









-
Fe-deficiency affects plant growth mainly by interfering with chlorophyll biosynthesis, photosynthesis, and respiration[1]. When plants are Fe-deficient, they exhibit leaf chlorosis, which in extreme cases can lead to complete plant death[2,3]. Yellowing is caused by the imbalance and breakdown of chlorophyll, the synthesis of which is regulated by abscisic acid. In recent years, ABA signaling, enhanced by low temperatures, can be transcriptionally activated by the downstream transcription factors known as AREBs (ABRE-binding proteins). These factors bind to ABRE (ABA response cis-elements) on the promoters of the CORs gene, leading to transcriptional activation and improved plant cold tolerance[4]. Abscisic acid (ABA) plays an important role in this signal transduction. In the presence of ABA, the binding of ABA to the PYR/PYL/RCAR receptor releases the restriction of SnRK2 by PP2Cs, and thus the SnRK2 protein kinase is activated and fulfills its role. These activated SnRK2s can phosphorylate the downstream transcription factors AREB/ABF, which in turn activate the ABRE-dependent expression of ABA-responsive genes such as RD29A, and CBFs[5,6]. The SnRK2 family is thus a key component in the ABA signaling pathway. The study of key genes in the Fe uptake process is therefore crucial for apples to counteract Fe-deficiency stress.
The protein kinase SnRK2 plays a crucial role in the ABA signaling pathway and is essential for plant resistance to stress, especially abiotic stresses. Transgenic plants of Arabidopsis thaliana, wheat, and poplar with SnRK2 exhibit enhanced drought resistance[7−9]. Although ABA does not induce Arabidopsis SRK2E/SnRK2.6 at the mRNA level, the protein is still capable of being activated in response to ABA and drought stress, suggesting post-transcriptional regulation mechanisms[10]. The mutant Arabidopsis SRK2E is significant in ABA signaling during water stress, preventing stomatal closure under rapid humidity reduction[11]. Three SnRK2 protein kinases, SnRK2.2, SnRK2.3, and OST/SRK2E/SnRK2.6, have been found to positively regulate ABA signaling in Arabidopsis thaliana[12]. Overexpression of SAPK8, SAPK9, and SAPK10 enhance ABA responses[13], the SnRK2 protein kinase-mediated processes such as seed germination, plant growth, and development[14−16]. Arabidopsis thaliana with AtSnRK2.8 transgene exhibits increased growth compared to the wild type, illustrating its growth-promoting potential[17]. Therefore, the SnRK2 family, which is widely present in higher plants, participates in various plant signaling pathways and contributes significantly to the plant stress tolerance mechanism[18].
Malus halliana Koehen, an apple rootstock native to the arid, saline-alkali areas of Gansu Province (China), has been the subject of relatively few and sporadic studies[19]. Despite this, the studies have shown it has excellent stress tolerance[20]. Moreover, the disadvantages caused by irrational human activities, such as irrational irrigation, irrational fertilizer application, etc., resulting in soil saline-alkali conditions has become increasingly severe, effective Fe use has decreased, and Fe-deficiency stress has become a major constraint to apple industry development[21]. Screening for genes related to Fe-deficient stress should help produce plants that are more resistant to Fe-deficiency. Based on qRT-PCR, the MhSnRK2.4 gene was screened, which was significantly up-regulated under Fe-deficiency stress, but the mechanism of its regulation under Fe-deficiency stress is still unknown. For this reason, the gene was cloned, and its function was characterized in Arabidopsis and apple calli.
-
For gene expression analysis, Malus halliana histoculture seedlings were utilized as the experimental material. Initially, these seedlings were cultivated on a basal medium for 3 weeks. Subsequently, they were subjected to uniform treatment conditions and maintained under regular control. In parallel, the wild-type (WT) Arabidopsis thaliana of the Colombian ecotype was employed as a comparative model. Additionally, 'Wang Lin' apple calli, which were originally sourced from the laboratory of Professor Wang Xiaofei at Shandong Agricultural University (Shandong, China), were also included in the present study. These calli were further grown in the laboratory under the conditions specified for the experiment.
For gene expression analysis, pest and disease-free Malus halliana seedlings with eight leaves were collected and precultivated for 7 d on Fe-supplemented (+Fe) MS medium. After 7 d of cultivation, Fe-deficiency stress treatments were applied, and the leaves were collected at 0, 12, 24, 48, and 72 h. In addition, Fe-deficient solutions were adjusted to 0 μM Fe (−Fe) according to the Fe assays of Han et al.[22]. Each treatment was replicated three times, with each replicate containing five plants.
Quantitative real-time PCR
-
Total RNA was extracted from the samples using an RNA extraction kit from BioTeke Corporation (Beijing, China). The RNA was then reverse transcribed using TaKaRa's PrimeScriptTM RT reagent Kit with gDNA Eraser (Perfect Real Time). The primers for the real-time PCR were designed with the assistance of Shanghai Sangon Biological Engineering Co., Ltd (Shanghai, China), and the sequences were searched in the NCBI database. The primer pairs used for the real-time PCR are listed in Table 1. In this study, cDNA from Malus halliana plantlets served as the template and GAPDH was used as a reference for quantitative real-time PCR analysis. The quantitative data obtained from the analysis was analyzed using the 2−ΔΔCᴛ method[23]. The reaction system for the real-time PCR included TB Green™ Premix Ex Taq II, 10 μmol·L−1 upstream primers, 10 μmol·L−1 downstream primers, cDNA template, and dd H2O at specific doses. The reaction conditions involved pre-denaturation at 95 °C for 3 min, which was followed by subsequent denaturation at 95 °C for 5 s, topped off by presoaking at 56 °C for 30 s, and elongation at 72 °C for 30 s, repeated for a total of 40 cycles. Each sample was run in triplicate to ensure the accuracy and reliability of the results.
Table 1. Using the NCBI database, the CDS sequences associated with the gene family were found, and real-time quantitative PCR was performed to analyze the primer list.
Gene
namePrimer sequence (5'-3') Forward primer Reverse primer MhSnRK2.1 GAGAACAAACTATGCGTCAGT
GGAGTGGTGTCCTTGCCTCGTGTATG MhSnRK2.2 GCAACCAGTTTGAAGAGCCAGATC CATGTACGAGCCCAGACCAAGG MhSnRK2.3 TGACGGACAAGCAGACCAAAGAG GATGTTCGGGTGCCTCAGAGAC MhSnRK2.4 ACCACAGATCGCTTCGCCATC CCACCAGCCGCATACTCCATC MhSnRK2.5 AGCGTGACCAGCCTAAACAGAG ACGGCTTGACCTCCTGCTTTG MhSnRK2.6 GCCTCGTCCTCGCTGAATCG CTCCGCTGCTGTCAATGTCAATG MhSnRK2.7 AACCACAGGTCCTTGAAGCATCC GCCTCGTCCTCGCTGAATCG MhSnRK2.8 ACCCCGAAAAGAGAATAACCA
TCCCACTGCCAACTTCCTCCTTCCATC MhSnRK2.9 ATCGCTTCGCCACCCAAACATC CCACCAGCCGCATACTCCATAAC Bioinformatics analysis of MSnRK2.4
-
The protein physicochemical data were obtained from the Protparam website (
http://web.expasy.org/protparam ). DNAMAN software was applied for the amino acid residue sequence comparison of the protein. The phylogenetic tree was established using MEGAX and the nearest neighbor (NJ) method. Cis-acting sites on MhSnRK2.4 promoters were coded using PlantCARE. Subcellular localization of tobacco was carried out according to the method of Sun et al.[24]. (Supplemental Table S1).MhSNRK2.4 and MhSnRK2.4-Anti gene cloning and vector construction
-
Primer design was performed using DNAMAN software to obtain the upstream primer of MhRnRK2.4 from the gene sequence taken in the NCBI database: ATGGAAAAGTACGAGGTTGTCAAGG, the downstream primer: TCAGCTGACGCTCACTTC, and the upstream primer of the antisense vector for the gene (MhRnRK2.4-Anti): TCATCTACTACAGAAAAGAAAACCCAA, and the downstream primer: TCAGCTGACGCTCACTTCTCC. Two genes were amplified using Malus halliana cDNA as a template. The PCR products were analyzed by 1% agarose gel electrophoresis to verify band size. Subsequently, the gel was recovered, and cloned into the pMD19-T cloning construct for use with the enzyme. E. coli was next converted and the gel analyzed. The correctly sequenced MhSNRK2.4 and MhSNRK2.4-Anti plasmids were amplified, followed by EcoRI and SalI restriction digestion, and ligated into the pRI101 plasmid. The plasmid was then inserted into E. coli. The single positive colonies were identified which were used to transform the plasmid into Agrobacterium tumefaciens LBA4404 for expression transformation using the freeze and thaw method.
Agrobacterium transformation of Arabidopsis thaliana, and apple calli
-
Following the method of Hu et al.[25], transgenic MhSNRK2.4 Arabidopsis seeds were acquired by genetic transformation, treated with 75% ethanol for 5 min, 26% sodium hypochlorite for 10 min, rinsed three to five times with deionized water, then they were sown on MS medium containing 30 mg·L−1 Kanamycin (Kan) and screened for resistant plants by PCR to obtain heterozygous transgenic plants of T3 generation after three successive generations of screening[26].
The infection of the apple calli was carried out according to the method of Hu et al.[27]. To obtain stable resistant apple calli, the overexpressed calli were incubated in medium containing 250 mg·L−1 cephalosporin(cef) and 30 mg·L−1 Kan. DNA was extracted and detected by qRT-PCR.
Treatment with Fe-deficiency in transgenic Arabidopsis, and apple calli
-
First, wild-type and transgenic Arabidopsis T3 seeds were sterilized and placed on MS solid medium and MS + 30 mg·L−1 Kan medium, respectively, followed by vernalization at 4 °C for 3 d and cultivation in a light source for 10 d. Seeds were then grown on MS solid medium with sufficient (+Fe: 40 μM Fe) and deficient (−Fe: 0 μM Fe) Fe for 20 d, phenotypes were observed and physiological indices were measured.
Finally, overexpressed and WT apple calli were cultured on MS solid medium for 15 d, followed by 20 d on Fe-sufficient (+Fe) and Fe-deficient (−Fe) solid medium.
Physiological index detection measurement
-
Chlorophyll measurement followed the method of Cheng et al.[28]. Diaminobenzidine (DAB) staining involved soaking leaf samples in a 50 mM DAB solution for either 12 or 24 h, followed by decolorization in 95% ethanol until the color changed to white. Similarly, Nitrotetrazolium Blue chloride (NBT) staining for root tip or leaf samples required immersion in a 50 mM NBT solution for 4 h, then decolorization with 95% ethanol until the color changed to white. Proline was measured according to the method of Júnio et al.[29]. The activities of superoxide dismutase (SOD), peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX) were measured using a spectrophotometer and a kit from Suzhou Kaiming Biological Co., Ltd (Suzhou, China). FCR activity was determined based on the method developed by Zhao & Ling[30]. The determination of Fe2+ content was as per the method of Gu[31]. Exocrine H+ was performed using the method of Zhao & Ling[30]. Three replicates were tested for each strain. In addition, following Sharma & Kumar[32], endogenous hormones, including growth hormone (IAA) and abscisic acid (ABA), were measured.
Construction and self-activation detection of pGBKT7-MhSnRK2.4 bait expression vector
-
The pGBKT7-MhSnRK2.4 and empty pGADT7 were transferred into yeast Y2H Gold, coated on SD/-Trp/-Leu solid medium, and incubated in inverted culture at 30 °C for 2 d. Then, the yeast was cultured on SD/-Leu/-Trp/-His/-Ade solid medium and incubated at 30 °C for 3−5 d. The growth of yeast was observed as a negative control, BD + AD was included in the experiment for comparison.
MhSnRK2.4 interaction protein screening
-
In the next Y2H co-transformation, yeast two-hybrid cDNA constructs were prepared utilizing the cDNA construction kit Clone Miner™ II from Invitrogen USA. The pGADT7 library plasmid was co-transfected with MhSnRK2.4 bait plasmid into Y2H gold-sensitive cells. The cells were first cultured in SD/-Leu/-Trp solid medium and inverted at 30 °C for 3 to 5 d. Positive clones in SD/-Leu/-Trp/-His/-Ade solid medium were collected and returned to SD/-Leu/-Trp/-His/-Ade/X-α-Gals solid medium for re-screening and then rotated at 30 °C for re-screening. Clones were replated on SD/-Leu/-Trp/-His/-Ade/X-α-Gals solid medium and next cultured in an incubated dish at 30 °C for 3 to 5 d. The sequencing outcomes were employed to synthesize primers, as listed in Supplemental Table S2. These primers were incorporated into the AD vector and co-transfected along with MhSnRK2.4 into the Y2H system to verify the interaction through a rotational confirmation method.
Statistical analysis methods
-
Analysis of variance was used to test for differences between treatments, and Duncan's test was used to compare means (p < 0.05). Software (OriginLab, Hampton, MA, USA) was used for statistical analysis and Origin 8.0 software (OriginLab, Hampton, MA, USA) for manipulation of figures.
-
Using the Malus halliana transcriptome database, eight Fe-deficiency functional genes, including MhSnRK2.1, MhSnRK2.2, MhSnRK2.3, MhSnRK2.4, MhSnRK2.5, MhSnRK2.6, MhSnRK2.7, MhSnRK2.8 and MhSnRK2.9, were selected for qRT-PCR. As shown in Fig. 1, MhSnRK2.4 expression increased dramatically at 12 h, reaching a peak at 48 h, 15.24 folds greater than at 0 h. In contrast, the changes in the expression of other genes under Fe-deficiency were not as significant as those of MhSnRK2.4 in Malus halliana, indicating that MhSnRK2.4 may play a critical role in the answer to Fe- deficiency.
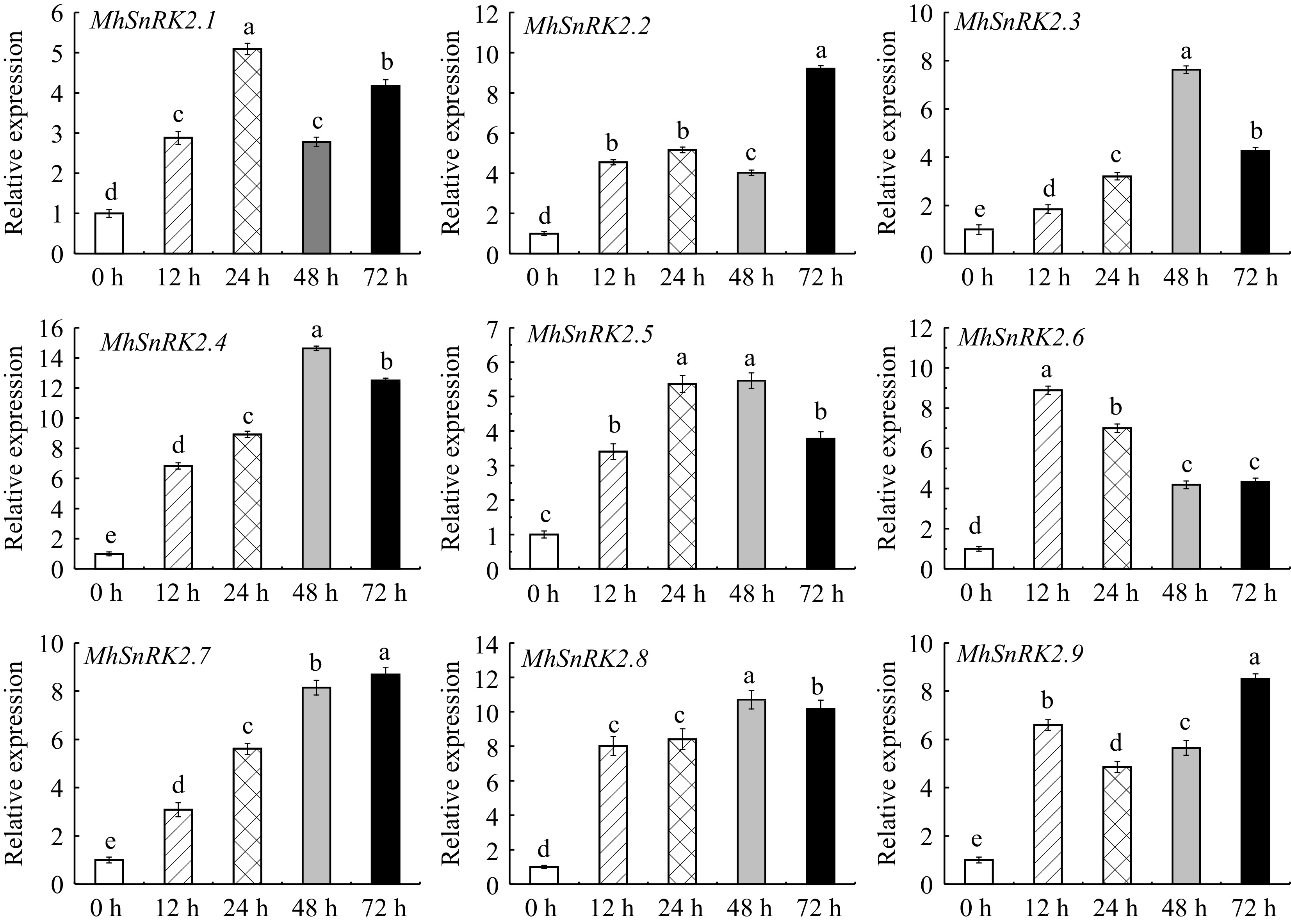
Figure 1.
Expression levels of eight Fe-deficiency functional genes on Fe-deficiency at 0, 12, 24, 48, and 72 h in Malus halliana. Different letters above the bars indicate significant differences (p < 0.05) as assessed by one-way ANOVA and the LSD test (p < 0.05).
Analysis of the MhSnRK2.4 gene
-
The MhSnRK2.4 gene was cloned from Malus halliana, with a band size of 1,059 bp (Fig. 2a). Transgenic MhSnRK2.4-Antiobtain (Anti) apple calli (Fig. 2b). The transgenic Arabidopsis, and overexpressed apple calli were characterized at the DNA level, and all matched the size of the bands of the gene (Supplemental Fig. S1). To investigate the expression site of MhSnRK2.4, a subcellularly localized carrier pART-CAM-EGFP was constructed by the homologous recombination method, and by injecting tobacco, it was observed that (Fig. 3), the green fluorescent fusion protein showed green fluorescence mainly in the cell membrane and nucleus, and a small amount of cytoplasmic distribution was also found. Therefore, it is suggested that this MhSnRK2.4 gene may be mainly expressed in the cell membrane, nucleus, and a small amount in the cytoplasm. The results presented in Table 2 revealed that the gene's open reading frame spans 1,059 base pairs, translating into 352 amino acids, possessing a molecular weight of 40.33 kD. The lipid coefficient was determined to be 62.48. Additionally, the charges on the protein were 49 positives and 55 negatives. The isoelectric point (PI) was calculated to be 6.15, indicating an alkaline nature. The protein was classified as hydrophilic with an average hydrophilicity value of −0.487. Furthermore, the instability coefficient indicated a value of 44.47, suggesting the protein's lack of stability. Comparison of the protein sequence of MhSnRK2.4 with sequences from other protein families revealed that both the N-terminal and C-terminal ends of MhSnRK2.4 are distinct from the corresponding ends in proteins from other families (Fig. 4). Furthermore, phylogenetic analysis revealed that MhSnRK2.4 and MhSnRK2.9 proteins shared high sequence identity (Fig. 5).
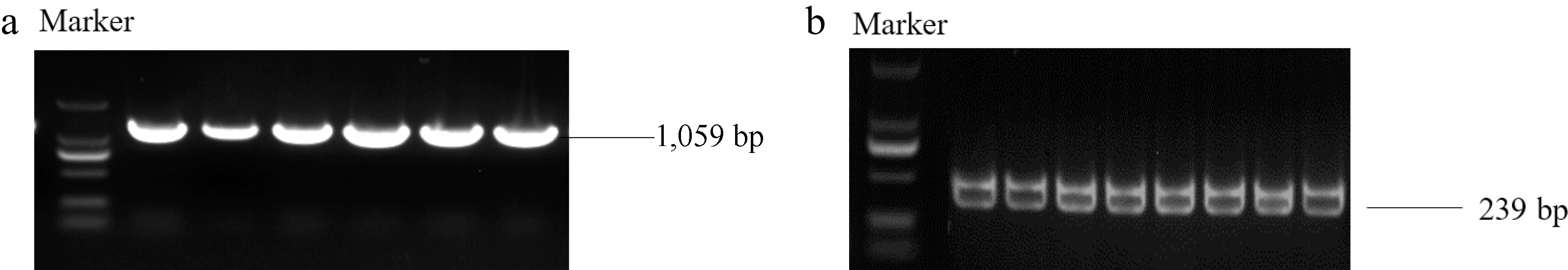
Figure 2.
PCR amplification of the MhSnRK2.4 gene and the MhSnRK2.4-Anti gene was performed using cDNA from Malus halliana as a template. (a) Electrophoresis of PCR products for cloning of MhSnRK2.4. (b) Electrophoresis images of MhSnRK2.4-Anti amplifcation.
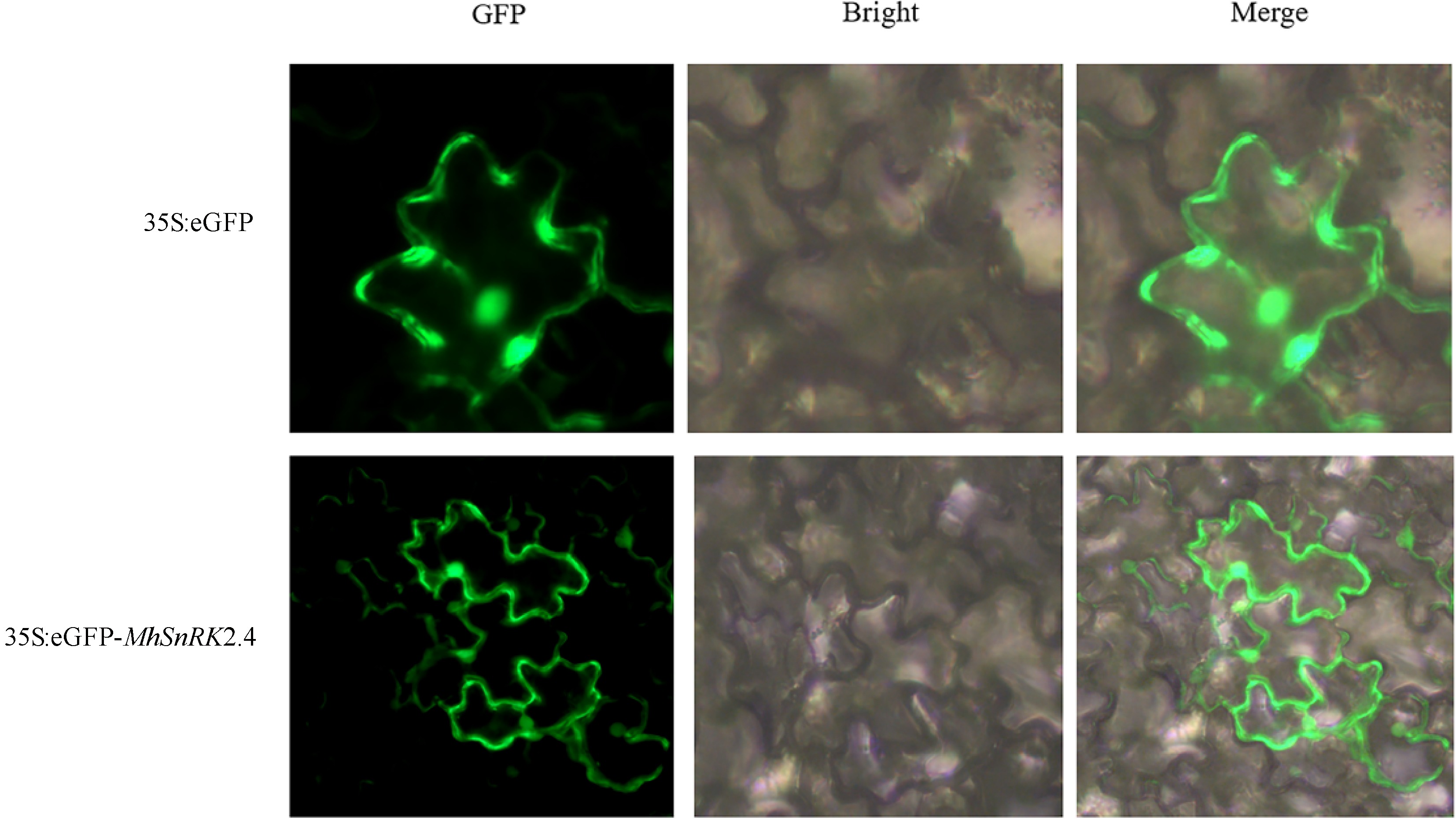
Figure 3.
Subcellular localization of MhSnRK2.4 genes. Tobacco transformed with 35S-GFP were used as the control. Co-transformed tobacco with 35S-MhSnRK2.4 -GFP, and 35S-GFP were used for subcellular localization. The nucleus (of cell) autofuorescence was also visualized by a laser scanning confocal microscopy.
Table 2. The protein sequence of MhSnRK2.4 was extracted, and the physicochemical properties of MhSnRK2.4 were analyzed using Expasy software.
Gene Amino acids Molecular weight (KD) Theoretical pI Positive residues Negative residues Aliphatic index Instability index Grand average of hydropathicity MhSnRK2.4 352 40.33 6.15 49 55 81.93 44.47 −0.487 
Figure 4.
The protein sequence of the Malus halliana MhSnRK2.4 gene was selected for analysis using DNAMAN, comparing it with protein sequences of other species.
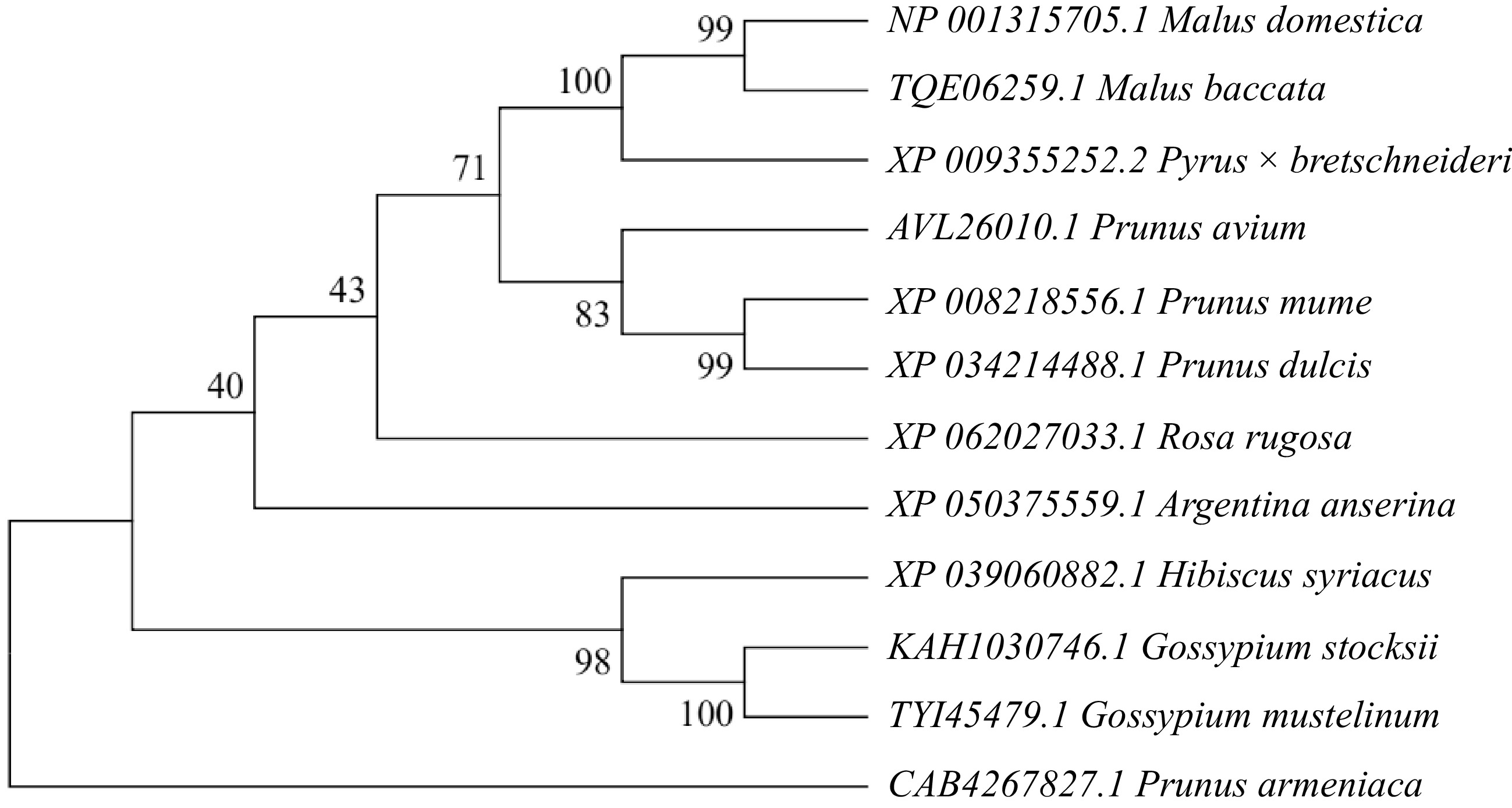
Figure 5.
Phylogenetic analysis of the Malus halliana MhSnRK2.4 gene was conducted in comparison with other species. The numbers at the branches of the evolutionary tree indicate the confidence levels of those branches; the larger the value, the higher the reliability.
Finally, the cis-elements in the 2,000 bp sequence upstream of MhSnRK2.4 were analyzed and a variety of abiotic stress-associated cis-elements were found, including those associated with low temperature and drought stress (Table 3). This implies that MhSnRK2.4 likely plays a key role in the abiotic stress tolerance response.
Table 3. Some important cis-regulatory elements regulated by the 2,000 bp genomic sequence upstream of MhSnRK2.4.
cis-Acting element Sequence Start sites (bp) End sites (bp) Function TGA-element AACGAC +68 +74 Auxin-responsive element LTR CCGAAA +1,403 +1,409 cis-Acting element involved in low-temperature responsiveness ABRE ACGTG −301 −306 cis-Acting element involved in the abscisic acid responsiveness A-box CCGTCC +1,458 +1,464 cis-Acting regulatory element G-box CACGTC +301 +307 cis-Acting regulatory element involved in light responsiveness O2-site GATGA +334 +342.5 cis-Acting regulatory element involved in zein metabolism regulation P-box CCTTTTG +1,572 +1,579 Gibberellin-responsive element GT1-motif GGTTAA −1,580 −1,586 Light responsive element MBS CAACTG +151 +157 MYB binding site involved in drought-inducibility MRE AACCTAA +1,723 +1,730 MYB binding site involved in light responsiveness Identification of transgenic Arabidopsis thaliana and overexpressed apple calli
-
qRT-PCR was used to determine expression levels of MhSnRK2.4 in transgenic Arabidopsis, and overexpressed apple calli. Results showed higher expression levels of MhSnRK2.4 in transgenic Arabidopsis and overexpressed apple calli compared to wild-type plants. Additionally, MhSnRK2.4-Anti expression was higher in overexpressed apple calli. Overexpression (OE) of MhSnRK2.4 was observed in Arabidopsis and apple calli, while OE of MhSnRK2.4-Anti was noted in apple calli (Fig. 6).
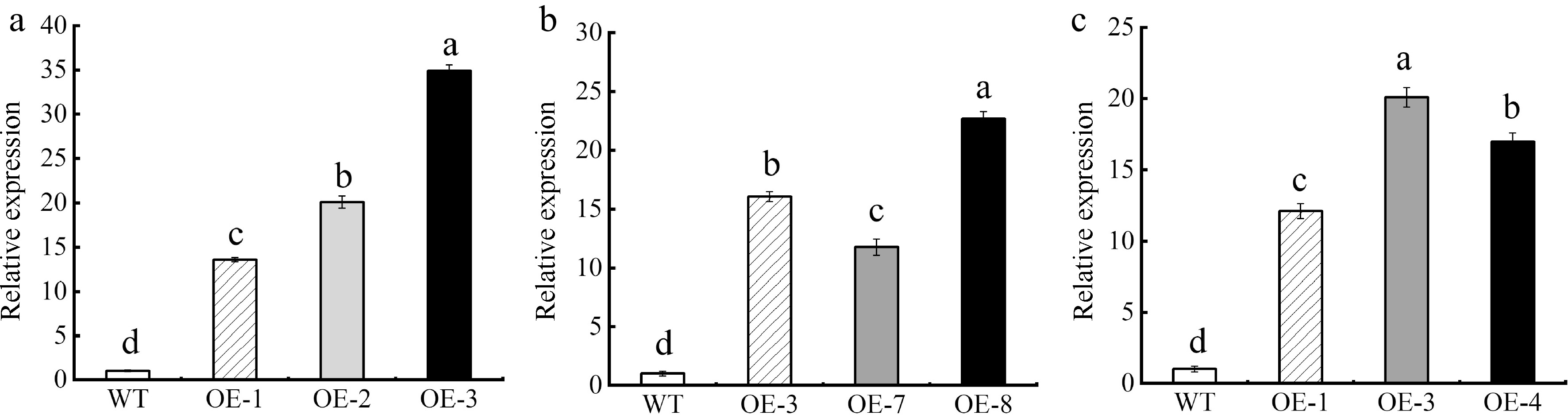
Figure 6.
Expression level of MhSnRK2.4. (a) Expression level of MhSnRK2.4 in Arabidopsis thaliana. (b) Expression level of MhSnRK2.4 in apple calli. (c) Expression level of MhSnRK2.4-Anti in apple calli. Resistance of transgenic MhSnRK2.4 Arabidopsis thaliana and apple calli to Fe-deficiency stress. Error bars denote the SD of three replicates. Different letters above the bars indicate significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test.
Resistance of transgenic MhSnRK2.4 Arabidopsis thaliana to Fe-deficiency stress
-
As depicted in Fig. 7, the transgenic Arabidopsis and the WT control Arabidopsis plants typically exhibited green leaves under Fe-supplemented conditions. In contrast, the WT Arabidopsis showed severe leaf chlorosis and stunted growth under Fe-deficient conditions. Furthermore, the chlorophyll a, chlorophyll b, chlorophyll (chlorophyll a + b), carotenoid levels (Supplemental Fig. S2), O2−, and H2O2 contents of the three transgenic lines (OE-1, OE-2, and OE-3) were not significantly different from those of WT plants from those of the WT plants under Fe-sufficient conditions. However, the transgenic Arabidopsis showed significantly increased photosynthetic pigment levels and reduced O2− and H2O2 contents under Fe-deficient conditions (Fig. 8). The genetically modified Arabidopsis plants, as depicted in Fig. 8, showed robust growth under Fe-sufficient conditions. However, under Fe-deficient conditions, the activities of POD, SOD, CAT, proline content, FCR, and Fe2+ levels were significantly higher in the transgenic lines OE-1, 2, and 3 than in the wild type. Additionally, the MDA content as shown in Fig. 8d was significantly increased relative to the WT control. These findings infer that the increased expression of MhSnRK2.4 fortified the Arabidopsis plants against the detrimental effects of Fe-deficiency.

Figure 7.
The accumulation of reactive oxygen species (ROS) and the resulting phenotypes in MhSnRK2.4 transgenic and wild-type (WT) Arabidopsis thaliana were assessed after 20 d of growth under both Fe-sufficient (+Fe) and Fe-deficient (−Fe) conditions. (a) Phenotypes. (b) NBT staining. (c) DAB staining.
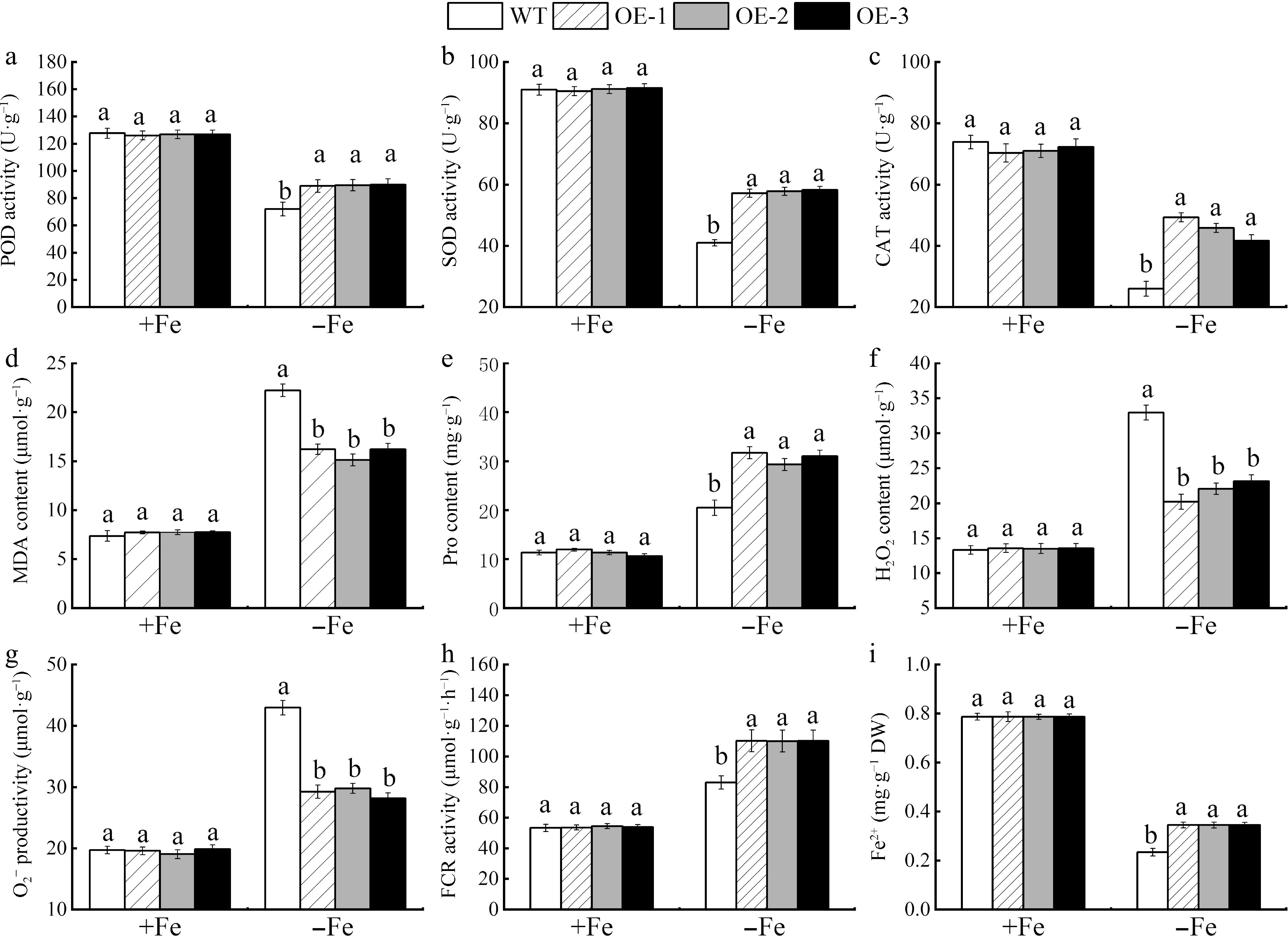
Figure 8.
The photosynthetic pigment content and enzyme activities were measured in MhSnRK2.4 transgenic and wild-type Arabidopsis thaliana plants that were grown for 20 d under either Fe-sufficient (+Fe) and Fe-deficient (−Fe) conditions. (a) POD activity. (b) SOD activity. (c) CAT activity. (d) MDA content. (e) Pro content. (f) H2O2. (g) O2−. (h) FCR activity. (i) Fe2+ activity. Different letters above the bars indicate significant differences (p < 0.05) as assessed by one-way ANOVA and the LSD test (p < 0.05).
Resistance of overexpressed MhSnRK2.4 apple calli to Fe-deficiency stress
-
As illustrated in Fig. 9, the growth of overexpressed MhSnRK2.4 apple calli did not differ significantly from that of WT apple calli under Fe-sufficient treatment, whereas under Fe-deficient treatment, overexpressed apple calli grew more vigorously than WT apple calli. The viability of overexpressed MhSnRK2.4-Anti was similar to that of overexpressed MhSnRK2.4 apple calli under Fe-sufficient conditions but was opposite to the viability of MhSnRK2.4 under Fe-deficient conditions. Bromocresol violet staining indicated that MhSnRK2.4 overexpressed apple calli compared to the WT control. MhSnRK2.4 overexpression calli were able to pump more H+ into the medium under Fe-deficient conditions (Fig. 9c, d), whereas MhSnRK2.4-Anti overexpression calli pumped less H+ into the medium under Fe-deficient conditions (Fig. 8e−h). In addition, the MhSnRK2.4 gene lines OE-1, OE-6, and OE-8 (Fig. 10) showed higher enzyme activities, including POD, SOD, CAT, APX, and FCR, compared to WT apple calli; and finally, apple calli that overexpressed the MhSnRK2.4 gene contained higher levels of Fe2+ and IAA than WT apple calli. In addition, the MhSnRK2.4-Anti gene lines OE-3, OE-7, and OE-8 showed the opposite effects to those of the MhSnRK2.4 gene line compared to WT apple calli (Supplemental Fig. S3).

Figure 9.
The phenotypes of apple calli overexpressing MhSnRK2.4 and MhSnRK2.4-Anti and the wild-type (WT) control, cultivated on Fe-sufficient or Fe-deficient media for 15 d, were evaluated through acidification analysis using a medium containing the pH indicator dye bromocresol violet. (a) MhSnRK2.4 lines. (b) MhSnRK2.4-Anti lines. (c) MhSnRK2.4 apple calli. (d) MhSnRK2.4-Anti apple calli.
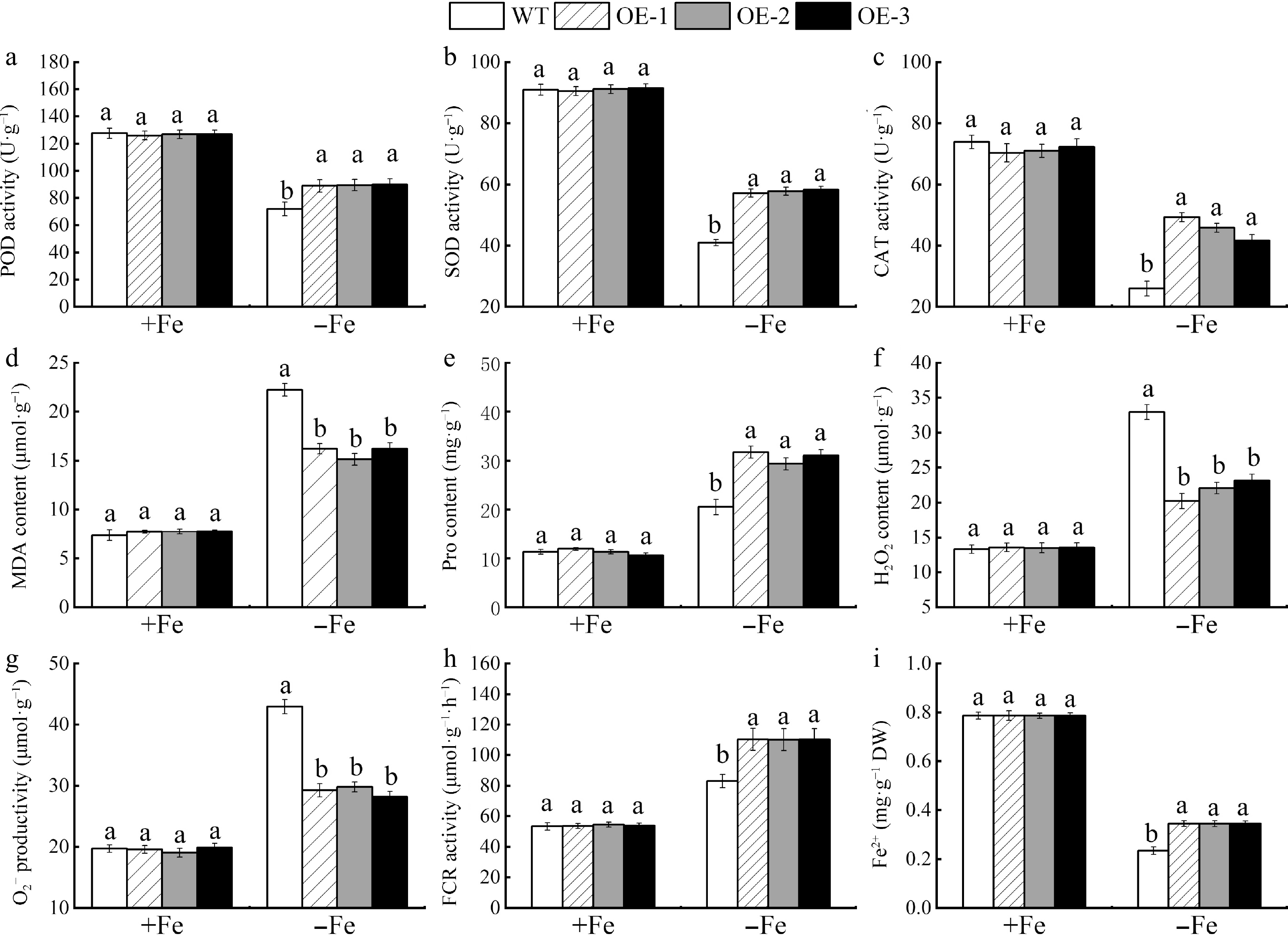
Figure 10.
In apple calli, the activities of antioxidant enzymes, the Fe-reducing capacity, and growth factors were assessed between MhSnRK2.4 overexpressing and wild-type (WT) samples under both Fe-sufficient and Fe-deficient condition. (a) POD. (b) SOD. (c) CAT. (d) MDA content. (e) Pro content. (f) H2O2 content. (g) O2−. (h) FCR. (i) Fe2+ content. Different letters above the bars indicate significant differences (p < 0.05) as assessed by one-way ANOVA and the LSD test (p < 0.05).
Screening of interacting proteins of MhSnRK2.4 by yeast two-hybrid
-
The recombinant plasmid MhSnRK2.4-pGBT7 was introduced into Y2H yeast recipient cultures and plated on SD/-Tp and SD/-Trp+x-a-gals plates, respectively. Yeast colonies grew within about 2 d. The recombinant plasmid MhSnRK2.4-pGBT7 was transferred into Y2H yeast recipient cells and plated on SD/-Tp and SD/-Trp+x-a-gals plates, respectively. As shown in Fig. 10, the recombinant plasmids MhSnRK2.4-pGBT7 and pGADT7-MhSCS, along with the positive control yeast cells, resulted in blue strains on the plates, indicating a strong interaction. In contrast, the negative control yeast cells grew on the two-deficient plates and appeared only white and did not grow on the four-deficient plates. This suggests that the MhSnRK2.4-pGBT7 and pGADT7-MhSCS proteins have a strong interaction (Fig. 11).
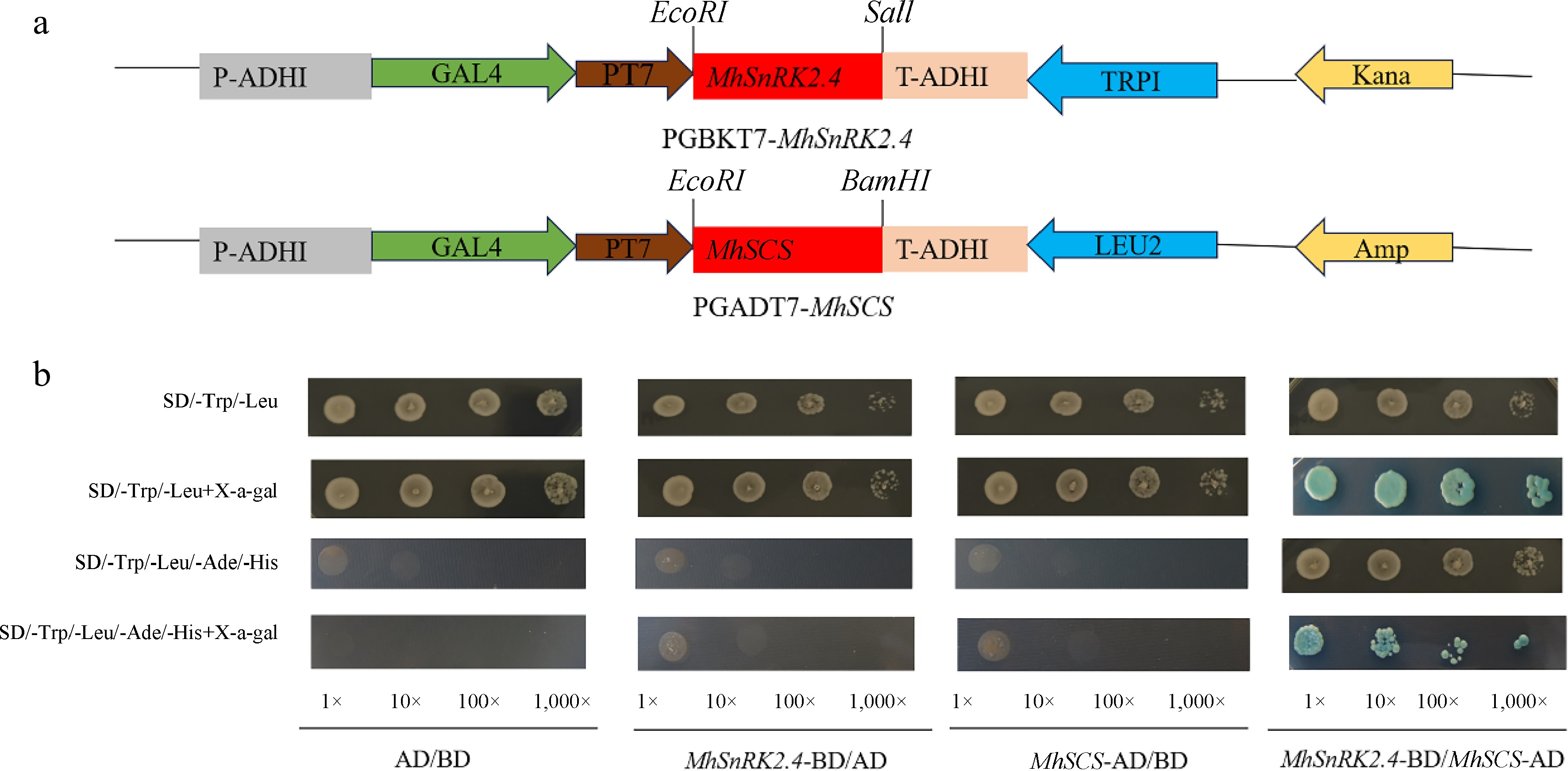
Figure 11.
Yeast two-hybrid assay showing the specific interaction of MhSnRK2.4 with MhSCS. (a) Model diagram of pGBKT7-MhSnRK2.4-BD vector construct, model diagram of pGADT7-MhSCS AD vector construct. (b) Yeast two-hybrid analysis of MhSnRK2.4 protein.
-
Iron (Fe) is the most abundant metallic element on earth and a key micronutrient necessary for human health and plant growth[33]. Its presence is intricately linked primarily to cell proliferation, DNA replication, photosynthesis, respiration, chlorophyll synthesis, and cellular oxidation[34]. While Fe is abundantly available in the earth's crust, its solubility in alkaline or calcareous soils is low, which results in limited uptake and use by plants[35]. The plant senses the adversity signals and then transmits them to different intracellular regions, causing the expression and regulation of adversity stress response genes, metabolite changes, physiological changes, and morphological changes in response to the unfavourable environment[36]. This process involves numerous phosphodiesterases and protein kinases, among which sucrose non-fermenting-related protein kinase 2 (SnRK2) is a relatively small family of plant-specific serine/threonine protein kinases[37]. Studies have shown that the expression of SnRK2s is differentially regulated by ABA, mannitol, and NaCl in different tissues and organs of Fe, some of which are up-regulated, while others are down-regulated[38]. High salt significantly induces SAPK4 expression in rice[39]. The SnRK2 member PKABA1 (protein kinase induced by abscisic acid1) first cloned from wheat (Triticum aestivum), and other wheat SnRK2s members TaSnRK2.4, TaSnRK2.7, TaSnRK2.8, and TaSnRK2.3 are up-regulated in response to different stress treatments (drought, low temperature, and salinity)[40]. Expression of maize SnRK2s genes in maize are up-regulated in response to NaCl (ZmSnRK2.3, ZmSnRK2.6, and ZmSnRK2.4), ABA (ZmSnRK2.2, ZmSnRK2.4, ZmSnRK2.5, ZmSnRK2.7, and ZmSnRK2.10) and low temperature stresses (ZmSnRK2.3, and ZmSnRK2.7), while some of them are suppressed at high temperatures[41]. The expression of four soybean (Glycine max) SnRK2 genes, SPK1, SPK2, SPK3, and SPK4[42,43], are up-regulated by hyperosmotic environments (drought and high salt). However, it is noteworthy that the expression of at least some of the SnRK2s are also regulated by photoperiod, such as SnRK2.4, and SnRK2.1[44]. This is evidence that members of the SnRK2 family have distinct functions. Based on previous transcriptome data and quantitative real-time PCR, the Fe-deficiency induced SnRK2.4 was screened and cloned to investigate its function in Fe-deficiency.
Although Fe is not part of plant chlorophyll, Fe-deficiencies may block the synthesis of pyrroles and porphyrins, which are the building blocks of chlorophyll synthesis[45]. It has been suggested that transgenic plants (Arabidopsis) have a lower chlorosis rate in their leaves than wild-type plants, which suggests that transgenic lines may have several sophisticated mechanisms to acclimate to maintain correct levels of Fe from the environment[46]. In contrast, the contents of chl a, chl b, chl a+b, carotenoids, O2− and H2O2 in wild-type Arabidopsis and transgenic Arabidopsis were analyzed, and it was found that the contents of the carotenoids in transgenic Arabidopsis were higher than in wild-type Arabidopsis under Fe-deficiency stress, whereas the contents of O2− and H2O2 were lower than in wild-type Arabidopsis. This was also verified by NBT and DAB staining, where the degree of staining represented the amount of accumulation of reactive oxygen species and hydrogen peroxide, respectively. At present, we have reported that transgenic Arabidopsis plants have an increased level of chlorophyll a and chlorophyll b suggesting that the wild-type photosynthetic system is more affected by Fe-deficiency stress and that Fe-deficiency affects chlorophyll synthesis. In turn, under Fe-deficiency stress, reactive oxygen species (ROS), mainly monolinear oxygen, are produced in chloroplasts to oxidize carotenoids and produce a variety of oxidative byproducts that can induce changes in gene activity and thus induce adaptations to environmental stress. Therefore, the experimental results can more accurately confirm that the carotenoid content of transgenic Arabidopsis was significantly higher than that of wild-type Arabidopsis, and the activity of FCR and the contents of Fe2+ of transgenic Arabidopsis were significantly higher than that of wild-type, suggesting that transgenic Arabidopsis was better adapted to Fe-deficiency stress.
Plants have evolved an ROS scavenging system not only to protect themselves from ROS toxicity but also to enable ROS to play an active role as signaling molecules during long-term development. Abiotic stress conditions can disrupt the balance between ROS production and removal in cells, resulting in increased ROS concentrations and oxidative damage to biomacromolecules, proteins, DNA, RNA, etc., which can inhibit plant growth and development and, in severe cases, lead to death[47]. POD, SOD, CAT, and APX are important enzymes in the plant antioxidant system and their activity levels can reflect the processes of plants affected by external stress[48]. In this study, it was observed that the activities of overexpressed apple calli enzymes (SOD, CAT, POD, and APX) were markedly higher than those of the respective wild-type plants under Fe-deficiency stress, which indicated that these overexpressed apple calli have a stronger antioxidant ability than the wild-type ones, which can reduce reactive oxygen radical (ROS) production. In this study, it was found that transgenic Arabidopsis stained lighter than wild-type Arabidopsis. In conclusion, through the above experiments, it was found that transgenic Arabidopsis can better maintain the stability of reactive oxygen species and protect plants from abiotic stress. Excessive accumulation of malondialdehyde caused cross-linking and polymerization of vital macromolecules such as proteins and nucleic acids, leading to changes in the structure and function of cell membranes. In this study, the MDA content of transgenic Arabidopsis was much higher than that of wild-type Arabidopsis. Proline, an osmoregulatory substance that induces the production of reactive oxygen species, was also found to be much lower in transgenic and wild-type Arabidopsis. In other words, Fe3+ was reduced to Fe2+ by the enzyme Fe chelate reductase (FCR enzyme) before it can enter various organelles and organs of the plant for transport and utilization. FCR activity and Fe2+ in Arabidopsis and apple calli were determined, and the present results showed that FCR activity and Fe2+ in transgenic Arabidopsis and overexpressed apple calli were significantly higher than those of the wild type during Fe-deficiency stress, whereas for the gene MhSnRK2.4-Anti, the results showed that FCR activity in overexpressed apple calli was significantly lower than that of the wild type. It has been shown that transgenic plants could promote Fe3+ reduction and improve Fe use by increasing FCR activity. In this way, Fe uptake in plants can be enhanced by transgenic plants.
Plant hormones play a crucial role in regulating plant responses to abiotic stress. IAA and ABA are closely related to the mechanism of Fe-deficiency in plants[49,50]. When Fe is deficient in the environment, the ABA content in plants increases and the IAA content decreases. The levels of ABA and IAA in apple calli overexpressing MhSnRK2.4 and overexpressing MhSnRK2.4-Anti were determined. The ABA content in transgenic apple calli overexpressing MhSnRK2.4 was significantly higher than in the wild-type control during Fe-deficiency stress. Conversely, the ABA content in calli overexpressing the antisense gene of MhSnRK2.4 was significantly lower than in the wild-type. Whereas the IAA content of overexpressed apple calli of transgenic MhSnRK2.4 was significantly lower than that of the wild-type control, it was determined that the IAA content of overexpressed apple calli of MhSnRK2.4-Anti was significantly higher than that of the wild type, which demonstrated that the transgenic plants acted as a positive regulator of Fe-deficiency stress in plants, whereas MhSnRK2.4-Anti acted as a negative regulator of Fe-deficiency stress. To summarize, transgenic MhSnRK2.4 improves resistance to Fe-deficiency.
-
In conclusion, the transgenic Arabidopsis thaliana MhSnRK2.4 gene and its overexpression in apple calli could cope with Fe starvation stress and enhance resistance, and its mechanism of action under Fe starvation stress was revealed in four aspects; chlorophyll synthesis, ROS, osmoregulation, and Fe reduction. Specifically, MhSnRK2.4 enhanced Fe-deficiency tolerance by increasing FCR and Fe2+ accumulation, while improving ROS scavenging efficiency, protecting membrane integrity and promoting chlorophyll synthesis. Finally, MhSnRK2.4 was shown to interact with SCS proteins. This provides a direction for further research into other functions of MhSnRK2.4 and a theoretical basis for breeding apple rootstocks with effective Fe-deficiency resistance.
-
The authors confirm contribution to the paper as follows: all authors contributed to the study's conception and design; funding acquisition: Wang X; writing original draft: Wang X; writing-review & editing: Wang X, Zhang Z, Gao Y, Dong Y; figure creation and adaptation, Wang X, Dong Y, Zhang Z, Zhao W, Zhang D, Ma X. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This work was supported by the National Natural Science Foundation of China (Project Number 31960581).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 List of primers for tobacco subcellular transformation.
- Supplemental Table S2 List of primers for yeast interactions.
- Supplemental Fig S1 Identification of genetically modified material. (a) Identification of transgenic MhSnRK2.4 Arabidopsis plants. (b) Identification of transgenic MhSnRK2.4 tobacco plants. (c) Identification of transgenic MhSnRK2.4 apple calli. (d) Identification of transgenic MhSnRK2.4-Anti apple calli.
- Supplemental Fig S2 The photosynthetic pigment content and enzyme activities were measured in MhSnRK2.4 transgenic and wild-type Arabidopsis thaliana plants that were grown for 20 days under either Fe-sufficient (+Fe) and Fe-deficient (-Fe) conditions. (a) Chla. (b)Chlb.(c) Car. (d) Chla+b.
- Supplemental Fig S3 Acidification analysis of transgenic MhSSnRK2.4-Anti apple calli on medium containing the pH indicator dye bromocresol violet. A yellow color around apple calli indicates acidification. (A). FCR (B). Fe2+. (C) ABA. (D) IAA.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang X, Zhang Z, Dong Y, Gao Y, Ma X, et al. 2024. Functional identification of protein kinase MhSnRK2.4 involved in Fe-deficiency stress in Malus halliana Koehne. Fruit Research 4: e032 doi: 10.48130/frures-0024-0026 

Functional identification of protein kinase MhSnRK2.4 involved in Fe-deficiency stress in Malus halliana Koehne
- Received: 23 March 2024
- Revised: 29 July 2024
- Accepted: 02 August 2024
- Published online: 04 September 2024
Abstract: SnRK (sucrose non-fermenting protein kinase family), a class of serine/threonine (Ser/Thr) protein kinases, is widely distributed in plants and is divided into three subfamilies: SnRK1, SnRK2, and SnRK3. The SnRK2 family, in particular, plays a significant role in osmotic stress resistance and ABA signaling pathways. However, research on the relationship between the SnRK2 gene and iron (Fe) deficiency in apples is limited. Studies have shown that the SnRK2.4 gene exhibited higher regulation of expression under Fe-deficient conditions compared to other genes. Yet, the mode of action of this gene in regulating Fe-deficiency stress remains unexplained. Therefore, the MhSnRK2.4 gene (Genbank ID:103411041) was cloned from Malus halliana in the present study. It was observed that transgenic Arabidopsis plants exhibited more robust growth and reduced chlorosis under Fe-deficiency stress when compared to the wild-type control. The overexpressed apple calli also exhibited enhanced growth Fe-deficiency stress. In addition, measurements of physiological indices showed that overexpression of the MhSnRK2.4 gene increased the tolerance of Arabidopsis thaliana to Fe-deficiency damage by promoting chlorophyll synthesis, Fe-deficiency damage by promoting chlorophyll synthesis, increasing the activity of antioxidant end enzymes, and promoting Fe reduction, and apple calli to Fe-deficiency damage by decreasing pH, increasing the activity of antioxidant end enzymes, and promoting Fe uptake. In conclusion, the overexpression of the MhSnRK2.4 gene enhanced the resistance to Fe-deficiency stress in Arabidopsis and apple calli, increasing antioxidant enzyme activity, and promoting Fe reduction.
-
Key words:
- MhSnRK2.4 gene /
- Gene cloning /
- Fe- deficiency stress /
- Yeast two-hybrid