









-
Wampee (Clausena lansium) is a valuable tropical-subtropical evergreen fruit tree belonging to the Rutaceae family, primarily cultivated in South China and Southeast Asia. Highly regarded for its nutritional and medicinal properties, almost every part of the plant—including its leaves, stems, roots, and fruits—contains abundant bioactive compounds[1]. Due to this, wampee has been widely applied in traditional Chinese medicine to treat various ailments, such as cough, asthma, and allergies[2].
Soluble sugars serve as the critical determinants of fruit quality by modulating both nutritional composition and sensory attributes. In the majority of fruit species, sucrose metabolism functions as the central regulatory mechanism for sugar homeostasis, orchestrating cellular sugar utilization, storage, and equilibrium[3]. This metabolic process relies heavily on sugar transporter proteins, which facilitate the bidirectional translocation of sugars across membranes during loading and unloading phases[4−6]. Notably, SWEET proteins (Sugar Will Eventually Be Exported Transporters) represent a unique class of transporters characterized by their capacity for bidirectional sugar movement.
Structurally, SWEET proteins in higher plants consist of seven transmembrane α-helices structural domains (TMs), organized into a 3-1-3 symmetric arrangement: two conserved triple-helix domains (N- and C-termini) connected by a variable transmembrane helix[7,8]. Phylogenetically, SWEETs are classified into four clades with distinct substrate preferences: the first clade (AtSWEET1−3) and the second clade (AtSWEET4−8) mainly mediate the transport of hexoses, especially glucose; the third clade (AtSWEET9−15) preferentially transports sucrose across membranes, the fourth clade (AtSWEET16−17) primarily transports fructose[4,9−11]. Functionally, SWEETs are pivotal for fruit sugar accumulation during fruit development. For instance, VvSWEET10 in grapes exhibits peak expression during ripening, and its overexpression significantly elevates glucose, fructose, and total sugar levels[12]. Beyond transporters, transcription factors (TFs) also critically regulate sugar metabolism. ClNAC68 in watermelon can suppress ClINV expression to modulate invertase activity and sugar content[13]. HpWRKY3 in pitaya could directly regulate HpINV2 and HpSuSy1 expression to control the fruit sugar accumulation[14]. In citrus, CitERF16 could activate the expression of CitSWEET11d and enhance the accumulation of sucrose in fruits[15]. These findings underscore the coordinated roles of SWEET transporters and TFs in fruit sugar regulation, offering potential targets for quality improvement in horticultural crops.
Current research on sugar accumulation in wampee fruits has made significant advances in elucidating its physiological basis, while molecular-level investigations are now gaining momentum. A significant advancement was made by Wei et al.[16], who successfully assembled the chromosome-level reference genome of the 'Jinfeng' wampee cultivar. Building on this genomic resource, Chen et al. identified two key regulatory genes, CIERF061 and CISWEET7, whose expression patterns showed strong correlation with sugar and acid accumulation patterns[17]. Their work further demonstrated that CIERF061 functions as a transcriptional activator of the CISWEET7 promoter. Despite these important findings, the comprehensive molecular mechanisms underlying sugar accumulation in wampee remain largely unexplored. Further elucidation of these mechanisms will provide crucial insights for targeted quality improvement and precision breeding strategies in this economically important fruit crop.
-
The wampee cultivars used in this study included the sweet-fleshed 'Huami' (HM) and sweet-sour 'Jinjixin' (JJX), cultivated in the experimental orchard of South China Agricultural University, Guangdong Province, China. Fruits were sampled at five developmental stages (71, 78, 85, 92, and 99 d after bloom (DAB)) corresponding to distinct physiological changes (Fig. 1). At each sampling point, 15 fruits of uniform size, consistent external appearance, and free from pest damage and disease symptoms were collected as one biological replicate. Three independent biological replicates were obtained for each time point to ensure statistical reliability. After sample collection, the fruits were immediately transported to the laboratory in a low-temperature container. Following brief cleaning and surface drying, the flesh was separated, flash-frozen in liquid nitrogen, and stored at −80 °C in a Haier ultra-low temperature freezer (model DW-86L626, Qingdao, China) for soluble sugar content determination, sucrose-metabolism-associated enzyme activity analysis, transcriptome sequencing, and gene expression analysis.

Figure 1.
Wampee fruits at different developmental stages and cross-sectional view of fruits. (a) 'Huami' wampee. (b) 'Jinjixin' wampee. d stands for days after bloom. Bar = 1 cm.
The virus-induced gene silencing (VIGS) in wampee fruits was conducted using JJX wampee plants cultivated at the experimental base of South China Agricultural University (80 DAB). Fruits with uniform growth status and free from pests and diseases were selected, and the pulp was separated for the experiment.
For the subcellular localization and dual-luciferase reporter assays, Nicotiana benthamiana plants were grown in a plant growth chamber maintained at 22 °C with a 15 h/9 h (light/dark) photoperiod and relative humidity of 70% ± 5%. The seeds were obtained from our laboratory stock.
Extraction and content measurement of soluble sugar and starch
-
The extraction and content measurement of sucrose, glucose, and fructose was carried out using HPLC (High Performance Liquid Chromatography), referring to the method of Hu et al.[18]. The extraction and measurement of total soluble sugars and starch were carried out with reference to the procedures as previously described by Li et al.[19].
Transcriptome sequencing of wampee
-
The sequencing service was provided by Novogene Co., Ltd. (Beijing, China). RNA integrity and quantity were precisely assessed using an Agilent 2100 Bioanalyzer. Non-specific library construction was performed, with a read length of 150 bp paired-end. The screening criteria for differentially expressed genes were |log2FC| ≥ 1 and padj ≤ 0.05. Qualified libraries were subjected to sequencing on the Illumina NovaSeq 6000 platform with different index tags.
The image data generated by high-throughput sequencing were base-called using CASAVA (Illumina Casava 1.8) to produce raw reads. Data preprocessing included removal of adapter-containing reads, reads with ambiguous bases (N), and low-quality reads (where > 50% of bases had Qphred ≤ 20), resulting in clean reads. These clean reads were then aligned to the reference genome of 'Jixin' wampee (determined by our laboratory) using HISAT2 (v2.0.5). Gene expression levels were quantified using featureCounts (v1.5.0-p3) to count reads mapped to each gene. Gene expression was normalized and expressed as FPKM (Fragments Per Kilobase of transcript per Million mapped reads), calculated based on gene length and the number of reads mapped to each gene.
Bioinformatics analysis
-
Analysis of transcriptome data. Differential expression analysis between the two comparison groups was carried out using DESeq2 software (1.20.0). Statistical enrichment of differentially expressed genes in the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway was analyzed using clusterProfiler (3.8.1) software. With the differential genes shared between HM and JJX at 78 DAB and 99 DAB as data sources, Weighted correlation network analysis (WGCNA) on the screened differential genes was conducted using the Novogene.
The phylogenetic tree was constructed using the Maximum Likelihood (ML) technique in MEGA-X software with 1,000 bootstrap replicates[20]. Get the phylogenetic tree's amino acid sequences for the Arabidopsis thaliana SWEET family from the TAIR website (
www.arabidopsis.org ). Analysis of conserved structural regions was conducted using the SMART web application (https://smart.embl.de/ ).DNA extraction
-
Sample genomic DNA was extracted utilizing the Plant DNA Extraction Kit (Aidlab, Beijing, China), following the instruction manual.
Total RNA extraction and reverse transcription
-
The total RNA of the samples was extracted using the Plant RNA Extraction Kit (Vazyme, Nanjing, China). Total RNA was reverse transcribed to cDNA using a cDNA Reverse Transcription Kit (TransGen Biotech, Beijing, China), referring to the instruction manual for specific steps.
Quantitative Real-Time PCR (qRT-PCR)
-
The Primer3 Input (website:
http://primer3.ut.ee ) was utilized to design qRT-PCR primers for the target genes. Primers were synthesized by Shanghai Generay Biotech Co., Ltd. (China) (Supplementary Table S1). The reaction system was prepared with RealUniversal Color PreMix (SYBR Green) reagent (TRANSGen, Beijing, China). Reaction programs were set up, and qRT-PCR reactions were performed on a Quantitative Real-Time PCR instrument (C1000 Touch Thermal Cycler, USA). Wampee Actin (GenBank accession number: MW532092) was used as internal reference genes, and the results were analyzed by the 2−ΔΔCᴛ method. Three replicates per sample.Gene cloning
-
Primer Premier5 software was used to create the primer based on the coding sequences of target genes found in the genome of 'Jixin' wampee. Primers were synthesized by Shanghai Generay Biotech Co., Ltd. China (Supplementary Table S1). Utilization of High-Fidelity cloning enzyme 2×Phanta® Max Master Mix (Dye Plus) (Vazyme, Nanjing, China), the cDNA of the 'Jinjixin' wampee pulp was used as a template for amplification. All PCR products were then electrophoresed in a 1.2% agarose gel to obtain the target genes. The target genes were ligated, transformed, and identified, and then sent to Guangzhou Aiki Biotechnology Co., Ltd., China. Sequencing results were compared with the 'Jixin' wampee genome sequence.
Subcellular localization analysis
-
The coding regions of the ClSWEET3a (without stop codons) with Hind III and BamH I restriction sites were cloned into the pC18-35S:GFP vector. The primers are listed in Supplementary Table S1. The recombinants were induced into Agrobacterium tumefaciens strain GV3101 (pSoup-p19) and used for transient expression in N. benthamiana. The infected N. benthamiana leaves were examined 48 h after infiltration. A fluorescence microscope (ZEISS LCM-800, Oberkochen, Germany) was used to capture the GFP signal. All assays were repeated three times.
Yeast functional complementation experiment
-
With reference to the method described by Feng[21], the yeast expression vector for the candidate gene (pDR195-ClSWEET3a) was constructed and transfected into the hexose transporter-deficient yeast EBY.VW4000. SC/-Ura + 2% Mal, SC/-Ura + 2% Glu, SC/-Ura + 2% Fru, and SC/-Ura + 2% Suc were cultured for 3−5 d on SD-ura selection medium containing a sole carbon source, respectively, and the growth of the yeasts was observed.
Validation of wampee VIGS
-
According to the method described by Xie et al.[22], the vectors pTRV1 and pTRV2 were selected and pTRV2 was double digested using BamH I and Sma I restriction endonucleases. The vector restriction sites in the ClSWEET3a primers were created based on the cleavage site, the target fragments amplified, and the recombinant plasmids constructed. Fruits of JJX at 80 DAB were selected, the bacterial fluids were injected from the top of the fruit to the middle of the fruit using a 1 mL syringe with a needle, and the fruits were covered with black plastic bags after the injection was completed. The plastic bags were removed after one day of dark culture and cultured for six days under normal conditions. Fruits that were still growing normally were picked and immediately placed in a low-temperature box to return to the laboratory. The pulp of the fruit samples was separated in the laboratory, frozen with liquid nitrogen, and kept in a refrigerator at −80 °C to facilitate the analysis of gene expression and the measurement of soluble sugar content.
Dual luciferase reporter assay
-
Referring to the approach of Cheng et al.[23] the pGreen II 62-SK vector was double digested with EcoR I and BamH I restriction endonucleases, candidate transcription factor primers were designed based on the enzyme cleavage sites, and the pGreen II 62-SK vector was constructed to serve as Effector. The pGreen II 0800-LUC vector was double-enzymatically cleaved with KpnI and NcoI restriction endonucleases, primers for ClSWEET3a promoter with vector cleavage site were designed according to the cleavage site, and the promoter of ClSWEET3a was ligated into the cleaved pGreen II 0800-LUC vector as Reporter. After transforming Agrobacterium, it was injected into N. benthamiana. The LUC/REN ratio was computed by measuring the fluorescence values of Firefly luciferase (LUC) and Renilla luciferase (REN).
Yeast one-hybrid assay
-
Referring to the method of Hu et al.[24], the pAbAi vector was double-enzymatically cleaved with Sac I and Xho I restriction endonucleases. ClSWEET3a was divided into three segments (neither BstB I nor Bbs I cleavage sites can be present in the promoter fragment), primers of promoter fragments with vector cleavage site were designed on the basis of the cleavage site, the target fragment was amplified using the cloned ClSWEET3a-pGreen II 0800-LUC template, and then ligated to the digested pAbAi vector. The pGADT7(AD) vector was double-enzymatically cleaved with EcoR I and BamH I restriction endonucleases, and primers for candidate transcription factors with vector cleavage sites were designed according to the cleavage sites. The target fragment was amplified and ligated with the cleaved pGADT7(AD) vector. The result was then co-transfected into yeast strain Y1HGold (with the AD empty vector as a control). The binding and activation capacity of transcription factors with the ClSWEET3a promoter was determined based on yeast growth at specific AbA concentrations. This was repeated in triplicate.
Statistical analysis
-
The data was processed using Excel 2010 software, and all data was statistically analyzed using IBM SPSS Statistics 25 (IBM, California, USA). The data analysis was conducted using a t-test and one-way analysis of variance (ANOVA), if p < 0.05, the result was considered to be significantly different. Graphing with GraphPad Prism 8.3 software (GraphPad Software, Inc).
-
The results demonstrated significant variations in sugar composition across different developmental stages within each cultivar (Fig. 2). During fruit development in wampee cultivars 'Huami' (HM) and 'Jinjixin' (JJX), sucrose content increased progressively, while hexose profiles showed cultivar-specific patterns. HM maintained a steady glucose increase, whereas JJX exhibited a transient decline at 85 DAB before rebounding. Fructose peaked at 85 DAB in HM but accumulated steadily in JJX thereafter. Notably, a metabolic shift occurred during ripening: hexoses dominated early development; however, the sucrose became predominant in mature fruits due to its accelerated accumulation coupled with slowed hexose accumulation. These differential sugar accumulation patterns between the sweet (HM) and sweet-sour (JJX) cultivars suggest distinct genetic regulation of sugar metabolism pathways during fruit ripening.
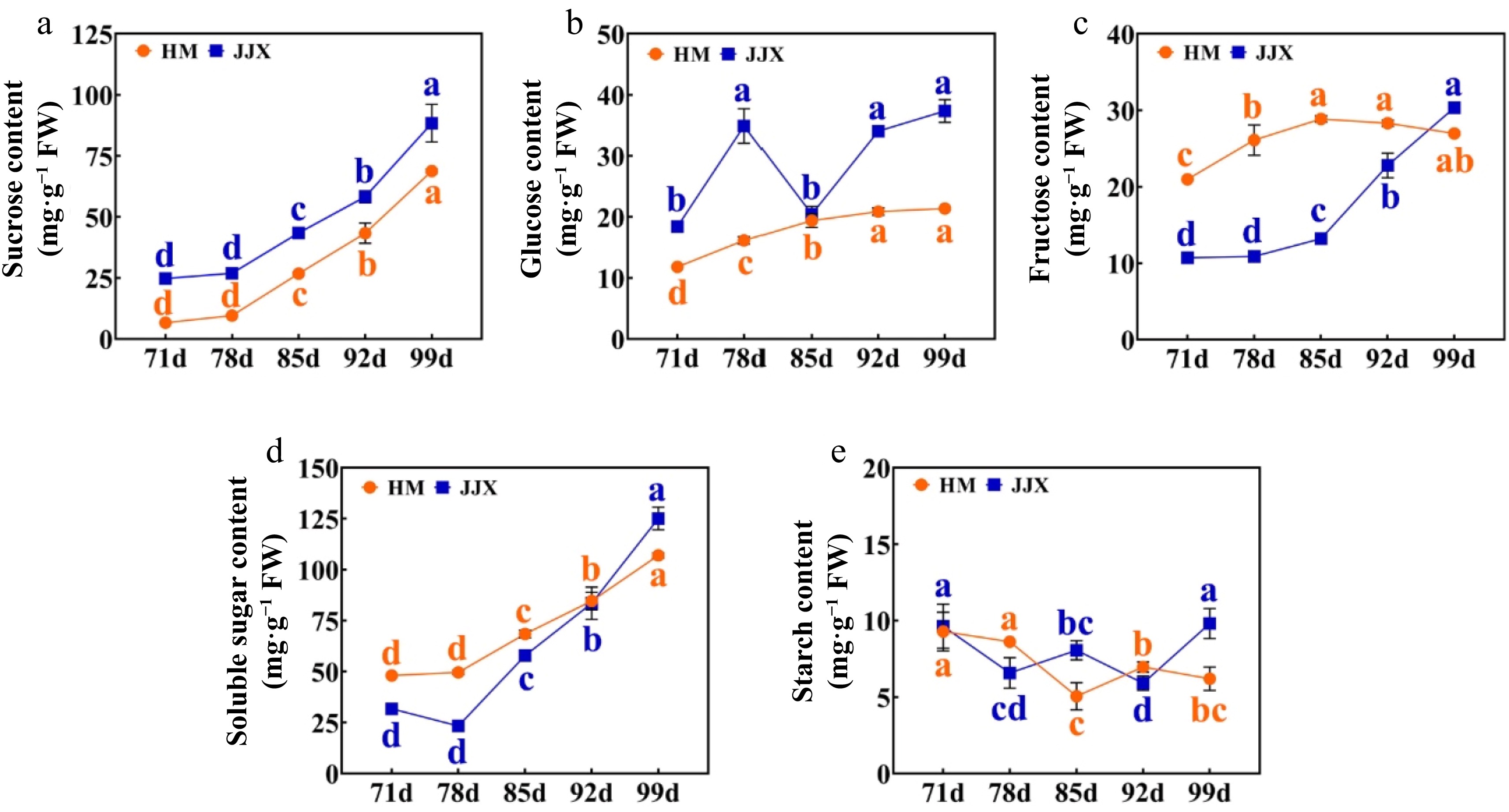
Figure 2.
Changes in sugar content of fruit during the development of wampee fruits. (a) Sucrose content. (b) Glucose content. (c) Fructose content. (d) Soluble sugar. (e) Starch content. HM stands for 'Huami' wampee; JJX stands for 'Jinjixin' wampee. d stands for days after bloom. Error bars represent the standard deviation of three biological replicates. Different lowercase letters represent the level of significance at p < 0.05 (one-way ANOVA with Waller-Duncan). Comparison was conducted at different stages of each cultivar.
Statistical analysis revealed a highly significant correlation between total soluble sugar and sucrose content in both HM and JJX fruits (Table 1). The rapid sucrose accumulation during late developmental stages and its predominance in mature fruits suggest its crucial role in determining final sugar composition during wampee fruit development.
Table 1. Correlation analysis of total soluble sugars with sugar component content in HM and JJX.
Varieties Categories Soluble sugar Sucrose Glucose Fructose HM Soluble sugar 1 0.999** 0.857* 0.549 Sucrose 0.999** 1 0.849* 0.535 Glucose 0.857* 0.849* 1 0.898* Fructose 0.549 0.535 0.898* 1 JJX Soluble sugar 1 0.993** 0.520 0.977** Sucrose 0.993** 1 0.592 0.979** Glucose 0.520 0.592 1 0.661 Fructose 0.977** 0.979** 0.661 1 HM stands for 'Huami' wampee; JJX stands for 'Jinjixin' wampee. * significant difference (p < 0.05); ** extremely significant difference (p < 0.01); t-test. Transcriptome sequencing analysis
-
Transcriptome sequencing was performed on HM and JJX fruits at three key developmental stages (78, 92, and 99 DAB) to exhibit significant sugar content variations (Supplementary Table S2). Based on the most pronounced differences in total soluble sugar and sucrose content between 78 DAB and 99 DAB in both cultivars, our bioinformatic analysis was focused on differentially expressed genes (DEGs) during this critical period. Comparative transcriptome analysis revealed substantial gene expression changes between 78 DAB and 99 DAB (Supplementary Fig. S1). In HM fruits, 3,631 DEGs were identified, comprising 1,555 upregulated and 2,076 downregulated genes. JJX fruits showed even more extensive transcriptional reprogramming, with 4,312 DEGs (2,068 upregulated and 2,244 downregulated). KEGG pathway enrichment analysis of these DEGs highlighted significant changes in starch and sucrose metabolism pathways. By cross-referencing with the 'Jixin' wampee genome, DEGs showing higher expression at 99 DAB compared to 78 DAB were specifically annotated (Supplementary Table S3).
Notably, 10 conserved DEGs shared between both cultivars were discovered, including: three 6-trehalose-phosphate phosphatase genes, one sucrose phosphate synthase gene, one starch synthase gene, and five genes with currently unknown functions.
These findings provide valuable insights into the transcriptional regulation of sugar metabolism during wampee fruit development and identify potential candidate genes for further functional characterization.
Weighted gene co-expression network analysis (WGCNA) was performed using the shared differentially expressed genes between the two critical developmental stages (78 vs 99 DAB) in both HM and JJX cultivars. The analysis revealed 2,103 conserved differentially expressed genes across both cultivars (Fig. 3), and the MEturquoise module demonstrated an extremely significant positive correlation with total soluble sugar content, suggesting its potential crucial role in regulating sugar accumulation during wampee fruit development. This conserved co-expression module may represent core regulatory networks involved in sugar metabolism that are shared between these two distinct wampee cultivars.
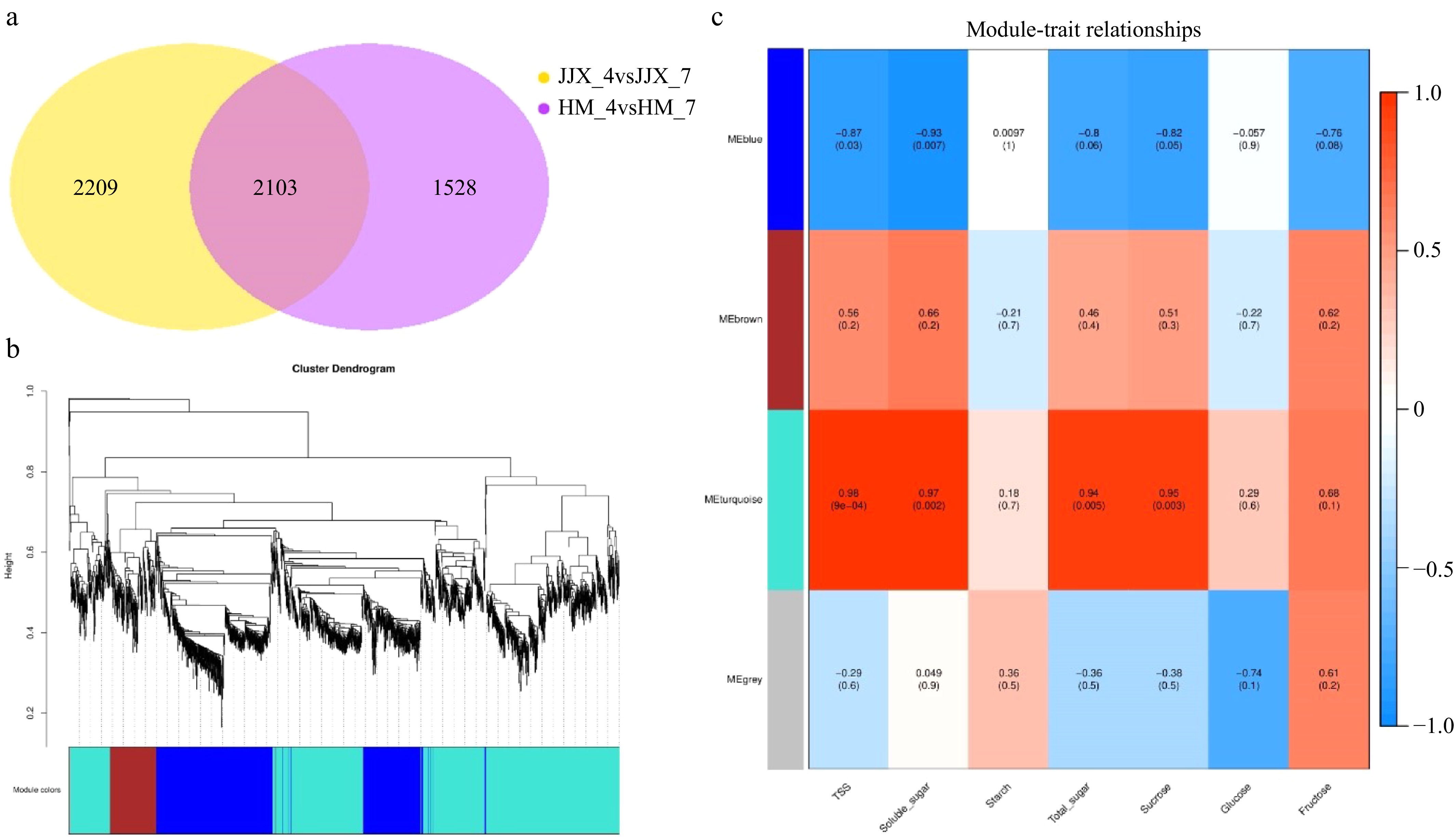
Figure 3.
Weighted gene co-expression network analysis (WGCNA) of differentially expressed genes in wampee fruits at 78 DAB and 99 DAB. (a) Venn diagram of differentially expressed genes. (b) Modular hierarchical clustering tree. (c) Module-trait correlation results plot (the upper number in the module represents the correlation coefficient r2 and the lower number represents the significance p. The color blue to red represents r2 from −1 to 1). HM_4 and HM_7 represent the 78 and 99 DAB 'Huami' wampee fruits, respectively; JJX_4, JJX_7 represent the 78 and 99 DAB 'Jinjixin' wampee fruits, respectively.
Identification of the candidate gene ClSWEET3a
-
Through comprehensive analysis of gene expression patterns in relation to total soluble sugar content (correlation coefficient threshold r2 ≥ 0.75) and annotation of the 'Jixin' wampee genome (Table 2), evm.model.Chr8.149 was identified as exhibiting the strongest correlation (r2 = 0.855) with sugar accumulation.
Table 2. Correlation between genes and soluble total sugar content and their annotations in the reference genome.
Gene ID Correlation coefficient r2 Annotation evm.model.Chr8.149 0.855* Bidirectional sugar transporter SWEET3 OS = Arabidopsis thaliana OX = 3702 GN = SWEET3 PE = 1 SV = 1 * Significant difference (p < 0.05); t-test. Phylogenetic analysis revealed that evm.model.Chr8.149 exhibited the highest sequence homology with AtSWEET3 from Arabidopsis thaliana (Fig. 4a), leading to its designation as ClSWEET3a. Expression profiling demonstrated a progressive upregulation of ClSWEET3a throughout fruit development, with maximal expression observed at 99 DAB (Fig. 4b).
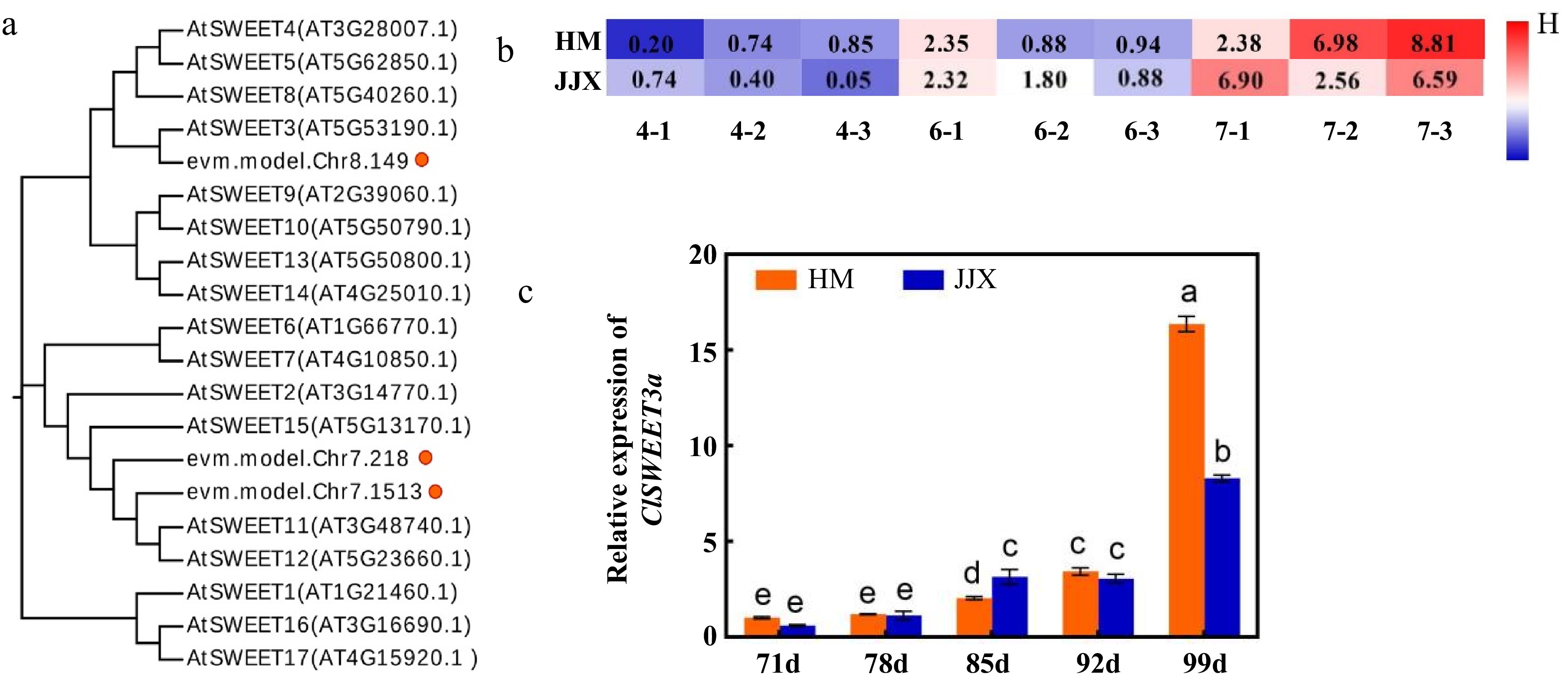
Figure 4.
Phylogenetic tree analysis of ClSWEET3a and expression patterns during fruit development. (a) ClSWEET3a gene and phylogenetic tree analysis of the SWEET family in Arabidopsis. (b) Expression levels of screened ClSWEET3a at different developmental stages of 'Huami' and 'Jinjixin' wampee. 4, 6, 7 represent wampee fruits at 78, 92, and 99 DAB, respectively. 1−3 means three replicates. (c) Expression pattern of ClSWEET3a during the development of wampee fruits. Comparison of the same varieties at different stages. Different lowercase letters represent significance levels at p < 0.05 (one-way ANOVA with Waller-Duncan). HM stands for 'Huami' wampee; JJX stands for 'Jinjixin' wampee. d stands for days after blooming. Error bars represent the standard deviation of three biological replicates.
The correlation analysis (Table 3) indicated that the coefficients between the expression level of ClSWEET3a and total soluble sugar, and sucrose content in HM and JJX were all over 0.85, with a significant positive correlation with sucrose content. These findings strongly suggest that ClSWEET3a plays a crucial regulatory role in sucrose accumulation and total soluble sugar content during wampee fruit maturation. The temporal coordination between ClSWEET3a expression and sugar accumulation patterns further supports its potential function as a key determinant of fruit sugar composition.
Table 3. Correlation analysis of ClSWEET3a expression level with total soluble sugar and sugar component content.
Varieties Gene Total soluble sugar Sucrose Glucose Fructose HM ClSWEET3a 0.876 0.894* 0.591 0.229 JJX ClSWEET3a 0.947* 0.969** 0.534 0.911* HM stands for 'Huami' wampee. JJX stands for 'Jinjixin' wampee. * Significant difference (p < 0.05); ** extremely significant difference (p < 0.01); t-test. Molecular cloning and sequence analysis of ClSWEET3a
-
Using cultivar-specific cDNA templates from both HM and JJX, full-length ClSWEET3a sequences were successfully amplified with designed cloning primers. Sequence alignment with the 'Jixin' reference genome revealed complete identity among all three sequences (750 bp), demonstrating high conservation of ClSWEET3a across wampee cultivars (Supplementary Fig. S2). These results confirm the accuracy of the present cloning strategy and the reliability of the reference genome annotation.
Subcellular localization of ClSWEET3a
-
Subcellular localization analysis provided clear evidence for membrane association of ClSWEET3a (Fig. 5). While control GFP signals appeared in both nucleus and plasma membranes, the ClSWEET3a-GFP fusion protein exhibited exclusive plasma membrane localization. This distinct membrane-specific fluorescence pattern conclusively demonstrates that ClSWEET3a functions as a plasma membrane protein, consistent with its predicted role as a sugar transporter.
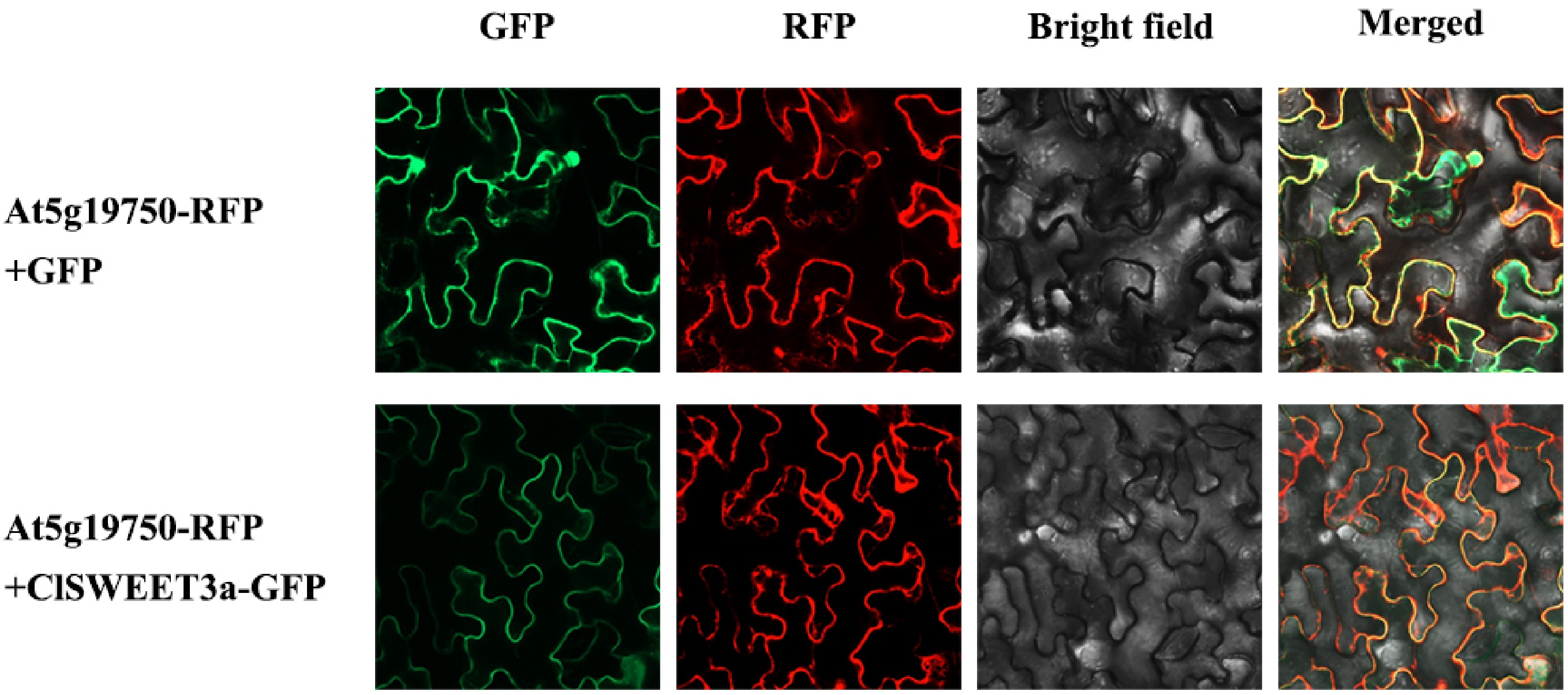
Figure 5.
Subcellular localization of ClSWEET3a in Nicotiana benthamiana. At5g19750-RFP, cytoplasmic 345 membrane marker; At5g19750-RFP + GFP, fusion of At5g19750-RFP and pC18-GFP; At5g19750-346 RFP + ClSWEET3a-GFP, fusion of At5g19750-RFP and ClSWEET3a-GFP. Bar = 20 μm.
Functional characterization of ClSWEET3a in yeast complementation assays
-
The yeast functional complementation assays demonstrated distinct carbon source utilization patterns (Fig. 6). All transformed yeast strains exhibited consistent growth on maltose-containing medium, serving as a positive control for cell viability. On the medium with sucrose and glucose as the sole carbon source, yeasts from the negative control grew weakly, while yeasts transformed with ClSWEET3a grew normally. All transformed yeasts grew weakly on the medium with fructose as the sole carbon source. The above results indicated that ClSWEET3a has the ability to transport sucrose and glucose.

Figure 6.
Transport properties of ClSWEET3a in the yeast EBY.VW4000. (a) The growth of EBY.VW4000 on selection medium with 2% maltose as the sole carbon source. (b) The growth of EBY.VW4000 on selection medium with 2% sucrose as the sole carbon source. (c) The growth of EBY.VW4000 on selection medium with 2% glucose as the sole carbon source. (d) The growth of EBY.VW4000 on selection medium with 2% fructose as the sole carbon source. pDR195 as a negative control. OD600 indicates the absorbance value of the bacterial fluids at 600 nm.
These findings provide compelling evidence that ClSWEET3a specifically facilitates the transport of sucrose and glucose, but not fructose, establishing its substrate preference among different sugar molecules. The differential growth responses clearly correlate with ClSWEET3a's transport capabilities, confirming its functional role in sucrose and glucose uptake.
Verification of the function of ClSWEET3a on sugar accumulation in wampee fruits
-
The expression of ClSWEET3a in wampee fruits was effectively silenced using the VIGS technique (Fig. 7). Compared to the empty vector control, the VIGS-treated fruits exhibited a highly significant reduction in ClSWEET3a expression. Concurrently, the sucrose and fructose contents were dramatically decreased, and glucose levels also showed a significant decline.
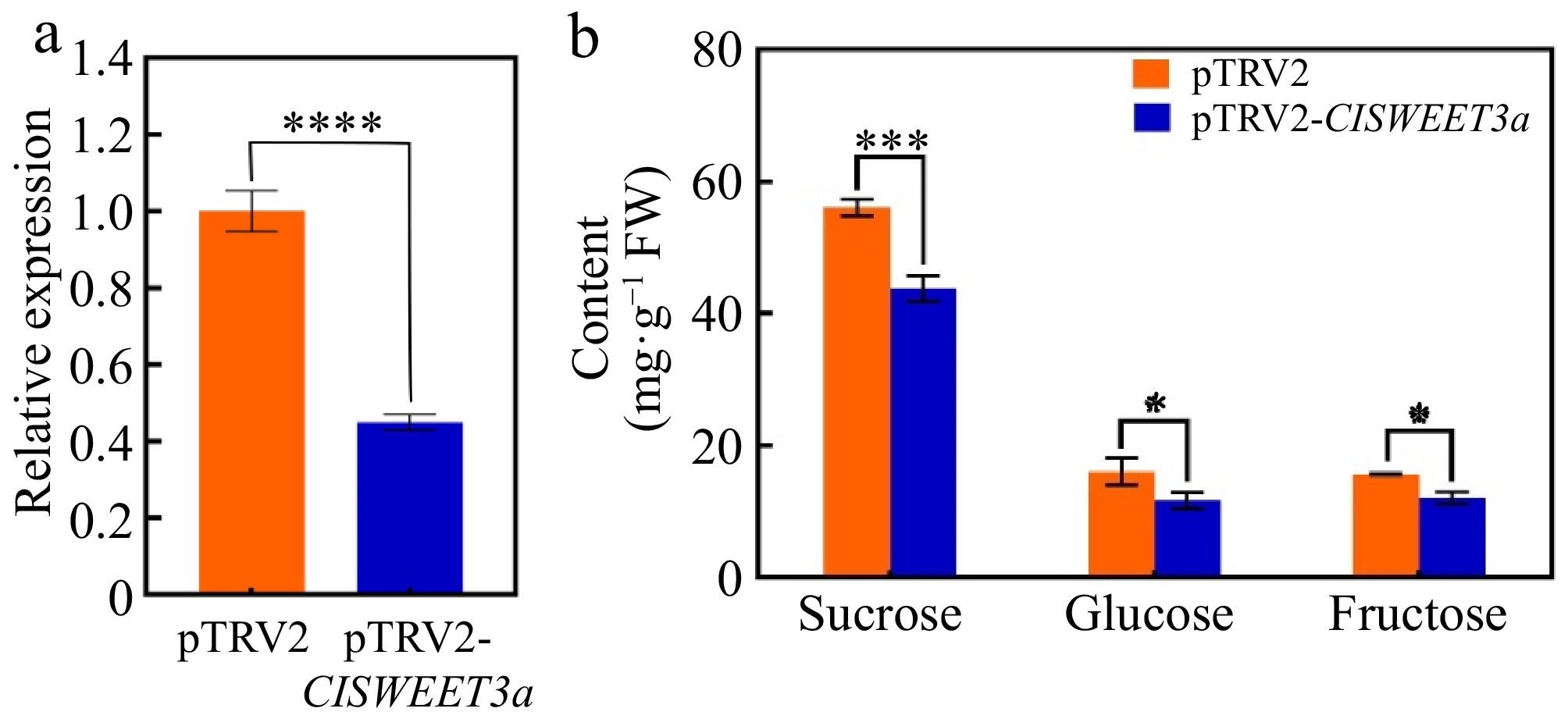
Figure 7.
Transient silencing of ClSWEET3a expression in wampee fruits. (a) Relative expression of ClSWEET3a in wampee fruits after transient silencing of ClSWEET3a. (b) Sucrose, glucose, and sucrose contents of wampee fruits after transient silencing of ClSWEET3a. * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001; t-test.
Dual luciferase reporter assay predicts transcription factors
-
Based on transcriptome data, 12 transcription factors (TFs) exhibiting high correlation (r2 > 0.70) with total soluble sugar content were selected for further analysis. These TFs belonged to multiple gene families, including ERF, bZIP, WRKY, C3H and so on (Supplementary Table S4).
To investigate their regulatory roles, a dual-luciferase reporter assay was conducted using the full-length promoter fragment of ClSWEET3a from JJX (Fig. 8a). Among the 12 TFs tested, ClERF016, ClERF062, ClbZIP3, ClbZIP4, ClRAP2-1, ClGT3A, and ClGATA1 significantly enhanced the activity of the ClSWEET3a promoter, as evidenced by their LUC/REN ratios being markedly higher than those of the negative control. Notably, ClbZIP3 exhibited the strongest transactivation effect (Fig. 8b).
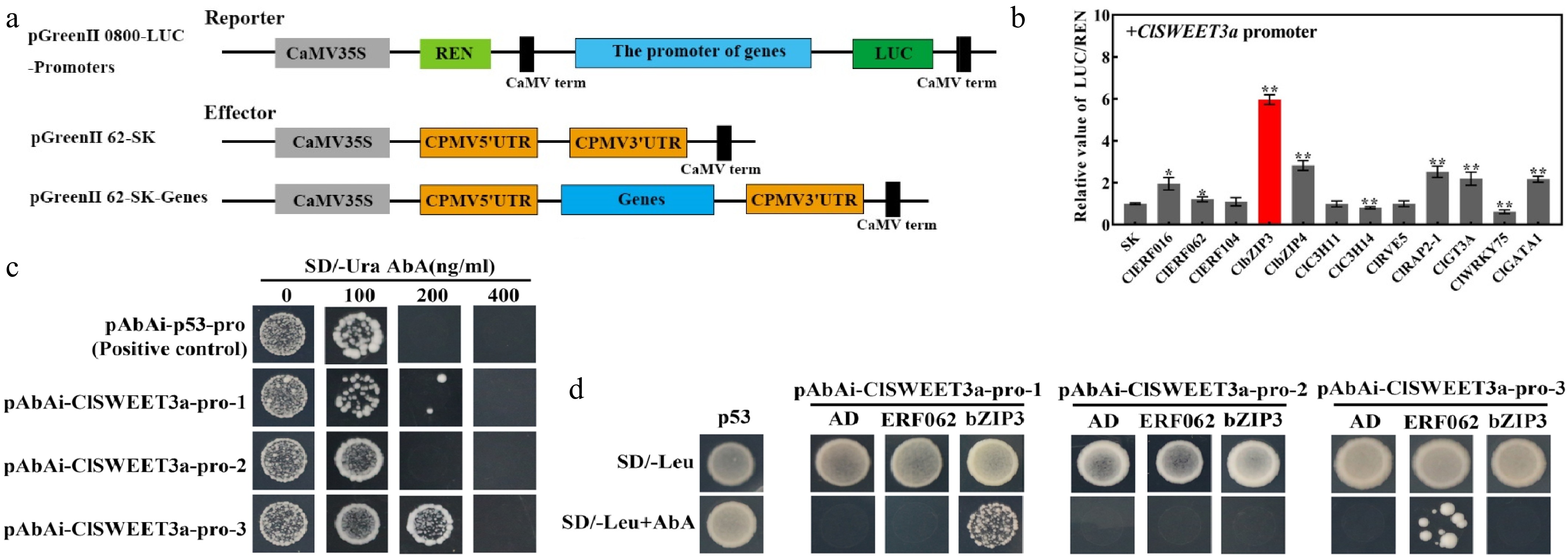
Figure 8.
Transcriptional regulation of ClSWEET3a by transcription factors and analysis of promoter interactions between transcription factor (ClbZIP3 and ClERF062) and ClSWEET3a. (a) Vector diagrams of Reporter and Effector. (b) Transcriptional regulation of the ClSWEET3a promoter by 12 transcription factors. The red bar represents the combination with the largest LUC/REN ratio. The LUC/REN ratio of the empty p62-SK vector was used as a control (set as 1). (c) Bait yeast AbAr expression level assay. (d) ClbZIP3 and ClERF062 binding to the promoter of ClSWEET3a. p53 was used as a positive control. AD was used as a negative control. Significance was analyzed by t-test. Error bars represent the standard deviation of three biological replicates. * p < 0.05; ** p < 0.01; one-way ANOVA with Waller-Duncan.
ClbZIP3 and ClERF062 bind to the key gene promoter of sugar accumulation
-
Dual luciferase reporter assay results indicated that ClbZIP3 had the strongest positive regulatory effect on the ClSWEET3a promoter, and ClERF062 also had a positive regulatory effect. The variation of these two TFs were consistent with the sugar accumulation (Fig. 9). To further investigate their regulatory mechanisms, yeast one-hybrid assays were performed to examine protein-DNA interactions. Both ClbZIP3 and ClERF062 demonstrated direct binding to the ClSWEET3a promoter (Fig. 8), confirming their potential roles in regulating sugar accumulation.
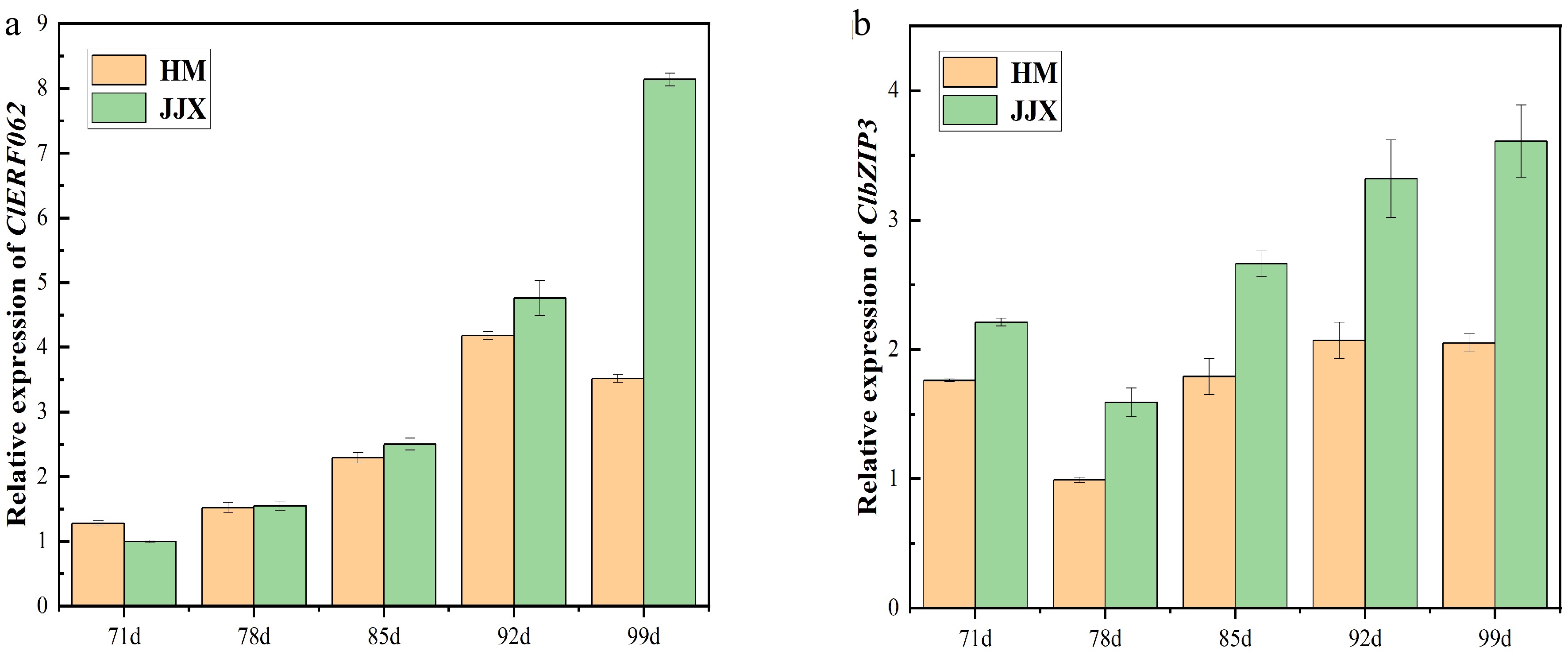
Figure 9.
The expression level of ClbZIP3 and ClERF062 during fruit development by qRT-PCR. (a)Expression of ClERF062; (b) expression of ClbZIP3. HM stands for 'Huami' wampee. JJX stands for 'Jinjixin' wampee. d stands for days after bloom.
-
As the predominant varieties for fresh consumption, sweet and sweet-sour wampee fruits derive their market competitiveness largely from edible quality, with sugar content being a crucial determinant. The present analysis of two cultivars (HM and JJX) revealed dynamic sugar accumulation patterns during fruit development, where hexoses dominated early stages while sucrose became predominant at maturity. These findings align with previous reports in wampee and mirror trends observed in other fruits, highlighting conserved metabolic shifts during ripening[25,26]. Compared with other fruits, such as citrus, the sucrose was dominant throughout the entire process of fruit development[27].
Functional characterization of ClSWEET3a in sugar transport
-
Phylogenetic analysis placed ClSWEET3a in Clade I of the SWEET family, showing high homology to Arabidopsis AtSWEET3, a known hexose transporter. Functional validation through VIGS silencing and yeast complementation assays confirmed its glucose transport activity, consistent with clade-based predictions[28]. Notably, ClSWEET3a also transported sucrose, suggesting functional divergence from its Arabidopsis ortholog. This functional divergence likely arose through evolutionary neofunctionalization following gene duplication, where key mutations in transmembrane domains (notably TM5 helix) expanded CISWEET3a's substrate-binding pocket. The dual-substrate specificity parallels reports also found in citrus (CitSWEET6), grape (VvSWEET10), pear (PuSWEET15), and apple (MdSWEET12a), where SWEET proteins exhibit clade-dependent yet sometimes expanded substrate ranges[12,29−31].
Transcriptional regulation model of sugar metabolism in wampee
-
Recent advances in transcriptional regulation have significantly enhanced our understanding of sugar accumulation mechanisms in fruits. Studies across various species have established clear connections between specific transcription factors and sugar metabolism. Lu et al. have found that citrus fruit sugar quality through the regulation of CsSPSs[27].
In pear, PuWRKY31 activates PuSWEET15 expression, thereby increasing fruit sucrose content, with its activity modulated by promoter acetylation[30]. Similarly, MdWRKY9 in apple promotes sugar accumulation by upregulating MdSWEET9b expression[32]. The present study advances understanding of sugar regulation in wampee by identifying ClbZIP3 and ClERF062 as direct activators of ClSWEET3a (Fig. 10).
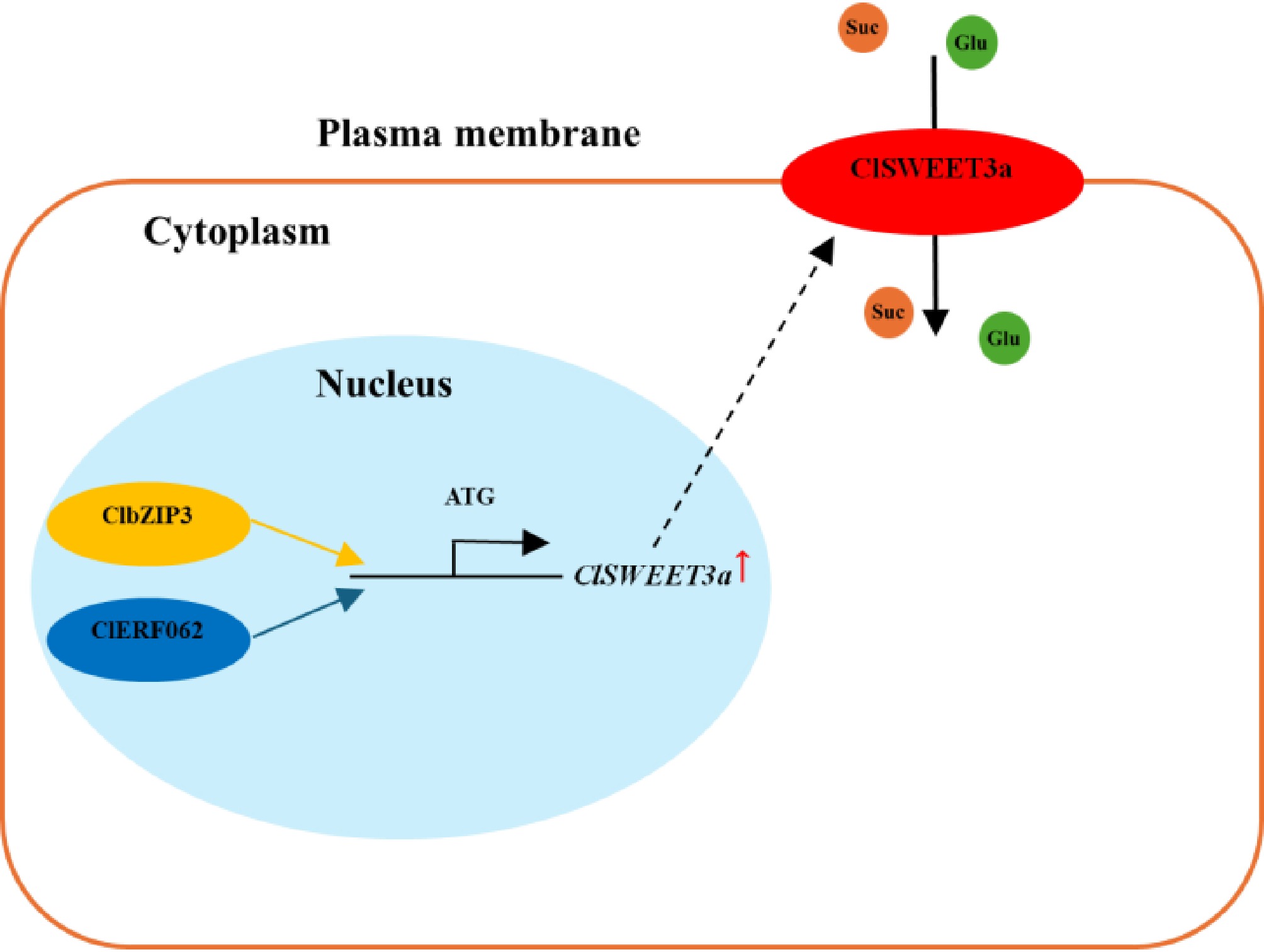
Figure 10.
Proposed model of ClbZIP3 and ClERF062 regulation of ClSWEET3a-mediated sugar accumulation in wampee fruits.
The functional characterization of these transcription factors reveals their potential integration with hormonal signaling pathways. Fruit ripening is a hormone-mediated process intricately coordinated by a network of signaling molecules. The signaling pathway—from receptor-mediated perception to the sequential activation of transcription factors (EIN3/ERFs)—forms a highly conserved regulatory framework[33]. As ERF family members typically function downstream of ethylene signaling[34,35], ClERF062 may connect sugar metabolism with ripening-related ethylene responses. This connection is exemplified in citrus, where ethylene-regulated CitERF6 and CitERF13 control peel degreening through chlorophyll degradation[36]. Similarly, bZIP transcription factors like ClbZIP3 often participate in ABA signaling and stress responses[37,38]. The apple homologs MdbZIP23 and MdbZIP46 illustrate this dual functionality, where they interact with MdWRKY9 to mediate ABA's influence on sugar accumulation[32]. These parallels suggest that ClERF062 and ClbZIP3 may similarly coordinate wampee sugar accumulation with ethylene and ABA signaling pathways, respectively, potentially linking fruit quality development with environmental and hormonal cues. Future studies should investigate these potential connections to fully elucidate the regulatory network controlling sugar accumulation in wampee fruits.
-
In this study, the mechanisms underlying sugar accumulation in wampee fruits were explored by comparing two cultivars— 'Huami' (sweet) and 'Jinjixin' (sweet-sour)—across five developmental stages. Transcriptome analysis identified ClSWEET3a as a key player, with its expression strongly correlated with total soluble sugar and sucrose content. Functional characterization revealed that ClSWEET3a, a plasma membrane-localized transporter, confirming its essential role in sugar accumulation. Furthermore, we uncovered a regulatory network involving two transcription factors (ClbZIP3 and ClERF062) that activate the ClSWEET3a promoter, with ClbZIP3 exhibiting stronger transcriptional control. The present findings provide a molecular framework for sugar accumulation in wampee, offering valuable insights for targeted breeding strategies aimed at enhancing fruit quality.
This work was financially supported by the Science and Technology Planning Project of Guangdong Province and Yunan County (Grant No. h20240204), and the Yonggen Sci-Tech Station Project (Grant No. 202308).
-
The authors confirm contributions to the paper as follows: study conception and design: Zhang Z, Qin Y; data collection: Wu L, Tang X; analysis and interpretation of results: Ding Y, Wei H; draft manuscript preparation: Chen S, Wu L; resources: Hu G, Zhao J, Qin Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Shiyi Chen, Liangfang Wu
- Supplementary Table S1 Summary of primers used in this study.
- Supplementary Table S2 Quality control results of transcriptome sequencing data.
- Supplementary Table S3 Differential gene annotation of wampee fruits in two stages (78 DAB and 99 DAB).
- Supplementary Table S4 Basic information of the screened transcription factors.
- Supplementary Fig. S1 KEGG enrichment analysis of differentially expressed genes in wampee fruits at 78 DAB and 99 DAB.
- Supplementary Fig. S2 Sequencing results of ClSWEET3a compared with the reference nucleotide sequence in the genome database. ClSWEET3a, genome reference sequence ('Jixin' wampee); ClSWEET3a-JJX, sequencing results of bacterial fluids ('Jinjixin' wampee); ClSWEET3a-HM, sequencing results of bacterial fluids ('Huami' wampee).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen S, Wu L, Tang X, Ding Y, Wei H, et al. 2025. ClSWEET3a mediates sucrose accumulation in wampee (Clausena lansium) fruit through transcriptional activation by ClbZIP3 and ClERF062. Fruit Research 5: e037 doi: 10.48130/frures-0025-0027 

ClSWEET3a mediates sucrose accumulation in wampee (Clausena lansium) fruit through transcriptional activation by ClbZIP3 and ClERF062
- Received: 18 June 2025
- Revised: 21 July 2025
- Accepted: 04 August 2025
- Published online: 10 October 2025
Abstract: Soluble sugar is an important factor in determining fruit quality and flavor. In this study, the sugar accumulation mechanisms in wampee (Clausena lansium) fruits were investigated by using two cultivars—'Huami' (sweet) and 'Jinjixin' (sweet and sour)—at five developmental stages. Transcriptome analysis revealed a strong correlation between ClSWEET3a expression and total soluble sugar/sucrose content. Functional characterization demonstrated that ClSWEET3a, a plasma membrane-localized transporter, facilitates glucose and sucrose uptake in yeast. Silencing of ClSWEET3a in 'Jinjixin' fruits significantly decreased both sucrose and hexose levels, confirming its role in sugar accumulation. Furthermore, we identified two transcription factors (ClbZIP3 and ClERF062) that activate the ClSWEET3a promoter, with ClbZIP3 exhibiting stronger regulatory effects. Yeast one-hybrid assays confirmed their direct binding to the ClSWEET3a promoter. Our findings elucidate a molecular framework for sugar accumulation in wampee and provide insights for targeted breeding strategies to improve fruit quality.
-
Key words:
- SWEET /
- Wampee (Clausena lansium) fruits /
- Sugar /
- Transcription factors /
- Fruit quality