









-
The apple (Malus domestica) is a widely cultivated and commercially important fruit[1]. The growth of apple trees is affected by many environmental factors, which also affect the quality and yield of the fruit. These factors include the nutritional conditions required by the plant[2]. Over the past few decades, nitrogen has played a significant role in increasing crop yields. Nevertheless, the excessive use of nitrogen fertilizers in agriculture has raised serious ecological concerns, including the accumulation of large quantities of reactive nitrogen in the soil, the atmosphere, and water sources[3]. Reduced nitrogen retention and increased nitrogen leaching rates pose significant challenges to crop cultivation, and hinder the development of agricultural practices[4]. Consequently, efficient nitrogen utilization is a pivotal determinant in achieving higher crop yields[2,5]. To enhance nitrogen use efficiency, it is imperative to elucidate the roles of genes involved in nitrogen assimilation, absorption, and transport.
Most land plants absorb nitrogen from the soil as nitrate and ammonium[2]. To cope with environmental fluctuations, plants modify their root structure and have evolved various absorption systems to maintain and optimize growth, enabling a rapid response[6]. Plants have developed two different nitrate uptake systems to efficiently absorb nitrate, including most of the NITRATE TRANSPORTER 1 (NRT1)/NITRATE PEPTIDE TRANSPORTER family (NPF), which plays a low-affinity transport function, and the high-affinity transporters family NITRATE TRANSPORTER 2 (NRT2) family[7]. Numerous studies have shown that synergistic activities between high- and low-affinity nitrate transport mechanisms increase the efficiency of nitrate uptake, which in turn increases the efficiency of nitrogen utilization. Variation in NRT1.1B (OsNPF6.5) between japonica and indica rice (Oryza sativa) has led to differences in nitrate utilization, and NRT1.1B-indica has the potential to improve nitrogen utilization efficiency[8]. The threonine of NRT1.1 is phosphorylated in cells in response to changes in nitrate, completing the transition from a low- to a high-affinity state. As plants have both high- and low-affinity nitrate absorption, they can absorb nitrate over a broad range of nitrate levels[9]. AtNRT2.1 and AtNRT2.3 are specifically expressed in roots, where they are associated with the nitrate transport system and participate in nitrate absorption by the root system[10]. Overexpression and genome-wide association analyses indicate that the high-affinity nitrate transport protein OsNRT2.3 has an important function in nitrogen yield and use efficiency in rice. In addition, two OsNRT2.3 mRNA isoforms have been identified, including OsNRT2.3a and OsNRT2.3b. The higher OsNRT2.3b rice genotypes have higher nitrogen absorption and transport efficiency compared to OsNRT2.3a[11−14]. In apple, the expression of MdNRT2.1 is mainly concentrated in the root system, where it plays a crucial role in the formation and development of adventitious roots (ARs). In addition, the expression of MdNRT2.1 is regulated by nitrate availability, with higher expression levels observed under high nitrate conditions. In apple roots, the expression of MdNRT2.1 is closely linked to nitrogen uptake, and its overexpression can significantly increase the plant's nitrate absorption capacity[15,16]. NRT2.4, a high-affinity nitrate transporter in Arabidopsis, regulates nitrate uptake and mediates source-to-sink nitrate translocation[17].
Following absorption, the nitrate is distributed to aboveground tissues or stored within the root system. The ability of nitrate translocation from roots to shoots is species-specific and modulated by various environmental factors[2]. The first step in nitrate transport from root to shoot is the root xylem loading, after which it continues to be transported upwards. The nitrate transporter protein NRT1.5 exhibits expression in the pericycle cells adjacent to the xylem, where it mediates the loading process in Arabidopsis[18]. Nitrate transport to the shoots is low in the nrt1.5 mutant, as is nitrate content in xylem sap, suggesting that AtNRT1.5 mediates nitrate efflux from root pericycle cells to the xylem for loading[18]. Additionally, under stress conditions, nitrate is transported from the shoot to the roots for storage[6]. The transporters NRT1.9 and NRT1.8 serve as key regulators in this transportation process, with NRT1.8 being particularly instrumental[2]. AtNRT1.8 is a low-affinity nitrate transport protein localized to the plasma membrane of the cell and is mainly expressed in the xylem parenchymal cells in roots[19]. NRT1.8 is upregulated in response to nitrate availability, as well as to an array of abiotic and biotic stresses, including exposure to low temperatures, cadmium (Cd2+), salt, and pathogens invasion[2,19]. The increased nitrate content in the xylem sap of the nrt1.8 mutant, along with the increased allocation of nitrate from the roots to the shoots, indicates that NRT1.8 mediates xylem sap nitrate retrieval by root cells[19]. Similar to the nrt1.8 mutant phenotype, the nrt1.9 mutant exhibits increased nitrate transport from the roots to shoots, indicating that NRT1.9 and NRT1.8 have functional similarities[19]. It is important to have efficient retrieval and storage of nitrate for maintaining plant growth under low-nitrate environmental conditions, including during transportation and storage in young leaves and seeds[2]. Previous research has shown that AtNRT1.7 localizes to minor phloem veins in senescent leaves, and that it helps nitrate remobilization to younger tissues[20]. Subsequent investigations have revealed that AtNRT2.4 localizes to leaf phloem parenchyma cells, where it mediates nitrate transport during nitrogen scarcity conditions[17]. Research involving Arabidopsis has revealed that NRT1.12/NPF1.1 and NRT1.11/NPF1.2 transport nitrate to developing tissues, whereas AtNRT1.6/NPF2.12 transports nitrate to growing embryos[21,22]. Furthermore, it has been elucidated that NRT1.4/NPF6.2 plays an integral role in the sequestration of nitrate within the petioles[23].
Nitrate is not immediately available to the plant after being transported into the cells, but is stored in cellular vesicles for use when needed[24]. However, nitrate can be reduced to nitrite by cytoplasmic nitrate reductase (NR) catalysis, and nitrite is potentially toxic and must be reduced to ammonium through NR in plastids or chloroplasts. Ammonium is involved in the synthesis of amino acids, which form the basis of protein synthesis[2]. The amino acids synthesized during assimilation are involved in carbon and nitrogen homeostasis, so nitrate assimilation is essential for life processes[6,24]. In cellular systems, nitrate is initially reduced to nitrite by cytoplasmic NR, which is a limiting factor for the nitrate assimilation pathway[25]. Previous studies have shown that disruption of the NIA1 and NIA2 genes reduces NR activity in the Arabidopsis nia1/nia2 double mutant. Other studies have reported that two genes encoding Arabidopsis NR, such as NIA1 and NIA2, have different sequences but similar structures[25]. A transgenic tobacco study showed that two Arabidopsis NR gene promoters (NIA1 and NIA2) are induced by nitrate[25]. The function of NIA1 is restricted to leaf tissue, whereas NIA2 is active in highly dividing regions of meristematic tissue, such as the shoot apical meristem and cambial cells[26]. When nitrate is reduced to nitrite, nitrite is transferred to chloroplasts, where nitrite reductase (NiR) reduces nitrite to ammonium, which is involved in the synthesis of amino acids[26]. The NIR gene encoding NiR has been cloned from different species with gene numbers ranging from one to two copies[24]. In Arabidopsis, the gene responsible for encoding nitrite reductase, known as NIR1, is essential and sufficient for the nitrate response, and plays a key role in nitrite assimilation processes[27]. In addition, NIR1 shows conservation across a wide range of higher plant species[27].
Nitrate functions as a signaling agent, triggering both cellular and long-distance signaling cascades that modulate gene expression, metabolism, physiological processes, and growth. A large number of genes have been identified as regulators within these nitrate-mediated pathways[28]. It has been shown that MdBT2 partially enhances nitrate-stimulated plant growth partly through downregulation of the DELLA protein MdRGL3a[29]. MdMYB10 transcriptionally upregulates MdNRT2.4-1 by directly binding its promoter region, thereby increasing nitrate uptake[30]. MdMYB88 and MdMYB124 regulate nitrogen use by regulating the expression of MdNRT1.5/1.7/1.8 and MdNRT2.4 to promote nitrate use under nitrogen-limited conditions[31]. The transcription factor MdHY5 directly interacts with the G-box motif within the promoter region of MdMYB10, thereby inducing its transcriptional activity and orchestrating the regulation of anthocyanin biosynthesis[30]. In addition, MdHY5 has been implicated in nitrate utilization by regulating the transcription of NR and nitrate uptake genes[32]. The MdNLP7 transcription factor in apple promotes nitrogen uptake and assimilation through transcriptional activation of MdNRT1.1 and MdNIA2, thus MdNLP7 promotes growth and substantial increases in biomass under low NO3- conditions[33].
Numerous phytohormones influence the expression of relevant genes via signaling cascades, thereby exerting further control over nitrate concentrations within the plant system. The auxin response factor 18 (ARF18) Apple ARF18 negatively regulates nitrate absorption and nitrogen use efficiency by directly inhibiting the expression of NRT1.1[34]. Cadmium and sodium stress initiate the ethylene/jasmonic acid (Eth/JA) signaling pathway and aggregate at the ETHYLENE INSENSITIVE3 (EIN3)/EIN3-like1 (EIL1) locus to regulate the expression of ethylene response factors (ERFs), thereby upregulating NRT1.8[35]. In addition, the Eth and JA signaling pathways further regulate stress tolerance and plant growth in an NR-dependent manner through EIN3/EIL1 and other unknown components that mediate the downregulation of NRT1.5[35]. ABA, a pivotal hormone in stress responses, plays an essential role in a myriad of growth, developmental processes, and adaptive stress responses[36,37]. ABA is hypothesized to be involved in nitrate signaling. Empirical evidence from a suite of experiments indicates that in Arabidopsis and wheat, ABA treatment triggers the induction of nitrate-responsive gene expression, suggesting an active participation of ABA in nitrate signaling pathways[38]. Research on Malus domestica has revealed that MdABI5, a component of the ABA signaling cascade, interacts with the MdNRT1.5 promoter and represses its expression, thereby hindering root-to-shoot nitrate translocation[39].
The mechanism of ABA signaling perception is widely known and involves three important core components, including PYR1/PYL/RCAR (Pyrabactin resistance1/PYR1-like/regulatory components of the ABA receptor), PP2C (Clade A type 2C protein phosphatases), and SnRK2s (subclass III sucrose non-fermenting-1 related protein kinase 2)[38]. PYR1/PYL/RCAR is an ABA receptor, which inactivates PP2C upon binding with ABA, thereby relieving the inhibition of SnRK2 activity[40]. Activated SnRK2 kinases (SnRK2.2/2.3/2.6) phosphorylate ABRE-binding factors (ABFs/AREBs), inducing expression of many ABA-responsive genes[41]. Among them, as the key factors in the ABA signaling pathway, ABFs participate in the regulation of adversity stress (abiotic and biotic stress), and biological developmental processes (senescence and early flowering)[42−44]. The role of ABFs in nutrient utilization remains ambiguous; however, emerging evidence hints at a transient interaction between ABFs and nitrate. Despite these indications, the precise molecular mechanisms underpinning this association have yet to be elucidated[45]. The ABFs generally tend to bind to specific classes of cis-acting elements, which play important roles in their regulatory networks[42,46]. Although the chromatin-binding profile of the MdABF1 gene in apple has not yet been elucidated, its homology with the Arabidopsis ABF genes suggests that MdABF1 may regulate the expression of downstream genes through similar mechanisms. In this research, we explored the molecular mechanism by which MdABF1 regulates nitrate utilization in apple. Our data revealed that MdABF1 directly interacted with the promoter regions of MdNIA1 and MdNRT2.3, resulting in the transcriptional repression of these genes. At the same time, MdABF1 had an indirect regulatory effect on MdNIA2 and MdNRT2.1, inhibiting nitrate absorption and assimilation. MdABF1 is a direct transcriptional activator of MdNRT1.8, but it exerts an indirect inhibitory effect on MdNRT1.5, thereby facilitating the transport of nitrate from the stem to the roots. In summary, MdABF1 regulates the MdNRTs/NIAs module, which in turn controls the processes of nitrogen absorption, assimilation, and allocation. This sheds light on the molecular mechanism by which MdABF1-mediated regulation of ABA influences nitrate utilization.
-
GL-3 apples, which are derived from 'Royal Gala' (Malus domestica), were used as the wild type. The apple seedlings were cultured on Murashige and Skoog (MS) medium supplemented with 0.1 mg/L gibberellic acid (GA3), 0.2 mg/L 1-naphthaleneacetic acid (NAA), and 0.6 mg/L 6-benzylaminopurine (6-BA) at 25 °C under a 16-h light/8-h dark photoperiod for 30 d. After rooting, the seedlings were grown in a basic nutrient solution with 5 mM KNO3 for 30 d. Subsequently, seedlings of a similar size and condition were transferred to nutrient solutions containing different concentrations of nitrate (0.5 and 15 mM KNO3) and different ABA concentrations (0 and 40 μM ABA), and incubated for 2 h under long-day conditions (25 °C, 16-h light/8-h dark). The seedlings were then rapidly transferred to liquid nitrogen and stored for quantitative real-time polymerase chain reaction (qRT-PCR). The gene expression of root seedlings at the same growth state was examined after they were treated with 0 and 0.5 mM KNO3 for 0, 3, 6, 9, 12, and 24 h.
Chinese crabapple (Malus hupenensis) was used in this experiment. The seeds were stored at 4 °C for 40 d, then germinated and grown in a 5 mM KNO3 nutrient solution for 5 weeks. Uniformly growing seedlings were selected and carefully transplanted into untreated vermiculite and treated with 0.5 mM KNO3 or 15 mM KNO3 and supplemented with 0 or 40 μM ABA for 45 d. The growth environment was long-day conditions (16-h light/8-h dark) at 25 °C. The solution was changed once a week. Samples were collected 45 d later, the roots were cleaned with ddH2O, dried using filter paper, and analyzed for the fresh weight, NR activity, and nitrate content.
Calli of the apple cultivar 'Orin' were maintained on MS medium with 1.5 mg/L 2,4-dichlorophenoxyacetic acid (2,4-D) and 0.4 mg/L 6-BA at 25 °C in the dark and at intervals of 16 d. Subsequently, the apple calli were treated with KNO3 (0.8 or 5 mM) nutrient solutions supplemented with ABA (0 or 20 μM), and their phenotypes were observed to detect relevant indicators. Transiently transformed transgenic calli were also treated with 0 or 20 μM ABA for 1 d and used for GUS staining experiments.
The Arabidopsis thaliana ecotype 'Columbia' (Col) was used for genetic transformation and functional identification. Arabidopsis seeds were grown on MS medium and vernalized at 4 °C for 3 d. The seeds were placed in an incubator for 1 week, and supplemented with 0.2 or 10 mM KNO3 and 0, 20, or 30 μM ABA under long-day conditions (16-h light/8-h dark) at 22 °C.
We used tobacco (Nicotiana benthamiana) grown for 1 month under long-day conditions (16-h light/8-h dark) at 24 °C for the transient dual luciferase assay.
The branches of Red 'Fuji' apple trees were taken from the Experimental Station of Shandong Agricultural University (Shandong, China). The branches were sprayed three times with 0 or 50 μM ABA. Subsequently, the two groups of branches were incubated at 25 °C for 16-h light/8-h dark and sampled at 0, 1, 3, 6, and 12 h for gene expression, and at 0, 2, 4, 7, and 10 d for nitrate content and NR activity analyses.
Phylogenetic tree analysis of the ABF family
-
A phylogenetic tree of ABF family proteins from apple and Arabidopsis was created using MEGA7.0, the tree was constructed by the Maximum Likelihood method (bootstrap = 1,000). Protein sequences were aligned with MUSCLE and analyzed using the p-distance model. Then, the ABF protein sequence information was annotated using iTOL (
https://itol.embl.de/itol.cgi ) as well as the overall landscaping.Plasmid construction and plant transformation
-
The MdABF1 full-length cDNA was inserted into the pRI101-AN (Takara, Dalian, China) containing the CaMV 35S promoter and the recombinant 35S::GFP-GW vector to construct the overexpressing vector. Consequently, the overexpressing vectors were transferred into apple calli and Arabidopsis by Agrobacterium LBA4404.
The 35S::MdABF1-GFP vector was transformed into 'Orin' apple calli via Agrobacterium-mediated transformation[29]. A mixture of 30 mg/L kanamycin and 250 mg/L carbenicillin was added to the medium to obtain transgenic apple calli[29].
The 35S::MdABF1-GW vector was transformed with Agrobacterium strain GV3101 on Arabidopsis (Col) using the flower dip method[47]. The seed stocks were subjected to transgenic screening on 1/2 MS medium supplemented with 30 mg/L glufosinate ammonium, resulting in a T3 pure transgenic strain for further studies[29].
RNA extraction and qRT-PCR analysis
-
Whole-plant tissues of apple and Arabidopsis were subjected to qRT-PCR analysis to assess systemic gene expression patterns. Total RNA was extracted from apple plants, calli, and Arabidopsis using the RNA Plant Extraction Kit (CWBIO, Beijing, China) according to the manufacturer's instructions[29]. Subsequently, reverse transcription was performed using the PrimeScriptTM RT First Strand cDNA Synthesis Kit (Takara, Dalian, China)[48]. Next, qRT-PCR was performed using UltraSYBR mix (SYBR Green I) (Takara) to detect the gene transcript levels[49]. Finally, relative gene expression was determined by the 2−ΔΔCᴛ method using 18S rRNA as the internal control[50]. Three independent biological replicates were performed for each sample separately. The qRT-PCR primers are listed in Supplementary Table S1.
Transient dual-luciferase assay
-
Tobacco (Nicotiana tabacum) leaves were used for the transient expression assay[51]. MdNRT1.8, MdNRT2.3, and MdNIA1 promoter fragments were cloned into the pGreenII 0800-LUC vector to generate luciferase reporter genes (proMdNRT1.8-LUC, proMdNRT2.3-LUC, proMdNIA1-LUC). MdABF1 was cloned into the pGreenII 62-SK vector to generate the effector (35S pro::MdABF1). The recombinant plasmids were transformed into Agrobacterium LBA4404, respectively. They were then injected into tobacco leaves, and incubation was performed as described previously[39,51]. Finally, fluorescence was detected using a live imager.
Promoter-GUS-staining assay
-
To assess the effect of MdABF1 on MdNRT1.8, MdNRT2.3, and MdNIA1 transcriptional activity under ABA treatment, these promoters were inserted into the p1300-GN-GUS reporter vector. Empty vector controls (promoter-free constructs with the GUS reporter gene only, called GUS in subsequent sections) were included in GUS assays to account for background activity. The recombinant vectors pMdNRT1.5::GUS, pMdNRT2.3::GUS, pMdNIA1::GUS, and GUS were transiently transformed into wild-type and 35S::MdABF1 transgenic apple calli via Agrobacterium (4404)[39]. These calli were transferred to medium with 0 or 20 μM ABA, respectively, and treated for 1 d. The GUS staining and activity assays were performed after treatment. The design of this GUS assay was conducted in accordance with a previously published study[39].
Electrophoretic mobility shift assay (EMSA)
-
An EMSA was completed as described previously[39]. The coding sequence of MdABF1 was cloned into pET32a and expressed as a His-tagged recombinant protein in BL21 (DE3), followed by purification. Oligonucleotide probes specifically tagged with 3'-biotin for the MdNRT1.8, MdNRT2.3, and MdNIA1 promoters were used as described previously[39]. The same unlabeled sequences were used to compete. Supplementary Table S1 lists the primers used.
Yeast one-hybrid (Y1H) assay
-
Y1H experiments were performed as described previously[39]. The MdABF1coding sequence was cloned into pGADT7, while promoter fragments of the MdNRT1.8, MdNRT2.3, and MdNIA1 containing the ABRE binding elements were inserted into the pHIS2 reporter vector. MdABF1 was co-transformed with reporter constructs into yeast strain Y187. Individual transformants were screened on SD/-Trp-Leu-His medium, then positive yeast cells were serially diluted 10-fold, 100-fold, and 1,000-fold. A 10 µL aliquot of the different concentrations of solutions was dropped onto selective medium (SD/-Trp-Leu-His with 90 mM 3-aminotriazole) and incubated at 28 °C for 2−3 d.
Nitrate content and nitrate reductase activities
-
To detect nitrate, Chinese crabapple apple seedlings were exposed to nitrogen-deficient conditions for 7 d and then treated with high (15 mM KNO3), or low (0.5 mM KNO3) nitrogen nutrient solutions and given 0 or 40 μM ABA, respectively. After 45 days of treatment, the fresh weights of the plants were determined, and nitrate content and NR activity were measured in the roots and stems of Chinese crabapple seedlings.
Arabidopsis seeds were stratified on MS medium and treated at 4 °C for 3 d. The seedlings were transferred to incubators for 4 d. Then, the seedlings were grown for 2 weeks under varying nitrate (10 or 0.2 mM KNO3), and ABA (0 or 20 μM) concentrations. Root length and fresh weight were determined, while the material was collected for nitrate content and NR activity. Untreated Arabidopsis in the incubator was cultured on MS medium for 2 weeks and then planted in vermiculite containing 2 mM KNO3 for 1 week, watered with 10 or 0.2 mM KNO3, and treated with 0 or 20 μM ABA for 2 weeks, respectively. Samples of the plants were assayed for fresh weight, nitrate content, and NR activity. The wild-type and MdABF1 transgenic calli cultured for 16 d were transferred to 5 or 0.8 mM KNO3 and treated with 0 or 20 μM ABA for 2 weeks. Then, samples were taken, and fresh weight, nitrate content, and NR activity were assayed. Nitrate content was determined using the hydrazine reduction method previously described[39]. The NR activity was assayed according to an NR activity assay kit (Keming, Suzhou, China).
The composition of the medium used to treat the Chinese crabapple apple seedlings and Arabidopsis included a basic nutrient solution containing 1.0 mM KH2PO4, 1.0 mM MgSO4, 1.0 mM CaCl2, 0.1 mM FeSO4·7H2O, 0.1 mM Na2EDTA·2H2O, 50 μM H3BO3, 50 μM MnSO4·H2O, 0.5 μM Na2MoO4·2H2O, 2.5 μM KI, 15 μM ZnSO4·7H2O, 0.05 μM CuSO4·5H2O, and 0.05 μM CoCl·6H2O, supplemented with 15, 10, 0.5, or 0.2 mM KNO3 as the sole N source, with KCl added to balance K concentrations[39].
MS Base Salts (-N) with 1.5 mg/L 2,4-D and 0.4 mg/L 6-BA plus 5 or 0.8 mM KNO3 as the sole N source.
The medium used to treat calli was MS Base Salts (-N) with 1.5 mg/L (2,4-D), and 0.4 mg/L 6-BA, along with 5 or 0.8 mM KNO3 as the sole N source.
Statistical analysis
-
A minimum of three biological replicates were performed for each experiment. Data were expressed as mean and standard deviation. The statistical analysis was performed using the LSD test in DPS software. A p-value ≤ 0.05 was considered significant.
-
ABA plays a pivotal role in plant growth and development, and is also crucial in mediating stress responses[36]. To study the role of ABA in nitrogen utilization, apple seedlings were exposed to either high (15 mM KNO3) or low (0.5 mM KNO3) nitrogen levels and received either 0 or 40 μM ABA treatments. After 45 d, the plants exposed to low nitrate concentration were smaller than those in the control group, which received high nitrate concentrations (Fig. 1a). Notably, the addition of ABA treatment accentuated the size reduction more markedly under both low and high nitrate conditions, indicating a significant interaction effect between nitrate concentration and ABA on plant growth (Fig. 1a). Fresh weight decreased significantly after the low nitrate and ABA treatments, based on the high and low nitrate treatments (Fig. 1b). Nitrate content and NR activity also decreased significantly under low nitrate and ABA treatments, while the root/shoot ratio of nitrate content increased (Fig. 1c−e). In addition, the MdABF1 expression increased following ABA treatment under both high and low nitrogen conditions, while the expression levels of nitrate absorption and assimilation genes decreased (Fig. 1f−n). These findings indicate that both low nitrate availability and ABA supplementation lead to reduced nitrogen uptake and assimilation in plants.
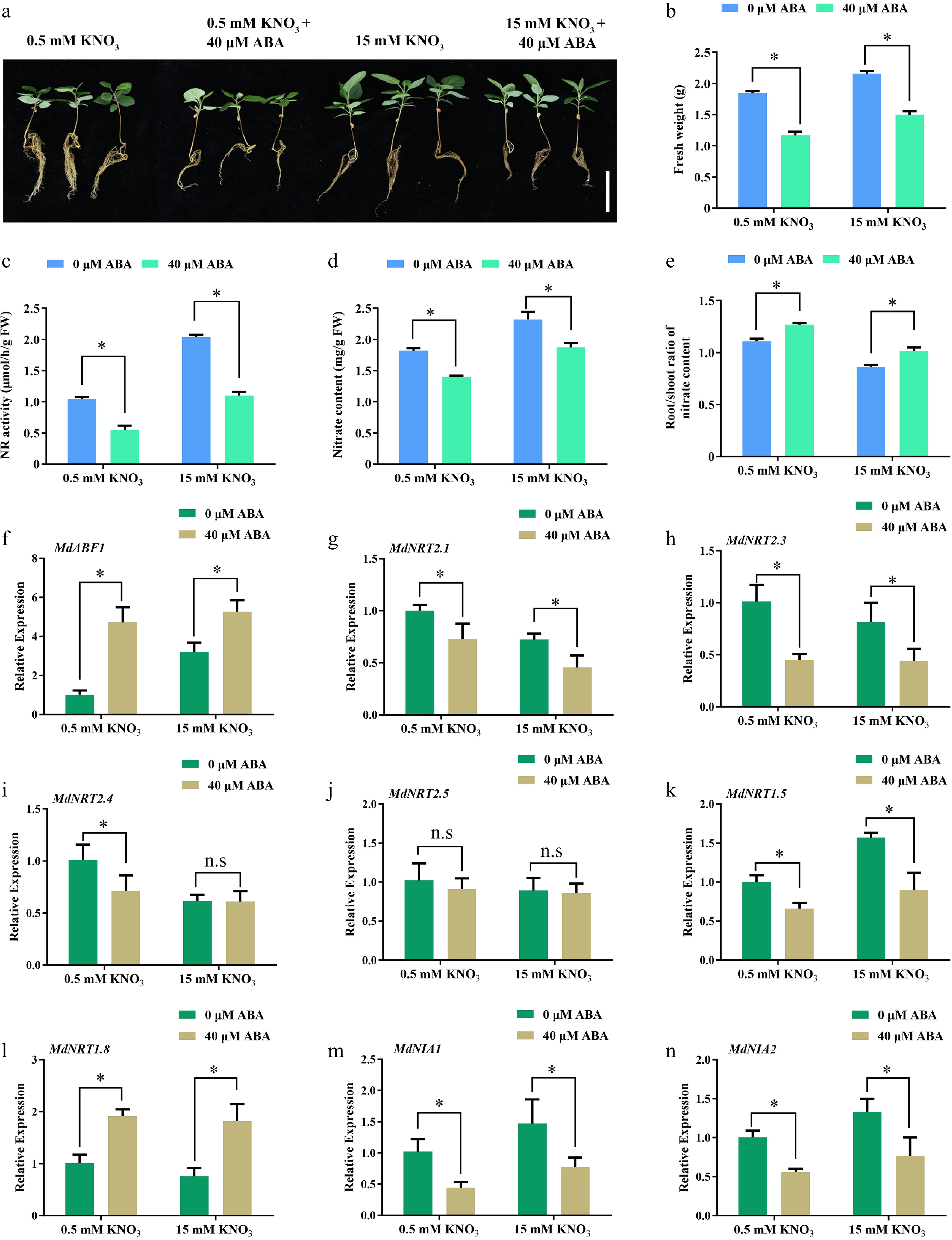
Figure 1.
ABA regulates nitrate absorption and assimilation. (a) Phenotype of apple seedlings grown on low (0.5 mM KNO3), and high (15 mM KNO3) nitrate nutrient solutions containing different ABA concentrations (0 or 40 μM). (b) Fresh weight, (c) nitrate reductase activity, (d) nitrate content, (e) root/shoot ratio of nitrate content in (a). The relative expression of (f) MdABF1, and (g)−(n) nitrate absorbing, transporting, and assimilation genes. Plants were treated with high (15 mM) or low nitrate (0.5 mM) concentrations containing different ABA (0 or 40 μM) for 2 h, and RNA was extracted for gene expression analysis. Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
Nitrate absorption and assimilation are regulated by ABA-mediated MdABF1
-
The regulatory network that governs nitrate has been extensively documented in the scientific literature. Changes in the nitrate-responsive transcriptome have been measured over time in the five main types of root cells in Arabidopsis. ABF2 and ABF3 serve as central transcriptional regulators in the endodermal cell layer[45]. This cell layer exhibits the most tightly interconnected gene network responsive to nitrate availability[45]. Therefore, it has been speculated that ABF transcription factors may participate in the nitrate utilization process. We examined the evolutionary relationship and expression of MdABFs in nitrate-treated apple seedlings; the results showed that MdABF1 was significantly induced by nitrate (Supplementary Fig. S1a−d). Furthermore, the ABF family is widely recognized as a pivotal element in the ABA signaling cascade[36,39]. Previous studies have found that MdABF1 can be induced by ABA and is significantly induced in the MdABFs family. Therefore, MdABF1 was selected for further investigation.
To elucidate the role of MdABF1 more comprehensively, we obtained MdABF1 overexpressing Arabidopsis and apple calli by the Agrobacterium-mediated method (Supplementary Fig. S1e−h). The overexpressed materials were treated with or without ABA under high and low nitrogen conditions. The results indicated that lines overexpressing MdABF1 in Arabidopsis and apple calli displayed significantly shorter primary roots (Fig. 2a−c, Supplementary Fig. S2a, b) and reduced biomass when subjected to low nitrate and ABA treatment compared to the wild-type, as evidenced by significantly lower fresh weights (Fig. 2d, e, Supplementary Fig. S2c, d, Fig. 3a−c, Supplementary Fig. S2i, j). Nitrate content and NR activity were decreased in overexpressed Arabidopsis and apple calli (Figs 2f−i and 3d−g, Supplementary Fig. S2e−h, S2k−n). The findings suggest that the overexpression of MdABF1 results in decreased nitrate content and NR activity, thereby impeding growth under conditions of low nitrate and ABA exposure, in both low and high nitrate environments.
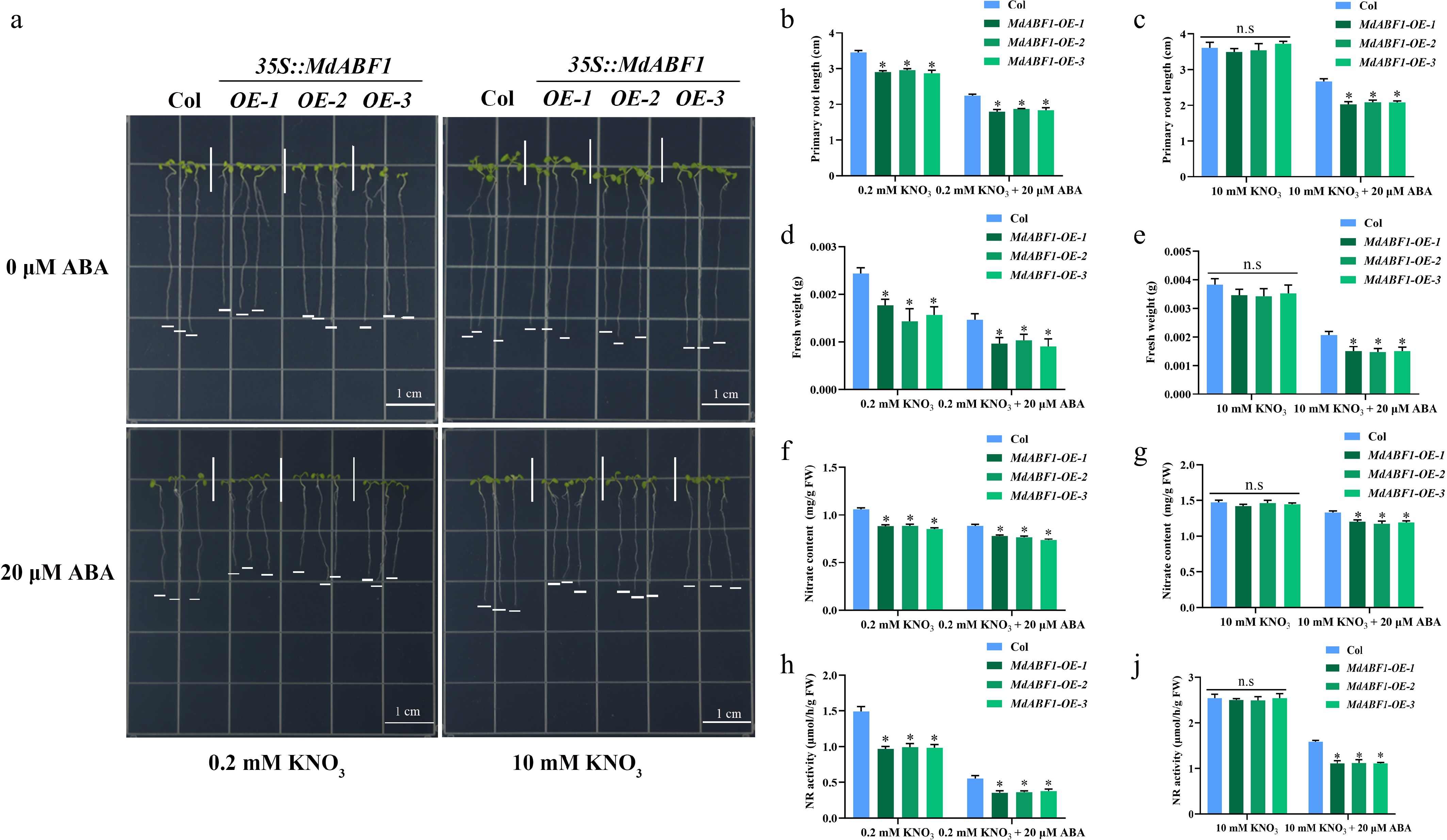
Figure 2.
Ectopic expression of MdABF1 in Arabidopsis regulates nitrate absorption and assimilation under ABA treatment. (a) Morphological characteristics of MdABF1 transgenic plants under low (0.2 mM), or high (10 mM) nitrate supplemented with 0 or 20 μM ABA treatment. Determination of (b), (c) primary root length, (d), (e) fresh weight, (f), (g) nitrate content, (h), (i) NR activity. Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
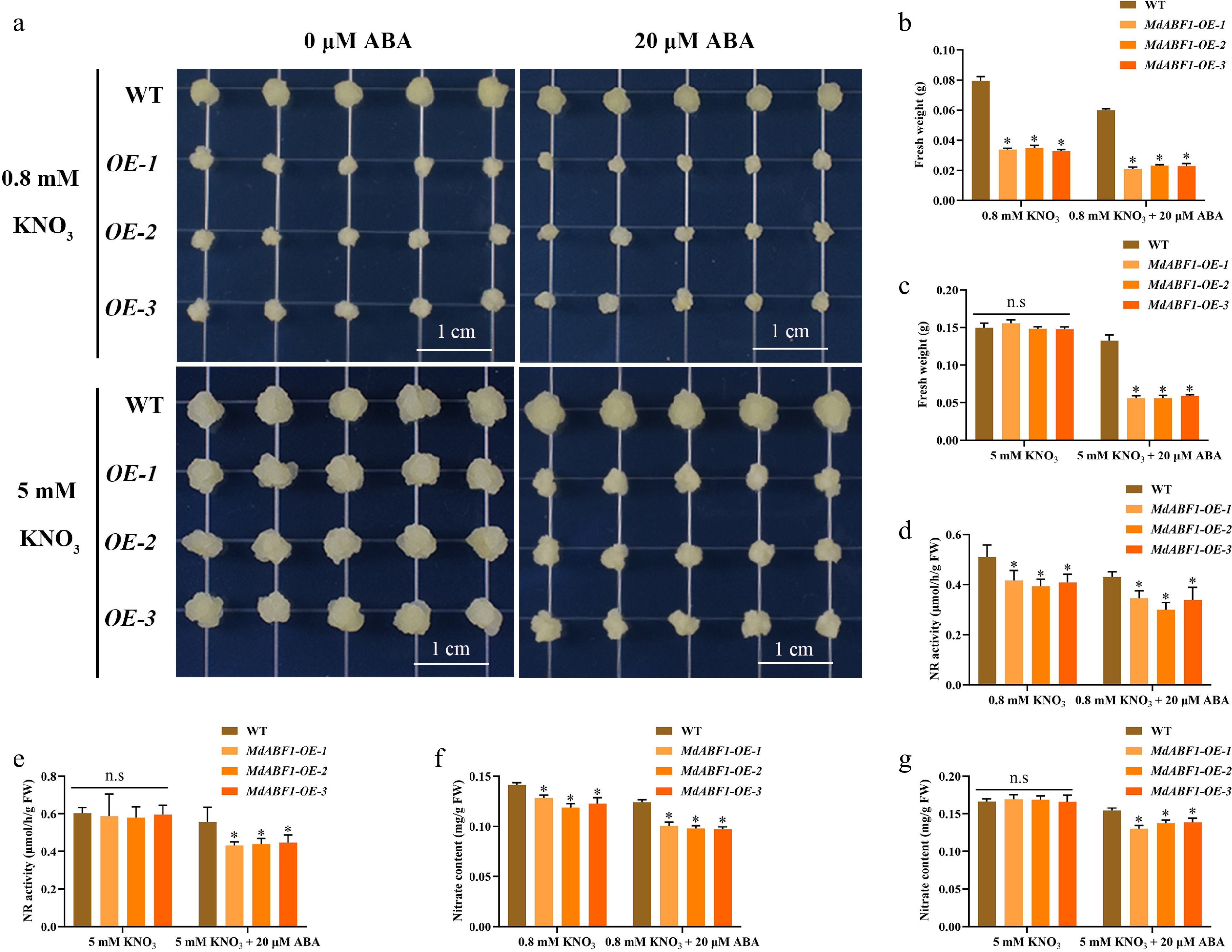
Figure 3.
Response of MdABF1 transgenic apple calli to different nitrate and ABA conditions. (a) Phenotypes of apple calli cultured under 0.8 mM and 5 mM KNO3 supplemented with 0 or 20 μM ABA. (b−g) Fresh weight (b, c), NR activity (d, e), nitrate content (f, g) in (a). Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
ABA-mediated regulation of the expression of genes related to nitrate uptake and assimilation by MdABF1
-
To further unravel the molecular mechanisms underlying MdABF1-mediated nitrate utilization, we analyzed data from previous research studies[45]. Chromatin immunoprecipitation (ChIP) and Transient Assay Reporting Genome-wide Effects of Transcription factors (TARGET) analyses identified NRT2.1, NIA1, and NIA2 as downstream targets of ABF involved in nitrate utilization. However, NIA1, NIA2, NRT2.1, and NRT2.3 were all involved in the nitrate utilization under the low nitrate and ABA treatment (Fig. 1f−n), and these genes were downregulated in the MdABF1 overexpressed materials (Fig. 4a−p, Supplementary Fig. S3a−p). MdABF1 was found to inhibit the expression of the NIA1, NIA2, NRT2.1, and NRT2.3 genes to a greater extent following exogenous ABA treatment in Arabidopsis and apple calli (Fig. 4a−p, Supplementary Fig. S3a−p). The data demonstrate that the MdABF1 gene suppresses the expression of NIA1, NIA2, NRT2.1, and NRT2.3 under low nitrate conditions, with the inhibitory effect being more pronounced in the presence of ABA.
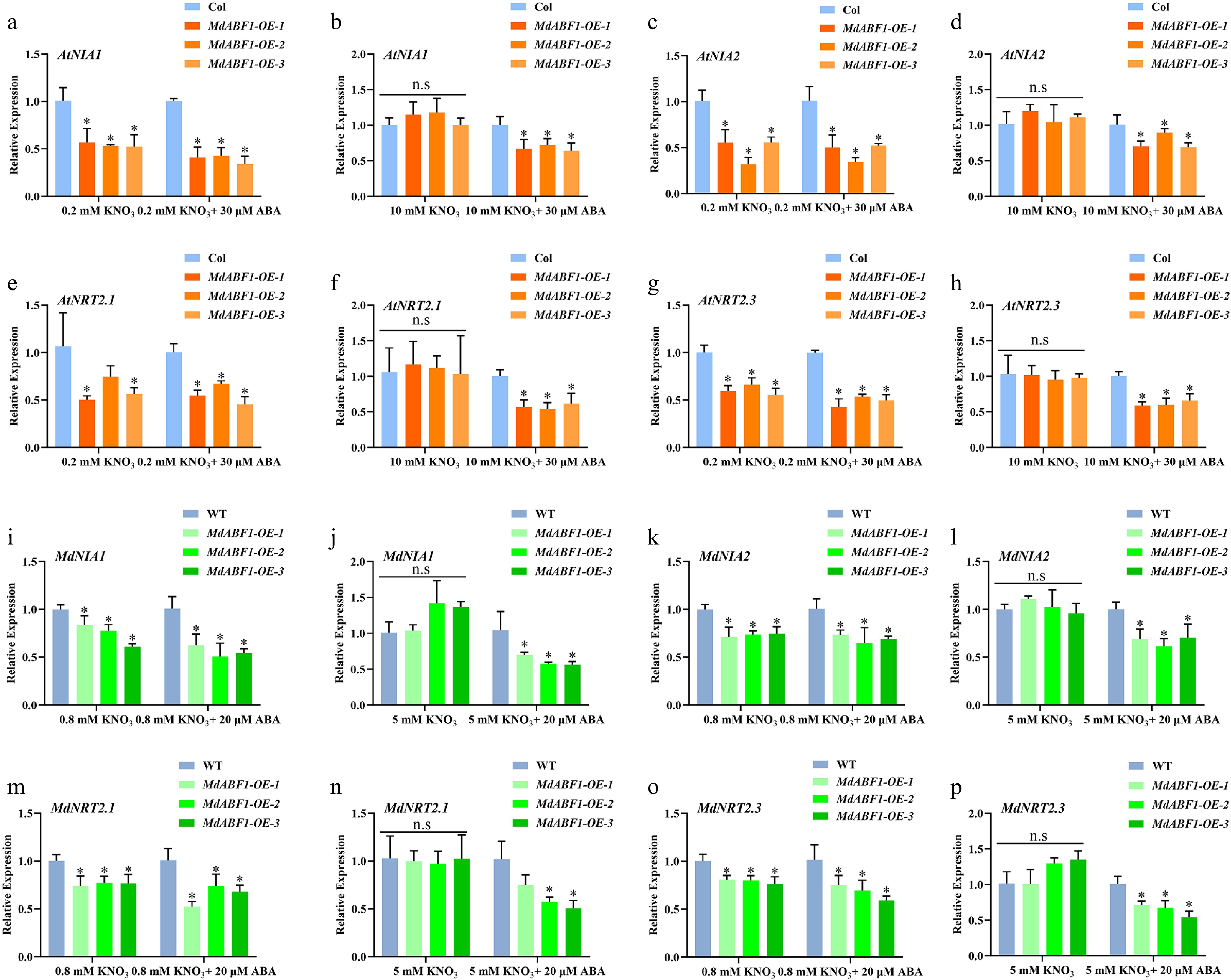
Figure 4.
Relative expression of NRT2s and NIAs in MdABF1 transgenic plants in response to nitrate and ABA. (a)−(h) Relative expression of AtNRT2s and AtNIAs in MdABF1 transgenic Arabidopsis under 0.2 mM and 10 mM nitrate conditions supplemented with 0 or 20 μM ABA. (i)−(p) Relative expression of MdNRT2s and MdNIAs in MdABF1 transgenic apple calli was incubated in 0.8 mM and 5 mM KNO3 with 0 or 20 μM ABA. Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
MdABF1 specifically binds the promoters of MdNRT2.3 and MdNIA1 and transcriptionally represses their expression
-
The expression of NIA1, NIA2, NRT2.1, and NRT2.3 was repressed by MdABF1. To further verify whether MdABF1 binds directly to the promoters, the MdNIA1, MdNIA2, MdNRT2.1, and MdNRT2.3 promoters with 2,000 bp sequences were selected. These promoters were analyzed to identify the presence of ABRE motifs, which are recognized by the MdABF1protein. To determine whether MdABF1 binds directly to the MdNIA1, MdNIA2, MdNRT2.1, and MdNRT2.3 promoters, we performed EMSA experiments using the MdABF1-His protein. The MdNIA1-DNA-MdABF1, MdNRT2.3-DNA-MdABF1 specific complexes were detected, and their frequency decreased with increasing amounts of unlabeled ABRE competitor added (Figs 5a, b and 6a). The MdNIA1-MdABF1 and MdNRT2.3-MdABF1 DNA-protein complexes did not appear when the ABRE cis-elements CACGTG were mutated to AAAAAA (Figs 5a, b and 6a).
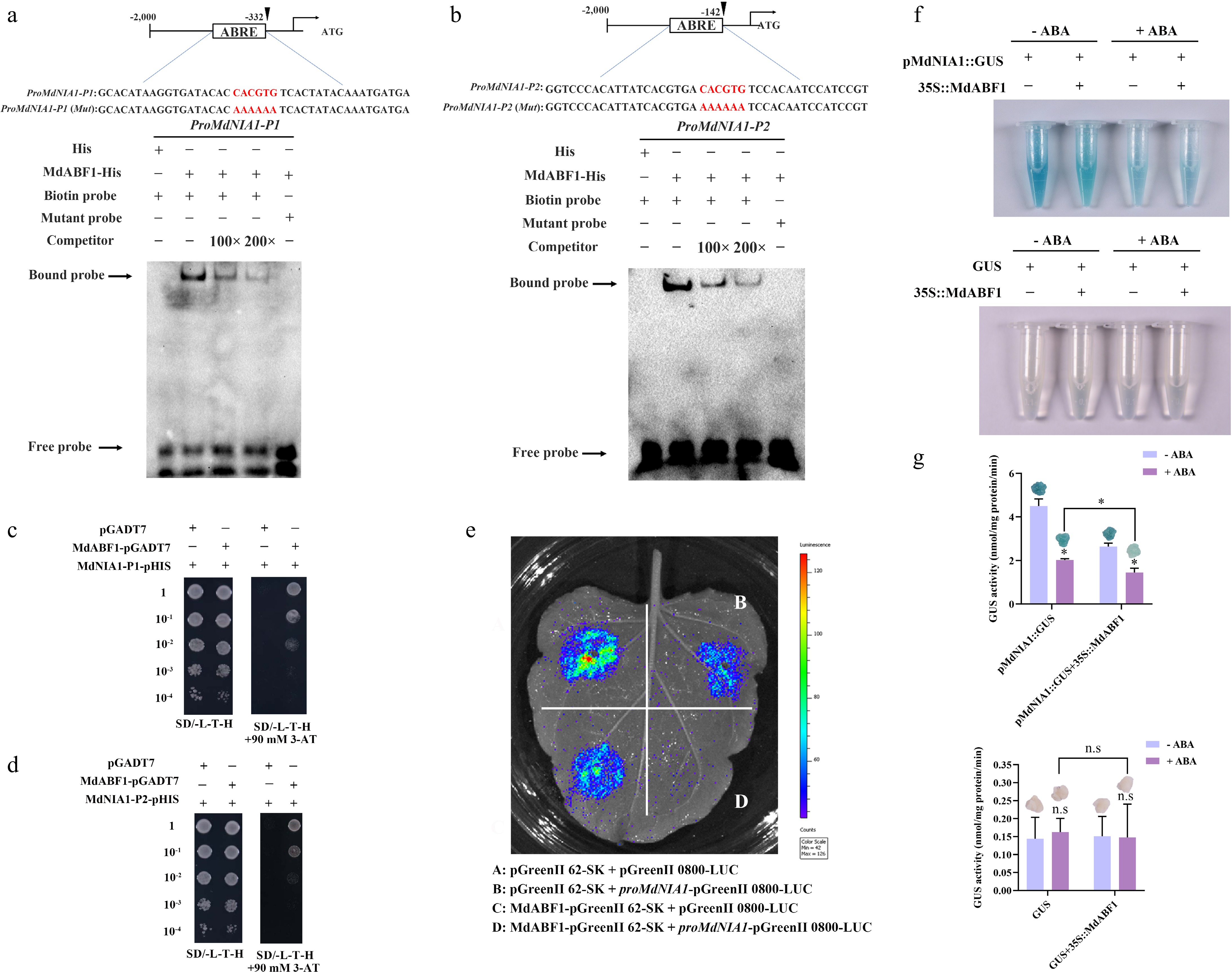
Figure 5.
MdABF1 binds to the promoter of MdNIA1. EMSA analysis showing the binding of MdABF1 to the (a) P1, and (b) P1 fragments of MdNIA1 promoter. Diagrams of the promoter of MdNIA1 are shown above. (c), (d) Y1H assay of the binding of MdABF1 to the MdNIA1 promoter in vitro. (e) Dual luciferase assays in tobacco leaves showing that MdABF1 transcriptionally repressed the expression of MdNIA1. (f) GUS staining experiment of apple calli showing that the repression of MdABF1 to the MdNIA1 promoter dependent on ABA. (g) GUS activity in (f). Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
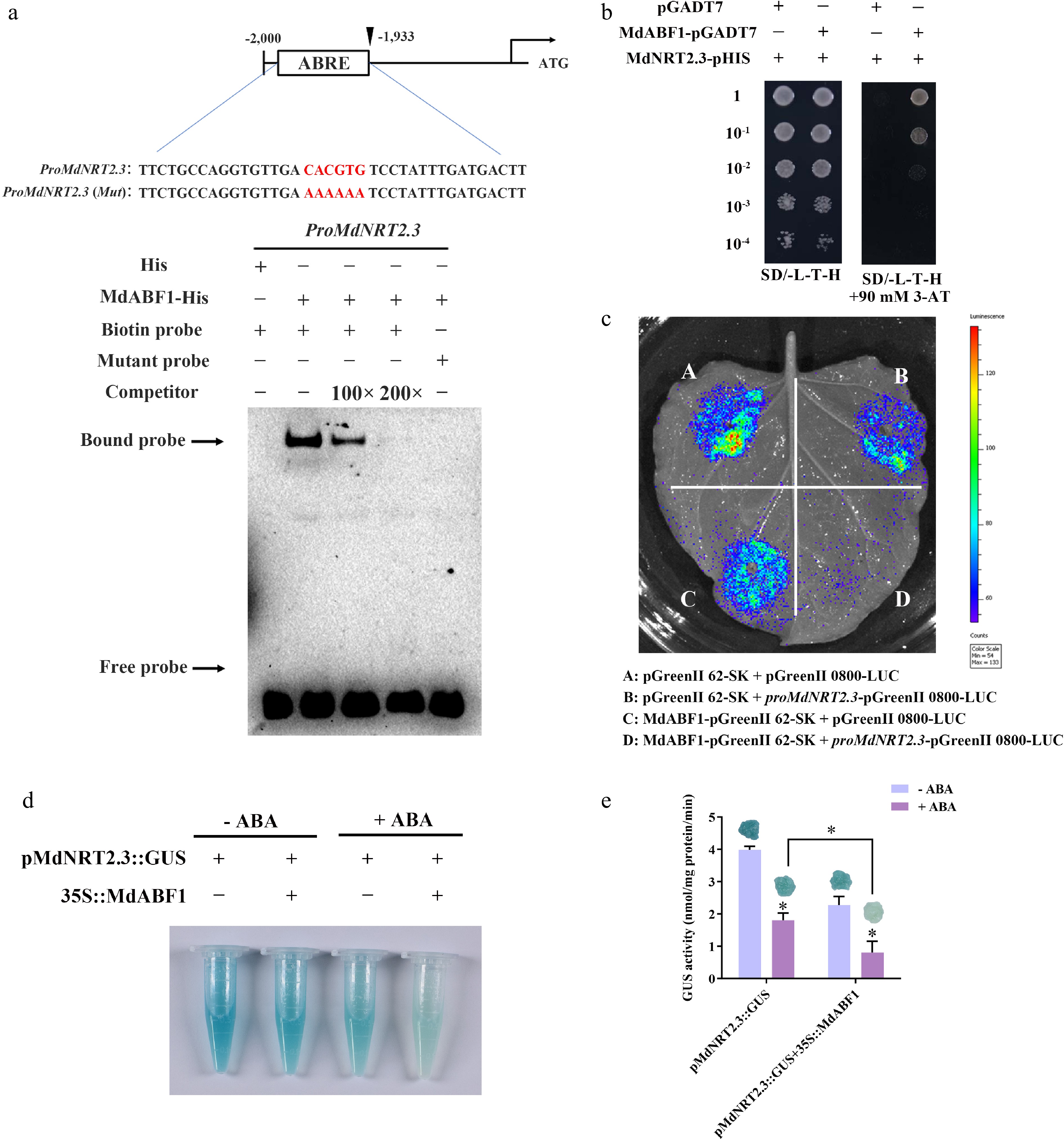
Figure 6.
MdABF1 binds to the promoter of MdNRT2.3. (a) EMSA analysis showing the binding of MdABF1 to the MdNRT2.3 promoter. Diagram of the promoter of MdNRT2.3 is shown above. (b) Y1H assay of the binding of MdABF1 to the MdNRT2.3 promoter in vitro. (c) Dual luciferase assays in tobacco leaves showing that MdABF1 transcriptionally repressed the expression of MdNRT2.3. (d) GUS staining experiment of apple calli showing the repression of MdABF1 to the MdNRT2.3 promoter. (e) GUS activity in (d). Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
The interaction between the MdABF1 protein and the promoters of MdNIA1 and MdNRT2.3 was further substantiated through Y1H assays. Co-transformation of MdNRT2.3-P-pHIS2, MdNIA1-P-pHIS2, and pGADT7 into yeast cells failed to support growth on selective SD medium. However, cells co-transfected with MdNIA1-P-pHIS2 and MdABF1-pGADT7 or MdNRT2.3-P-pHIS2 and MdABF1-pGADT7 exhibited normal growth on the selective medium (Figs 5c, d and 6b). This finding indicates that the MdABF1 protein directly binds to the promoters of MdNIA1 and MdNRT2.3.
We verified whether MdABF1 is a transcriptional activator or repressor of MdNIA1 and MdNRT2.3 using the dual luciferase assay. Compared to the other combinations, the fluorescence intensity was largely absent following the co-injection of proMdNIA1-pGreenII 0800-LUC, proMdNRT2.3-pGreenII 0800-LUC and MdABF1-pGreenII 62-SK into tobacco (Figs 5e and 6c). Consequently, the data suggest that the MdABF1 protein exerts an inhibitory effect on the transcription of MdNIA1 and MdNRT2.3.
Furthermore, we examined the expression of MdNRT2.3 and MdNIA1 in MdABF1 by transiently transforming MdNRT2.3 and MdNIA1 into MdABF1 overexpressing calli, which results in decreased expression of MdNIA1 and MdNRT2.3. The expression of MdNIA1 and MdNRT2.3 decreased further in the presence of ABA (Figs 5f, g and 6d, e). These findings demonstrate that protein MdABF1 directly interacts with the promoters of MdNIA1 and MdNRT2.3, which leads to the repression of their gene expression.
ABA regulates the reallocation of nitrate
-
Previous research has documented that ABA influences the redistribution of nitrate within plant tissues[39]. Accordingly, we employed a comparable methodology to investigate this phenomenon in annual apple branches. The results revealed a significant reduction in nitrate content within the branches on the second day after the application of ABA spray treatment compared to the control. There was then a progressive decline in nitrate content over time (Fig. 7a). After nitrate is absorbed by the roots, it is transported to the shoots and back again via the NRT1.5 protein, which mediates this process in Arabidopsis[18]. NRT1.8 is also involved in the reallocation of nitrate[19]. ABA treatment significantly induced MdABF1 expression (Fig. 7b), while gradually downregulating MdNRT1.5 (Fig. 7c). Meanwhile, the expression of MdNRT1.8 increased by around 14-fold (Fig. 7d). The nitrate root/shoot ratio of apple seedlings was significantly higher after ABA treatment than the control group (Fig. 1e). Consequently, the expression profiles of MdNRT1.5 and MdNRT1.8 aligned with the patterns observed in the annual branches subjected to ABA treatment (Fig. 1k−l). MdNRT1.5 expression decreased following ABA treatment, regardless of nitrogen availability (Fig. 1k), whereas MdNRT1.8 expression increased (Fig. 1l). To further elucidate the molecular mechanism of ABA-regulated nitrate reallocation, we treated the MdARF1 overexpressing Arabidopsis under high and low nitrogen conditions with or without ABA. The fresh weight was lower in the MdABF1 transgenic lines than in the WT under ABA treatment, regardless of nitrate content (Fig. 8a−c, Supplementary Fig. S4a, b). Accordingly, the roots to shoots nitrate ratio in the overexpressed MdABF1 Arabidopsis was remarkably higher under low nitrate and ABA treatment than in the wild type (Fig. 8d, e, Supplementary Fig. S4c, d). Interestingly, the expression of NRT1.5 and NRT1.8 in MdABF1 overexpressing material treated with nitrate and ABA was consistent with the above expression pattern (NRT1.5 expression was downregulated, while NRT1.8 expression was upregulated) (Fig. 8f−m, Supplementary Fig. S4e−l). We speculated that MdABF1 might directly regulate the expression of MdNRT1.5 and MdNRT1.8. To test this speculation, we analyzed the promoters of MdNRT1.5 and MdNRT1.8. EMSA revealed specific binding of MdABF1 to the MdNRT1.8 promoter (Fig. 9a). This result was further confirmed by the Y1H assay (Fig. 9b). The dual luciferase and GUS staining assays demonstrated that MdABF1 transcriptionally activated MdNRT1.8 expression (Fig. 9c−e). The findings demonstrate that MdABF1 directly interacts with the promoter region of MdNRT1.8, thereby activating its transcription and affecting the return flow of nitrate from shoots to roots.
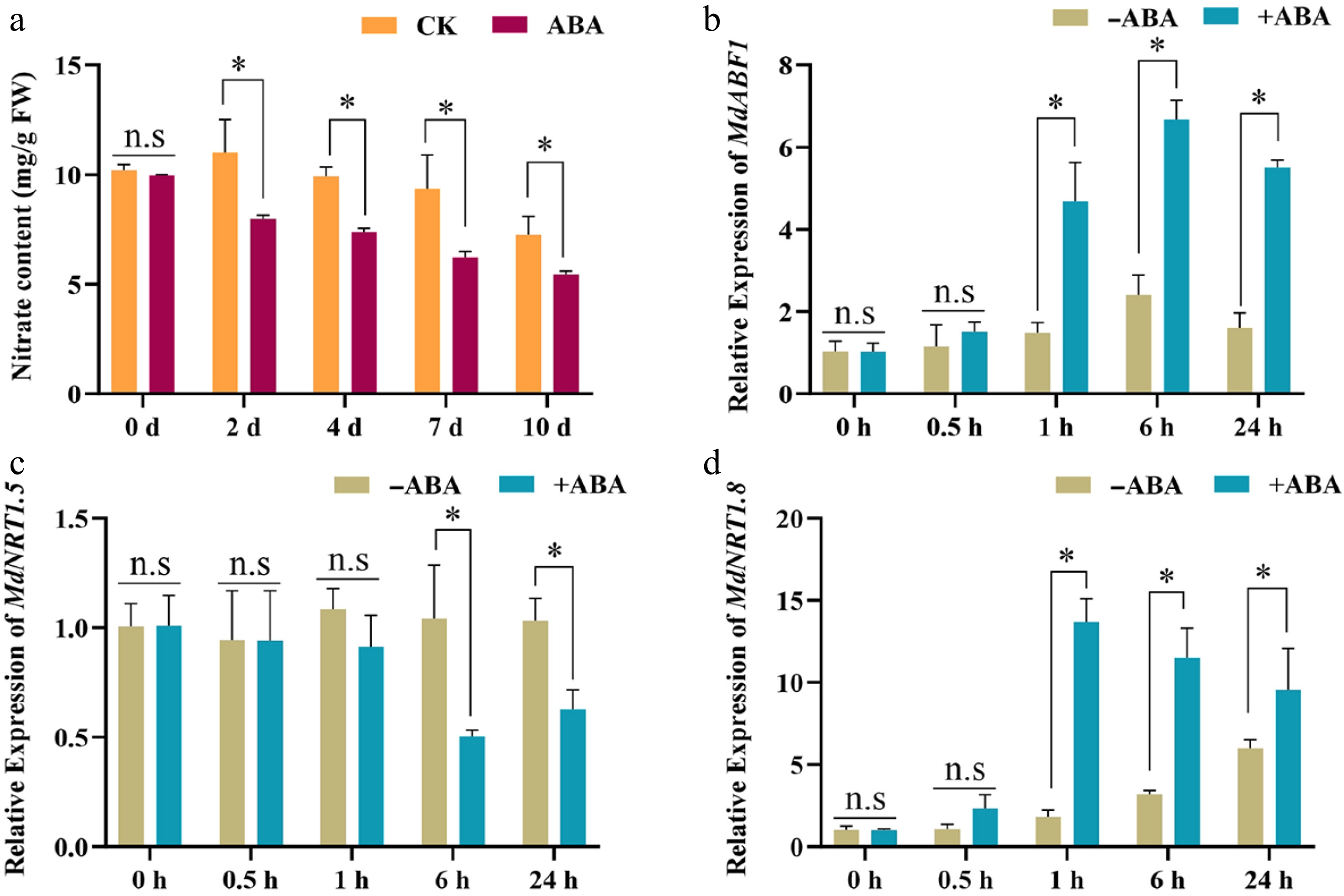
Figure 7.
ABA regulates nitrate allocation. (a) The nitrate content of apple branches. The samples were collected and analyzed for nitrate content treated by ABA for 2, 4, 7, and 10 d. (b)−(d) Expression analysis of MdABF1, MdNRT1.5, and MdNRT1.8 in apple branches treated by ABA. Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
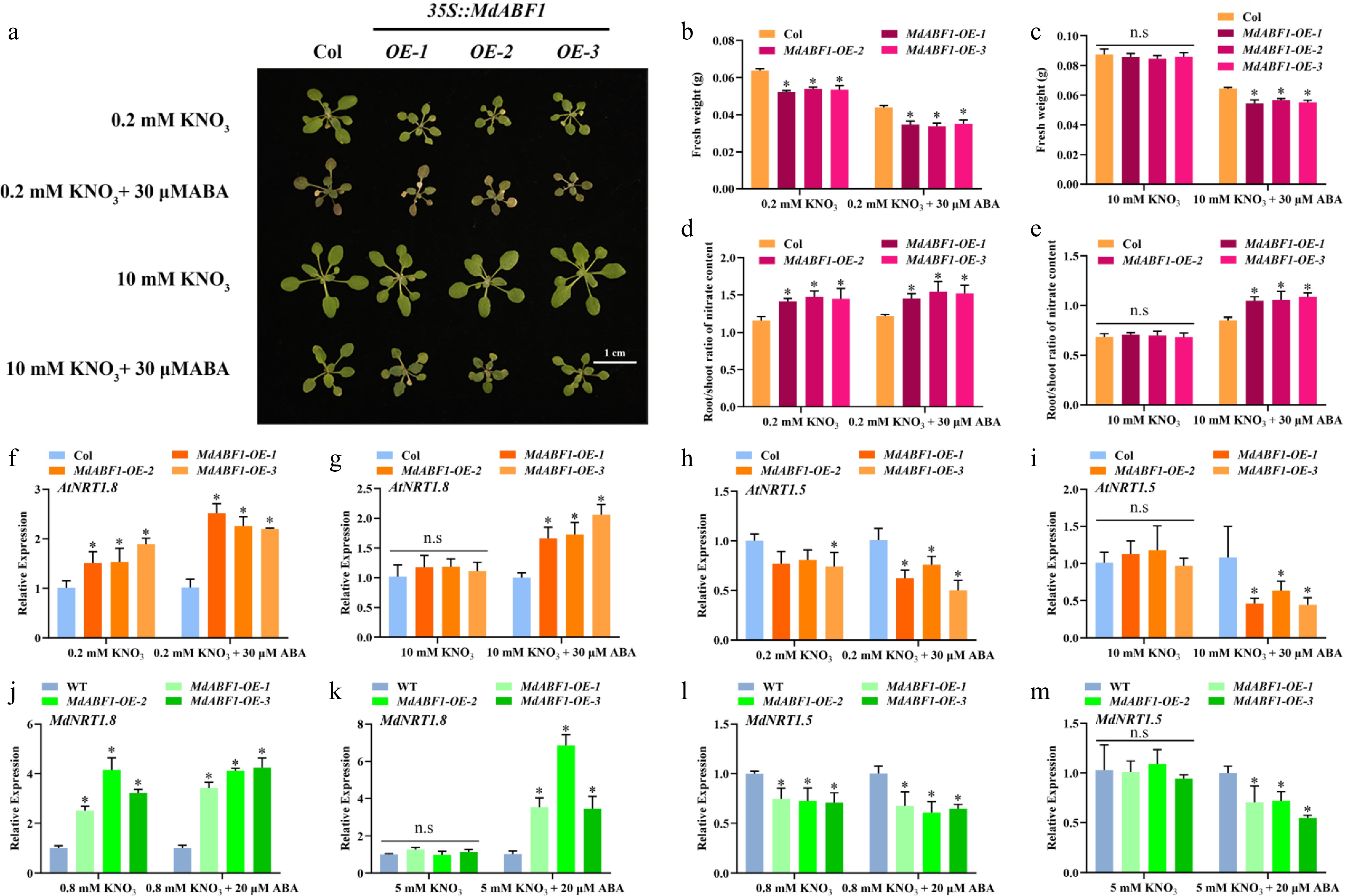
Figure 8.
MdABF1 plays a positive role in nitrate transport. (a) Phenotype of MdABF1 overexpression Arabidopsis treated with different concentrations of nitrate (0.2 or 10 mM KNO3), and ABA (0 or 30 μM). (b), (c) Fresh weight and (d), (e) root/shoot ratio of nitrate content in (a). Expression of (f), (g) AtNRT1.8 and (h), (i) AtNRT1.5 genes in Arabidopsis. Expression of (j), (k) AtNRT1.8 and (l), (m) AtNRT1.5 genes in apple calli. Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
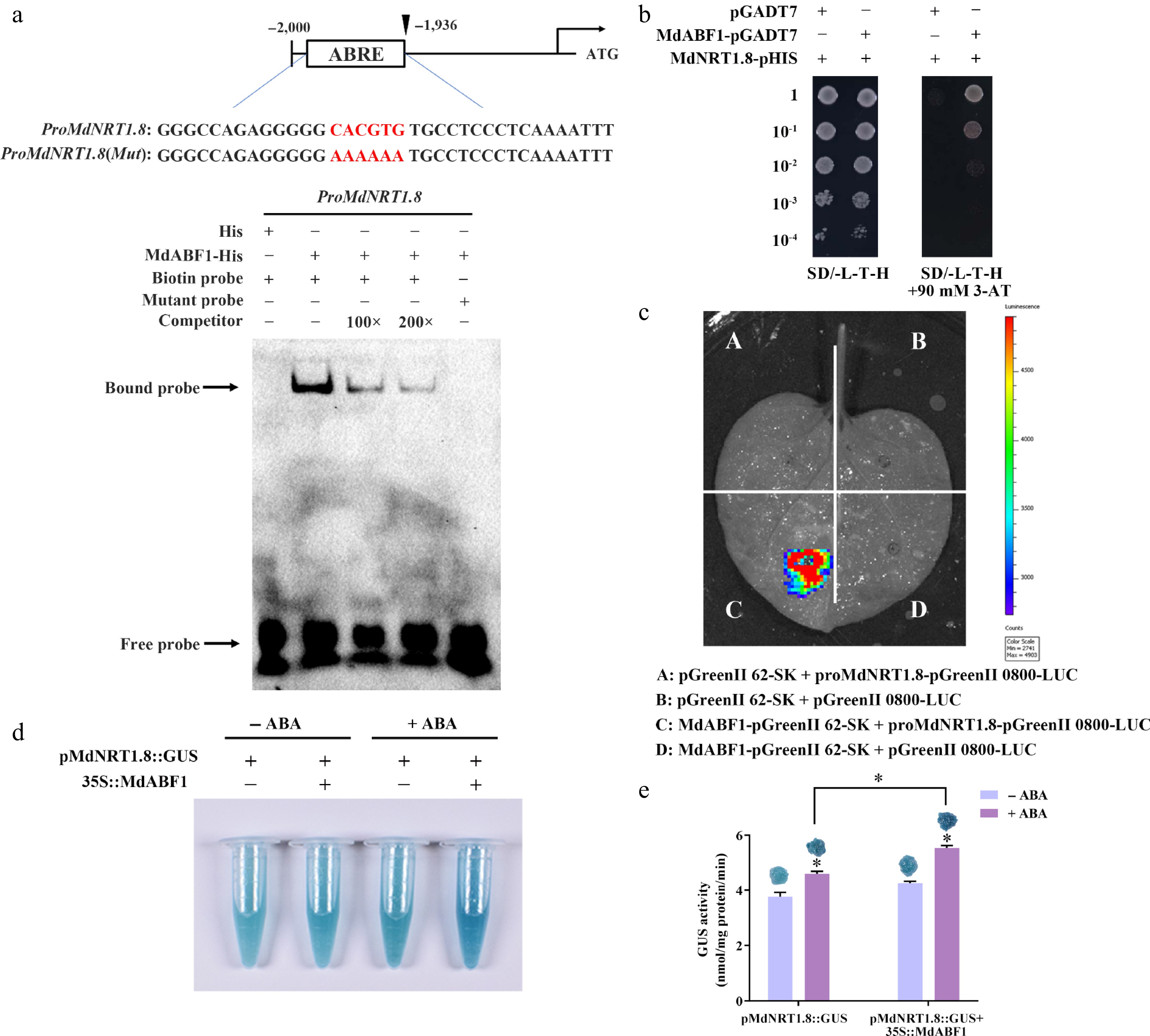
Figure 9.
MdABF1 binds directly to the promoter of MdNRT1.8 and activates its expression. (a) EMSA analysis showing MdABF1 binding to the MdNRT1.8 promoter. Diagram of the promoter of MdNRT1.8 are shown above. (b) Y1H assay of MdABF1 binding to the MdNRT1.8 promoter in vitro. (c) Dual luciferase assays in tobacco leaves showing that MdABF1 transcriptionally repressed the expression of MdNRT1.8. (d) GUS staining experiment of apple calli showing the repression of MdABF1 to the MdNRT2.3 promoter. (e) GUS activity in (d). Error bars exhibit the means ± SD of three independent replicates. Asterisks indicate significant differences (LSD test, *p < 0.05).
-
Efficient nitrogen utilization is indispensable for the realization of sustainable agricultural practices[7]. Compared to annual model plants like Arabidopsis and rice, the study of nitrogen utilization and its regulatory networks is markedly under-researched in perennial fruit trees[33]. In recent years, the interplay between hormone signaling and nitrate utilization has become clearer, with accumulating evidence suggesting that hormone signaling may influence plant growth and development via modulating the effects of nitrate use[39]. The NRT/NPF gene family is crucial for nitrate utilization and is involved in various hormone signaling pathways, including those of auxin, ethylene (Eth), jasmonic acid-isoleucine (JA-Ile), gibberellins (GAs), and ABA[6,52]. Among them, NPF6.3 regulates the growth of lateral roots by modulating auxin transport in response to different nitrate concentrations[9]. Conversely, AtNPF3 is a nitrate- and nitrite-specific transporter protein that governs the sequestration of nitrite within the foliage[53]. Later studies identified NPF3 as a GA transporter protein, as it affects GA accumulation in the root endodermis, and it functions as a crosstalk regulator of the crosstalk between GA and nitrite during hypocotyl[54]. Analysis of the npf2.1 mutant revealed that NPF2.10 plays a role in the translocation of JA/JA-Ile from damaged to undamaged leaves during damage response. The responses to nitrate are of great interest, including those relating to lateral root length and gene expression (NRT2.1 and NRT1.1), which are strongly affected in Ethylene Insensitive2 (EIN2) and Ethylene Receptor1 (ETR1) mutants[52]. The study provided evidence for an association between nitrate signaling and the biosynthetic and response pathways of the plant hormones auxin, GA, Eth, and JA-Ile.
ABA is closely related to nitrate. Previous research has demonstrated that knocking down TabZIP60, which is a member of the bZIP family gene closely related to the AtABF gene family, leads to a significant increase in nitrogen uptake and yield[55]. The result of overexpressing TabZIP6 was the opposite[55]. Furthermore, the npf4.6 mutant exhibits a more sensitive response to nitrate-mediated seed germination[56]. Analysis of the nitrate transcriptome using ChIP-seq and TARGET revealed that nitrate response genes and their associated biological processes were regulated in a coordinated manner that was both spatial and temporal, highlighting a key hub of nitrate regulation[45]. The gene regulatory network (GRN) model was used to determine that the endodermis serves as the regulatory hub for the nitrate response. Furthermore, ABF2 and ABF3, which are significant genes in the ABA pathway, were identified as transcription factors that played crucial roles in regulating the nitrate response within the endodermis[45]. Multiple investigations have consistently indicated a close relationship between ABA signaling and nitrate, highlighting the need for a detailed exploration of the precise regulatory network. In this study, we identified the ABA-regulated regulatory module of MdABF1-MdNRTs/NIAs (Fig. 10), which targets genes in the presence of ABA, including the repression of MdNRT2.1, MdNRT2.3, MdNIA1, MdNIA2, MdNRT1.5 expression, as well as the promotion of MdNRT1.8 function. MdNRT2.1 and MdNRT2.3 are involved in nitrate absorption, MdNIA1 and MdNIA2 are responsible for nitrate assimilation, and MdNRT1.8 acts as a positive regulator of nitrate reallocation. In contrast, MdNRT1.5 has an inhibitory effect. Ultimately, the inhibitory effect of MdABF1 was attributed to the suppression of nitrate absorption and assimilation, accompanied by the promotion of nitrate retrieval, resulting in impaired utilization and growth (Fig. 9).
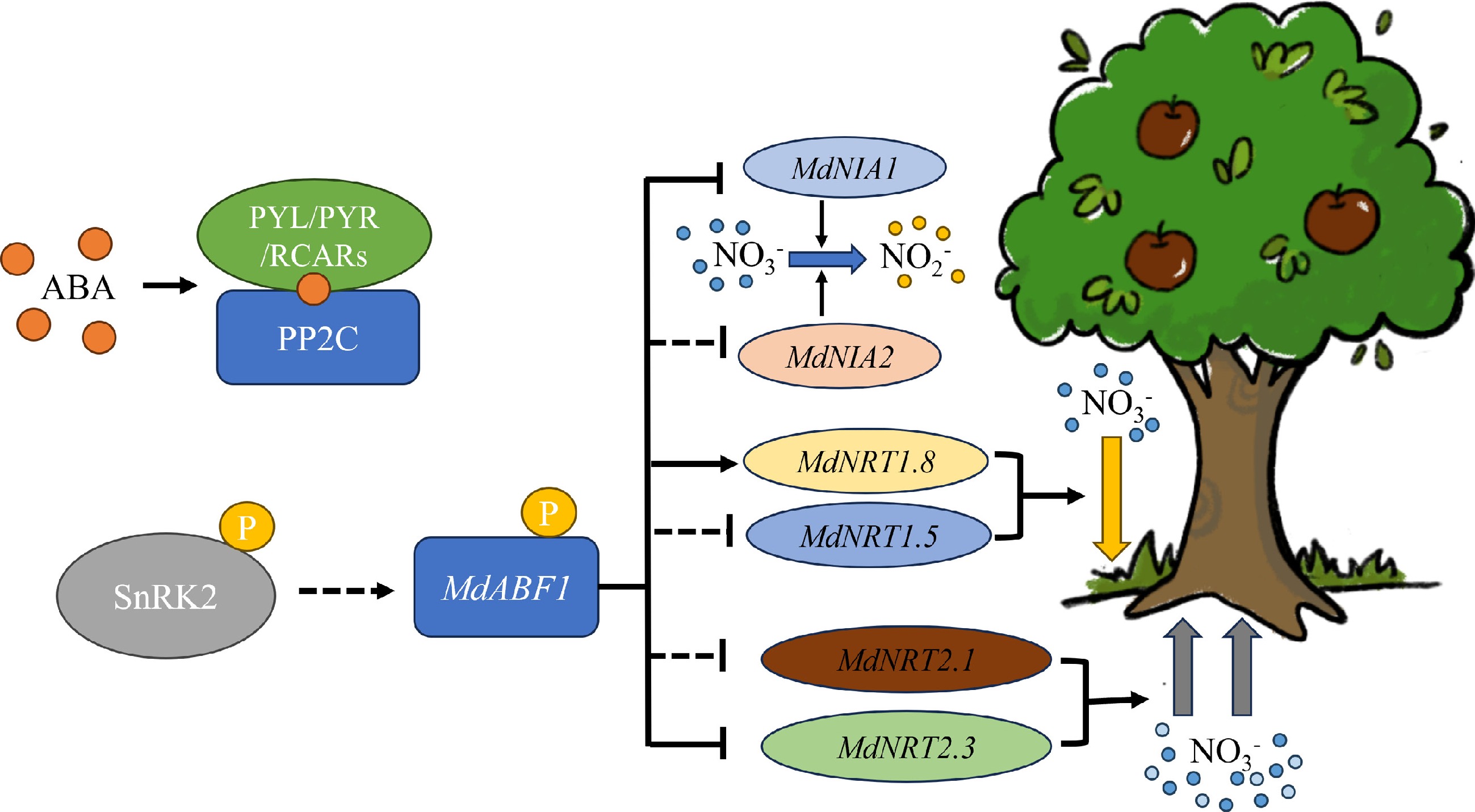
Figure 10.
Working model of an ABA mediated MdABF1-MdNRTs/NIAs module in regulating nitrate utilization in apple. In the presence of nitrate, ABA-mediated MdABF1 can be activated. This occurrence has a direct negative moderator effect on MdNIA1 and MdNRT2.3, and a positive moderator effect on MdNRT1.8. At the same time MdABF1 has an indirect regulatory effect on MdNIA2, MdNRT1.5, and MdNRT2.1. Nitrate absorption was inhibited by downregulation of MdNRT2.1 and MdNRT2.3. The down regulation of the MdNIA1 and MdNIA2 transcripts inhibited the process of nitrate assimilation. In contrast, increasing the level of MdNRT1.8 and decreasing the level of MdNRT1.5 favored the reallocation of nitrate to the root system.
Several NRT/NPF members play a role in the absorption of nitrate in both Arabidopsis and rice[2]. NRT1.1/NPF6.3 was the first protein identified in higher plants that is involved in nitrate absorption, which exhibits both high- and low-affinity nitrate absorption[23]. Unlike NPF6.3, the transporter protein NRT1.2/NPF4.6 is constitutively expressed and functions as a low-affinity nitrate transporter[9,56]. However, some of the NRT2s exhibit high-affinity nitrate transport activity. Well-documented evidence shows that NRT2.1 and NRT2.2 are important genes that are responsible for the high-affinity absorption of nitrate in Arabidopsis[57]. NRT2.4 and NRT2.5 also participate in high-affinity nitrate absorption, but only when nitrogen is deficient[17,58]. Studies on rice have shown that NRT2.1, NRT2.2, and NRT2.3 play a role in high-affinity nitrate absorption[38]. The OsNRT2.3 gene generates two transcripts, OsNRT2.3a and OsNRT2.3b, by alternative splicing. High expression of OsNRT2.3b improves the pH buffering capacity of rice and increases the absorption of N, P, and Fe[14]. In Arabidopsis, NIA1 and NIA2 encode nitrate reductases that are involved in multiple hormone signaling pathways[59]. Our data support the ABA-mediated inhibition of nitrate assimilation and absorption by MdABF1 in several ways. (1) When ABA was given at high nitrate concentrations, the biomass of apple seedlings decreased significantly, as did their fresh weight. This was also demonstrated at low nitrate concentrations (Fig. 1a, b). MdABF1 was overexpressed in Arabidopsis and apple calli, and the biomass and fresh weight of the plants decreased significantly after the low nitrate and ABA treatment at low and high nitrate levels (Fig. 2a, d, e, Supplementary Fig. S2c, d, Fig. 3a−c, Supplementary Fig. S2i, j). (2) In nitrate- and ABA-treated apple, the nitrate content, and NR activity decreased significantly (Fig. 1c, d), and the levels of both decreased significantly in MdABF1-overexpressing Arabidopsis and apple calli (Fig. 2f−i, Supplementary Fig. S2e−h, Fig. 3d−g, Supplementary Fig. S2k−n). These results suggested that MdABF1 inhibits nitrate absorption and assimilation when ABA and low nitrate levels are present. A previous study showed that ABA signaling is recruited to inhibit the energy-demanding nitrogen assimilation pathway, if the plant is under drought stress[52]. (3) Genes related to nitrate absorption and assimilation were repressed under ABA treatment (Fig. 1f−n). The expression of genes related to nitrate absorption and assimilation was also repressed in MdABF1-overexpressing plants (Fig. 4 and Supplementary Fig. S3). We demonstrated that MdABF1 binds directly to the promoters of MdNIA1 and MdNRT2.3 genes, thereby repressing their expression (Figs 5 and 6). In a previous study on wheat, the ABF-like gene TabZIP60 was found to play a negative regulatory role in nitrate utilization[55]. This function was similar to that of MdABF1, which is the subject of the present study.
Once nitrate has been absorbed into the root cells, it is either stored in the root or transported to the shoot[2]. Of the two main genes identified as being associated with long-distance transport, NRT1.5 mediates loading xylem with nitrate, whereas NRT1.8 is responsible for unloading it[18,19]. The process of reallocating nitrate to the roots usually occurs under stressed conditions[39]. This study found that MdABF1 was involved in the reallocation process under ABA-mediated conditions. (1) ABA treatment significantly enhanced nitrate allocation to roots relative to shoots in apple seedlings (Fig. 1e). A similar redistribution pattern was observed in Arabidopsis plants overexpressing MdABF1 (Fig. 8d, e). A previous investigation also demonstrated that ABA signaling is involved in nitrate retrieval[39]. These results revealed that the MdABF1 gene was also involved in nitrate retrieval. (2) The transcriptional level of nitrate assimilation genes was detected in apple seedlings treated with low levels of nitrate and ABA. MdNRT1.5 expression was downregulated, while MdNRT1.8 expression was significantly upregulated (Fig. 1i−l, Supplementary Fig. S4i−l). The expression patterns of these two genes were similar in Arabidopsis and apple calli (Fig. 8f−l, Supplementary Fig. S4e−l). Further experiments revealed that MdABF1 bound directly to the MdNRT1.8 promoter and activated its transcription (Fig. 9). As ABA stimulates the production of JA, the two are synergistically involved in biological processes[60]. Some genetic analyses have shown that cadmium and sodium stress trigger the Eth/JA signaling pathway, which accumulates EIL1 to regulate ERF expression and subsequently increase NRT1.8 expression[35]. In contrast, the downregulation of NRT1.5 mediated by Eth and JA signaling through EIN3/EIL1 and other unknown components[31]. NRT1.8 and NRT1.5 are both synergistically regulated by ABA[35]. Thus, we conclude that ABA-regulated MdABF1 promotes the nitrate retrieval process by promoting the expression of MdNRT1.8.
In conclusion, this article shows that MdABF1 plays a negative role in nitrate absorption and assimilation by negatively regulating MdNRT2.3 and MdNIA1, while also activating MdNRT1.8, which positively regulates nitrate allocation. MdABF1 was also found to negatively regulate MdNRT2.1, MdNIA1, and MdNRT1.5. Overall, this study unveils the regulatory module for nitrate utilization in perennial apple trees, offering new insights into the molecular mechanisms through which ABA regulates nitrate utilization.
-
Apples are one of the most popular and widely consumed fruits world wide. Belonging to the species Malus domestica in the rose family, they have been cultivated in Asia and Europe for thousands of years and were later brought to North America by European colonists. As an essential substance for plant growth and development, nitrate plays a crucial role throughout the entire lifecycle. However, research into the relationship between hormones and nitrogen is limited. This paper investigates the connection between ABA and nitrate. Our results demonstrated that ABA-regulated MdABF1 suppressed nitrate absorption and assimilation through directly downregulating MdNIA1 and MdNRT2.3 expression, but activated the expression of the MdNRT1.8 gene to promote nitrate retrieval. These results elucidate the molecular regulatory module of MdABF1-MdNRTs/NIAs and provide groundwork for understanding how ABA regulates nitrogen utilization.
This work was supported by the National Natural Science Foundation of China (32272683), the Natural Science Foundation of Shandong Province (ZR2024JQ036), the China Postdoctoral Science Foundation (2024M761850), and the China Agriculture Research System of MOF and MARA (CARS-27).
-
The authors confirm their contributions to the paper as follows: study conception and design: Zhang T, Wang X; experiment and technical assistance: Rui L, Liu G; data collation and analysis: Rui L, Wang X, Li H, Yang Y; sample culturing and collection: Rui L, Liu R, Li H, Liu G; manuscript writing and revising: Rui L, Zhang T, Wang X. All authors read and approved the final manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used in the article.
- Supplementary Fig. S1 Phylogenetic tree analysis of ABF family, the expression of MdABF1 in apple treated by nitrate and identification of transgenic material.
- Supplementary Fig. S2 Overexpression of MdABF1 negatively regulates nitrate absorption and assimilation genes in Arabidopsis and apple calli under ABA treatment.
- Supplementary Fig. S3 Relative expression of NRT2s and NIAs in transgenic plants of MdABF1 treated by different concentrations of nitrate and ABA.
- Supplementary Fig. S4 MdABF1 plays a positive role in nitrate retrieval.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Rui L, Wang X, Liu G, Li H, Yang Y, et al. 2025. ABF1-MdNRTs/NIAs module mediates ABA-regulated nitrate utilization in apple. Fruit Research 5: e035 doi: 10.48130/frures-0025-0026 

ABF1-MdNRTs/NIAs module mediates ABA-regulated nitrate utilization in apple
- Received: 12 June 2025
- Revised: 10 July 2025
- Accepted: 18 July 2025
- Published online: 12 September 2025
Abstract: Apples are a globally prominent fruit, valued for their texture, flavor, and nutritional value. Apple trees rely heavily on nitrogen, a vital nutrient essential their growth and development. However, the low absorption of nitrogen fertilizers is a significant agricultural challenge, prompting research into improving nitrogen use efficiency. Nitrate is a key form of nitrogen that serves as both a nutrient and a signaling molecule, influencing plant processes. Although hormones such as abscisic acid (ABA) participate in nitrate signaling, the specific mechanisms in apples remain unclear. In this work, we discovered that ABA-regulated MdABF1 negatively regulated nitrate absorption and assimilation by directly downregulating the expression of the MdNIA1 and MdNRT2.3 genes, but activated the expression of MdNRT1.8 to promote nitrate retrieval. These results illustrated the molecular regulatory network through which ABA regulated the utilization of nitrate by the MdABF1-MdNRTs/NIAs module. Taken together, our study offers novel insights into how ABA orchestrates nitrate utilization in apple plants.
-
Key words:
- MdABF1 /
- ABA /
- Nitrate /
- Absorption and assimilation /
- Apple