









-
Saline-alkali stress is one of the major adverse factors affecting plant growth and yield[1], and about 10% of the world's land (more than 1.1 billion hectares) is threatened by saline-alkali[2]. Saline-alkali stress poses a severe threat to plants. It disturbs the equilibrium of water and ions within plants, hampers both photosynthesis and respiration, initiates oxidative harm, suppresses growth and development, leading to stunted plants with yellowing leaves and potentially resulting in plant death[3,4]. To adapt to growth and development in adverse conditions, plants boost cell wall firmness by controlling the accumulation of cell wall constituents like pectin and lignin, which prevents water loss and reduces the entry of excess ions.
Glycosyltransferases (GTs) constitute a large enzyme family, which catalyze the transfer of activated sugars onto diverse acceptor molecules[5,6]. They play a vital role in maintaining the metabolic balance of various substances through glycosylation reactions in plants[7,8]. The GT8 family of proteins all contain the Glycosyl-transferase 8 structural domain[9], and the GT8 gene family features a complex composition and a substantial number of members[10−12], with 41 genes belonging to the GT8 family in Arabidopsis alone and 39 family genes in rice[13]. Based on the protein phylogenetic relationships, the GT8 family genes can be categorized into four subfamilies[14], galacturonosyltransferase (GAUT), galacturonosyltransferase-like (GAUT-like, GATL), galactoside synthase (GolS), and glycogen starch synthesis initiation protein (PGSIP), of which two subfamilies, GAUT and GATL, are mainly involved in the biosynthesis of pectin and xylan in the cell wall. For example, GAUT1 in Arabidopsis is involved in pectin biosynthesis[15], mutations in AtGAUT12 drastically reduce xylan content in plants[16,17], and GUX1, GUX2, and GUX5 are all involved in the phytosynthesis of xylan[18]. MicroRNA775 modulates the inherent leaf size and decreases cell wall pectin content through targeting the galactosyltransferase gene (GATL9) in Arabidopsis[19]. In poplar, PdGATL1.1 and PdGATL1.2 might be involved in the synthesis of xylan and other cell wall polymers[20]. Pectin and xylan are required for cell wall synthesis, suggesting that GT8 family genes play an important role in plant pectin synthesis[21].
Pectin is an acidic polysaccharide rich in galacturonic acid, which is mainly found in the primary cell wall and intercellular layer, accounting for up to 40% of the dry weight of the cell wall[22]. The extracellular space, which is mainly composed of the cell wall, acts as the foremost hurdle to block the ingress of salt ions into the cell[23]; therefore, pectin can be used as a salt-resistant barrier to regulate the entry of salt ions[24]. The GATL-catalyzed formation of homogalacturonan (HG) is the most important component of the pectin main chain[25]. HG pectin can reversibly bind metal cations such as Ca2+ and Na due to the large number of carboxyl groups and the negative charge upon dissociation, which endows it with an important ion-balancing function. Under low-temperature conditions, pectin and non-cellulose components of the cell wall components tend to accumulate, of which the increase in pectin content can enhance the compressive strength of plant cells and the thickness of the cell wall, reducing the degree of cellular exposure to low-temperature hazards+[26]. Under drought conditions, plants may also be modified by pectin to produce cell wall remodeling and improve drought tolerance[27]. Salt-tolerant faba bean varieties had significantly higher pectin content in the cell walls of root tip cells than sensitive strains[28]. Most AtGATL genes are highly expressed in stem and hypocotyl tissues and are also crucial for HG biosynthesis, AtGATL1 may function as a galactosyltransferase involved in pectin synthesis[20], and AtGATLT5 can also alter the size of Rhamnogalacturonan II (RGII). Multiple OsGATL genes were also found to be induced by low temperature, drought, and ABA treatments in rice[13]. Similarly, in Eucalyptus megacephalus EgrGATL1 was induced by low temperatures to undergo cell wall remodeling to influence plant abiotic adversity resistance[29].
Malus halliana Koehne is a high quality apple rootstock native to arid saline zones. Currently, the academic research on M. halliana is insufficient and there are fewer relevant results, but in the available studies, it has been found to exhibit excellent adversity tolerance, a characteristic that makes it uniquely valuable and promising in the field of apple rootstocks[30]. In view of this, we decided to deepen our research work on M. halliana. In this experiment, we screened and cloned the gene MhGATL1 (MD02G1155400), which is associated with the pectin synthesis pathway significantly induced by saline-alkali stress, based on the previous qRT-PCR data from members of the apple GT8 family clade under saline-alkali stress and drought stress (Supplementary Fig. S1)[31]. The functions of it in Arabidopsis, tobacco, and apple callus tissues were studied, and Y2H technology was further used to screen genes that may interact with it. This research provides stronger support and guarantee for the development of the apple growing industry.
-
Gene expression analysis was performed on M. halliana seedlings. The plant materials used for genetic transformation were Arabidopsis thaliana var Columbia, Honghua Dajinyuan tobacco (Nicotiana tabacum L.), and Orin apple calli tissue.
M. halliana seedlings were first placed in MS medium for 30 d. After initial growth stabilization, the seedlings were transferred to rooting medium. After rooting, the seedlings were transplanted to Hoagland nutrient solution for 15 d to allow further development and acclimatization to the subsequent treatments. The response mechanisms of M. halliana seedlings were investigated by using 100 mM 1:1 NaCl : NaHCO3 (pH 8.0) to simulate saline-alkali stress and 15% PEG 6000 (w/v) to simulate drought stress. Seedling samples were collected at 0, 6, 12, 24, 48, and 72 h after the stress treatments for subsequent analysis. The incubation conditions were as follows: 26 °C, 80%−85% relative humidity, 6,000 Lx light intensity, and 16 h/8 h of light/dark time.
Acquisition of transgenic Arabidopsis, tobacco, and apple calli tissues and stress treatment
-
Transformation of Arabidopsis by inflorescence infestation, was carried out with specific reference to the methods of Wang et al.[32]. The detailed steps of Arabidopsis cultivation are as follows: seeds of the wild-type are first disinfected and rinsed, then plated on MS medium and incubated at 26 °C under a 16 h light/8 h dark photoperiod. After 30 d, the seedlings are transferred to 7 cm × 7 cm plastic pots filled with a substrate mixture (3 substrate : 1 vermiculite) for further growth. During the flowering stage, Agrobacterium-mediated inflorescence infiltration is carried out. Subsequently, the seeds are collected and screened on Kanamycin (Kan)-resistant medium for three generations to obtain homozygous transgenic seeds. The T3 generation of transgenic Arabidopsis was transplanted into the substrate and incubated for 30 d to fully acclimatize the growth environment. Subsequently, the plants were subjected to saline-alkali stress (100 mM 1:1 NaCl : NaHCO3), and the plants were watered with fresh water as a control treatment.
According to the method proposed by Zhang et al.[33], transgenic tobacco was obtained by selecting young leaves from tobacco tissue culture seedlings, cutting them into small pieces, placing them in the infection solution for 8 min, and suctioning off the excess bacterial liquid. The leaves were then placed on MS medium without and with antibiotics (250 mg/L Cef + 30 mg/L Kan) for cultivation. When the regenerated buds grew to 1.5 cm, they were transferred to tobacco rooting medium for growth to promote the growth and development of their roots, thus completing the entire cultivation process of tobacco transgenic seedlings. Transgenic tobacco was treated in liquid MS medium with and without the addition of 100 mM 1:1 NaCl : NaHCO3 for 3 d, respectively.
The well-developed callus tissues were prepared by placing them in the bacterial solution, incubated with shaking for 15 min, the excess bacterial solution was aspirated, and then incubated on MS medium for 24 h. It was then transferred to medium containing antibiotics and incubated at 25 °C under dark conditions until transgenic callus tissue was obtained. Orin apple calli tissues were treated in solid MS medium with and without the addition of 60 mM 1:1 NaCl : NaHCO3 for 15 d, respectively. Three replicates of each of the above treatments were carried out.
Quantitative real-time PCR
-
Total RNA was extracted from M. halliana leaf blades using an RNA extraction Evo M-MLV qRT-PCR kit (Accurate Biotechnology Co., Ltd, Changsha, China). cDNA was obtained by reverse transcribing the RNA using the Evo M-MLV Reverse Transcription Kit (Accurate Biotechnology Co., Ltd, Changsha, China). Primers were designed using the DNAMAN software after searching the apple genome database for MhGATL1 sequences (Supplementary Table S1). GAPDH served as the reference gene for quantitative real-time PCR (qRT-PCR). Each sample had three replicates. Eventually, the data were analyzed by the 2−ΔΔCᴛ method[34], and the difference was evaluated using the Duncan test of single-factor ANOVA with a significance level of p < 0.05.
Bioinformatics analysis and cloning of the GATL1 gene
-
The NCBI online database (
www.ncbi.nlm.nih.gov ) was utilized to identify the sequence information of GATL1 proteins in other species. The ProtParam website (https://web.expasy.org/protparam/ ) was utilized to analyze the physical and chemical properties of proteins. Alignment operations on the amino acid sequence of proteins were performed using DNAMAN software. A phylogenetic tree was constructed using the maximum likelihood (ML) method in MEGA-X software[35]. Using the Plantcare website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) the cis-acting elements on the MhGATL1 promoter were predicted. Analysis of the subcellular localization of the GATL1 protein was carried out using the Predict Protein website (https://predictprotein.org ).The cDNA obtained by reverse transcription was used as a template for PCR amplification of MhGATL1. Reaction steps: 94 °C for 5 min for pre-denaturation; 94 °C for 30 s for denaturation, 58 °C for 30 s for annealing, 72 °C for 90 s for extension, for 42 cycles; and 72 °C for 10 min for final extension. The PCR product was subjected to 1.25% agarose gel electrophoresis and the target band was recovered, ligated with the pMD19-T cloning vector (Bioscience Biotechnology Co., Ltd, Shanghai, China), and then transformed into E. coli (Trelief 5α), the colonies were identified by PCR, and the positive cloned colonies were sent for sequencing. The precisely sequenced MhGATL1 plasmid was extracted and ligated into the pRI101 overexpression vector by double digestion with Sma I and Kpn I to construct the pRI101-MhGATL1 fusion plasmid, transformed into E. coli, screened for positive single colonies, and then transferred into Agrobacterium verticillioides (GV3101) by the freeze-thawing method for genetic transformation.
Yeast two-hybrid (Y2H) assays
-
The yeast system was employed to evaluate the transactivation ability of the GATL1 protein. The MhGATL1 CDS were cloned and inserted into the pGBKT7 (BD) plasmid (Clontech, Shanghai, China) to obtain the BD-MhGATL1 recombinant vector. The empty BD served as the negative control. The BD-MhGATL1 plasmid was transferred into yeast strain Y2H Gold following the lithium acetate-polyethylene glycol-mediated transformation protocol. The colonies were incubated inverted on SD/-Leu/-Trp (DDO) and SD/-Trp/-Leu/-Ade/-His/X-α-Gal (QDO/X) culture substrates, respectively, for 3 d at 30 °C, and colony growth was observed and recorded for transcriptional self-activation.
Yeast two-hybrid cDNA libraries were generated with the Clone Miner™ II cDNA Library Construction Kit (Thermofisher, MA, USA). The pGADT7(AD) library plasmid was co-transfected with the BD-MhGATL1 bait plasmid into Y2H Gold receptor cells, which were subsequently coated on DDO solid medium and incubated in inverted culture at 28 °C for 3 to 5 d. Positive clones on DDO plates were then selected and transferred to QDO/X solid medium and rescreened 2−3 times. The colonies on the QDO/X plates were amplified by PCR, and positive clones with PCR products longer than 500 bp were selected and then sent to the company for sequencing, and the sequencing results were analyzed for candidate interacting proteins by comparison on NCBI Blastx. Candidate interacting proteins with correct ORFs were screened, and subsequently, specific primers were designed based on their respective sequence information (Supplementary Table S1). The full-length fragment of the coding sequence of the prey gene was amplified by PCR using the cDNA of M. halliana as a template, and the pGADT7 vector was reconstructed (with the enzymatic cleavage sites of EcoR I and BamH I). The amplified full-length fragment of the prey gene was ligated into the digested vector to reconstruct the pGADT7 fusion vector. Finally, pGADT7-Prey recombinant plasmid containing the prey gene and BD-MhGATL1 recombinant plasmid containing the bait gene were co-transfected into Y2H Gold yeast receptor cells.
Determination of related indexes
-
The chlorophyll content was detected by the acetone method[36]. The relative electrical conductivity (REC) was measured using a DDS-11A conductivity meter. MDA content was determined by the thiobarbituric acid method[37]. Proline content was evaluated by the acid ninhydrin method, following the method of Ferreira Júnior[38]. The determination of superoxide dismutase (SOD) activity was carried out through the photochemical reduction approach with the application of nitroblue tetrazolium[31]. Catalase (CAT) activity was measured via ultraviolet absorption. One unit of enzyme activity was defined as a 0.1 OD240 change within 1 min[31]. Peroxidase (POD) activity was determined with the guaiacol method by detecting the absorbance at OD470[31]. The levels of Na+ and K+ ions were detected using the flame spectrophotometer method[39]. Nitrotetrazolium Blue chloride (NBT) and Diaminobenzidine (DAB) staining was carried out according to the method of Wang et al.[40]. Determination of pectin content was carried out with a Solarbio Pectin Content Kit Item No. BC1400 (Solarbio Science and Technology Co., Ltd. Beijing, China). Three replicates were performed per line.
Statistical analyses
-
In this study, all data were obtained from three independent experiments, with each experiment having three technical replicates. Treatment effects were tested by analysis of variance, and means were compared using Duncan's test (p < 0.05). Significance of differences was analyzed using SPSS software (IBM, Armonk, NY, USA), and plotted using Origin 8.0 (OriginLab, Hampton, MA, USA).
-
To clarify the transcript levels of MhGATL1 gene (LOC103401949) under different abiotic stress times (Supplementary Fig. S1)[31], qRT-PCR was performed and it was found that the expression levels of MhGATL1 under both saline-alkali and drought stresses, compared with the control group (0 h), both showed an increasing trend and reached a peak at 48 h, which were 69.15 times and 13.7 times of 0 h, respectively.
Identification of the MhGAT1 gene
-
The cDNA of M. halliana was used as a template, and a 1071 bp band was amplified by PCR (Supplementary Fig. S2a), sequence analysis showed that the band was MhGATL1, which was homologously recombinantly ligated with the plant expression vector pRI101 to obtain the fusion plasmid pRI101-MhGATL1 (Supplementary Fig. S2b).
As shown in Supplementary Table S2, MhGATL1 consisted of 357 amino acids, had a relative molecular mass of 39.75 kDa, and a protein isoelectric point of 6.30, thus being classified as an acidic protein. In addition, MhGATL1 has a positive residue number of 28 and a negative residue number of 31, an instability coefficient of 42.30, which makes it an unstable protein; a fat coefficient of 90.45, which indicates good lipid solubility; and an average hydrophilicity of 0.014, which indicates a hydrophobic protein. This indicates that the protein encoded by MhGATL1 is an unstable acidic hydrophobic protein.
Protein sequence analysis of the MhGATL1 gene
-
The hydrophobicity of GATL1 proteins was analyzed using the Protscale online website, and the results showed that the polypeptide chain encoded by the GATL1 gene showed the strongest hydrophilicity at position 55, taking the value of −2.100, and the strongest hydrophobicity at position 8, taking the value of 3.111. The polypeptide chain of the amino acid encoded by GATL1 showed hydrophilicity (Supplementary Fig. S3a). The transmembrane region of GATL1 protein was predicted using TMHMM (Supplementary Fig. S3b), and no transmembrane sequence existed for MhGATL1, which was hypothesized to be a possible secreted protein.
The GATL1 protein sequences of other species were obtained from the database, and the phylogenetic tree was constructed using the maximum likelihood (ML) method in MEGA-X software. The analysis results showed that MhGATL1 is highly evolutionarily related to Pyrus × bretschneideri, and the two are clustered together as a subfamily, whereas MhGATL1 is relatively distantly related to other species compared to them (Supplementary Fig. S3c) . The results of multiple sequence comparison showed that the MhGATL1 protein sequence differed from those of other species to a certain extent at the N-terminal end, and the C-terminal end was relatively conserved (Supplementary Fig. S4).
Analysis of cis-acting elements of the MhGATL1 promoter
-
Analysis of the cis-acting elements within the 2,000 bp upstream sequence of the MhGATL1 promoter showed that there are multiple hormones as well as adversity-related regulatory elements in the MhGATL1 promoter region as shown in Supplementary Table S3. It is hypothesized that the MhGATL1 gene is regulated by environmental signals including abscisic acid and gibberellin, and that it possesses light-responsive, defense, and stress-responsive regulatory elements as well as being involved in the induction of low temperature, MeJA-responsive regulatory loci.
Screening and identification of transgenic plants
-
To validate the successful acquisition of MhGATL1 transgenic material, it was characterized at both the transcriptional and DNA levels. DNA from transgenic and WT material was extracted as a template, PCR was conducted to perform the characterization at the DNA level. The results are presented in Supplementary Fig. S5a. When 18S was employed as a primer for Arabidopsis and apple calli, and NtActin was used as a primer for tobacco, bands were visualized within the PCR product fragments of both the WT and transgenic materials. This signified that the DNA was extracted correctly and could be utilized for subsequent verification. PCR with the PRI-MhGATL1 primer generated distinct PCR bands, whereas WT showed no band, suggesting that the PRI-MhGATL1 vector was successfully constructed. At the RNA level, qRT-PCR was used to identify whether the transgenic material was successfully obtained or not. As shown in Supplementary Fig. S5b, MhGATL1 exhibited high transcriptional expression in the transgenic materials, which demonstrated that transgenic tissues of Arabidopsis, tobacco, and apple calli were successfully acquired.
MhGATL1 overexpression increases pectin content in Arabidopsis
-
To further explore the function of MhGATL1 in the process of plant adversity tolerance, the research was started with gene overexpression in Arabidopsis. Changes in pectin content of wild type (WT), and MhGATL1 (OE-1, -4, -6) overexpressing Arabidopsis lines were determined under normal and saline-alkali stress conditions. The results showed (Fig. 1) that the pectin content was increased in the transgenic lines compared with the WT under normal conditions, which indicated that the expression of the MhGATL1 gene could promote the synthesis of pectin. Under saline-alkali stress, the pectin content of MhGATL1-OE Arabidopsis lines significantly accumulated in roots and leaves, and the pectin content accumulated more in leaves.
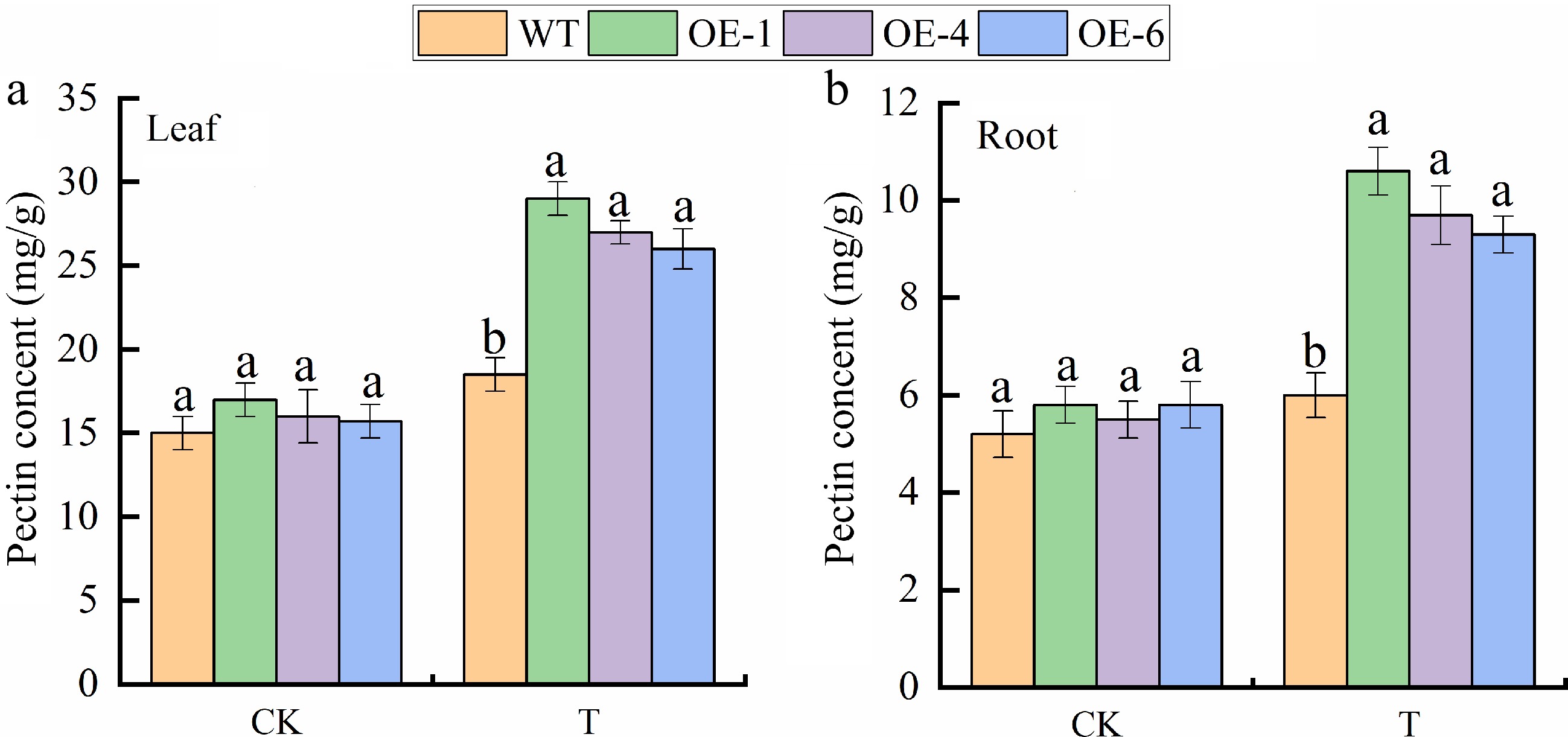
Figure 1.
Pectin content in (a) leaves, and (b) roots of MhGATL1-OE and wild-type (WT) Arabidopsis under normal conditions (CK), and saline-alkali stress (T). Error bars denote the SD of three replicates. Different letters above the bars indicated significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test.
MhGATL1 improves resistance in Arabidopsis under saline-alkali stress
-
To further investigate the role of MhGATL1 in regulating saline-alkali, WT and MhGATL1-OE Arabidopsis were subjected to saline-alkali treatment. As shown in Fig. 2, under saline-alkali stress, MhGATL1-OE plants grew better (Fig. 2a) and had longer root length and more lateral roots (Fig. 2b, c), and NBT and DAB staining of Arabidopsis leaves revealed lighter staining in MhGATL1-OE Arabidopsis leaves under stress (Fig. 2d, e), suggesting that overexpression of MhGATL1 scavenges excess ROS accumulated by saline-alkali stress. As shown in Fig. 3, the content of Relative Conductivity (REC) and MDA in MhGATL1-OE were lower than that of WT and the content material of Pro was greater than that of WT under saline-alkali stress; the antioxidant enzyme of MhGATL1-OE Arabidopsis was greater than that of the control, among which the SOD activities in the transgenic Arabidopsis OE-1, OE-4, and OE-6 had elevated SOD activities by 64.7%, 61.4%, and 46.6%, POD activities by 35.9%, 39.1%, and 28.1%, and CAT activities by 92.1%, 82.8%, and 77.1%, respectively, compared with the WT. Under normal conditions (CK), the changes in Na+, K+ content, and Na+/K+ in the MhGATL1-OE and WT Arabidopsis root mesophyll were not significant. The Na+ content in the WT and MhGATL1 roots exhibited an upward trend, whilst the K+ content material confirmed a downward trend, ensuing in an improved Na+/K+ ratio. However, the Na+/K+ ratio in the roots of MhGATL1-OE Arabidopsis were extensively decreased compared to that of the WT. This shows that the overexpression of MhGATL1 helps keep the Na+/K+ ratio, thereby enhancing the saline-alkali tolerance of Arabidopsis. According to these findings, we proposed a hypothesis that the heterologous overexpression of MhGATL1 augments the resistance of Arabidopsis to saline-alkali stress.
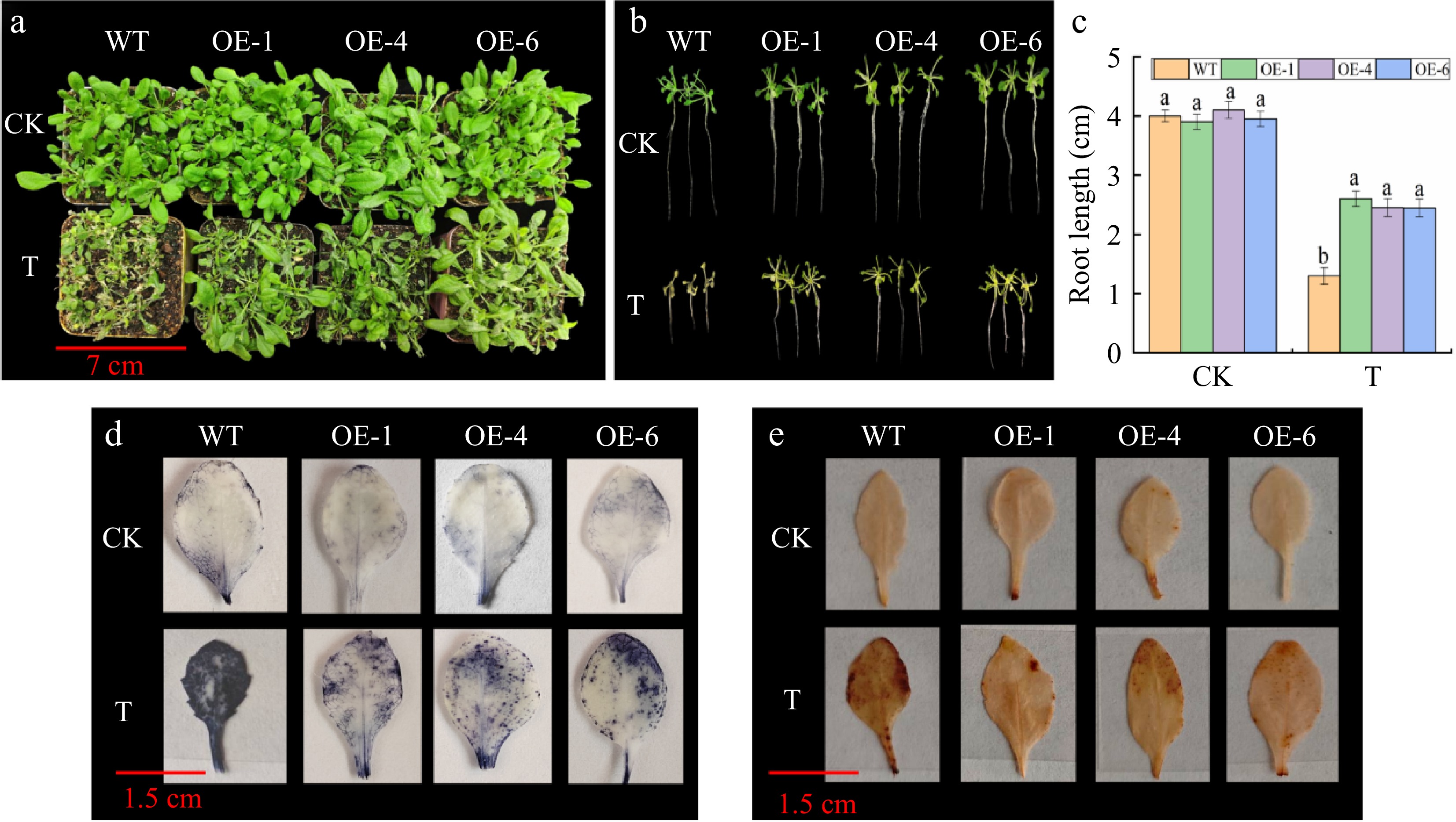
Figure 2.
The phenotype, root length, NBT, and DAB staining of MhGATL1-OE and wild-type (WT) Arabidopsis under normal conditions (CK), and saline-alkali stress (T). (a) Phenotype. (b) Phenotypes of root length. (c) Root length. Different letters above the bars indicated significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test. (d) NBT staining. (e) DAB staining.
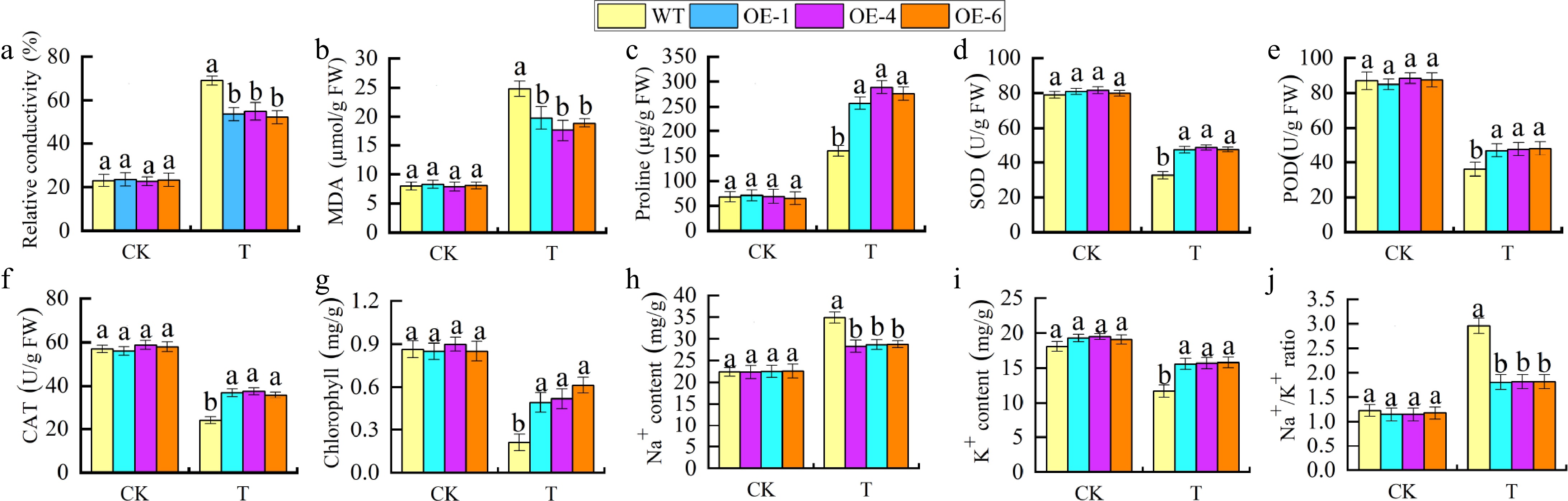
Figure 3.
Physiological indicators of saline-alkali stress in MhGATL1-OE Arabidopsis. (a) Relative conductivity. (b) MDA content. (c) Pro content. (d) SOD activity. (e) POD activity. (f) CAT activity. (g) Chlorophyll content. (h) Na+ content in roots. (i) K+ content in roots. (j) Na+/K+ ratio. Different letters above the bars indicated significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test.
To elucidate the molecular mechanism of MhGATL1 in saline-alkali response, the expression of saline-responsive representative genes and key genes for pectin synthesis at the transcriptional level were analyzed by qRT-PCR (Fig. 4). Salinity-responsive representative genes included RD29A, NCED3, NHX1, KIN1, and P5CS, and all salinity-responsive representative genes were up-regulated in MhGATL1 transgenic lines under saline-alkali conditions compared with the WT. The expression stages of key genes for pectin synthesis (AtMUM4, AtGATL5, AtQUA2, and AtPME1) had been additionally accelerated in Arabidopsis strains overexpressing MhGATL1 under saline-alkali stress. The outcomes indicated that overexpression of the MhGATL1 gene provided stronger tolerance by way of inducing the expression of saline-alkaline stress-related consultant genes, and genes associated to the pectin synthesis pathway.
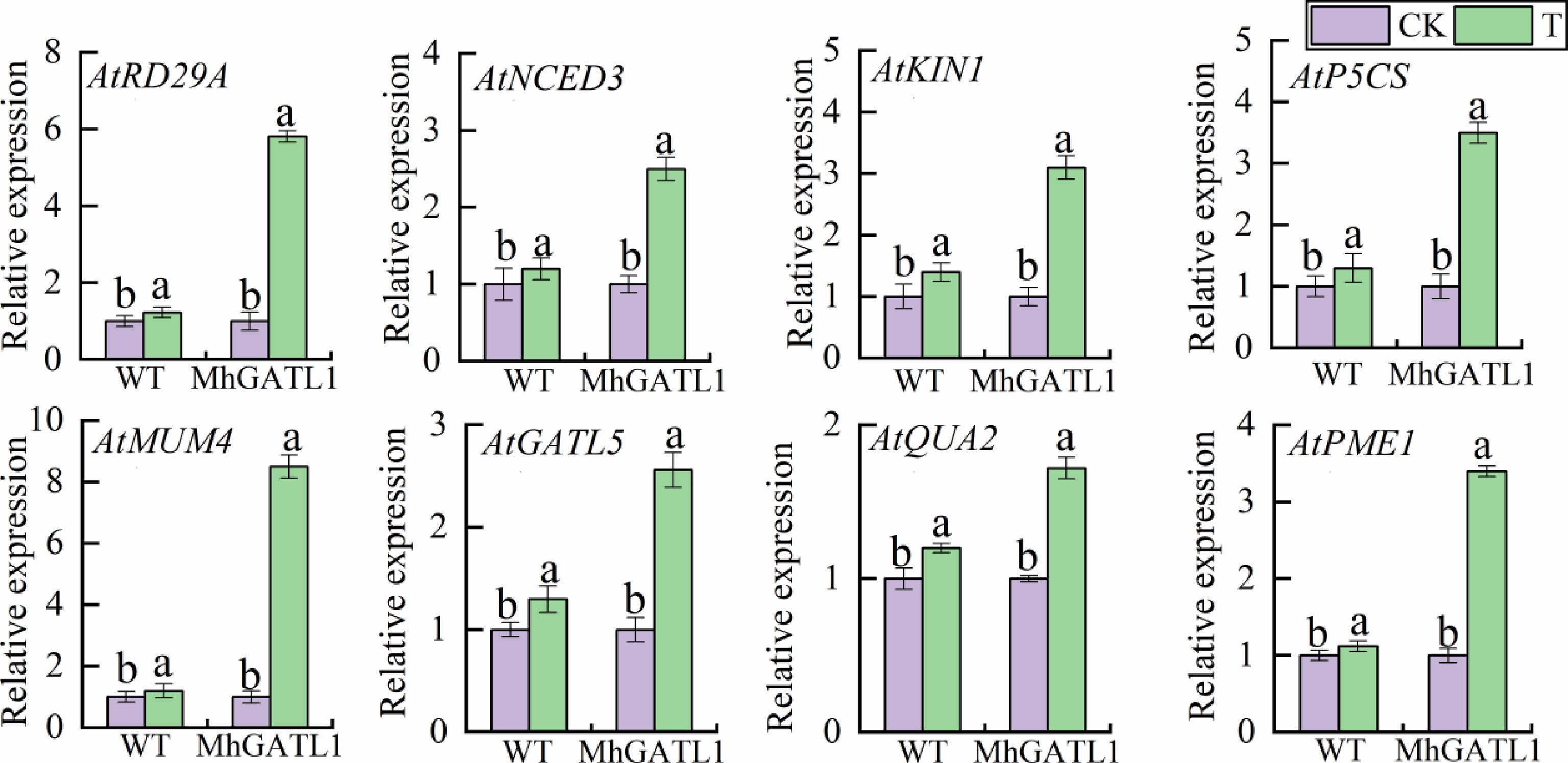
Figure 4.
Expression levels of MhGATL1-OE and WT Arabidopsis pectin synthesis pathway and saline-alkali stress response genes under normal conditions (CK), and saline-alkali stress (T). The expression levels of MhGATL1-OE and WT Arabidopsis under normal conditions were set to '1'. Different letters above the bars indicated significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test.
MhGATL1 improves resistance in tobacco under saline-alkali stress
-
As proven in Fig. 5a, each MhGATL1-OE (OE-1, -5, -6) and WT tobacco grew generally below ordinary conditions, and after saline-alkali treatment, WT showed obvious wilting and loss of green color, whereas the transgenic tobacco wilted to a lesser extent. DAB staining of tobacco leaves revealed (Fig. 5b) that there was no significant difference between the staining results of MhGATL1-OE and WT tobacco leaves under normal conditions; whereas, after saline-alkali stress, WT showed a dark brown color, and MhGATL1-OE was lighter in color, which indicated that MhGATL1-OE scavenged ROS accumulated by saline-alkali stress. Based on the results of DAB staining, physiological indexes of tobacco leaves were determined (Fig. 5c–h), and the chlorophyll contents in transgenic lines OE-1, OE-5, and OE-6 were increased by 29.75%, 32.78%, and 30.28% under stress. Meanwhile, the contents of REC and MDA in transgenic tobacco decreased significantly after saline-alkali treatment, while the content of Pro in leaves increased significantly. As indicated with the aid of antioxidant enzyme recreation assay, SOD, POD, and CAT things to do in MhGATL1-OE tobacco have been greater than these in WT manage below saline-alkali stress. Based on these results, we hypothesized that MhGATL1 heterologous overexpression improved the resistance of tobacco to saline-alkali stress.
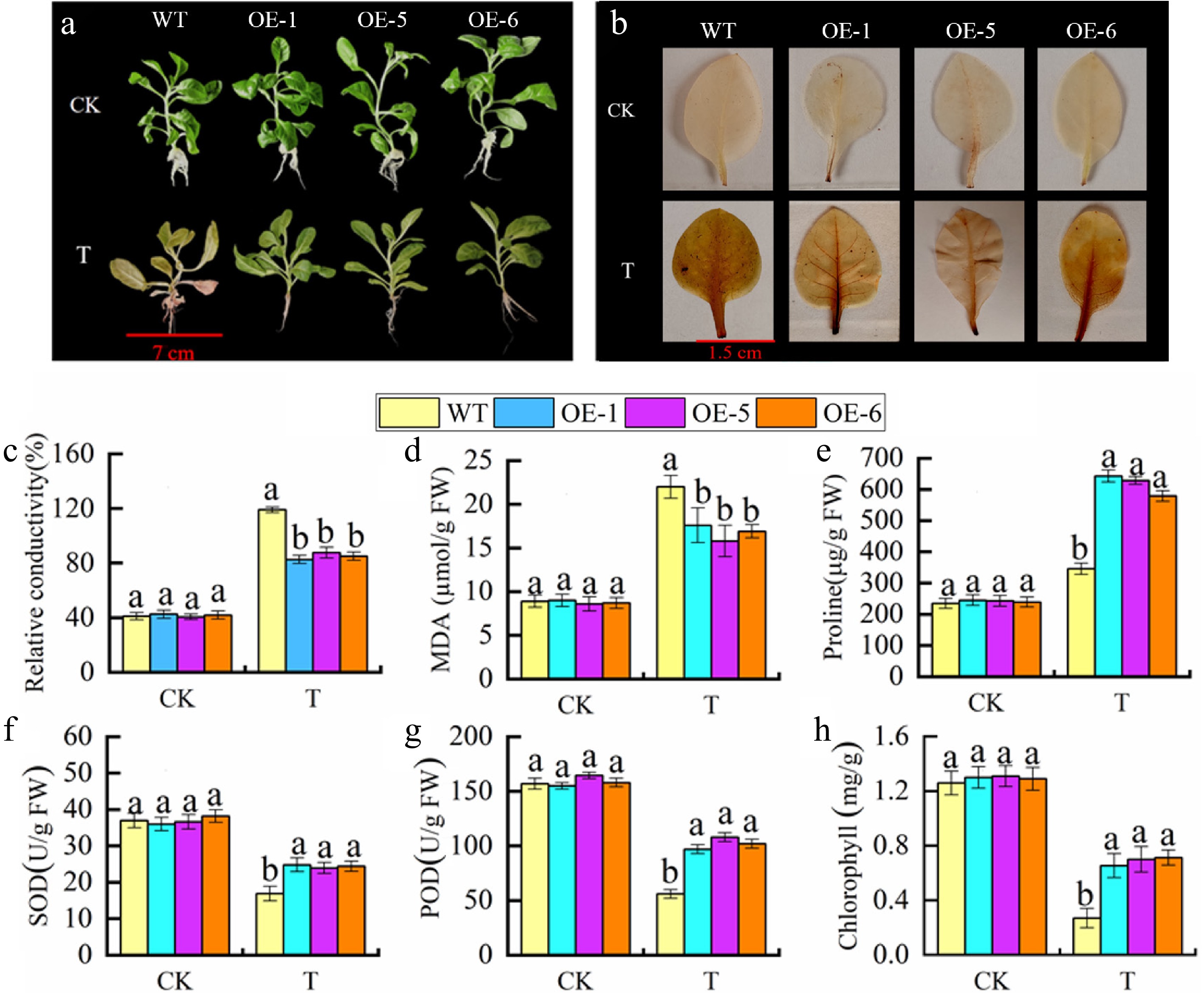
Figure 5.
Overexpression of MhGATL1 improves saline-alkali resistance in tobacco. (a) Phenotypes. (b) DAB staining. (c) Relative conductivity. (d) MDA content. (e) Pro content. (f) SOD activity. (g) POD activity. (h) Chlorophyll content. Different letters above the bars indicated significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test.
MhGATL1 improves resistance in apple calli under saline-alkali stress
-
As shown in Fig. 6a, the growth status of MhGATL1-OE and WT apple calli exhibited no significant disparity under normal circumstances. Nevertheless, the increase of each MhGATL1-OE and WT apple calli used to be impeded below stress conditions. As proven in Fig. 6b–g, under saline-alkali conditions, the overexpressed calli tissues introduced a greater clean weight than the WT. Additionally, the MDA content material inside the MhGATL1-OE calli tissues used to be conspicuously decreased; the pro content material was once raised; and the things to do of the SOD, POD, and CAT enzymes had been extensively enhanced. Such effects endorse that MhGATL1-OE is capable of positively modifying the tolerance of apple calli tissues to stress.
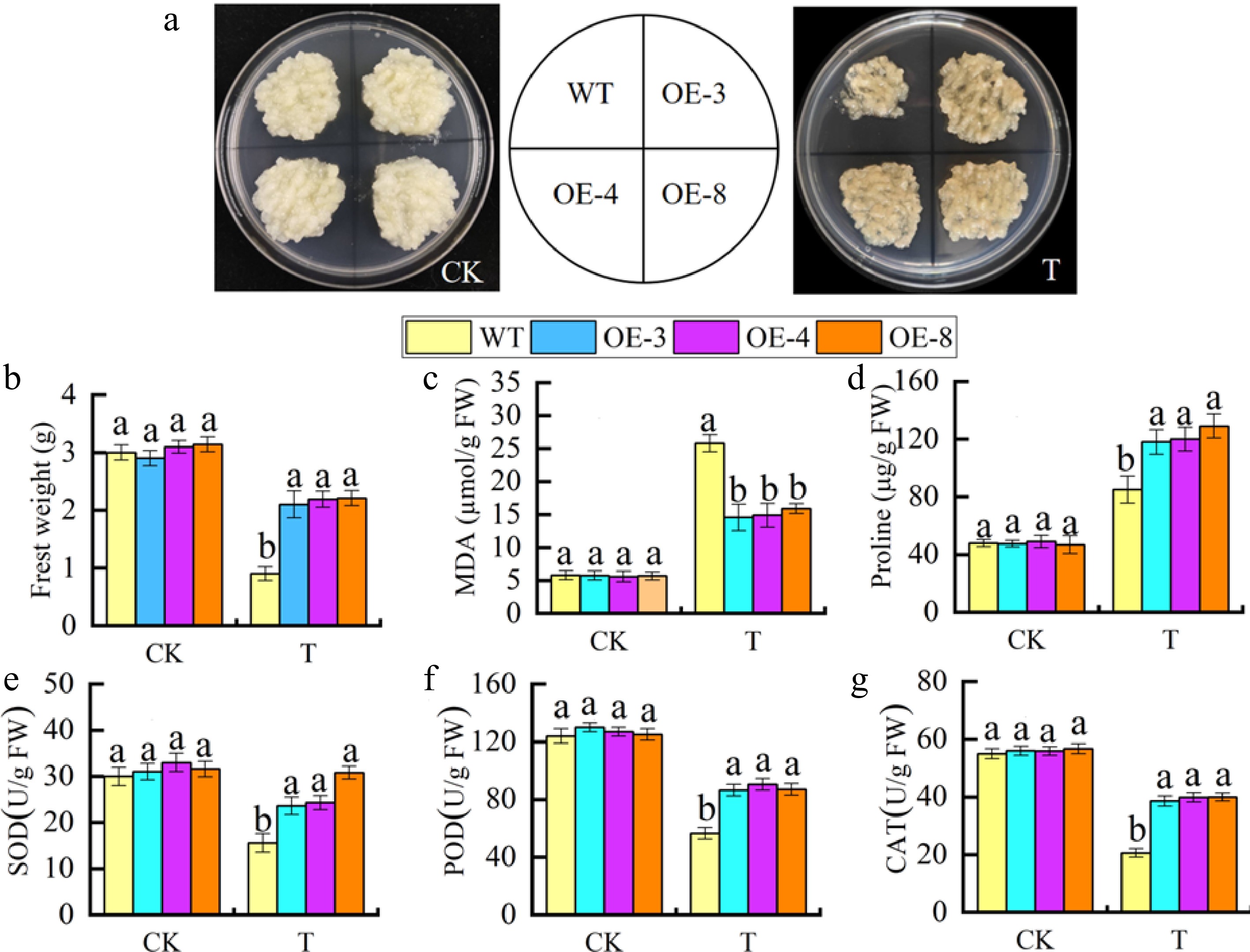
Figure 6.
Overexpression of MhGATL1 improves saline-alkali resistance in apple calli. (a) The phenotypes of MhGATL1-OE and WT apple calli under normal conditions (CK), and saline-alkali stress (T). (b) Fresh weight. (c) MDA content. (d) Pro content. (e) SOD activity. (f) POD activity. (g) CAT activity. Different letters above the bars indicated significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test.
Yeast two-hybrid screening for MhGATL1-interacting proteins
-
Before screening the library, the constructed BD-MhGATL1 decoy vector needs to be subjected to self-activation assay. As illustrated in Fig. 7, the BD-MhGATL1 decoy recombinant plasmid and the prey empty vector were transfected into Y2H Gold. Then, the transfected products were placed onto DDO and QDO/X media separately. All combinations were able to grow normally on DDO medium, suggesting that the recombinant bait plasmids had been successfully introduced into the host bacteria without exerting toxicity on them. In contrast, none of the samples could grow in the QDO/X medium and did not display a blue color. This implies that the BD-MhGATL1 plasmid was once correctly transfected into the yeast lines and did no longer possess self-activating activity, enabling its direct utility in the subsequent screening (see Fig. 7, the first two rows).

Figure 7.
Yeast two-hybrid screening for MhGATL1-interacting proteins. DDO and QDO/X represent SD/-Leu/-Trp and SD/-Leu/-Trp/-His/-Ade/X-α-Gal medium, respectively.
The BD-MhGATL1 bait plasmid and the built Y2H library plasmid have been collectively brought into Y2H Gold yeast receptor cells. Subsequently, the converted merchandise have been unfolded on DDO plates and cultured for a duration of 2–3 d. The preliminary screening yielded 48 clones. Subsequently, these effective clones from the DDO screening plate had been transferred to the QDO/X screening plate, and the screening way of life used to be repeated for three generations. Eventually, 28 blue colonies have been received (Supplementary Fig. S6a). PCR amplification was once carried out on the 28 blue colonies the use of the primer AD-F/R. The PCR amplification outcomes are introduced in Supplementary Fig. S6b. Subsequently, the legitimate clones with electrophoretic bands longer than 500 bp have been chosen for DNA sequencing. The sequencing consequences have been analyzed via Blastx comparison, and initially, seven candidate proteins that may have interaction with MhGATL1 had been received (Supplementary Table S4).
To verify whether MhGATL1 interacts with the candidate proteins, the CDS sequences of MhDIN10, MhOST1, MhUGE5, MhbZIP23, MhSOS2, MhCHI, and MhABI3 were cloned. The cloned candidate genes were combined with AD vectors to form prey proteins. Subsequently, the prey proteins and BD-MhGATL1 have been mutually delivered into the yeast sensory kingdom for mutual verification. As depicted in Fig. 7, all mixtures grew except abnormality on DDO medium, suggesting that each the bait protein and the prey protein have been correctly developed and efficiently changed into the Y2H Gold sensory state. Blue colonies grew on QDO/X medium BD-MhGATL1+AD-MhbZIP23, BD-MhGATL1+AD-SOS2, BD-Mh GATL1+AD-MhUGE5, BD-MhGATL1+AD-MhABI3, and BD-MhGATL1+AD-MhCHI, and the blue colonies reduce with growing dilution, suggesting that MhGATL1 interactswith MhbZIP23, MhSOS2, MhUGE5, MhABI3, and MhCHI proteins, but not with MhOST1 and MhDIN10.
Interacting proteins of MhGATL1 are induced by saline-alkali
-
To study the role of the candidate interacting genes of apple MhGATL1 in the signaling pathway of saline-alkali stress, the transcript levels of the candidate interacting genes were detected by qRT-PCR under saline-alkali stress. The results, as shown in Fig. 8, showed that the expression levels of the candidate genes under saline-alkali stress showed a bimodal peak, except for MhUGE5 and MhCHI, which showed an increasing and then decreasing trend, which was similar to that of MhGATL1 by saline-alkali induced expression trend. Among them, after 24 h of saline-alkali stress treatment, MhbZIP23 had the highest expression level, which was 11.2-fold that of the control (0 h). The above results indicated that the candidate interactions of MhGATL1 encoding genes were induced by saline-alkali, and MhbZIP23 was the most significant in response to saline-alkali.

Figure 8.
Expression analysis of candidate interacting genes of MhGATL1 under saline-alkali treatment. Different letters above the bars indicated significant differences (p < 0.05) as assessed by one-way ANOVA and the least significant difference (LSD) test.
-
The cell wall, as a barrier for plant cells, plays an important role in maintaining cell expansion and integrity[23]. GATL genes play an important role in cell wall biosynthesis, especially that of pectin and xylan, which constitute the basic constituent materials of the cell wall. Pectin, a polysaccharide mainly composed of galacturonic acid, is vital for controlling cell wall porosity, intercellular adhesion, cell swelling, and pathogen resistance[41,42]. The relationship between pectin content and plant stress tolerance has been a subject of interest. For instance, pectin accumulation is associated with root cell elongation under stress[43]. In rice[13] and cotton[14], certain GATL genes are induced by various stresses and hormones, indicating their potential role in abiotic stress response. However, the response of GATL1 to stress in apple remains unreported. In this study, we conducted bioinformatic analysis and genetic transformation of MhGATL1 to elucidate its molecular mechanism in response to saline-alkali stress.
In this experiment, we cloned MhGATL1 from M. halliana and conducted bioinformatics analysis on it. Moreover, through genetic transformation, we obtained Arabidopsis and tobacco with heterologous overexpression of the MhGATL1 gene, as well as apple callus with homologous overexpression of this gene. Several investigations have indicated that the correlation between the expression of the GATL gene and the accumulation of pectin has an impact on the plant's reaction to stress conditions[44,45]. In the current experiment, it was observed that the overexpression of MhGATL1 enhanced the accumulation of pectin in the roots and leaves of Arabidopsis under saline-alkali stress conditions. It is hypothesized that the increased pectin accumulation triggers certain physiological and biochemical alterations within the plants, thereby improving their saline-alkali tolerance, which is consistent with the research on Eucalyptus grandis by Cheng et al.[29]. Subsequent size of the physiological and biochemical parameters of the MhGATL1 transgenic substances prove that both transgenic Arabidopsis and tobacco strains manifested most advantageous boom and elongation of principal roots when subjected to stress. This suggests that the transgenic flora probably possess an augmented capability to soak up water and nutrients, conferring a gain for withstanding saline-alkali stress conditions[46]. Transgenic apple callus tissue has a higher fresh weight than WT. The accumulation of ROS (O2−, H2O2) in leaves could be visualized by DAB and NBT staining, and the staining results revealed that saline-alkali stress led to the accumulation of ROS in the plants[47], while the overexpression of MhGATL1 could remove part of the ROS accumulation. Under stress conditions, the MhGATL1 transgenic lines exhibited lower levels of REC and MDA, suggesting that the MhGATL1-OE transgenic plants suffered relatively less damage. The content material of Pro, an osmoregulatory substance, is a indispensable indicator of plant stress tolerance[48]. In this research, the Pro contents in the MhGATL1 transgenic substances have been greater than these in the WT, indicating that the transgenic substances may want to more efficiently improve the content material of osmoregulators to relieve saline-alkali stress. Under adverse conditions, plants promptly boost the activity of antioxidant enzymes like POD, SOD, and CAT, safeguarding the organism from harm[49]. The enzyme activities within the MhGATL1 transgenic material were substantially greater than those of the WT, thus enhancing the plant's tolerance to saline-alkali stress. The balance of Na+/K+ is significant in a plant's resistance to saline-alkali stress. Under normal circumstances[50], the roots of MhGATL1-OE exhibited a high K+ content and a low Na+ content. Nonetheless, in the presence of saline-alkali stress, the Na+ content within the roots of both WT and MhGATL1 witnessed a substantial increase, thereby inducing a rise in the Na+/K+ ratio. However, the Na+/K+ ratio in the roots of MhGATL1-OE Arabidopsis was remarkably lower compared to that of WT. This implies that MhGATL1-OE bolsters the saline-alkali tolerance of Arabidopsis by sustaining the Na+/K+ ratio, which aligns with the outcomes observed in barley[51]. These results indicated that the MhGATL1 transgenic lines enhanced the saline-alkali tolerance of the plants by promoting pectin synthesis, decreasing REC and MDA contents, increasing Pro and chlorophyll contents, increasing antioxidant enzyme activities (POD, SOD, CAT), and maintaining lower Na+/K+ ratios during stress.
To elucidate the molecular mechanism of MhGATL1 in saline-alkali response, we analyzed the expression of saline-alkali response representative genes and key genes for pectin synthesis at the transcriptional level by qRT-PCR. The expression levels of salinity-responsive representative genes (AtRD29A, AtNCED3, AtNHX1, AtKIN1, and AtP5CS) as well as key genes for pectin synthesis (AtMUM4, AtGATL5, AtQUA2, and AtPME1) were significantly increased in overexpressing MhGATL1 lines under saline-alkali stress. The results indicated that overexpression of MhGATL1 gene enhanced tolerance by inducing the expression of salinity stress-related representative genes and genes related to pectin synthesis pathway.
Protein interactions play a crucial role in understanding intracellular signaling mechanisms[52]. Previous research has indicated that GATL engages with other genes to modulate plant responses to abiotic stress conditions. In cotton, the GhGATL4_Dt gene is highly expressed in 0-DPA ovules and regulated by GbPDF1, which has been shown to be involved in fiber initiation and early elongation[53]. In this study, we showed that the apple MhGATL1 protein itself was not self-activating by Y2H assay, and screened to obtain seven candidate proteins that might have interactions with MhGATL1. Pairwise validation results indicated that MhbZIP23, MhSOS2, MhUGE5, MhABI3, and MhCHI proteins interact with MhGATL1 in yeast. These interactions are considered to play a significant part in plant tolerance to saline-alkali conditions. However, it should be noted that the Y2H method only verified the interactions in an in vitro context. Consequently, their functions and action mechanisms within plants remain to be further investigated.
-
MhGATL1 is highly expressed under abiotic stress, therefore, MhGATL1 was isolated from M. halliana. Overexpression of MhGATL1 in Arabidopsis, tobacco, and apple callus tissues responds to saline-alkali stress and increases its resistance, and exhibits its mechanism of motion underneath saline-alkali stress in four aspects: reactive oxygen system, ion homeostasis, osmoregulation, and pectin accumulation. Simultaneously, MhGATL1 was capable of enhancing saline-alkali tolerance by augmenting the expression of genes associated with the pectin synthesis pathway and promoting the accumulation of pectin content. Finally, MhGATL1 has been shown to interact with MhbZIP23, MhSOS2, MhUGE5, MhABI3, and MhCHI proteins. Therefore, the up-regulation of MhGATL1 under stress not only points out a direction for further exploring other functions of MhGATL1 but also lays a theoretical foundation for breeding apple rootstocks with efficient saline-alkali tolerance.
This work was funded by the National Natural Science Foundation of China (32460735).
-
The authors confirm contribution to the paper as follows: study conception and design: Wang Y; data collection: Wang X; analysis and interpretation of results: Li J, Zhang Z; draft manuscript preparation: Wang X. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 List of primers used for qRT-PCR and cloning of MhGATL1.
- Supplementary Table S2 List of GATL1 gene and information on Its encoded proteins.
- Supplementary Table S3 Major cis-acting elements of the promoter sequence of the MhGATL1 gene.
- Supplementary Table S4 Candidate interacting proteins obtained by Y2H screening of MhGATL1 protein.
- Supplementary Fig. S1 Expression analysis of MhGATL1 under different saline-alkali stress (a) and drought stress (b) time points.
- Supplementary Fig. S2 MhGATL1 gene amplification map. (a) PCR amplification product of MhGATL1 gene. (b) Electrophoresis images of pRI101-MhGATL1 PCR amplification.
- Supplementary Fig. S3 Protein sequence analysis of MhGATL1. (a) Schematic diagram of MhGATL1 protein affinity/hydrophobicity analysis. (b) Transmembrane structural domains of MhGATL1. (c) Phylogenetic analysis of this protein of MhGATL1 from M.halliana and other species.
- Supplementary Fig. S4 Sequence comparison between the MhGATL1 and other species.
- Supplementary Fig. S5 Identification of MhGATL1 transgenic materials. (a) DNA level identification. (b) RNA level identification.
- Supplementary Fig. S6 Screening and PCR detection of prey proteins for MhGATL1. (a) Screening of positive clones on plates. (b) Colony PCR detection of MhGATL1 positive clones.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang X, Li J, Zhang Z, Wang Y. 2025. Functional identification of saline-alkali stress enhancing gene GATL1 in apple (Malus halliana). Fruit Research 5: e010 doi: 10.48130/frures-0025-0001 

Functional identification of saline-alkali stress enhancing gene GATL1 in apple (Malus halliana)
- Received: 29 September 2024
- Revised: 27 December 2024
- Accepted: 06 January 2025
- Published online: 03 March 2025
Abstract: Pectin is an essential component of the plant cell wall and is critical for maintaining the structural integrity of the plant and protecting it from biotic and abiotic stresses. Galacturonosyltransferase-like (GATL), a crucial enzyme within the pectin synthesis route, governs the biosynthesis and accumulation of pectin. In this study, MhGATL1, cloned from Malus halliana, was significantly upregulated under saline-alkali stress conditions. The MhGATL1 gene was transformed into Arabidopsis, tobacco, and apple callus through Agrobacterium-mediated transformation to obtain transgenic materials. Their tolerance to saline-alkali stress was appraised through various physiological and biochemical assays. Meanwhile, the yeast two-hybrid (Y2H) approach was adopted to identify and corroborate the interacting proteins. The results revealed that transgenic plants exhibited superior growth under stress conditions, as manifested by increased pectin content, elevated levels of chlorophyll and proline, enhanced activities of antioxidant enzymes (SOD, POD, CAT), decreased conductivity and MDA content, and a more favorable Na+/K+ ratio in comparison to wild-type plants. Furthermore, genes related to salinity stress and key pectin synthesis pathway genes were significantly upregulated in the overexpression lines compared to the wild type. Y2H experiments further demonstrated that MhGATL1 interacts with several proteins, including MhbZIP23, MhSOS2, MhUGE5, MhABI3, and MhCHI. These results suggest that MhGATL1 positively regulates plant saline-alkali tolerance. Thus, this gene holds great promise as a useful genetic resource for the development of apple varieties with enhanced tolerance to saline-alkali conditions.
-
Key words:
- Malus halliana /
- MhGATL1 /
- Pectin synthesis /
- Saline-alkali stress