmonate. Abbreviations: Ft – fruitlet, IMG – immature green, MG – mature green, Br – breaking, FR – full ripe.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ma S, Ali MM, Gull S, Shi M, Wang X, et al. 2024. Unveiling the influence of methyl jasmonate on key sugar metabolism genes in developing loquat fruit. Fruit Research 4: e016 doi: 10.48130/frures-0024-0009 

Unveiling the influence of methyl jasmonate on key sugar metabolism genes in developing loquat fruit
- Received: 06 October 2023
- Revised: 14 January 2024
- Accepted: 23 January 2024
- Published online: 24 April 2024
Abstract: The evaluation of fruit quality greatly relies on the presence of soluble sugars. In this study, we investigated the dynamics of soluble sugars and basic fruit quality of 'Zaozhong 6' loquat after the application of methyl jasmonate (MeJA). As loquats approached full ripeness, the study unveiled a notable increase in their soluble sugar content, predominantly attributed to the rise in fructose levels. MeJA application accelerated sugar accumulation, suggesting its potential for expediting fruit maturation and enhancing sugar content. Importantly, MeJA-treated fully ripe loquats exhibited a significantly higher sugar-acid ratio. In addition to compositional analyses, the study delves into the molecular aspects of sugar metabolism. Despite the absence of detectable sucrose and glucose, the expressions of FK, HK, SPP, SPS, and SS genes were notably influenced, emphasizing the complex regulatory mechanisms governing sugar metabolism in loquat fruit. This research provides valuable insights into the temporal changes in sugar composition during loquat fruit development, underscores the role of MeJA in shaping these dynamics, and highlights the significance of the sugar-acid ratio in fruit quality determination.
-
Key words:
- Fruit quality /
- Plant growth regulator /
- Fructokinase /
- Zaozhong /
- Sugar-acid ratio