









-
Strawberry (Fragaria × ananassa) is an important worldwide horticultural crop with many advantages, including: (1) fruit abundant in nutrition, antioxidants, vitamins, fiber and flavonoids; (2) easy cultivation, successive flowering, and high fruit yield; (3) a model plant for studying the ripening of non-climacteric fruit; and (4) its higher economic benefits by winter greenhouse cultivation. In China, the annual planting area of the strawberry industry reaches 173,000 hectares, resulting in a fruit yield of 5 billion kg and generating an income of one hundred billion. China has become the world's largest producer and consumer of strawberries, accounting for more than 50% of the world's total output. The most extensively cultivated is 'Benihoppe', introduced from Japan and constitutes over 50% of total plantings in China[1].
Owing to strawberry fruit without postharvest ripening, it must be harvested after ripening on plants[2], consequently, the harvested fruit are apt to decay[3,4]. Thus in the past years, extensive research has been conducted on the storage techniques and theory underlying harvested strawberry fruit. These include packing materials such as Active Packaging System[5,6]; chemical and physical control, such as hydrogen sulfide[7], high CO2[8], ozone[9], phytosulfokine α (PSKα)[10], triacontanol[4], and UV-C rays[11]; as well as studies on phytohormones and signaling molecules, such as ABA (abscisic acid), IAA (auxin), ethylene (Eth), JA (jasmonic acid), SA (salicylic acid), ROS (reactive oxygen) and NO (nitric oxide)[9,11−17]. However, a comprehensive understanding of the defined physiological and molecular mechanisms for the senescence of harvested strawberry fruit remains ambiguous.
Currently, it is widely accepted that the ripening of strawberry fruit is dominantly controlled by ABA at both biosynthesis and signaling levels, associated to key enzymes such as NCED (9-cis-epoxycarotenoid dioxygenases) and CYP707A (8'-hydroxylases), as well as two core signaling pathways including the 'ABA-FaPYR1-FaPP2Cl-FaSnRK2' and 'ABA-FaABAR-FaRIPK1-FaABI4' pathways[18]. In contrast, post-harvest of strawberry fruit is primarily influenced by hormones, polyphenols, antioxidant enzymes, and softening enzymes[4,11,16,17]. For example, previous reports found that H2O2 mediated anthocyanin degradation and polyphenol formation during strawberry fruit senescence, and redox homeostasis formed a metabolic interface between oxidative stress perception and physiological responses regulated by ROS, remarkably affecting strawberry fruit postharvest senescence[13,14]. The initial burst of ROS accelerates the senescence, while the ROS-mediated feedback control of anti-oxidative activation is involved in the activity of POX and SOD by post-transcriptional regulation[11]. To some extent, ethylene-promoted strawberry fruit senescence is involved in endogenous NO[12], meanwhile ethylene induced ABA biosynthesis rather than ABA catabolism in the receptacle tissue, confirms that postharvest senescence is induced by the interaction of ABA with ethylene[15]. Interestingly, the postharvest ripening of strawberry fruit is delayed by exogenous IAA, while promoted by exogenous ABA; however, the ripening process is not affected by the combination of exogenous IAA + ABA. These findings suggest a vital role of the interaction of IAA with ABA in regulating fruit ripening[9]. Together, ROS, ABA, ethylene and IAA may play important roles in the senescence of strawberry fruit.
At the gene level, it is now reported about 3,264 coding genes annotated in development and ripening of strawberry fruit[19], among which the expression of FaPE1 is inhibited by IAA and promoted by ethylene, confirm that FaPE1 is a key gene for the softening during fruit senescence[20]. Also, FcNAC1 interacts with FcPL (encoding Fragaria chiloensis pectin lyase) to regulate cell wall remodeling[21]. The expression levels of WRKY31, WRKY70, and NAC83 are correlated with the delayed strawberry fruit senescence, to some extent, NAC83 is used in fruit storability[22]. An enone oxidoreductase (FaEO) contains a calmodulin-binding region and catalyzes the last reductive step in the biosynthesis of 4-hydroxy-2,5-dimethyl-3(2H)-furanone, a major component in the characteristic flavor of strawberry fruit[23,24]. At the molecular level, IAA upregulates the expression of genes associated with the IAA signaling pathways (AUX/IAA, ARF, TOPLESS), E3 ubiquitin protein ligase and annexin. Meanwhile the downregulated genes are involved in pectin depolymerization, cell wall degradation, sucrose and anthocyanin. In contrast, exogenous ABA induces genes related to softening and signaling components (SKP1, HSPs, CK2, and SRG1), showing the cooperative and antagonistic actions between IAA and ABA. This highlights a cross-talk between IAA and ABA potentially by receptor-like kinases and ubiquitin ligases[9]. To a larger extent, secondary metabolites originated from sugars and organic acids play important roles in fruit senescence during storage, including carbohydrate and energy metabolism, volatile biosynthesis, and phenylpropanoid activity[25].
Although much progress has been made on the understanding of the rapid senescence of strawberry fruit, the defined mechanisms are not fully upstanding. In this study, we used the widely-cultivated strawberry 'Benihoppe' under room-temperature conditions to rapidly assess the natural senescent processes and mechanisms of the harvested strawberry fruit. We comprehensively investigated a series line of morphological and physiological changes, including decay extension, weight loss rate, firmness, brightness, soluble sugar content, titratable acid levels, soluble solid content, red/green difference, amino acid content and aroma changes. Additionally, we analyzed phytohormones levels such as abscisic acid (ABA), indole-3-acetic acid (IAA), brassinolide (BR), gibberellic acid (GA), and ethylene (Eth). Furthermore, to understand the physiological variations at the molecular level, we performed RNA-seq and proteome analysis. Finally, based on our gained data set, a physiological, molecular and metabolic model underlying the strawberry fruit senescence was proposed.
-
In this study, the fruits of 'Benihope' strawberry were collected from the Larsen Canyon Strawberry Demonstration Garden in Changping District, Beijing, China. The strawberry fruits were cultivated under controlled greenhouse conditions (20–25 °C, relative humidity of 70%–85%, a natural light/dark cycle of 14/10 h). According to the developmental stages of strawberry fruit[26], the primarily-ripening red fruit at about 28 d after anthesis were transferred from the plants to explore the senescence of harvested strawberry fruit. The fruits were put in plastic boxes covered by white film with numerous holes and stored under room temperature conditions (23–25 °C, 60%–70% humidity).
A total of 270 strawberry fruits were divided into nine groups of biological replicates, with each group consisting of 30 fruits stored in transparent plastic shockproof packaging boxes. Samples were collected at different time points including 0, 2, 4, 6, and 8 d. For physiological index determination and histological sequencing, three groups of storage repetitions were used at each sampling time point, while the remaining three groups were utilized for volatile component analysis. Biochemical indicators were determined by taking five fruits from each storage repeat group on the same day as collection within the same storage stage. These fruits were cut into small pieces and thoroughly mixed to form a composite sample, which was then divided into three groups for detection.
Physicochemical fruit analysis
-
Decay index was according to the determination method[27] with slight modifcation. The spoilage of strawberry fruit was recorded from three groups of biological replicates every 48 h at the sampling time, and the spoilage rate was calculated. Firmness was measured according to the method of Zhang et al.[28] with a slight improvement. The CTX texture instrument (AMETK Broofield, USA) was used to measure the firmness of strawberry fruit. Newton (N) represents the firmness unit. Weight loss rate according to the weighing method[29]. Soluble solid concentration (SSC) was determined using a digital refractometer (TD-45, Zhejiang, China).
The color measurement was conducted using the Minolta Colorimeter CR-300 from Japan, following the method described in Quek et al[30,31]. This colorimeter is based on the CIE (Commission International de L'Eclairage) L*a*b* color space. This system uses three values (L*, a* and b*) to precisely define the location of a color inside a three-dimensional visible color space. Prior to analyzing the samples, calibration was performed on the white color tile with coordinates (L* = 91.0, a* = +0.3165, b* = +0.3326). Duplicate samples were analyzed and the mean was recorded.
Sugars and organic acid extraction and quantification
-
The content of soluble sugar was determined by the plant soluble sugar detection kit (A14511, built in Nanjing, China). The anthrone colorimetric method was used to determine the blue-green furfural derivative generated according to its detection principle. Within a certain range, the color depth is proportional to the sugar content, so the sugar can be quantified.
The manual titration method was modified with reference to Lu et al.[32]. The acid-base neutralization titration method was used for determination. Five strawberry fruits were taken from each of the three groups of biological replicates for sample mixing. The sodium hydroxide standard solution was prepared according to the report above.
Determination of free amino acid content
-
The extraction of free amino acids in fresh fruit was carried out in accordance with the methods of Keutgen & Pawelzik[33] with slightly improved using an automatic amino acid analyzer (L-8900, Hitachi). Referring to the experimental method of He et al.[34] with some improvements, using fluorescence detector (VIS), 570-nm and 440-nm chromatographic column for ion exchange column, column temperature 60 °C, injection volume for 20 μL, mobile phase for citric acid buffer solution, flow rate 0.4 mL/min, column temperature of reaction column after column 135 °C.
Volatile compounds
-
The headspace solid phase micro-extraction method and gas chromatography-mass spectrometry analysis method was used according to the method of Zhao et al.[35] with slight modification. The method including headspace solid phase microextraction (SPME), analysis method of gas chromatography-mass spectrometry (GC-MS), and chromatographic conditions for carrier gas of He. The flow rate is 1 mL/min. The program was set as the temperature rise program: the initial temperature was set to 35 °C, hold for 3 min, rise to 80 °C at 3 °C/min, rise to 200 °C at 9 °C/min, hold for 10 min, and then rise to 210 °C at 10 °C/min, hold for 3 min, and the temperature of the sample inlet was 230 °C. As to mass spectrum conditions, EI was ionization mode, the temperature was set at 230 °C, the electron energy is 70 eV, and the scanning mass range was set at 30−450 nm.
To match the total ion flow diagram with the NIST library mass library, and to qualitatively analyze the identification results of volatile organic compounds in the storage process of fresh strawberry fruit with the matching degree greater than 99% in the mass library, the quantitative analysis was performed by the area normalization method. SPSS software was used to analyze the volatile matter data of 'Benihope' strawberry fruit during storage.
Extraction and quantification of strawberry phytohormones
-
Phytohormones (except ethylene) were extracted from the whole fresh fruit and analyzed as previously described by Maldiney et al.[36]. Finally, the optical density of each hole in the reaction plate was measured at 410 nm using an enzyme-labeled spectrophotometer.
Determination of ethylene production was based on the method published by Zou et al.[37]. Gas chromatography conditions: Agilent 5190-1535 gas injection probe (2.5 mL), Agilent 6890 N gas chromatograph. Hydrogen ion flame (FID) detector, HP-55% Phenol Metal Siloxane chromatographic column. The temperatures of the injection port, detector, and column box were 260, 250, and 40 °C respectively. Hydrogen gas with a flow rate of 30 mL/min is used as the carrier gas. Nitrogen gas with a flow rate of 25 mL/min was used as the tail blowing gas, and the sample volume was retained for 5 min, with an injection volume of 1 mL. The experiment to determine the ethylene release rate of strawberry fruit during harvest storage was conducted at five sampling time points, including 0, 2, 4, 6, and 8 d of storage, divided into five groups of treatments. The strawberry fruits were approximately 153 g from each group and stored in a 1-L aluminum foil composite membrane gas sampling bag. The bag was be placed at room temperature for 2 h to collect gas. A 2.5 mL gas injection needle was employed to extract 1 mL of gas samples from each sample bag, and the ethylene concentration was subsequently measured by a gas chromatograph. The experiment was repeated three times and the average values were taken. The results of the significance analysis were expressed as 'mean ± standard deviation'.
RNA extraction and RNA-seq
-
Based on our previous report on RNA-seq analysis of strawberry fruit[38], 15 harvested strawberry ripe fruits (the 0-d, 2-d, 4-d, 6-d and 8-d after harvest; five groups and three biological replicates) were prepared for transcriptome sequencing. Total RNA was extracted using a RNeasy Plant Mini Kit (Qiagen, Dusseldorf, Germany). RNA purity and quantification were evaluated using a NanoDrop 2000 spectrophotometer (Termo Scientifc, USA). RNA integrity was assessed using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). The libraries were constructed using TruSeq Stranded mRNA LT Sample Prep Kit (Illumina, San Diego, CA, USA) and sequenced on an Illumina NovaSeq 6000 platform by OE Biotech Co., Ltd. (Shanghai, China). The clean reads were assembled using Trinity software (
https://github.com/trinityrnaseq/trinityrnaseq/releases ) to obtain the long-contig transcriptome. Hierarchical cluster analysis of differentially expressed genes (DEGs) was performed to investigate the genes expression pattern of different samples. After reads reassembled by StringTie software, novel transcripts identification was performed by comparing the octoploid strawberry genome (Fragaria ananassa,www.rosaceae.org/Analysis/12335030 ) using Cufcompare software. ASprofile software was used to analyse the alternatively splicing of differentially regulated transcripts isoforms. According to the expression level of the protein-coding gene among the different treatments, a significant difference in expression was obtained as |log2 fold change| ≥ 2. The RNA-seq was performed with three repeats.Proteome analysis of stored strawberry fruit
-
Based on our previous report on proteome analysis of strawberry fruit[39], 15 ripe fruits (the 0-d, 2-d, 4-d, 6-d and 8-d after harvest; five groups and three biological replicates) were used for DEP (differentially expressed proteins) by proteome analysis with TMT labeling technique in JINGJIE PTM Biolab Co. Ltd. (Hangzhou, China). Differentially abundant proteins were defined as those with abundances that were > 1.3-fold (p < 0.05) different among fruit developmental stages. Proteins were functionally classified based on Gene Ontology terms from the following three main categories: biological process, cellular component, and molecular function. Additionally, the enriched KEGG pathways (
www.genome.jp/kegg ) among the differentially abundant proteins were identified based on a two-tailed Fisher's exact test. The GO/KEGG annotations with a corrected p-value < 0.05 were considered significant.Statistical analysis
-
ANOVA was performed using the SPSS version 23.0 (SPSS Inc., NY, USA). Data means were compared using Duncan's multiple range test at p < 0.05. OriginPro2019b (OriginLab Institute Inc., Northampton, MA, USA) was used to create graphs. Experimental results were presented as the mean of three biological replicates ± standard deviation.
-
The results showed a series of obvious morphological and physiological changes in ripe fruit were observed over time (Fig. 1). The freshness gradually lost, as evidenced by the combination of the continually-increased decay index (Fig. 1b) and water loss (Fig. 1c) and the continually-decreased firmness (Fig. 1d) and brightness (Fig. 1e). Interestingly, the most important quality parameters (including sugar, acid and soluble solids concentration) exhibited complex variations, such as the soluble sugar content showed first increased during 0−2 d then continually decreased, while it remained high level at 6 d in comparision to 0 d (Fig. 1f). In contrast, although both titratable acid content (TAC) and soluble solids concentration (SSC) also were increased during 0−2 d, hereafter declined rapidly (Fig. 1g & h). Given de-greening and red-coloring were coupled with strawberry fruit ripening, thus we detected the red and green color difference during storage, but not showing significance (Fig. 1i).
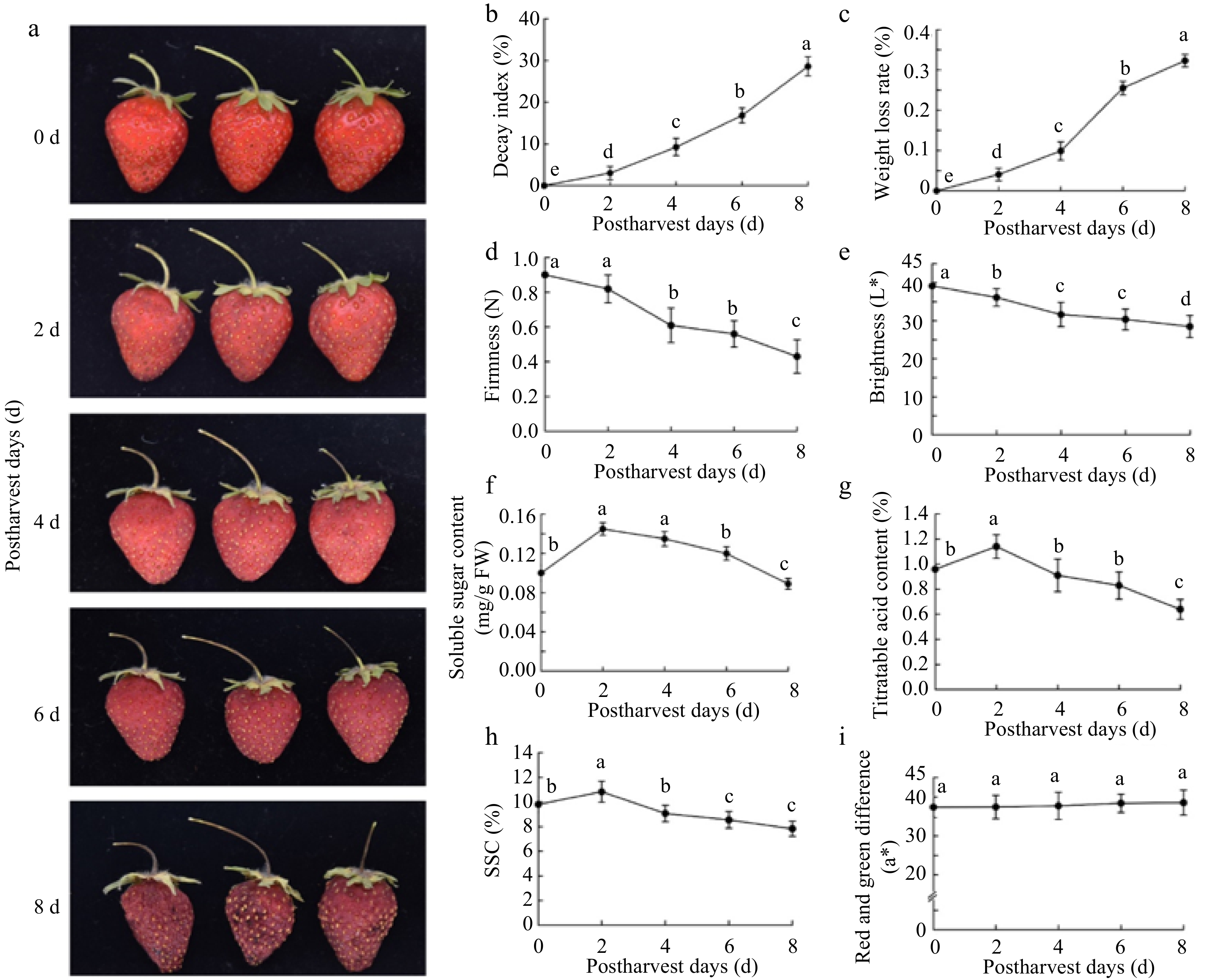
Figure 1.
Morphological and physiological changes of the strawberry fruit at room temperature during 0−8 d storage. The parameters include, (a) external morphological changes, (b) decay index, (c) weight loss rate, (d) hardness, (e) brightness, (f) soluble sugar content, (g) titratable acid content, (h) soluble solid content, and (i) red and green difference. The error bars represent the standard error (n = 3). For each storage day, different letters above lines indicate significant differences according to Duncan's multiple range test, at p < 0.05.
Changes in phytohormones levels
-
To further understand the physiological mechanism of strawberry fruit senescence, the levels of phytohormones including ABA, IAA, BR, GA3, MeJA and ethylene, were investigated over time during storage. Despite ABA which showed a constant-increase over time (Fig. 2a), other hormones showed complex variations during storage (Fig. 2b−f), especially IAA indicative of complex changes by a 'W' curve. Notably, apart from a similar trend during 2−4 d stages, BR and MeJA showed different variations (Fig. 2c & e), while GA3 and ethylene (Eth) increased only after 4 d, especially Eth increased rapidly (Fig. 2d & f). These data suggested an important role of ABA in the senescence of harvested strawberry fruit over time (0−8 d). It was concomitant with the potential role of MeJA at early (0−2 d), IAA at prometaphase (2−4 d), BR at the middle (2−6 d), GA3 at postmetaphase (4−6 d), and Eth at later (4−8 d) during the senescent stages, respectively. Given the rapid senescence occurred at 2 d after harvesting, hereafter the rapidly-accumulated hormones including ABA, BR, MeJA and IAA were involved in the early senescence, while ethylene was implicated in the later postharvest senescence of the harvest fruit. Altogether, ABA is a critical regulator for the senescence of harvested strawberry fruit, concomitant with its early collaboration with BR, MeJA and IAA, latter with ethylene.
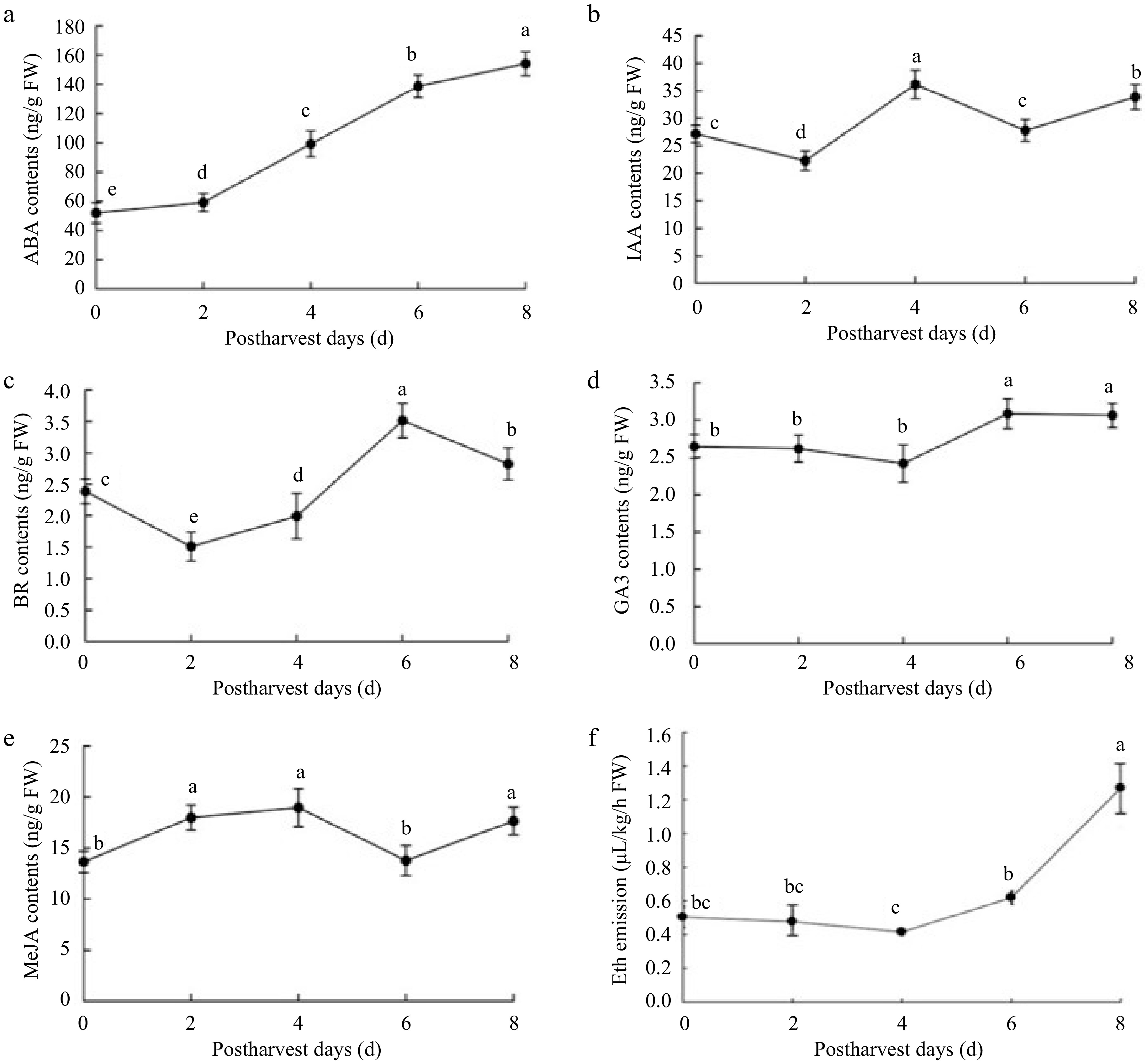
Figure 2.
Hormonal concentration changes in strawberry fruit at room temperature during 0−8 d storage. The parameters included (a) abscisic acid (ABA), (b) indole acetic acid (IAA), (c) brassinolide (BR), (d) gibberellic acid (GA3), (e) MeJA, (f) ethylene (Eth). The error bars represent the standard error (n = 3). For each postharvest day, different letters above lines indicate significant differences according to Duncan's multiple range test, at p < 0.05.
RNA-seq for differentially-expressed genes and pathways relate to the senescence of strawberry fruit
-
After removing low-quality and primer/adapter-containing reads, 95.05 Gb clean reads were generated with an average of 6 Gb per sample, and Q30 levels reaches over 92% (Supplemental Table S1). A total of 66,533 unigenes were obtained, reaching match ratios of 88.96%−89.28% to the octaploid strawberry reference genome[40]. These data indicate the RNA-seq quality to meet with subsequent biological analyses.
On the basis of our RNA-seq data and pathway enrichment analyses, a maximum number of 3,861 DEGs (differentially expressed genes) were screened between 2-d unigenes and 8-d unigenes, here named as 2−8 (3,861), after a combination of the pair analyses among 0-d, 2-d, 4-d, 6-d and 8-d unigenes and five top pair DEG analyses, including 0−2 (663), 2−6 (981), 0−8 (1,928), 4−8 (2,866), 2−8 (3,861) screened by |log 2 FC| > 2 and p-adjust < 0.05 (Fig. 3a). Indeed, this notion is confirmed by the analysis of 12 DEG clusters, varied with different turning points over time by the Fuzzy cMeans method (Fig. 3b), among which the three pathways of DEGs were involved in plant hormone signal transduction, phenylpropanoid biosynthesis, and flavonoid biosynthesis (Supplemental Fig. S1). Next, we focused on the potential DEGs key to plant hormone metabolism and signal transduction pathways, highlighting on ABA with NCEDs and PYLs, auxin with IAAs and AUXs, ethylene with ACSs, EIN3 and ERFs, BR with BZRs, and JA with JMTs (Supplemental Table S2), suggesting a potential role of ABA, linked to a concerted regulation with Eth, IAA, BR and JA in the senescence of postharvest strawberry fruit.
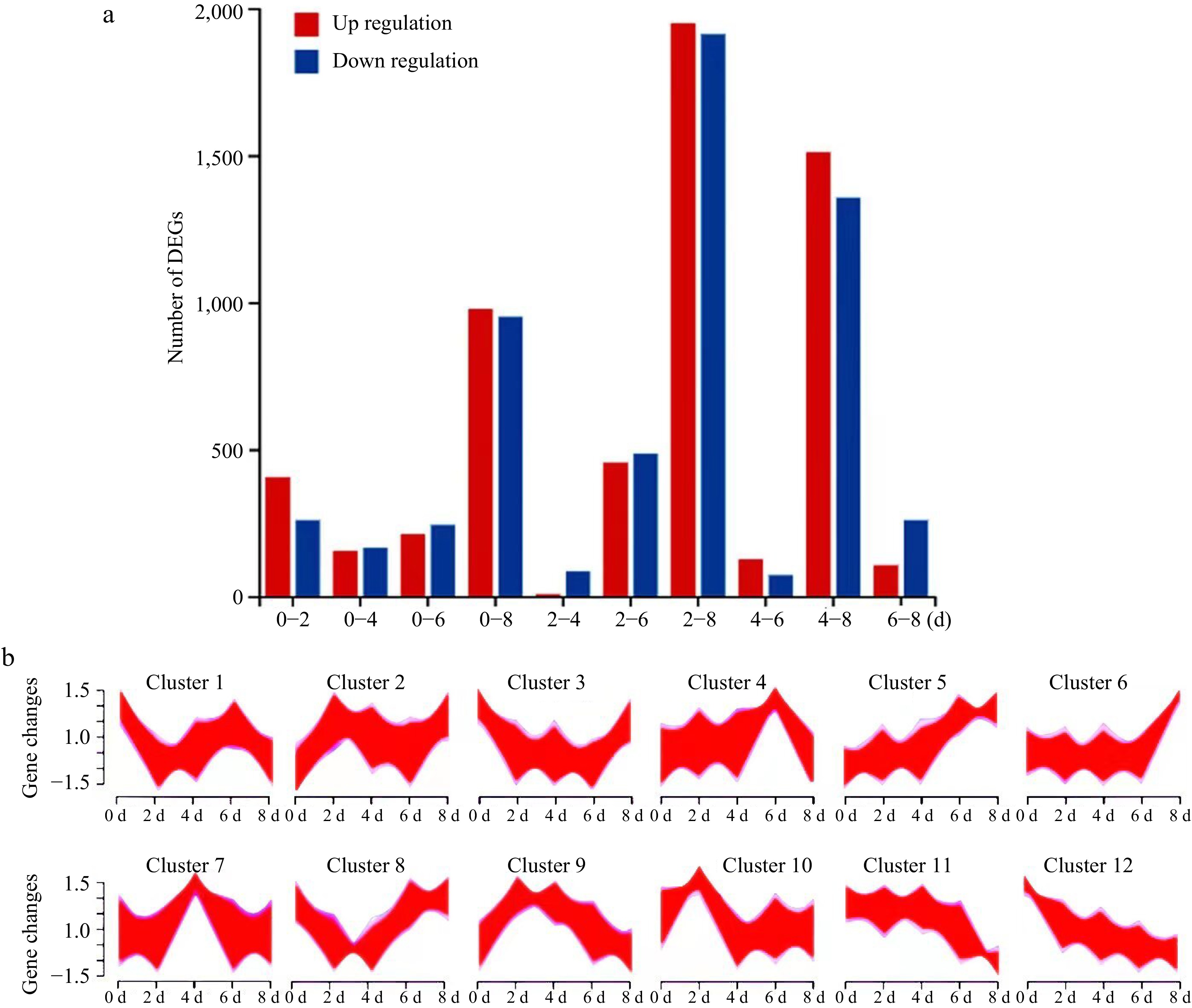
Figure 3.
The numbers and clusters of DGEs in strawberry fruit over time at room temperature during 0−8 d storage. (a) DGEs by pair comparison. The '0−2' represents DGEs by pair comparison between 0-d identified proteins and 2-d identified proteins, and so on for 0−4, 0−6, and 0−8 DGEs by pair comparison. The '0−2' represents DGEs by pair comparison between 0-d unigenes and 2-d unigenes, and so on for 0−4, 0−6, 0−8, 2−4, 2−6, 2−8, 4−6, 4−8, and 6−8. (b) DGE clusters over time. DGEs: differentially expressed genes.
Proteome-analysis of differentially-expressed proteins and pathways during the senescence of strawberry
-
A total of 33,138 peptides were identified, from which a total 21,253 unique peptides were screened, and 8,208 proteins could be quantified by the TMT labeling technique, among which 648 DEP (differentially expressed proteins) could be obtained. The results showed that the numbers of DEPs increased over time (Fig. 4a), which could be largely divided into seven clusters (Fig. 4b). An increased-expression trend was included Cluster 2 (85 proteins), highlighting on plant hormone signal transduction, α-linolenic acid metabolism involved in two jasmonate methyltransferases (JMTs) and four lipoxygenases (LOXs). A decreased-expression trend was included Cluster 7 (106 proteins), highlighting on phenylpropanoid biosynthesis, starch and sucrose metabolism, steroid biosynthesis, and SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) interactions in vesicular transport. There was a higher expression at 2 and 4 d which included Cluster1 (90 proteins) and Cluster 5 (103 proteins), highlighting on the Cluster 1 of pentose and glucuronate interconversions, MAPK (mitogen-activated protein kinase) signaling pathway, starch and sucrose metabolism, plant-pathogen interaction, the Cluster 5 of amino sugar and nucleotide sugar metabolism, photosynthesis, and phagosome. A lower expression at 2 and/or 4 d was included in Cluster 3 (80 proteins) and Cluster 4 (70 proteins), highlighting on the Cluster 3 of citrate cycle (TCA cycle), glycolysis/gluconeogenesis pentose, pyruvate metabolism and spliceosome, the Cluster 4 of protein processing in endoplasmic reticulum. An increased-expression trend after 4 d was included in Cluster 6 (114 proteins), highlighting on glutathione metabolism, monoterpenoid biosynthesis, and RNA transport. Notably, the plant hormone signal transduction was highlighted on ABA with PYL, SnRK2, ABF; Eth with ETR, CTR1, EIN2; BR with BSK; GA with GID1; JA with JAR1, JAZ (Fig. 5), while IAA-related DEPs were not screened, suggesting an important role of ABA, JA and Eth in senescent metabolism.
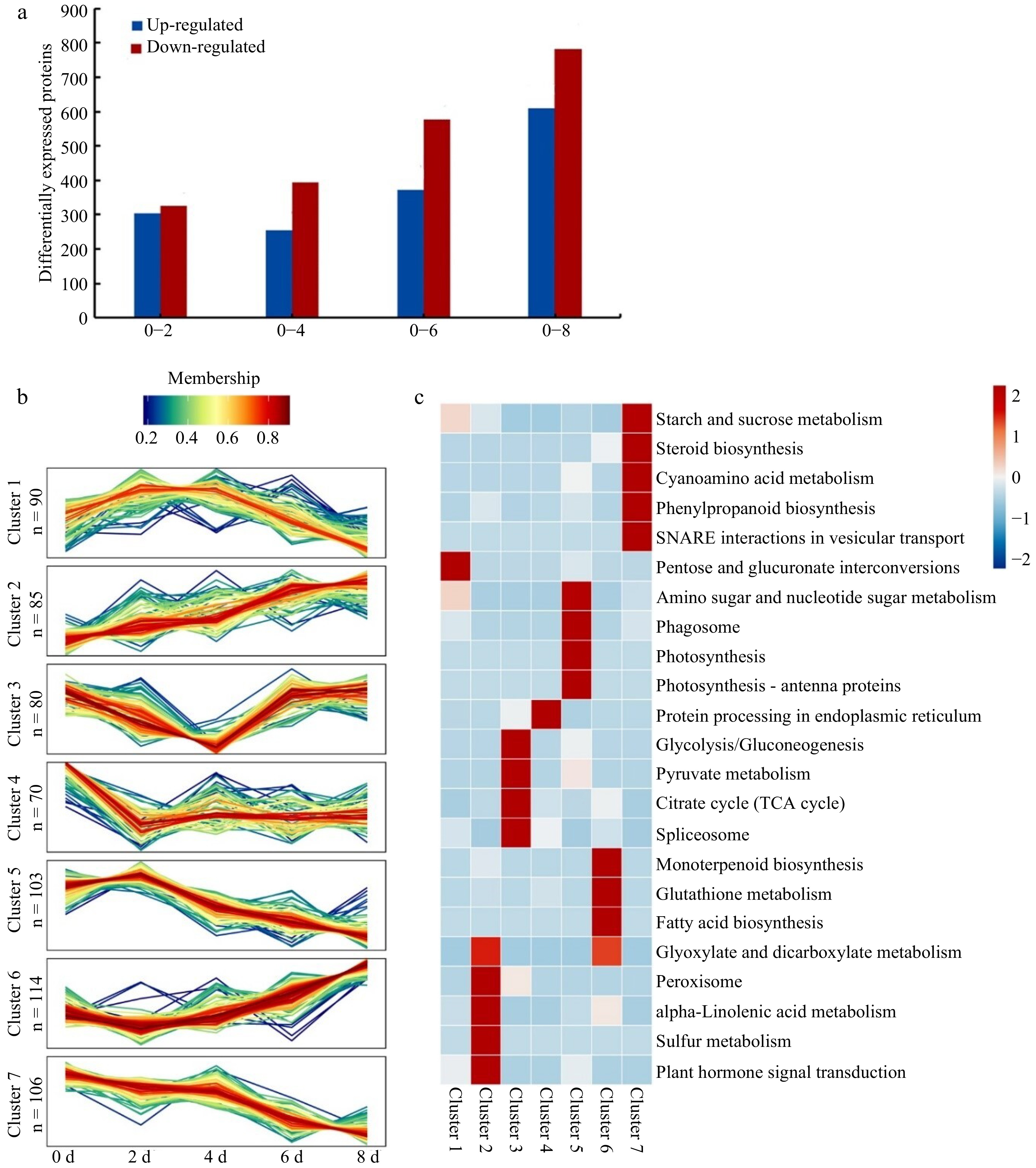
Figure 4.
The numbers, pathways and clusters of DEPs in strawberry fruit over time at room temperature during 0−8 d storage. (a) Differentially expressed proteins (DEPs) by pair comparison. The '0−2' represents DEPs by pair comparison between 0-d identified proteins and 2-d identified proteins, and so on for 0−4, 0−6, and 0−8. (b) The DGP clusters and highlighting pathways over time. DEPs: differentially expressed proteins. The n represents the numbers of DGPs in a cluster. (c) Enrichment of differentially expressed protein cluster KEGG pathway.
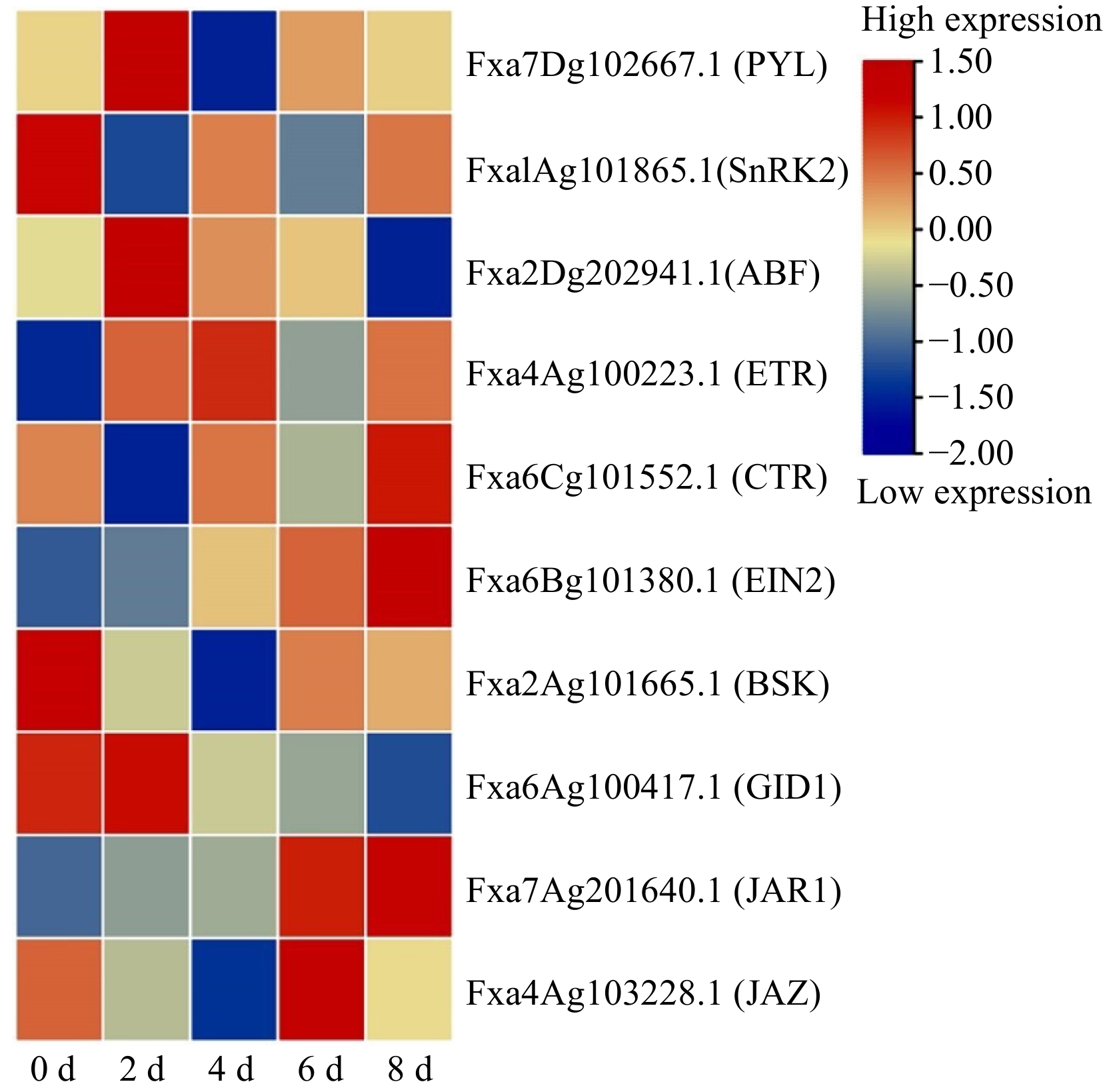
Figure 5.
Heatmap analysis of differentially expressed proteome related to strawberry fruit senescence during storage. PYL: PYR1-like/regulatory components of ABA receptor proteins (PYR/PYL/RCARs); SnRK2: SNF1-related kinase 2; ABF: ABRE (ABA-responsive element binding protein)-binding factor); ETR: ethylene triple response; ETR2: CTR: constitutive triple response; EIN2: Ethylene Insensitive 2; BSK: brassinolide signal kinase; GID1: gibberellin-insensitive dwarf1 (gibberellin receptor); JAR1: jasmonate resistant 1; JAZ: jasmonate-ZIM domain.
The 'V' curve in Cluster 3 (Fig. 4b & c) might represent a metabolism transition at 4 d. It showed high pentose and glucuronate interconversions and MAPK signaling pathway before the point of the 4th day. However, it showed a high glutathione metabolism and terpenoid biosynthesis after the point of the 4th day. We proposed that the senescence is coupled with sugar metabolism and gluconeogenesis, signaling, anti-stress, rancidity, and aroma, which were involved in ABA, Eth, BR, JA, and GA. Altogether, these data suggested that the senescence of strawberry fruit was controlled by post-transcriptional regulation, mostly involved in the high turnover of the core metabolisms concerning starch and sucrose as well as phenylpropanoids (mostly by phenylalanine aminolyase 2, PAL2). Meanwhile, it was coupled with high metabolisms of alpha-linolenic acid, glyoxylate and dicarboxylate, peroxisome, and vitamin B6.
Concentration changes in volatiles and free amino acids during the senescence of harvested strawberry
-
To understand some important metabolic changes related to the senescence, we investigated the contents of the volatiles and amino acids in strawberry fruit. The results showed that only 16 volatile substances were detected at 0 d, among which nerolidal accounted for 57.96%. The ethyl caproate was the dominant component at 2 d. Whereas the number of volatile substances increased to 68 species at 8 d that mainly included esters and alcohols (Fig. 6). It was worth noting that volatile acids, such as 2-amino-6-methylbenzoic acid, began to appear at 6 d, which indicated that the fruit would be initiated to deteriorate. Altogether, nerolidal and ethyl caproate represent the characteristic aroma of the ripe 'Benihope' strawberry fruit, concomitant separately with the ripening and senescent fruit.
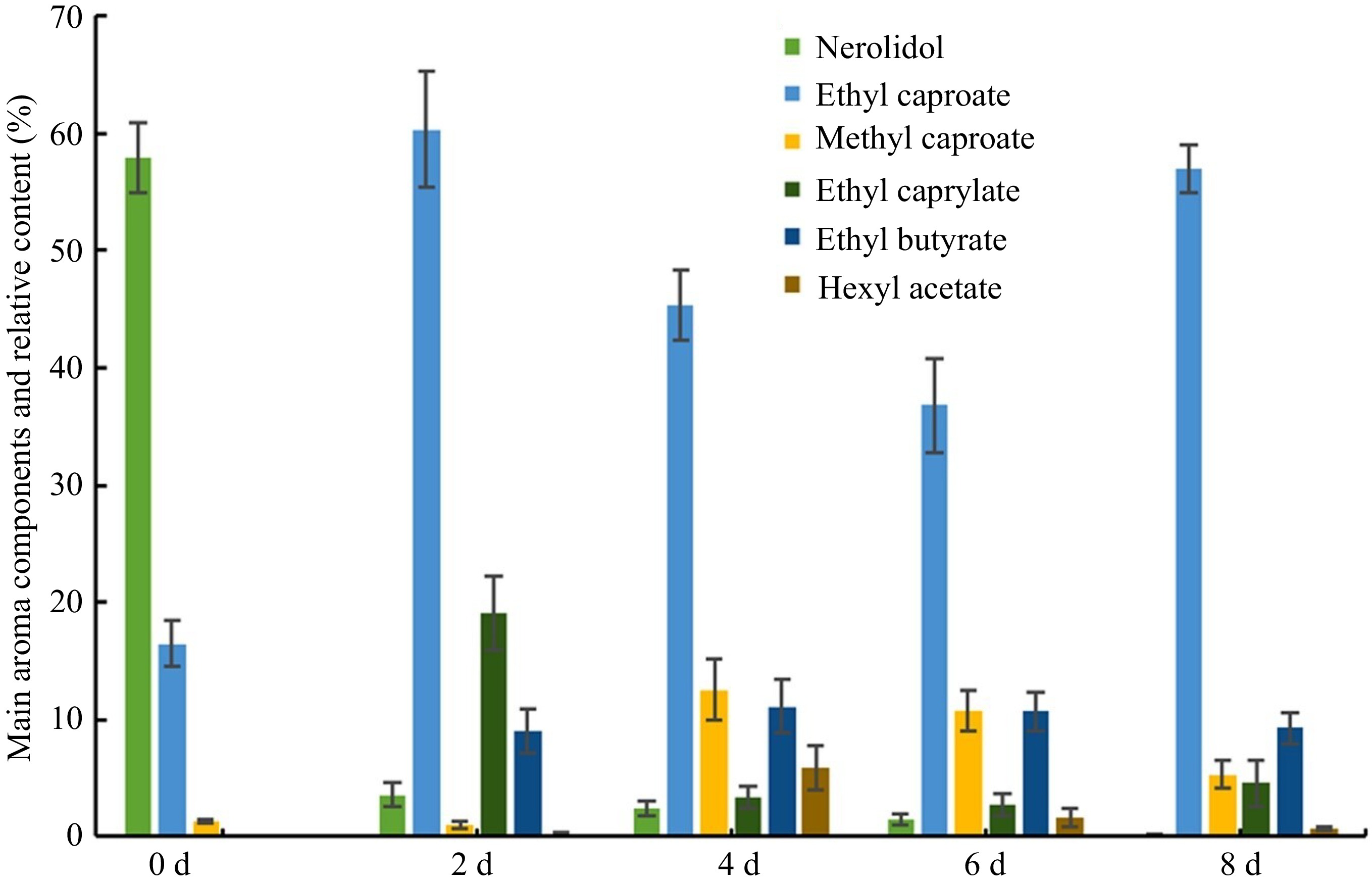
Figure 6.
Changes in the relative contents of the main aroma components in strawberry fruit over time.
In addition, a total of 16 amino acids were detected during 0−8 d (Table 1). After harvesting strawberry fruit, the total amino acid content decreased with the extension of time. The total content of amino acids at 0 d (3.904 µg/g) was significantly higher than that at 8 d (2.309 µg/g). Notably, Thr showed the highest level (2.085 µg/g), accounting for 53.4% of the total content at 0 d (3.904 µg/g). While cysteine (Cys) was detected only at the later stage of storage (6 and 8 d) (Table 1). Altogether, the higher levels of nerolidal, ethyl caproate, and Thr might represent the characteristic components of the ripe 'Benihope' strawberry fruit, while the detection of Cys and ethyl caproate represent the severe senescence.
Table 1. Changes in free amino acid content in the 'Benihope' strawberry fruit during storage.
Free amino acid (µg/g) 0 d 2 d 4 d 6 d 8 d Threonine (Thr) 2.085 ± 0.091a 1.539 ± 0.045b 1.085 ± 0.062c 1.463 ± 0.079b 0.824 ± 0.025c Alanine (Ala) 0.901 ± 0.028a 0.735 ± 0.032b 0.487 ± 0.024d 0.549 ± 0.018c 0.717 ± 0.010b Glutamic acid (Glu) 0.313 ± 0.012a 0.272 ± 0.027ab 0.234 ± 0.008b 0.268 ± 0.040ab 0.216 ± 0.052b Aspartic acid (Asp) 0.165 ± 0.042a 0.165 ± 0.040a 0.185 ± 0.046a 0.188 ± 0.037a 0.127 ± 0.047a Serine (Ser) 0.152 ± 0.026a 0.139 ± 0.028a 0.134 ± 0.034a 0.158 ± 0.082a 0.184 ± 0.058a Valine (Val) 0.127 ± 0.035a 0.141 ± 0.023a 0.107 ± 0.028a 0.111 ± 0.066a 0.091 ± 0.029a Phenylalanine (Phe) 0.037 ± 0.042a 0.035 ± 0.044a 0.029 ± 0.027a 0.035 ± 0.014a 0.017 ± 0.032a Glycine (Gly) 0.026 ± 0.015a 0.021 ± 0.011a 0.025 ± 0.012a 0.027 ± 0.014a 0.031 ± 0.017a Leucine (Leu) 0.024 ± 0.011ab 0.015 ± 0.021b 0.029 ± 0.035ab 0.011 ± 0.006b 0.039 ± 0.008a Lysine (Lys) 0.021 ± 0.020a 0.024 ± 0.013a 0.019 ± 0.015a 0.024 ± 0.012a 0.015 ± 0.014a Tyrosine (Tyr) 0.015 ± 0.005a — — — 0.009 ± 0.001a Isoleucine (Ile) 0.014 ± 0.002a 0.015 ± 0.008a — — — Histidine (His) 0.013 ± 0.007a 0.015 ± 0.009a — 0.009 ± 0.005a 0.008 ± 0.003a Methionine (Met) 0.011 ± 0.005a — — 0.011 ± 0.009a 0.014 ± 0.006a Arginine (Arg) — 0.009 ± 0.004b 0.024 ± 0.083a — 0.012 ± 0.008a Cysteine (Cys) — — — 0.006 ± 0.001a 0.005 ± 0.002a Total amino acids 3.904 ± 1.52a 3.124 ± 1.26ab 2.358 ± 0.34ab 2.855 ± 0.45ab 2.309 ± 0.27b —, representing not detectable. For each free amino acid, different letters within a row indicate significant differences according to Duncan's multiple range test at p < 0.05. -
It is commonly accepted that the ripening of strawberry, which serve as a model for non-climacteric fruit, is primarily regulated by ABA[18,26]. Furthermore, considerable advancements have been made in recent years towards comprehending the senescent mechanisms in postharvest fruits[4,9,11,16,17,41], which involves hormones, phenylpropanoid/flavonoid, antioxidant regulators, and softening enzymes, serving as common events in ripening and senescence[4,11,13,16,17, 42−46], where ROS, ABA, ethylene and IAA are regarded as being key players[9, 11,15,25].
In the present study, our results confirmed that the rapid softening and water loss of the postharvest fruit led to shorter storage, which was coupled with an increase in soluble sugars during the early stage within 2 d (Fig. 1), suggesting the good quality with the highest levels of nerolidal and Thr; while the detection of Cys and ethyl caproate represents severe senescence (Fig. 6 & Table 1). These processes were concomitant with a continually-increased trend of ABA, which coordinated JA, BR and Eth separately with the early, middle, and later senescent stages; while IAA participated in senescent metabolic transition but a lesser role is played by GA (Fig. 2). Thus, we consider that ABA may be critical to the senescence of harvested strawberry fruit.
Interestingly, our transcriptome and proteome revealed that the senescence-related top pathways by DEGs pointed to plant hormone signal transduction, phenylpropanoid biosynthesis, and flavonoid biosynthesis, highlighting on a role of ABA (NCEDs, PYLs), IAA (IAAs, AUXs), Eth (ACSs, EIN3 and ERFs), BR (BZRs), JA (JMTs) in the senescence (Supplemental Table S2); meanwhile DEPs pointed to plant hormone signal transduction, α-linolenic acid, phenylpropanoids, and MAPK signaling, highlighting on a role of ABA (PYL, SnRK2, ABF), Eth (ETR, CTR1, EIN2), JA (JAR1, JAZ), BR (BSK), and GA (GID1) in the senescence (Fig. 5). Taken together, the combination of omics and hormone levels strongly suggest that ABA, Eth and JA may play significant roles during harvested strawberry fruit senescence, among which ABA is a central player, channeling to regulation of aroma by LOXs, phenylpropanoids by PAL2 and anti-stresses by MAPK signaling (Fig. 4).
Since fruit ripening and senescence are an irreversible biological process, in a combination of our previous studies[26] and the present study, we consider that ABA not only plays an important role in regulating strawberry fruit ripening, but also controls strawberry fruit senescence, mostly linked to rapid softening and decay, as well as aroma emission mostly by coordination with JA and Eth. Finally, on the basis of an integration of RNA-seq and proteome data as well as hormone content, a physiological, molecular and metabolism model for strawberry fruit senescence is proposed (Fig. 7).
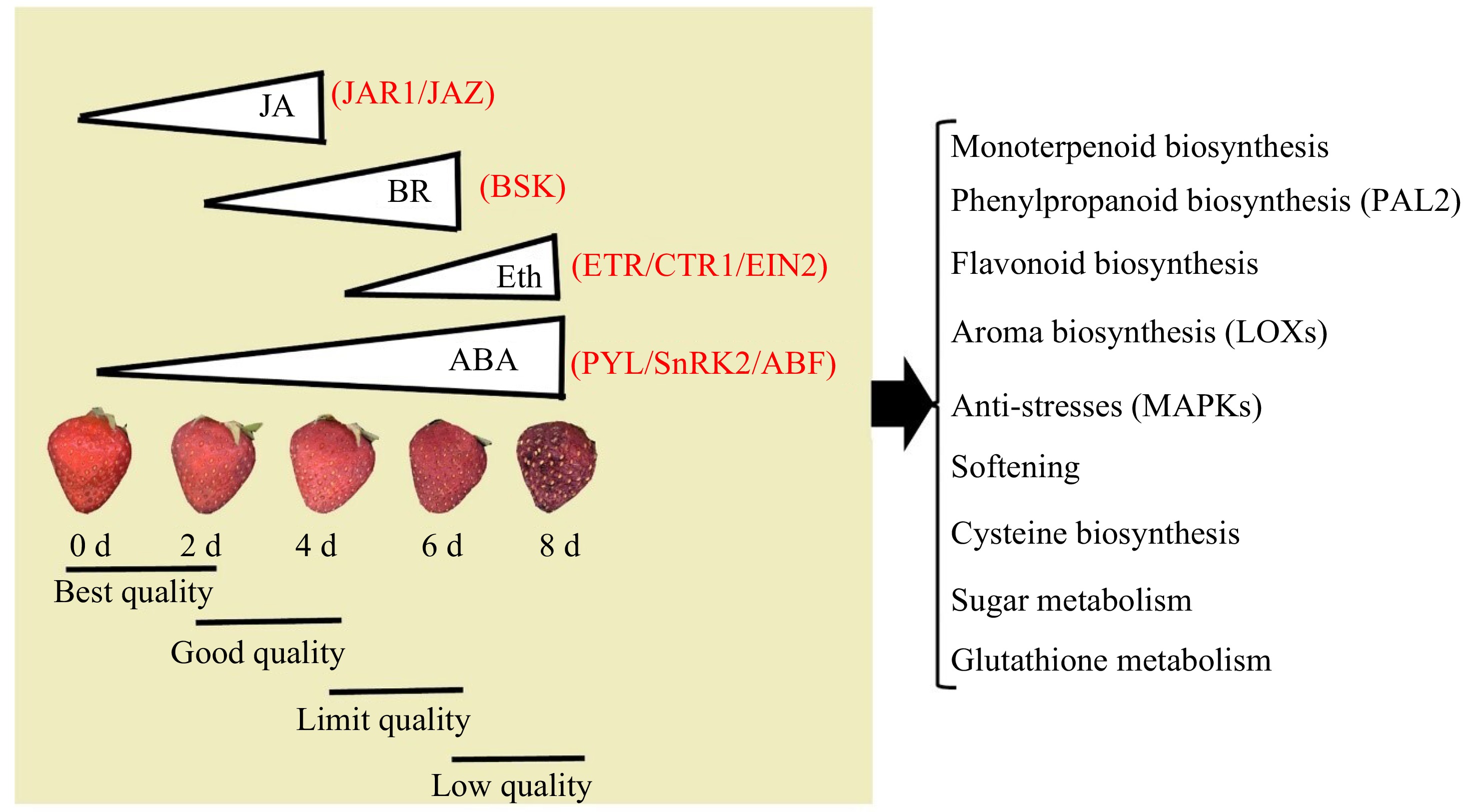
Figure 7.
A physiological, molecular and metabolism model for strawberry fruit senescence from the best quality to low quality. The central hormone of ABA promotes the fruit rapid senescence from the best, good, and limited quality to low quality during 0−8 d, by a coordinated regulation of the early stage (0−4 d) with JA, middle stage (2−6 d) with BR, and later stage (4−6 d) with Eth. They trigger a series of signaling transduction and metabolisms: the former includes ABA with PYL/SnRK2/ABF, JA with JAR1/JAZ, BR with BSK, Eth with ETR/CTR/EIN2; the later includes monoterpenoid, phenylpropanoids, and flavonoid biosynthesis, as well as sugar metabolism, together with aroma and cysteine biosynthesis, anti-stresses, and softening. PYL: PYR1-like/regulatory components of ABA receptor proteins (PYR/PYL/RCARs); SnRK2: SNF1-related kinase 2; ABF: ABRE (ABA-responsive element binding protein)-binding factor); ETR: ethylene triple response; CTR: constitutive triple response; EIN2: Ethylene Insensitive 2; BSK: brassinolide signal kinase; GID1: gibberellin-insensitive dwarf1 (gibberellin receptor); JAR1: jasmonate resistant 1; JAZ: jasmonate-ZIM domain.
-
Overall, our current study demonstrated that the continuously elevated central hormone ABA promoted rapid senescence in harvested strawberry 'Benihoppe', resulting in varying fruit qualities categorized as best, good, limit, and low. This finding was consistent with the coordinated regulation of early-stage JA signaling, middle-stage BR signaling, and late-stage Eth signaling by a series of core components. Ultimately, these hormonal signals regulate multiple biochemical metabolisms including monoterpenoid biosynthesis, phenylpropanoid biosynthesis, flavonoid biosynthesis, sugar metabolism, aroma and cysteine biosynthesis; they also contribute to anti-stress responses and softening processes. These findings provided a comprehensive understanding that fruits aged 2-d are preferred while those at 4-d day are optimal, and the 6-d fruit are limited under given storage conditions.
-
The authors confirm contributions to the paper as follows: study conception and design: Shen Y, Duan L, Chen X; data collection: Yan B, Bai Y, Liu Z; analysis and interpretation of results: Yan B, Wang Y, Bai Y; draft manuscript preparation: Shen Y, Yan B; review and editing: Liu H; project administration and funding acquisition: Shen Y. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This study was funded by the China National Science Foundation (Projects 32030100).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Boqian Yan, Ye Wang, Yuan Bai
- Supplementary Fig. S1 Differential gene KEGG Pathway enrichment.
- Supplementary Table S1 Assemble and compare transcriptome data of strawberry.
- Supplementary Table S2 Differential gene related to plant hormone metabolism and its signal transduction pathway.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yan B, Wang Y, Bai Y, Liu Z, Liu H, et al. 2024. Insights into the senescent mechanisms of harvested strawberry fruit at the physiological, molecular and metabolic levels. Fruit Research 4: e018 doi: 10.48130/frures-0024-0011 

Insights into the senescent mechanisms of harvested strawberry fruit at the physiological, molecular and metabolic levels
- Received: 26 December 2023
- Revised: 20 February 2024
- Accepted: 26 February 2024
- Published online: 06 May 2024
Abstract: Strawberry (Fragaria × ananassa) is a worldwide cultivated horticultural crop, however, its short preservative life of the harvested fruit remains a challenge to be addressed. Currently, although much progress has been made toward understanding the senescence of harvested strawberry fruit, the defined mechanisms remain unclear. Therefore, we performed a series of morphological, physiological and biochemical, as well as transcriptome and proteome analyses using the widely-cultivated strawberry 'Benihoppe' during 0−8 d at room temperature. The results showed not only the shorter storage of harvested strawberry fruit resulted from the rapid perishability, softening, and water loss, but also an increase in soluble sugars within 2 d and a coordination of ABA with JA at the early stage, BR at the middle stage and ethylene at the later stage, respectively. The RNA-seq data highlighted on ABA with NCEDs and PYLs, auxin with IAAs and AUXs, ethylene with ACSs, EIN3 and ERFs, BR with BZRs, and JA with JMTs; while proteome data highlighted on ABA with PYL/SnRK2/ABF, JA with JAR1/JAZ, GA with GID1, BR with BSK, and ethylene with ETR/CTR/EIN2, suggesting an important role of ABA, JA, and ethylene in the senescence of harvested strawberries. Interestingly, higher contents of nerolidalyl caproate and threonine represented characteristic signs of ripening and senescence. Finally, a physiological, molecular and metabolic model for strawberry fruit senescence is proposed, providing comprehensive insights into the preservative mechanisms.
-
Key words:
- Strawberry fruit /
- Postharvest /
- Senescence /
- Phytohormone