









-
With the continuous warming of the global climate, the worsening of soil salinization has severely threatened the ecological environment, and the sustainable development of human society[1]. More than 800 million hectares of soil worldwide are affected by salinization[2], and saline-alkali land has become a major factor limiting agricultural and forestry production in China[3].
Studies have demonstrated that salt stress inhibits plant growth, disrupts physiological processes, reduces soil osmotic potential, causes ion imbalance, and can even lead to plant death under high salt concentrations[4]. Although technological innovations and scientific advances in managing saline-alkali land have made progress, they are still restricted by regional and resource limitations. Therefore, enhancing plant salt tolerance and exploring the mechanisms behind it are of great significance[5]. Plants adapt to saline-alkali stress by modifying their morphology[6]. Salt stress inhibits plant height, branch number, stem diameter, root surface area, total root length, and biomass. It also causes varying degrees of leaf surface damage, weakens root vitality, and may lead to plant death when stress exceeds tolerance thresholds[7]. The root system is the first to perceive stress signals, which are then gradually transmitted to the above-ground parts. Studies have found that growth and biomass changes under salt stress are related to rootstock varieties, as rootstocks with different salt tolerance capacities show varying growth and biomass to increasing salt stress[8].
At the cellular and tissue levels, salt stress induces a series of physiological and biochemical responses in plants[9], including osmotic, ionic, and oxidative stress[10]. Salt stress leads to osmotic imbalance, prompting plants to accumulate osmoregulatory substances such as proline, soluble sugars, and soluble proteins to mitigate its effects[11]. It reportedly disrupts ionic balance by increasing Na+ and Cl− concentrations while decreasing Ca2+, K+, and NO3− contents[12]. It also elevates reactive oxygen species (ROS) levels, including superoxide radicals, hydrogen peroxide, and hydroxyl radicals[13]. To mitigate ROS damage, plants adjust antioxidant enzyme activity—such as superoxide dismutase, catalase, and peroxidase—based on salt concentration and exposure time[14]. Some studies suggest that plants enhance salt tolerance by maintaining pH and ion homeostasis[15], while others have suggested that some salt-tolerant or salt-sensitive plants have evolved different mechanisms to adapt to salt-alkali stress[16].
Apples are among the most widely cultivated fruit crops worldwide, and nutrient transport and metabolism significantly influence their yield and quality[17]. As sessile organisms, apple trees must adapt to abiotic stresses such as soil salt-alkali damage and extreme temperatures. Rootstocks affect scion growth, fruit yield, and quality[18]. Dwarfing rootstocks are widely used for their ability to induce early flowering, control tree size, reduce labor requirements, and improve fruit yield[19]. In apple cultivation, salt-tolerant rootstocks are increasingly used to improve plant resistance[20]. However, studies on rootstock resistance to combined salt and alkali stress are limited, even though these stresses often occur together in nature. Therefore, identifying suitable rootstocks for comprehensive stress resistance evaluation is crucial. Systematic research on the abiotic stress responses of apple rootstock resources can help identify high-quality, stress-tolerant candidates, and improve overall apple resilience[21]. Existing research on salt tolerance in apples typically relies on controlled NaCl solution treatments or focuses solely on a single rootstock/scion combination, failing to simulate the complex stress environment of natural saline-alkali soil in apple production (such as mixed ion stress, soil structure changes, and nutrient imbalance). To our knowledge, few studies have systematically compared the salt tolerance of different apple rootstock-scion combinations under real saline-alkali field conditions. The present study specifically uses natural saline-alkali soil as the stress medium, and focuses on three combinations with the same scion (ensuring consistent genetic background of the aboveground part) but different rootstocks, thereby enabling the quantification of the impact of rootstock characteristics on the salt tolerance of the entire grafted system independently. This design provides more ecologically and agriculturally relevant insights for the practical cultivation of apples in saline-alkali areas. In this study, three different rootstocks were used to graft the 'Luli' cultivar. By comprehensively evaluating how each rootstock affects the salt tolerance of 'Luli' scions, this study aimed to identify optimal salt-tolerant rootstock combinations. Simultaneously, it provides deeper insights into the differences in salt tolerance mechanisms among rootstocks, offering a scientific basis for optimizing grafting strategies.
-
The scion varieties of experimental materials were all Luli. As a scion variety, Luli is a representative variety of early ripening and high quality apples, which fills the vacancy of early ripening and high quality apples in the market. The rootstocks were Qingzhen-1, M9-T337, and PYTC with different salt sensitivity, respectively. From the Tianping Lake experimental base of Shandong Fruit Research Institute (Shandong, China), located at 36°12'53" N and 117°5'40" E, annual seedlings of about 40 cm were selected with the same growth and good growth. The soil required for the experiment was collected from the Tianping Lake Base in Tai'an, and the Yellow River Delta Development Zone in Dongying. Among them, Dongying is located in the heart of the Yellow River Delta. It is a coastal saline land with severe soil salinisation, making it one of the most challenging delta saline lands in China (and the world). This experiment selected natural saline-alkali soil for two core reasons: First, it simulates the complex real-world conditions of apple production in saline-alkali regions. Natural saline-alkali soil not only contains high concentrations of soluble salts like Na+ and Cl−, but also suffers from degraded soil structure, low nutrient availability, and imbalanced ion ratios—factors that cannot be fully replicated by simple NaCl solution treatments in controlled environments. These combined factors significantly impact plant growth. Second, it avoids the limitations inherent in studies using sole NaCl stress. Previous studies often overlooked the synergistic or antagonistic effects of other ions and soil property changes under natural saline-alkali conditions. Using natural saline-alkali soil more accurately reflects the overall stress response of apple rootstock-scion combinations under actual field conditions, providing more ecologically and agriculturally practical references for enhancing apple yields in saline-alkali areas.
Taking the soil samples from Tianping Lake base in Tai'an as the control (A), the saline soil samples from Dongying Yellow River Delta were washed with tap water to soil with corresponding salt concentration, and 0.0, 2.5 and 4.0 L tap water were washed for every 10 kg of saline soil, and soil materials with salt concentration of (B) 10‰, (C) 6‰, and (D) 3‰ were obtained (Table 1). The salt concentration, ion concentration, pH value, and organic content of the obtained soil were measured. The measured pH of the experimental soil was classified as highly saline and slightly alkaline, since the present experiment centered around salt stress[22].
Table 1. Soil salt concentration and organic matter content.
Sample Saltness (‰) N (g/kg) P (g/kg) K (mg/g) Na (mg/kg) pH Organic carbon (g/kg) Organic matter (g/kg) A 1 ± 0.32 1.2 ± 0.3 1.15 ± 0.27 2.1 ± 0.26 140 ± 29 6.8 ± 0.2 8.21 ± 1.14 12.68 ± 1.35 B 10 ± 0.8 0.49 ± 0.11 0.37 ± 0.18 0.35 ± 0.08 738 ± 23 7.4 ± 0.1 5.91 ± 0.58 9.67 ± 1.26 C 6 ± 0.5 0.38 ± 0.09 0.45 ± 0.25 0.42 ± 0.09 426 ± 36 7.3 ± 0.2 6.07 ± 0.56 9.24 ± 1.52 D 3 ± 0.28 0.35 ± 0.076 0.61 ± 0.21 0.56 ± 0.05 316 ± 27 7.3 ± 0.1 6.05 ± 0.42 9.11 ± 1.22 The transplanted plants were potted into soil holes of 40 cm × 60 cm, with mulch film at the bottom, rain shelter for cultivation, bottom watering, and infiltration irrigation. Soil samples in the pot were taken regularly for salt content detection to ensure the accuracy of the experimental gradient. Photosynthesis, and various biochemical indicators of leaves at the same position with different gradients were measured regularly (Table 2).
Table 2. Test design table.
Saline
concentrationApple variety (plant/pot) Subtotal Luli/Qing Zhen 1 Luli/M9-T337 Luli/PYTC 1‰ (CK) 30 30 30 90 3.0‰ 30 30 30 90 6.0‰ 30 30 30 90 10.0‰ 30 30 30 90 Total 120 120 120 360 Measurement of indicators
Statistics of mortality and survival rate
-
According to the method of Tahir et al.[23], the mortality rate is the number of deaths per plant variety as a percentage of the total number of the plant variety.
Determination of root morphology
-
According to the instructions of the WinRHIZO root scanner, the cleaned roots were placed horizontally, and the physiological data of the scanned roots were measured using ImageJ software[24].
Growth index measurement
Plant height
-
The distance from the base of the plant to the top of the main stem of the plant was measured using a tape measure with an accuracy of 1 mm.
Ground diameter
-
A Vernier caliper, with an accuracy of 0.01 mm was used to determine the diameter of the plant, 5 cm from the ground.
Crown width
-
A tape measure with an accuracy of 1 mm was used to measure the average north-south and east-west width of the plant.
Branch length
-
The distance from the base of the longest branch to the top of the branch is determined using a tape measure with an accuracy of 1 mm.
Leaf area
-
The leaf area of the leaves was scanned, and the leaf area calculated using ImageJ software.
Blade thickness
-
Blade thickness is measured with a vernier caliper with an accuracy of 0.01 mm.
Relative conductivity measurement
-
The relative electrical conductivity of the plants was measured according to the method of Jia et al.[25]. The electrical conductivity of the solution was measured by using Shanghai Leizhi DDS-307 conductivity meter (produced by Yi Electrical Scientific Instrument Co., Ltd.). The conductivity of ultra-pure water, denoted as EC0, was measured, then 0.1 g fresh root sample was soaked in ultra-pure water for 4 h, and the conductivity of the obtained solution was measured and recorded as EC1. Finally, the solution obtained after soaking for 4 h was boiled for 30 min and cooled to room temperature. The conductivity of the solution was measured and recorded as EC2.
Determination of relative water content
-
The relative water content of leaves of plants was determined according to the method of Xue et al.[26]. After cleaning, six leaves were taken from each treatment group, and their fresh weight was weighed on the analytical balance as Wf; then the leaves were immersed in pure water for 24 h, wiped clean and their saturated fresh weight recorded as Wt; the sampleds were immediately weighed, then dried at 105 °C for half an hour, dried at 80 °C to constant weight, and their dry weight measured as Wd.
Toluidine blue staining
-
Slices of apple leaves were stained with 0.05% toluidine blue (TB) staining solution at 25 °C, and the leaf tissues after staining were observed and determined with a microscope[27].
Measurement of stomatal pore size
-
The stomatal closure experiment was conducted according to the method of Wang et al.[28]. Apple leaves of 4-week-old plants at the same developmental stage were obtained. The leaves were immersed in a buffer (10 mM KCl, 7.5 mM iminodiacetic acid, 10 mM MES-Tris, pH 6.15) for 2 h under light. The stomatal opening of the epidermis was observed and determined using a microscope. The pore size was detected using Image J software. At least 10 stomatal diameters were calculated for each apple leaf, and each calculation was repeated three times.
Endogenous hormone measurement
-
The contents of abscisic acid (ABA), gibberellin (GA4), indoleacetic acidv (IAA), jasmonic acid (JA), trans-zeatin-riboside (ZR), and salicylic acid (SA) were determined by enzymo-linked immunosorbent assay (ELISA). Phytohormones contents were detected by Guocangjian (
www.targetcrop.com ) based on the Sciex 4500 LC-MS/MS platform. Three or more replicates of each assay were performed[29].Determination of osmotic regulatory substances
Proline content
-
The proline content was determined using proline (PRO) Content Determination Kit and according to the operating instructions (Keming, Suzhou, China). The instruments used were: a visible spectrophotometer, a water bath, a bench top centrifuge, an adjustable pipette, a micro glass cuvette, acetic acid, a mortar/homogeniser, ice, and distilled water.
Soluble sugar content
-
0.5 g dry leaf sample was accurately weighed, centrifuged with 75% methanol at 8,000 × g at 70 °C for 10 min, and soluble sugar in the sample was extracted three times, combined with the supernatant, and adjusted to a fixed volume of 25 mL. The absorbance of the supernatant at 450 nm was determined using an enzymoleter (INFINITE 200 PRO, Austria) with standard glucose as the internal reference, and soluble sugar content expressed in mg/g FW.
Soluble protein content
-
Soluble protein was determined using Coomassie bright Blue G250 as the color reagent, and bovine serum albumin as the internal reference protein to measure the soluble protein content. Soluble protein content is expressed in mg/g FW.
Determination of oxidase activity
-
SOD activities in leaves were determined using superoxide Dismutase (SOD) Kit, according to the operating instructions (Keming, Suzhou, China).
POD activities in leaves were determined using peroxidase (POD) Kit, according to the operating instructions (Keming, Suzhou, China).
CAT activities in leaves were determined using catalase (CAT) reagent kit, according to the operating instructions (Keming, Suzhou, China).
The instruments used were: visible spectrophotometer, benchtop centrifuge, adjustable pipette, 1 mL glass cuvette, mortar and pestle, ice, and distilled water.
Malondialdehyde determination
-
Apple leaves (0.5 g) were weighed into a cold mortar and ground with 2 mL 0.05 mol/L phosphoric acid buffer. The homogenate was transferred into a test tube, 5 mL 0.05% thiobarbituric acid solution was added, boiled for 10 min, cooled and then centrifuged at 5,000 × g for 10 min. The supernatant was determined using 0.5% thiobarbituric acid solution as a blank, with the absorption values at 450, 532, and 600 nm[30].
Determination of chlorophyll content
-
Apple leaves (0.5 g), were weighed and transferred to a test tube containing 1.5 mL 95% ethanol for dark culture, for one day. The absorbance at 470, 649, and 665 nm was measured by spectrophotometry, and the contents of chlorophyll a and b were calculated[31].
Measurement of photosynthetic parameters
-
According to the instructions of the photosynthesometer Li-6800 (Li-COR, USA), the photosynthesis of plants were measured. The photosynthetic parameters were measured from 9:30 to 12:00 in the morning, and from 14:30 to 16:00 in the afternoon. Healthy and mature leaves of fruit trees were selected to measure the light and parameters of the system leaves, the data read, and the net photosynthetic rate, stomatal conductance, intercellular carbon dioxide concentration, and transpiration rate recorded.
Determination of ion content
-
The dried leaf sample was boiled with HNO3 and H2O2 at 220 °C for 60 min using a flame photometer (M410, Sherwood Scientific, Cambridge). The content of Na+ and K+ and other elements were determined[32].
Expression of related genes
-
RNA was reextracted from apple leaves using the OmniPlant RNA Kit (DNase I) (Conway Century Biology, Jiangsu, China). Reverse transcription was performed using the PrimeScript First Strand cDNA Synthesis Kit (TaKaRa, Dalian, China), to obtain cDNA. The gene expression levels were measured using the ABI7500 qRT-PCR instrument by real-time fluorescence quantitative PCR. Gene expression analysis was conducted using the Ct (cycle threshold) 2−ΔΔCᴛ technique, with 18S serving as the internal reference. Each qRT-PCR reaction sample was replicated a minimum of three times. Supplementary Table S1 lists the pertinent primers[33].
Data analysis
-
The data of the statistical analysis were analyzed by Excel, and SPSS 22.0 software. Duncan multiple mean comparison test was used to distinguish the significant difference between different letters, p < 0.05. Triplicate biological and technical repetitions were carried out for each experiment. GraphPad Prism software was used for mapping[34].
-
The plant growth index is the most intuitive index of plant salt tolerance[35]. After the salt stress treatment, the growth status of the three rootstock-scion combinations was observed (Fig. 1a). Plant height, crown width, stem diameter, leaf number, and branch length all showed a decreasing trend with increasing salt concentrations (Fig. 1b−d, f−g), whereas mortality increased with salt concentration (Fig. 1e). Under control conditions, all three rootstock-scion combinations grew well, with no plant mortality observed. At 3‰ salt concentration, the mortality rate of Luli/Qingzhen-1 was nearly zero, while Luli/M9-T337 and Luli/PYTC showed mortality rates of approximately 52% and 47%, respectively. In contrast, at 10‰ salt stress, the growth of Luli/Qingzhen-1 remained significantly better than that of Luli/M9-T337 and Luli/PYTC, with a mortality rate of only about 10%, compared to 72% and 75% in Luli/M9-T337 and Luli/PYTC, respectively. These phenotypic observations confirmed that salt stress significantly inhibited the growth of Luli/M9-T337 and Luli/PYTC more than Luli/Qingzhen-1. The results demonstrated that Luli/Qingzhen-1 exhibited good salt tolerance.
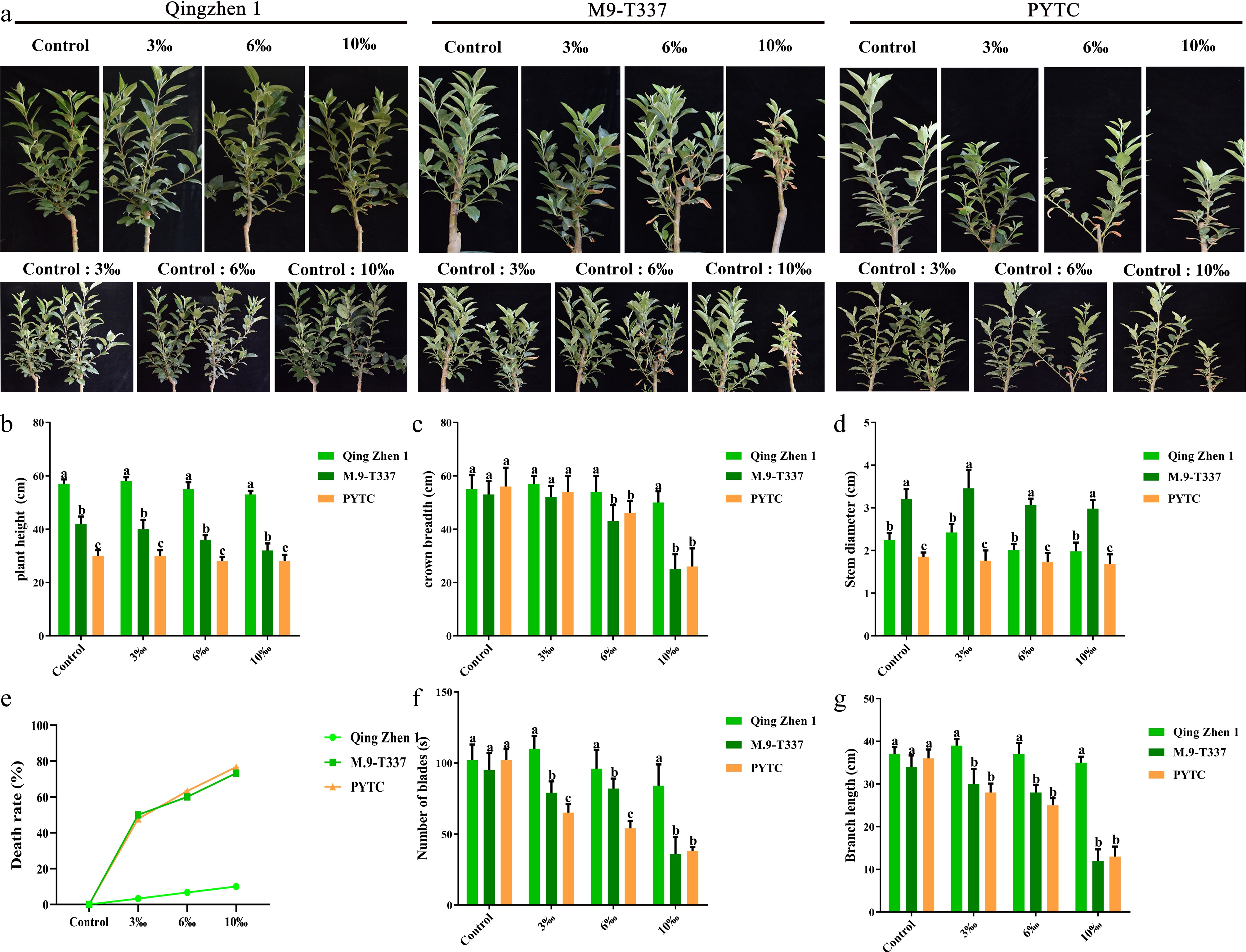
Figure 1.
Plant growth indicators under salt stress. (a) The growth phenotypes of Luli/Qingzhen-1, Luli/M9-T337, and 'Luli'/PYTC were studied (1 month after transplanting into saline-alkali soil). (b) Plant height, (c) crown width, (d) stem diameter, (e) mortality rate, (g) branch length, the value is the average± standard error (n ≥ 3), and the letter indicates that there is a significant difference at the p < 0.05 level under each concentration treatment.
Effects of salt stress on plant leaves
-
The quality of plant leaf growth is one of the important indicators reflecting plant survival under stress, ensuring normal physiological activity[36]. Under salt stress, the leaf phenotype of the three rootstock-scion combinations is shown in Fig. 2a. Under normal growth conditions, the leaves of the three rootstock-scion combinations grew evenly and well. Under 10‰ salt stress, the plant leaves were damaged; only slight wilting appeared around the leaf margins of Luli/Qingzhen-1 anvil combined seedling, and the leaf death rate was about 17%, while the leaves of other anvil combined seedlings were severely damaged, with shriveling, curling, and even drying, and the leaf death rate reached about 48% and 54%, respectively (Fig. 2k). Leaf fresh weight (Fig. 2b), leaf area (Fig. 2c), chlorophyll content (Fig. 2j), and leaf water content (Fig. 2i) were all significantly reduced. Under 6‰ salt stress, the leaves of Luli/Qingzhen-1 seedlings grew well, while the leaves of Luli/M9-T337 and Luli/PYTC seedlings showed slight yellowing around the margins. At 3‰, leaf phenotype returned to normal, but internal physiological indices remained adversely affected compared with Luli/Qingzhen-1 anvil combined seedling.
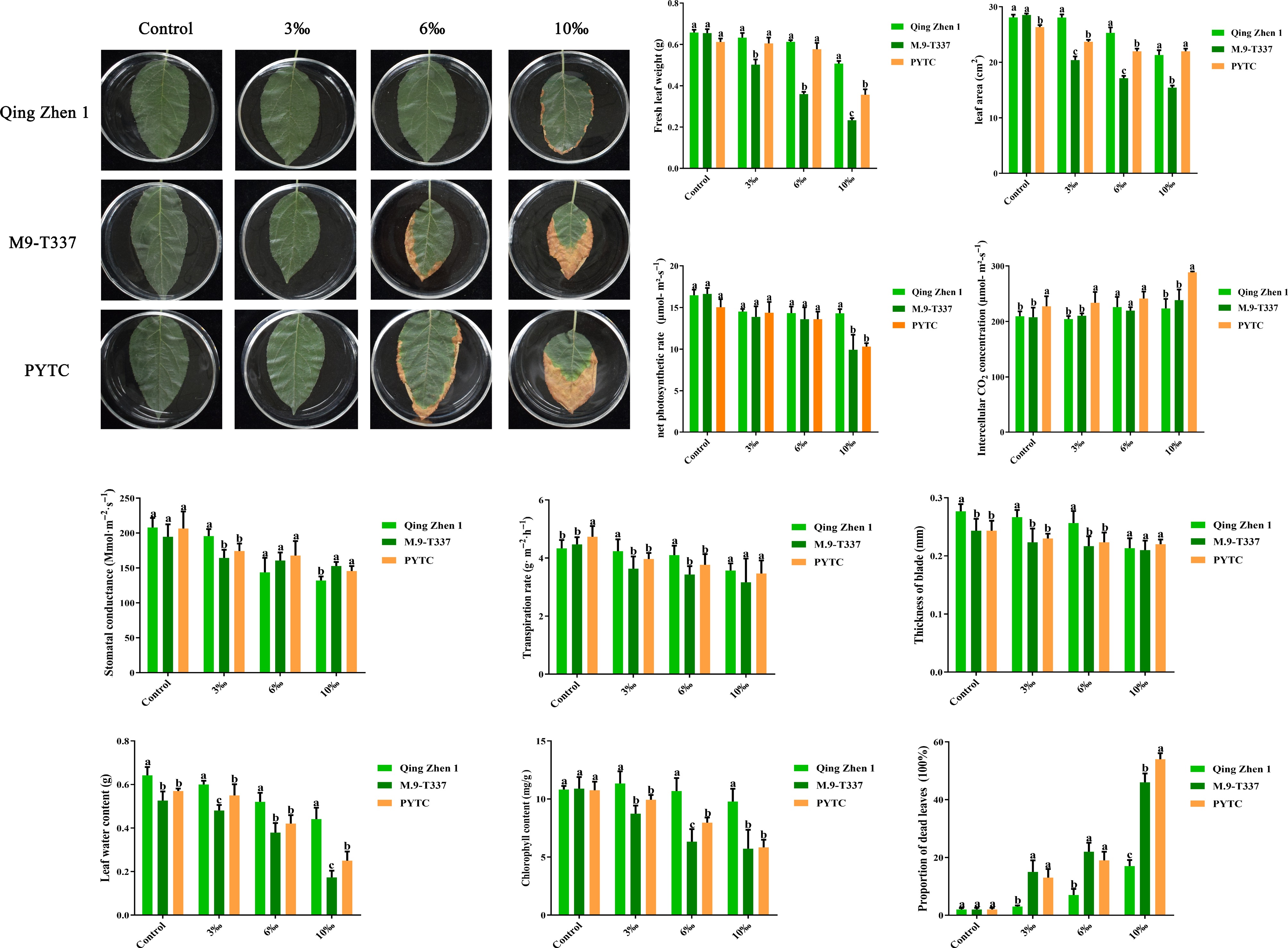
Figure 2.
Effects of salt stress on plant leaves. (a) Leaf damage degree phenotype (2 months after transplanting into saline-alkali soil). (b) Leaf dry weight, (c) leaf area, (d) net photosynthetic rate (Pn), (e) intercellular carbon dioxide concentration (Ci), (f) stomatal conductance (Gs), (g) transpiration rate (E), (h) leaf thickness, (i) leaf water content, (j) chlorophyll, (k) proportion of dead leaves. Values are mean ± standard error (n ≥ 3), and letters indicate significantdifferences at p < 0.05 levels under each concentration treatment.
Photosynthesis is an important index of plant stress resistance and reflects the adaptability of plants to the environment to a certain extent. Under normal growth conditions, photosynthetic indices were relatively normal with no significant differences, but under salt conditions, net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular carbon dioxide concentration (Ci), and transpiration rate (E) of the three rootstock-scion combinations showed a decreasing trend with increasing stress concentration, although the decline in Luli/Qingzhen-1 was relatively low, and that of Luli/M9-T337 and Luli/PYTC was more pronounced. Under 6‰ and 3‰ salt conditions, photosynthetic indices were less affected, and Luli/Qingzhen-1 showed the smallest decline (Fig. 2d−g). The results showed that the leaf indices of Luli/Qingzhen-1 reflected good adaptability to salt stress.
Effects of salt stress on plant leaf structure
-
Continuous salt stress reduces plant cell turgor pressure, thereby limiting cell growth; plants must perform osmotic adjustment to maintain cell expansion and growth[37]. The leaf anatomical structure of the three rootstock-scion combinations was observed using an optical microscope. The results showed that under normal conditions, the leaves of all three combinations had good structure and no tissue damage. Under 3‰ salt stress, the leaf structures remained similar to the control group, with no significant damage. At 6‰, compared with the control, the tissue structure of Luli/Qingzhen-1 was clear and its cells were closely arranged, while Luli/M9-T337 and Luli/PYTC showed slight damage. Under 10‰ salt stress, Luli/Qingzhen-1 leaves had less structural damage and more abundant cells. In contrast, the internal structure of Luli/M9-T337 and Luli/PYTC leaves was severely damaged, with fewer, irregularly shaped cells and loosely arranged, incomplete palisade tissues (Fig. 3). The results showed that under low salt stress, all three combinations experienced slight structural damage, while under high salt stress, Luli/Qingzhen-1 maintained relatively intact leaf structure, indicating strong resistance to salt stress.
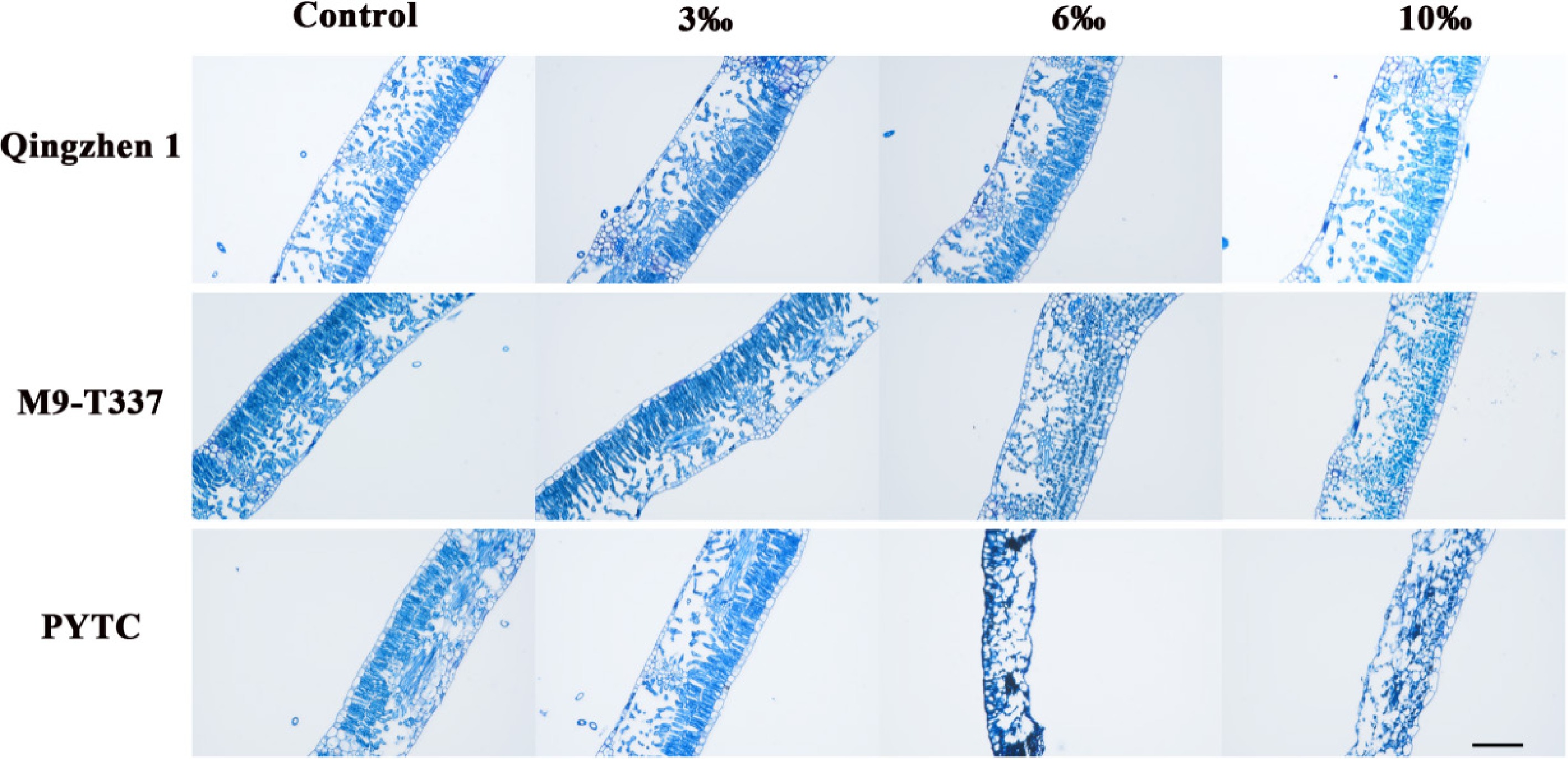
Figure 3.
Effects of salt stress on plant leaf structure. Scale bars: 500 µm (2 months after transplanting into saline-alkali soil).
Effects of salt stress on element contents in plant leaves
-
Through experiments under different salt concentrations, the elemental contents of the shoot leaves of the apple rootstock-scion combinations were analyzed (Fig. 4). Under control conditions, the K+ content of Luli/M9-T337 was significantly lower than that of Luli/Qingzhen-1 and Luli/PYTC. However, under salt stress, Na+ content increased gradually with salt concentration. At 10‰, Na+ content in Luli/Qingzhen-1, Luli/M9-T337, and Luli/PYTC increased by 120%, 210%, and 225%, respectively, compared with the control, and the K+ ions showed a gradually decreasing trend with increasing salt concentration. Compared with the control, the K+ content decreased gradually: by 26%, 51%, and 70% in Luli/Qingzhen-1, Luli/M9-T337, and Luli/PYTC, respectively. In addition, under salt stress, the contents of several trace elements (N, P, Mn, Mo, Mg, and Zn; Fig. 4c−g) showed a downward trend, with Luli/Qingzhen-1 experiencing the smallest decrease. The contents of Fe and Cu (Fig. 4h, i), however, increased under salt stress. Based on the analysis, compared with Luli/M9-T337 and Luli/PYTC, Luli/Qingzhen-1 exhibited a smaller increase in Na+ and a smaller decrease in other ion contents, indicating less sodium uptake and lower ion toxicity, and thereby a better growth trend.
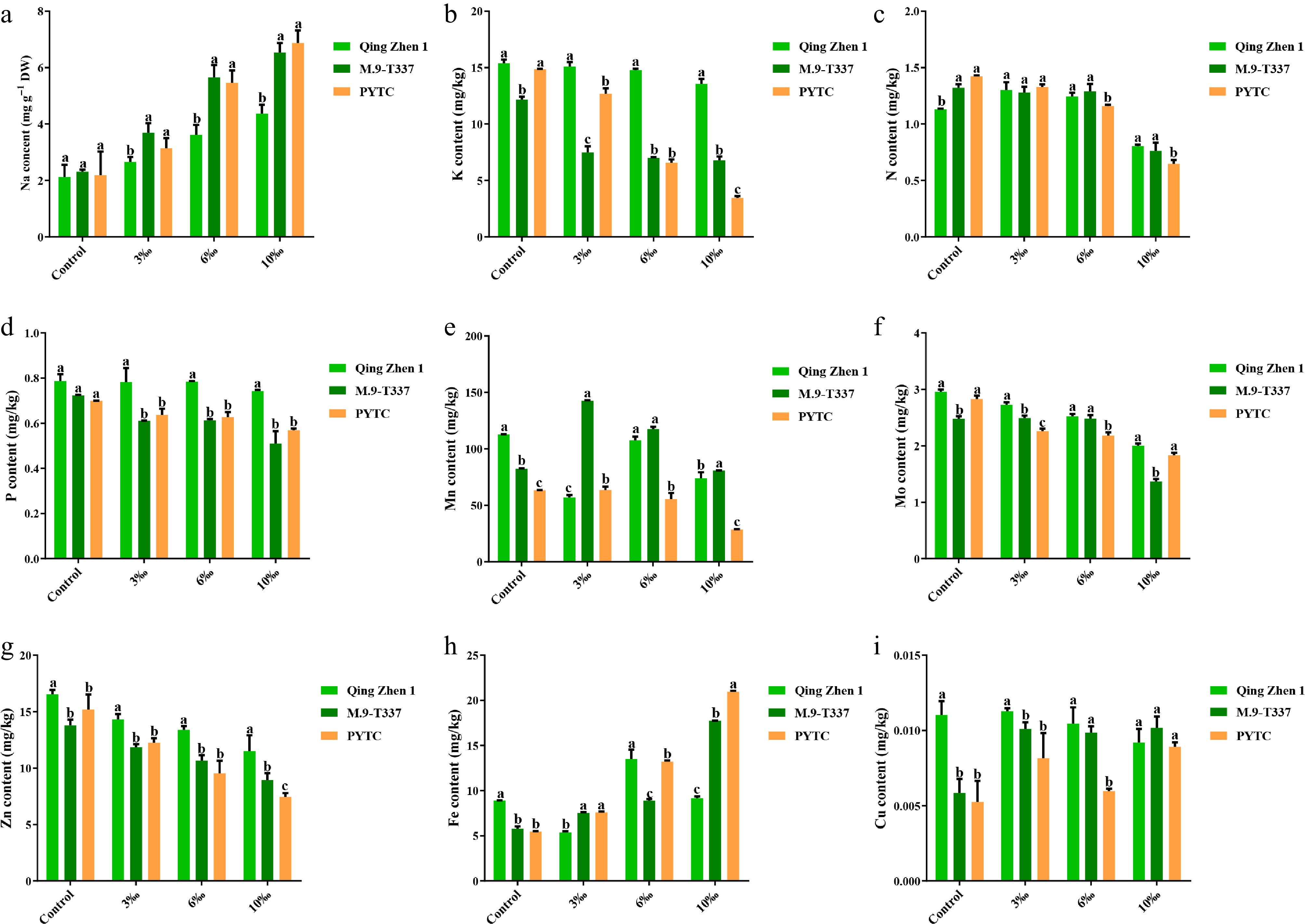
Figure 4.
Metal ion content (2 months after transplanting into saline-alkali soil). (a) Na ion content, (b) K ion content, (c) N ion content, (d) P ion content, (e) Mn ion content, (f) Mo ion content, (g) Zn ion content, (h) Fe ion content, (i) Cu ion content. The value is the average ± standard error (n ≥ 3), the letter indicates that there is a significant difference at the p < 0.05 level under each concentration treatment.
Effects of salt stress on activities of antioxidant enzymes, MDA, and REC in plants
-
The activities of antioxidant enzymes and the contents of MDA and REC were measured under salt stress (Fig. 5). Under normal conditions, antioxidant enzyme activities varied among the combinations, likely due to differences in rootstock genotypes. However, with increasing salt stress, CAT, POD, and SOD activities in Luli/Qingzhen-1 showed a gradual increase compared with the control group, while those inLuli/M9-T337 and Luli/PYTC decreased gradually (Fig. 5a−c). This indicated that Luli/Qingzhen-1 had a stronger ability to scavenge ROS under salt stress, thereby better maintaining redox homeostasis. We also measured hydrogen peroxide content (Fig. 5f). The contents of MDA and relative conductivity (REC) in Luli/M9-T337 and Luli/PYTC increased significantly under salt stress, while the increase in Luli/Qingzhen-1 was smaller. In addition, under normal conditions, there were no significant differences in MDA and REC contents among the three combinations. Under salt stress, MDA and REC increased with rising salt concentration in all three combinations, but the values in Luli/Qingzhen-1 were significantly lower than those in Luli/M9-T337 and Luli/PYTC (Fig. 5d, e). These results indicate that Luli/Qingzhen-1 may resist salt stress by regulating its antioxidant system and maintaining membrane stability.
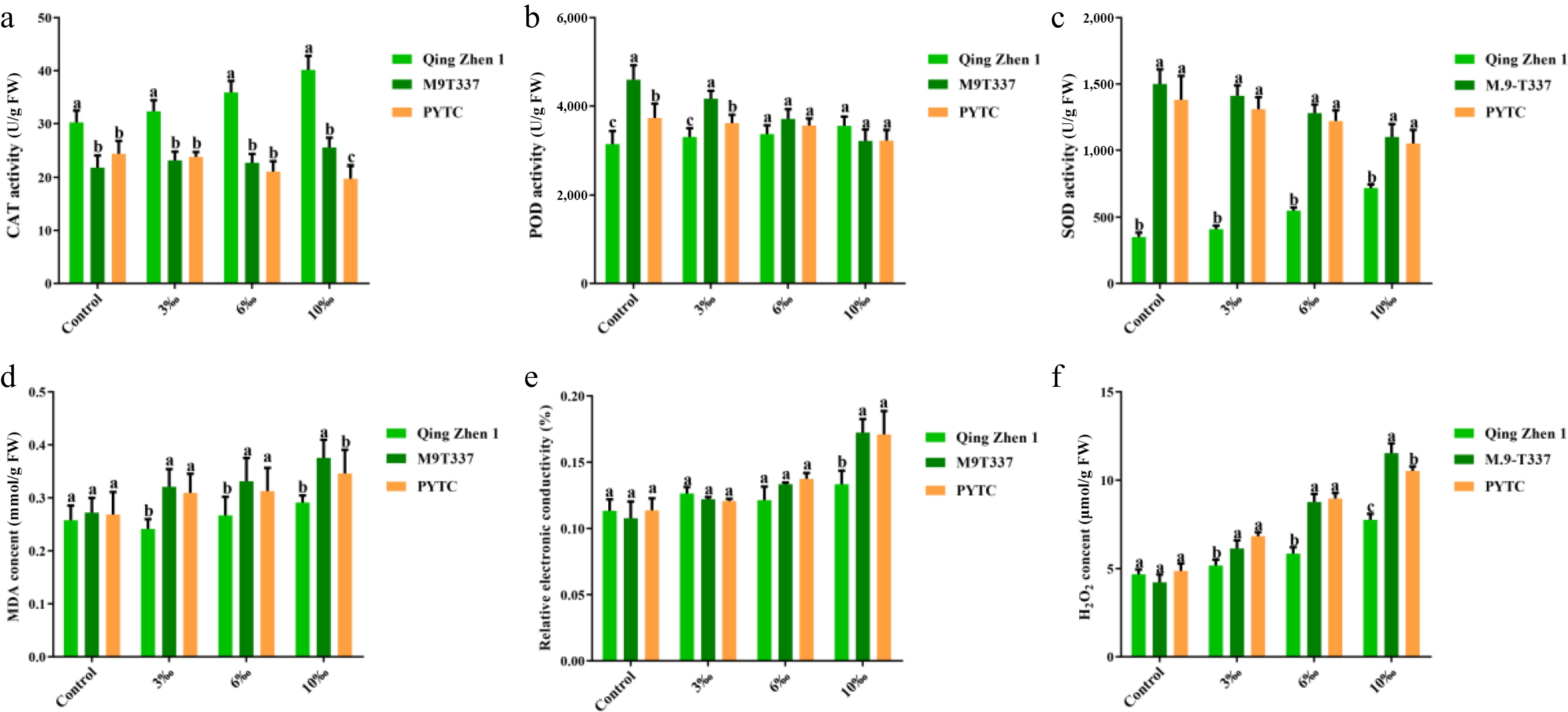
Figure 5.
Activity of antioxidant enzymes and contents of MDA and REC in plants under salt stress (2 months after transplanting into saline-alkali soil). (a) CAT activity, (b) POD activity, (c) SOD activity, (d) MDA content, (e) relative electrolytic leakage, (f) hydrogen peroxide content. The value is the average ± standard error (n ≥ 3), the letter indicates that there is a significant difference in the p < 0.05 level under each concentration treatment.
Effects of salt stress on osmoregulatory substances in plants
-
Osmoregulatory substances in plants can reflect plant resistance to some extent[38]. Under salt conditions, the starch content of Luli/Qingzhen-1 significantly increased (Fig. 6a), and the soluble protein (Fig. 6b), soluble sugar (Fig. 6c), and proline (Fig. 6d) contents in Luli/M9-T337 and Luli/PYTC also increased significantly, showing a direct correlation with salt concentration. However, the increases in Luli/M9-T337 and Luli/PYTC were more substantial, which may reflect a need to mobilize more internal nutrients to actively resist salt stress. This indicates that plants may enhance their osmotic regulation capacity by accumulating small molecular organic compounds, and that Luli/Qingzhen-1 may accumulate fewer osmotic regulatory substances, possibly because it requires fewer nutrients to resist adverse environmental factors.
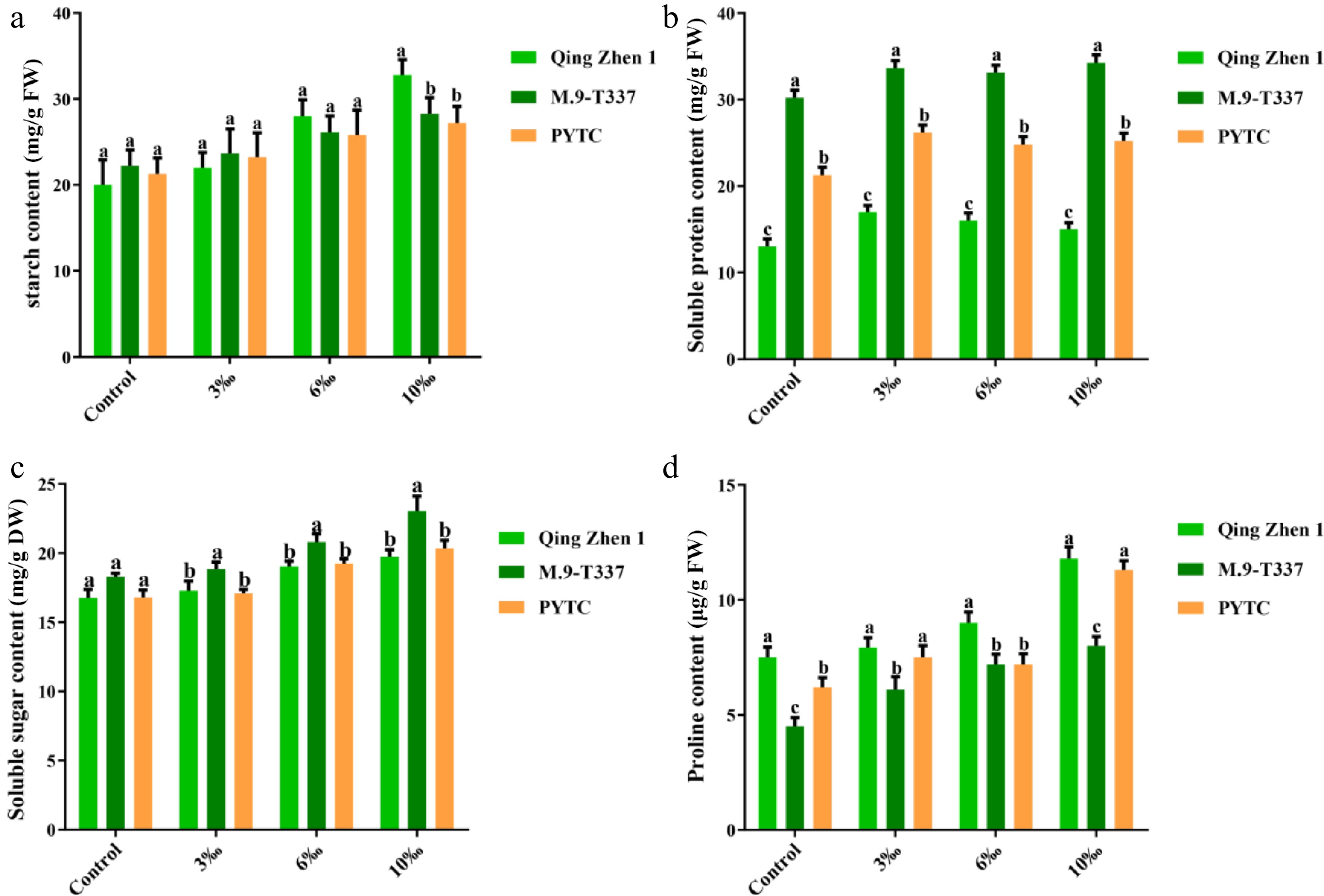
Figure 6.
Contents of osmoregulatory substances in plants under salt stress (2 months after transplanting into saline-alkali soil). (a) Starch content, (b) soluble protein content, (c) soluble sugar content, (d) proline content, the value is the average ± standard error (n ≥ 3), the letter indicates that there is a significant difference in the p < 0.05 level under each concentration treatment.
Expression of salt-resistant genes in plants under salt stress
-
Salt stress induces the expression of stress signal transduction-related genes, which ultimately activate physiological and biochemical responses[39]. To gain insight into the underlying molecular mechanisms, common key genes associated with stress responses were selected (Fig. 7), and their expression levels analyzed in different apple rootstocks by qRT-PCR. It was found that with increasing salt concentration, the expression levels of stress signal transduction-related genes were significantly upregulated. At 10‰ salt concentration, the expression levels of MdAREB1B, MdDREB2A, MdRD22, MdRD29A, MdRD29B, and MdSOS2 were significantly increased and upregulated in Luli/Qingzhen-1 (Fig. 7a−f), and MdRD22 was significantly upregulated in Luli/M9-T337 (Fig. 7c). The upregulation of MdAREB1B, MdDREB2A, and MdSOS2 was relatively minor (Fig. 7a, b, f). The expression levels of MdAREB1B, MdRD29A, MdRD29B, and MdSOS2 in Luli/PYTC were only slightly upregulated compared with the control (Fig. 7a, b, e, f). However, the degree of upregulation in some genes of Luli/M9-T337 and Luli/PYTC was lower than that in Luli/Qingzhen-1, indicating that Luli/Qingzhen-1 may resist salt stress more rapidly by significantly inducing the transcription of stress-related genes under salt stress.
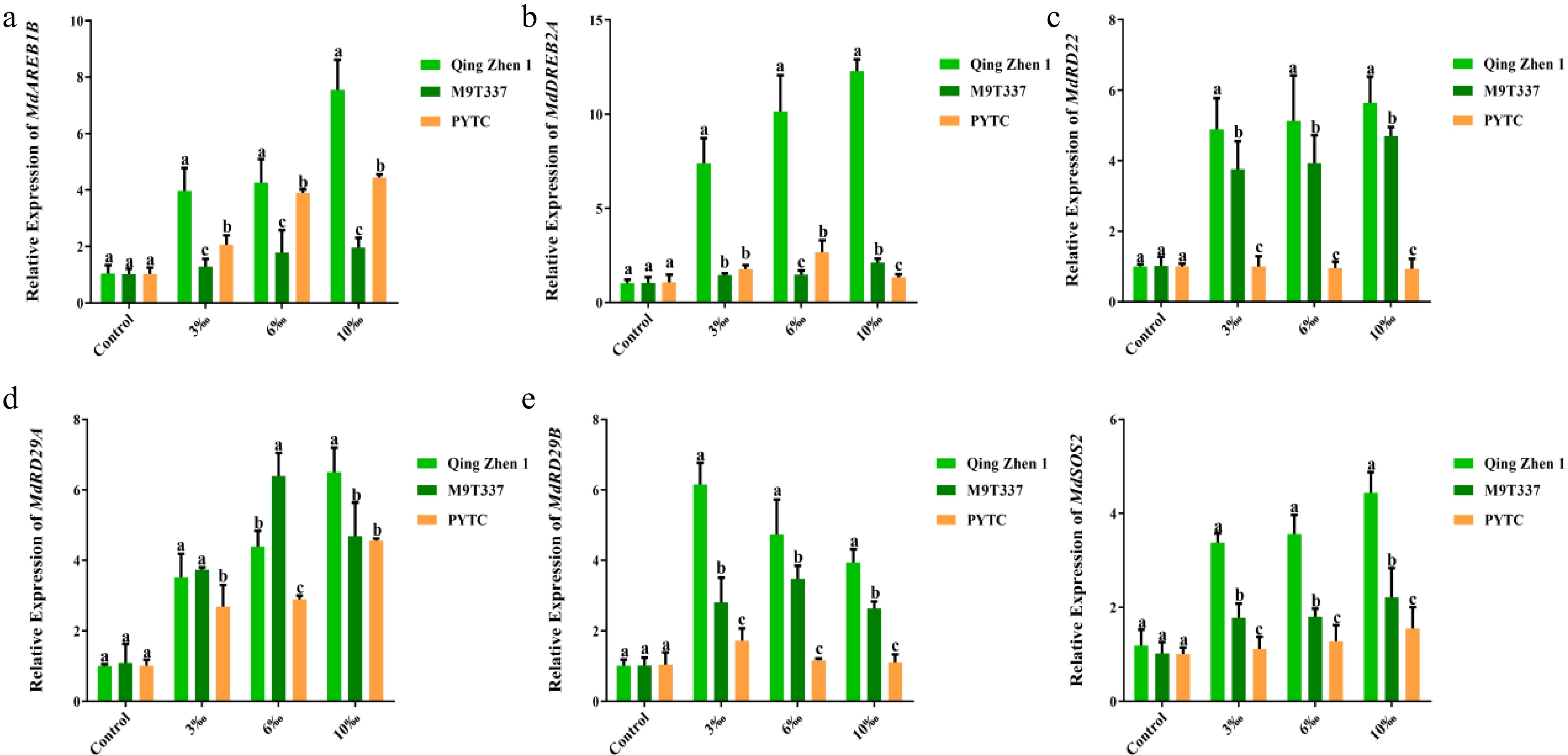
Figure 7.
Expression of related genes in plants under salt stress (2 months after transplanting into saline-alkali soil). Expression of (a) MdAREB1B, (b) MdDREB2A, (c) MdRD22, (d) MdRD29A, (e) MdRD29B, (f) MdSOS2. The value is the mean ± standard error (n ≥ 3), letters indicate significant differences at the p < 0.05 level for each concentration treatment.
Effects of salt stress on plant stomata
-
The stomata movement is also regulated by several environmental factors, such as light intensity, drought, and salt stress[40]. Stomata play a key role in gas exchange between the plant and the environment. Under normal conditions, the stomatal opening and closing of the three rootstock–scion combinations were roughly the same, with no significant differences (Fig. 8). However, under 10‰ salt stress, the stomatal closure of Luli/Qingzhen-1 was greater than that of the other two combinations. These results suggest that Luli/Qingzhen-1 may resist salt stress by regulating the degree of stomatal opening and closure.
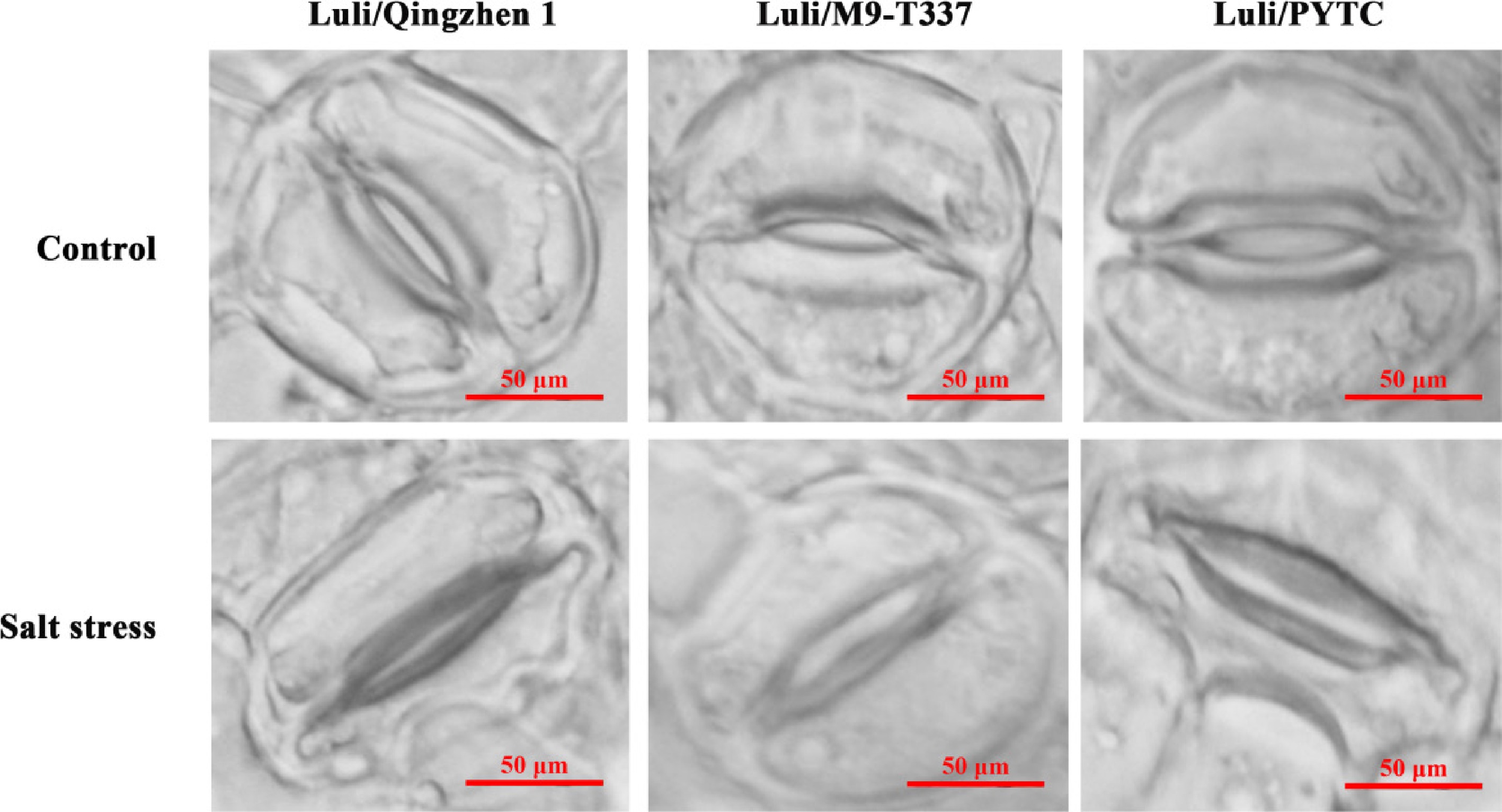
Figure 8.
Effect of salt stress on stomata opening and closing of leaves with different rootstock-scion combination. (Bar = 50 μm) (2 months after transplanting into saline-alkali soil).
Effects of salt stress on plant endogenous hormones
-
Under salt stress conditions, plants undergo specific changes in their growth hormones to cope with the stress. The reduction in growth-related hormones (such as auxins and gibberellins) helps restrict excessive growth and conserve energy. Therefore, we certain determinations of plant hormones were conducted[41]. After the salt stress phenotype was determined, leaves from the scion Luli/Qingzhen-1, which showed strong salt tolerance, were selected as the control, and the dynamic changes of endogenous hormones (ABA, IAA, GA4, ZR, JA, and SA) in leaf samples were examined (Fig. 9a−f). During salt stress treatment, ABA content was significantly higher than that of the control (Fig. 9a), while other hormones showed a decreasing trend. The variation in ABA content at each sampling point was similar to that of the control, showing an increasing trend over time. The GA4 content in the control sample increased initially, and then stabilized (Fig. 9b). IAA content increased slowly in both the control and treatment (Fig. 9c). ZR content in the control sample increased initially, and in the salt-treated sample it also showed an upward trend (Fig. 9e). JA content in the control sample fluctuated across sampling points, while it decreased under salt stress (Fig. 9d). SA content in the control sample remained relatively stable, while in salt-treated samples, SA showed a decreasing trend over time (Fig. 9f).
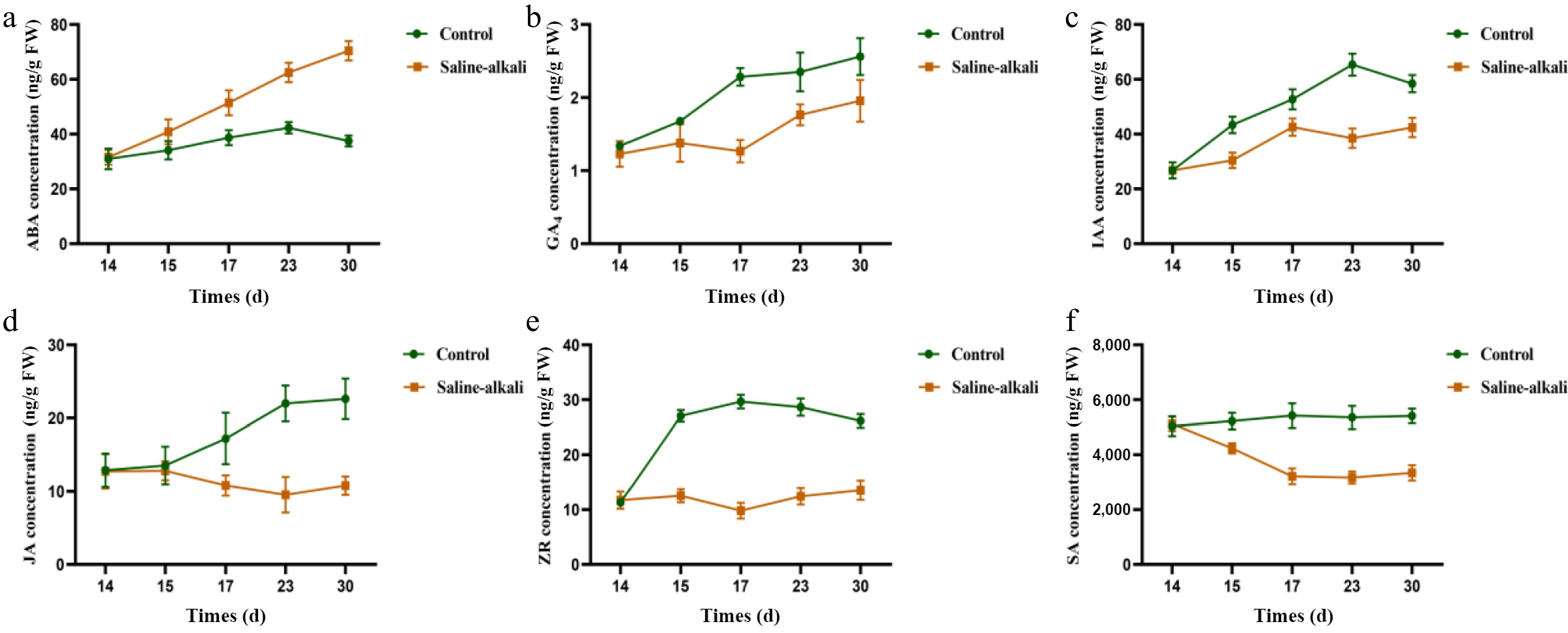
Figure 9.
Effects of more resistant rootstock hormones under salt stress (2 months after transplanting into saline-alkali soil). (a) ABA content, (b) GA4 content, (c) IAA content, (d) JA content, (e) ZR content, (f) SA content. The value is the mean ± standard error (n = 10).
Effects of salt stress on plant growth after 6 months
-
The growth of the three rootstock–scion combinations were observed and recorded at regular intervals (Fig. 10). Under normal conditions, the trees grew well and relatively uniformly. However, under 10‰ salt stress, the tree structure of Luli/Qingzhen-1 remained intact, while the plant height of Luli/M9-T337 and Luli/PYTC was about 15% lower than that of the control. The plant heights of M9-T337 and PYTC were 41% and 37% lower than the control, respectively (Fig. 10b), showing significant differences compared with the former. In addition, Luli/Qingzhen-1 also showed notable advantages in crown width (Fig. 10d), and branch length (Fig. 10e), both of which were greater than those of Luli/M9-T337 and Luli/PYTC. Luli/M9-T337 and Luli/Qingzhen-1 also had greater stem diameters, which were significantly larger than those of Luli/PYTC (Fig. 10c). As the salt concentration decreased, physiological indicators gradually improved, and tree growth was negatively correlated with salt concentration. Photosynthetic parameters in the three combinations were then assessed. The net photosynthetic rate (Fig. 10f), stomatal conductance (Fig. 10i), and transpiration rate (Fig. 10h) decreased with increasing salt concentration, while intercellular CO2 concentration (Fig. 10g) increased slightly and remained relatively stable. However, the photosynthetic rate of Luli/Qingzhen-1 remained higher than that of Luli/M9-T337 and Luli/PYTC.
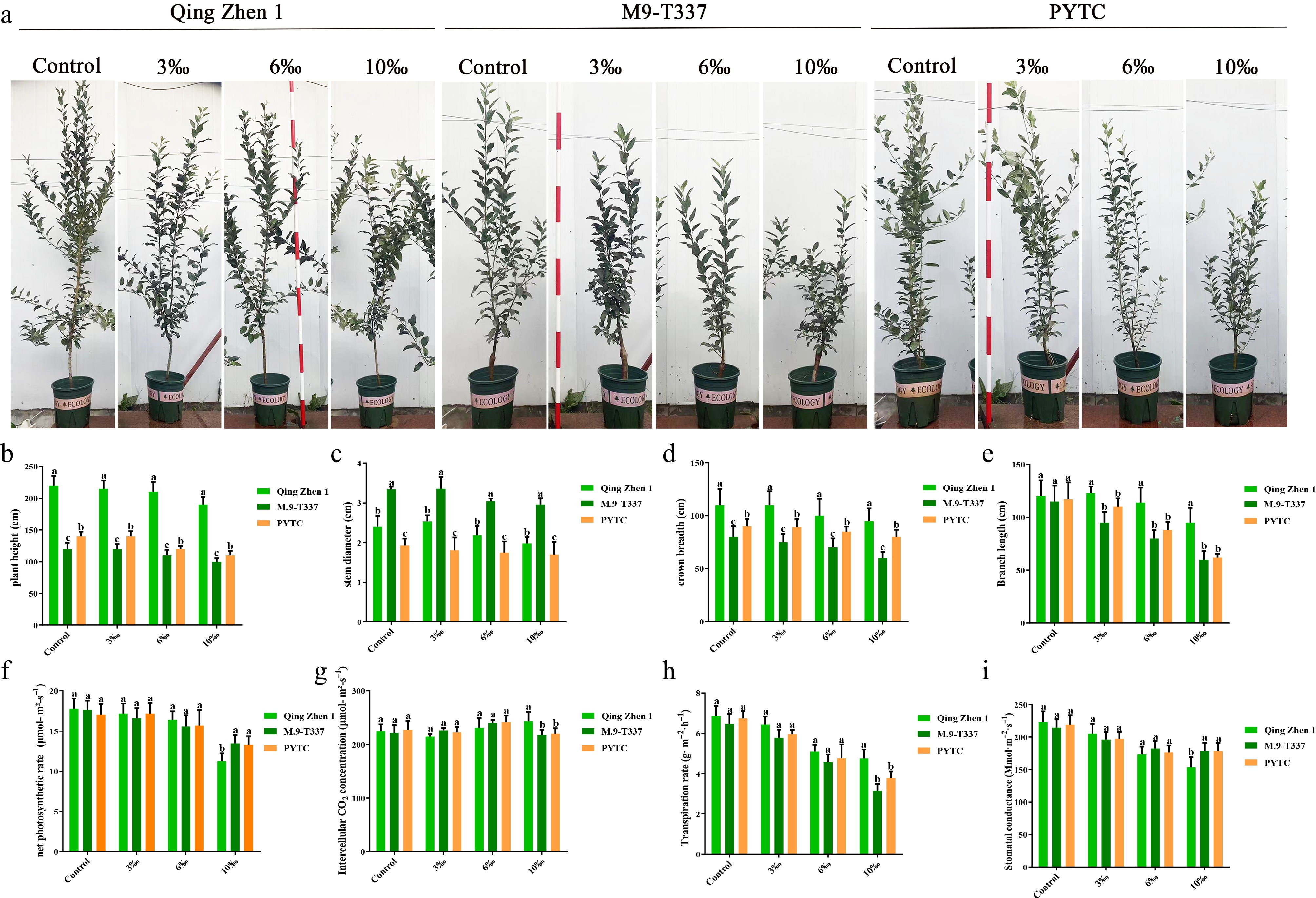
Figure 10.
Effects of salt stress on plant growth after 6 months. (a) Rootstock growth performance in later period, (b) tree height, (c) stem diameter, (d) crown width, (e) branch length, (f) net photosynthetic rate, (g) intercellular carbon dioxide concentration, (h) transpiration rate, (i) stomatal conductance. The value was the average ± standard error (n ≥ 3), letters indicate significant differences at p < 0.05 levels.
Effects of salt stress on plant roots
-
Plant roots are the first to sense harmful substances in the soil. In this study, root damage and decay under salt stress were more severe in Luli/M9-T337 and Luli/PYTC than in Luli/Qingzhen-1 (Fig. 11a), and the content of Na+ ions in the roots was positively correlated with salt concentration (Fig. 11b). The K+ ion content in roots was negatively correlated with salt concentration (Fig. 11c), likely due to excessive Na+ interfering with water uptake, resulting in water deficit and osmotic stress. The results confirmed that the root water content of Luli/Qingzhen-1 was higher, while the root water content of Luli/M9-T337 and Luli/PYTC was lower (Fig. 11d), and the root area (Fig. 11g), and root fresh weight (Fig. 11h) were significantly lower than those of Luli/Qingzhen-1. Moreover, MDA content (Fig. 11e) and relative conductivity (Fig. 11f) were significantly higher in Luli/M9-T337 and Luli/PYTC than in Luli/Qingzhen-1. These results indicate that salt stress inhibited apple rootstock growth by damaging root structure, and reducing both root and shoot biomass. Overall, Luli/Qingzhen-1 exhibited better root activity and root traits under salt stress.
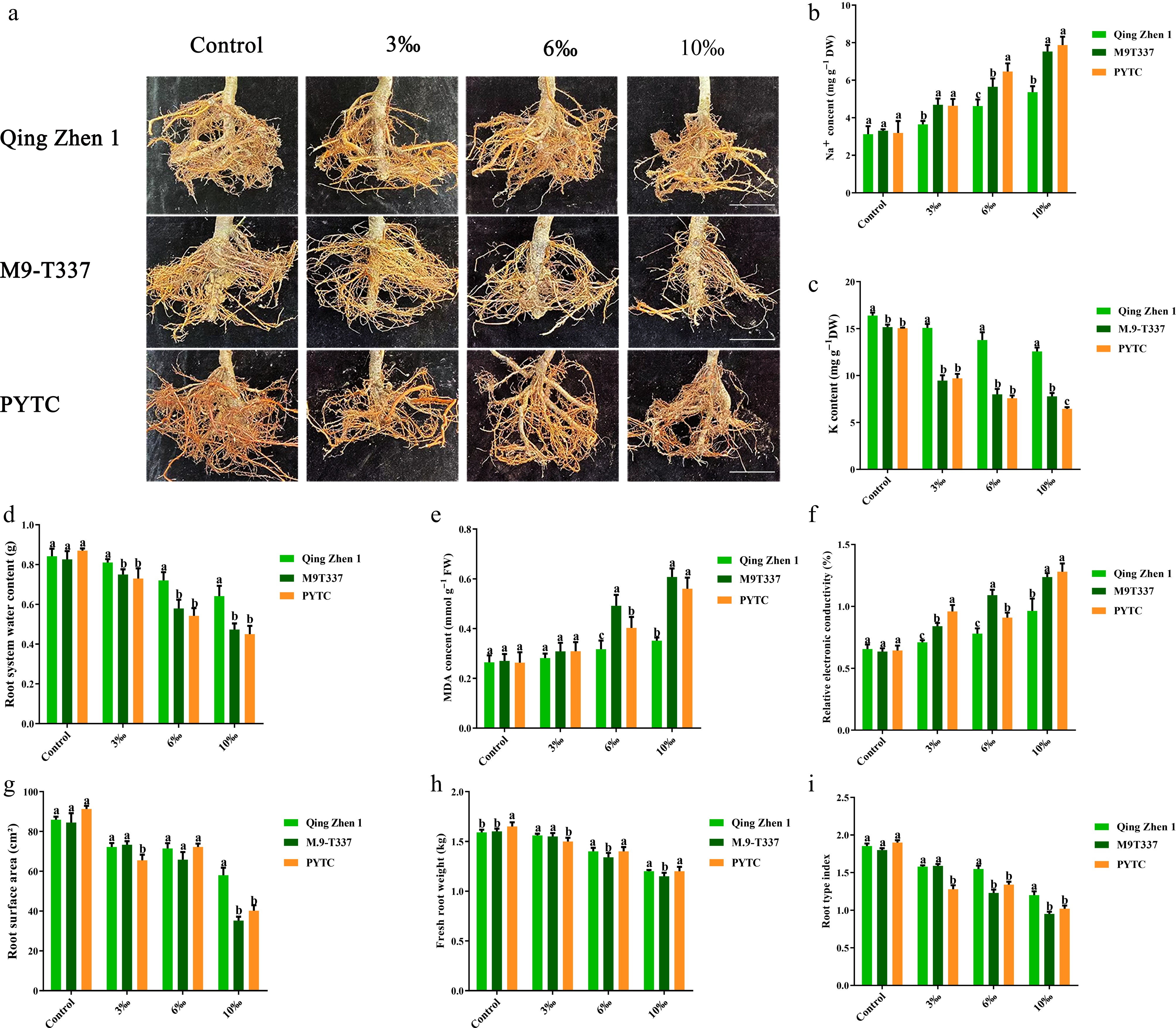
Figure 11.
Effects of salt stress on plant roots. (a) Rootstock root phenotype (effects of salt stress on plant growth after 6 months). (b) Na ion content, (c) K ion content, (d) root water content, (e) malondialdehyde content, (f) relative electrical conductivity, (g) root surface area, (h) fresh root weight, (i) plant type index. The value is the mean ± standard error (n ≥ 3), and the letters indicate significant differences at p < 0.05 levels under each concentration treatment. (Bar = 5 cm).
-
The results showed that salt stress significantly inhibited apple plant growth in terms of height, stem diameter, and crown width. At the same time, it caused leaf damage, root structure damage, plant dehydration, and, in some cases, plant death. The degree of inhibition was positively correlated with the concentration of salt stress, with variations observed among different salt-tolerant apple plants. In addition, research has shown that apple salt tolerance is closely related to rootstock varieties[42]. Apple rootstocks can be classified into three types: highly salt-tolerant, moderately salt-tolerant, and salt-sensitive. In this study, differences in growth indicators between high and low salt-tolerant rootstocks were primarily reflected in plant mortality, the extent of leaf damage, and root structure damage. The growth performance of the Qingzhen-1 rootstock-scion combination was significantly better than that of M9-T337 and PYTC, indicating that Qingzhen-1 belongs to the highly salt-tolerant group. The growth inhibition observed in saline-alkali soils may be attributed to the combined effects of saltstress (such as ionic toxicity and osmotic stress).
Under salt-alkali stress, high Na+ concentrations in the environment disrupt ion homeostasis in plant cells, leading to oxidative damage to cell membranes, ion toxicity, and metabolic disorders. Numerous studies have shown that salt stress increases Na+ accumulation in plants, resulting in osmotic stress that causes leaf water loss, reduced metabolic activity, and the exclusion of other nutrients[43,44]. In this study, as salt concentration increased, Na+ content in the rootstocks of all apple genotypes gradually rose, while K+ content showed a decreasing trend. The reduction in K+ may be linked to disruptions in osmotic balance and cell metabolism, suggesting that salt stress disturbed the Na+/K+ balance and triggered sodium ion toxicity (Fig. 4a & b). Notably, the accumulation of Na+ and the reduction of K+ in Luli/Qingzhen-1 under salt stress were relatively low, indicating reduced osmotic stress in this combination. Moreover, leaf damage (Fig. 2), root structure damage (Fig. 11), MDA content, and REC (Fig. 5d & e) in the Luli/Qingzhen-1 rootstock-scion combination were significantly lower than in Luli/M9-T337 and Luli/PYTC under moderate and high salt stress. These findings align with previous results, suggesting that Luli/Qingzhen-1 may actively resist salt stress by regulating ion balance and maintaining membrane lipid homeostasis. However, the specific mechanisms of ion uptake and utilization in this response require further investigation.
Osmoregulatory substances in plants also serve as important indicators under stress[45,46], reflecting the adaptability and resistance of plants to adverse environments[47]. To cope with osmotic stress caused by salinity, plants adjust their internal osmoregulatory substances to mitigate salt-induced damage. Under salt and alkali stress, wheat and beans resist increasing salinity by elevating levels of proline, soluble sugars, and polyol[48,49]. In this study, the contents of proline, soluble sugars, and soluble proteins in the three rootstock-scion combinations were higher than in the control group under salt stress (Fig. 6). These results indicate that all three combinations could regulate plant osmotic pressure by altering the contents of these substances. The levels of proline, soluble sugars, and soluble proteins increased significantly in the salt-tolerant rootstocks M9-T337 and PYTC. However, the increase was less pronounced in Luli/Qingzhen-1, which also exhibits high salt tolerance. This suggests that Luli/Qingzhen-1 may not require as many small organic molecules to cope with salt stress, indicating its inherently strong salt tolerance.
Stomata are the channels through which plants exchange gas with the external environment[50]. They play a key role in regulating gas exchange between the interior of the plant and the atmosphere[51]. Through stomata, plants absorb large amounts of CO2 for photosynthesis and O2−; this gas exchange is fundamental to plant life and a crucial process for maintaining ecological balance on Earth[52]. This study showed that under salt stress, the relative water content in the leaves of Luli/Qingzhen-1 was significantly higher than in those of Luli/M9-T337 and Luli/PYTC (Fig. 2i). This may be due to Luli/Qingzhen-1 regulating stomatal opening and closing under stress to control respiration and reduce water loss (Fig. 8), thereby mitigating salt stress damage. In addition, many studies have shown that ABA is closely related to stomatal movement and may be regulated by the stress-responsive hormone ABA[53]. In fact, stomatal regulation likely involves multiple plant hormones, including ethylene, gibberellin, and salicylic acid[54], a view supported by this study. Under high salt stress, Luli/Qingzhen-1 leaves exhibited reduced stomatal opening and increased ABA content (Figs 8 & 9), suggesting that ABA may act in coordination with other hormonal pathways to regulate stomatal movement and enhance plant salt tolerance.
Saline-alkali stress induces the expression of genes related to stress signal transduction and affects the physiological and biochemical responses reactions of plants[55,56]. Plants have evolved regulatory mechanisms to adapt to salt stress, which are crucial for their growth and development. To date, the roles of several stress-responsive genes in plant salt stress resistance have been extensively studied. In apples, these genes have been shown to play important roles in stress signal transduction[21,30,57]. Under salt and drought stress, the SOS2–SOS3 complex phosphorylates and activates the transport activity of the SOS1 antiporter. Previous studies have shown that MdSOS2 is expressed at varying levels across all tested organs and responds positively to salt stress[58]. In this study, it was found that salt stress induced the expression of MdSOS2, with significantly higher induction levels observed in Luli/Qingzhen-1 compared to the other two rootstock–scion combinations. This may be one of the reasons why Luli/Qingzhen-1 exhibits greater salt tolerance than Luli/M9-T337 and Luli/PYTC. In addition, several key stress-related genes are involved in the plant response to abiotic stresses. Studies have shown that the expression of MdDREB2A, MdDREB2B, MdRD29A, and MdRD29B may be up-regulated under stress conditions such as drought, salinity, temperature, and osmotic stress[59,60]. In our experiment, we observed significant up-regulation of MdRD22, MdRD29A, MdRD29B, MdDREB2A, and MdAREB1B in Luli/Qingzhen-1 (Fig. 7), indicating that this rootstock–scion combination rapidly transmits stress signals and that these stress-related genes respond strongly to external stimuli. Although these genes were also up-regulated in Luli/M9-T337 and Luli/PYTC, the extent of up-regulation was lower than in Luli/Qingzhen-1. Notably, the expression of MdDREB2A was even down-regulated in Luli/PYTC. The level of transcriptional activation of these genes positively correlated with salt resistance. The salt-tolerant rootstock Luli/Qingzhen-1 appears to more effectively induce and transmit stress signals, thereby initiating physiological and biochemical responses to actively resist salt stress.
Differences in rootstock growth vigor and root size may also impact salt tolerance, prompting us to conduct targeted research on the root systems of three rootstock-scion combinations. The three rootstocks—'Qingzhen 1' (presumed to have moderate growth potential, with undefined but functionally adaptive root traits), M9-T337 (a dwarf variety with a small root system), and Malus hupehensis* (characterized by vigorous growth, a large root system, and salt sensitivity)—exhibit significant variations in growth potential and root characteristics. These differences are likely to influence variations in salt tolerance through multiple interconnected mechanisms.
Our findings align with this inference: the moderate growth potential of 'Qingzhen 1' and its presumably balanced root size/structure correlate with superior ion homeostasis, osmotic adjustment, and photosynthetic stability under salt stress. In contrast, the large root system and high growth vigor of PYTC result in increased salt accumulation and resource allocation conflicts, while the small root system and dwarfing traits of M9-T337 reduce salt uptake but limit defensive capacity. These patterns collectively demonstrate that rootstock salt tolerance is not determined by root size or growth potential alone; rather, it is regulated by their interaction with physiological defense mechanisms, such as ion transport efficiency and flexibility in resource allocation.
The salt tolerance of the Qingzhen -1 anther-spike combination is not driven by a single isolated mechanism, but rather achieved through the synergistic action of multi-level regulatory processes spanning physiological, metabolic, and molecular dimensions. Plant responses to salt stress trigger a series of adaptive changes: First, enhanced ion homeostasis is achieved by activating sodium efflux and compartmentalization, which maintains potassium-sodium balance while reducing cytoplasmic sodium toxicity, thereby laying the foundation for cellular function maintenance (Fig. 4). Subsequently, increased synthesis of osmoprotectants (proline, soluble sugars) and upregulation of antioxidant enzyme activity mitigate osmotic stress and ROS damage, protecting membrane integrity and reducing electrolyte leakage (Figs 5, 6). The cell membrane and stomata interact physiologically, jointly regulating plant water balance, gas exchange, and stress responses. Thus, we observe that stomatal regulation mechanisms balance water loss (Fig. 8). The relationship between hormones and stomata—such as elevated ABA levels triggered by stress signaling—optimizes resource allocation between stress defense and growth maintenance (Fig. 9). Salt stress induces expression of key stress response genes, rapidly transmitting stress signals that subsequently influence plant growth and development (Fig. 7).
Overall, 'Qingzhen-1' to minimize growth inhibition under natural saline-alkali conditions, highlighting that the salt tolerance of apple rootstock-scion combinations arises from the integration of multiple pathways rather than the action of isolated traits.
-
This study analyzed and compared the salt tolerance of three apple rootstock–scion combinations. The results revealed that differences among Luli/Qingzhen-1, Luli/M9-T337, and Luli/PYTC were mainly reflected in plant mortality, leaf and root structure damage, ion content, membrane lipid peroxidation, osmotic adjustment substances, endogenous hormone levels, and the expression of stress signal transduction-related genes induced by salt stress. The rootstocks identified in this experiment provide a reference not only for selecting and breeding salt-tolerant combinations, but also for germplasm innovation. These findings are of both strategic and practical significance for safeguarding and promoting the sustainable development of China's modern apple industry. Moreover, they offer important support for advancing high-quality, efficient, and distinctive agricultural practices and accelerating the development of a strong agricultural nation.
This work was supported by the National Key Research and Development Program of China (2022YFD1201700), National Natural Science Foundation of China (32172538), and thw Key Research and Development Plan (Major Scientific and Technological Innovation Project) of Shandong Province (2022LZGC010).
-
The authors confirm their contributions to the paper as follows: conceptualization, writing - original draft: Chen GL; investigation: Chen GL, Liu JY, Wang HB; formal analysis: Chen GL, Wang DR, Liu X, Liu HF; methodology, validation: Wang DR, Liu X; data curation: Liu X, Li L, Wang S; software: Li L, Qu MS, Liu JY; visualization: Qu MS; supervision: Li LG, You CX; resources, writing - review & editing: Chen GL, Wang X, Li LG, You CX; funding acquisition: You CX. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers for quantitative real-time qRT-PCR.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen GL, Wang DR, Liu X, Wang X, Li L, et al. 2025. Evaluation of salt stress tolerance of three apple genotypes rootstock combinations in 'Luli' apple. Fruit Research 5: e042 doi: 10.48130/frures-0025-0033 

Evaluation of salt stress tolerance of three apple genotypes rootstock combinations in 'Luli' apple
- Received: 08 July 2025
- Revised: 29 August 2025
- Accepted: 11 September 2025
- Published online: 27 November 2025
Abstract: In rootstock-scion combinations, the rootstock is the key factor determining a plant's salt tolerance, primarily through its root physiology and soil interaction capabilities. However, the interaction between rootstock and scion in influencing salt tolerance in apple trees remains underexplored. In this study, three one-year-old dwarf rootstocks—'Qingzhen-1', 'M9-T337', and '(Malus hupehensis Rehd, var. Pingyiensis, PYTC)'—were selected, with 'Luli' used as the scion. The salt tolerance was compared in natural soil (1‰), and saline-alkali soil from Dongying (3‰, 6‰, 10‰). The results showed that salt stress inhibited plant growth and damaged root structure. Among the combinations, Luli/Qingzhen-1 exhibited superior growth performance, with reduced malondialdehyde content, relative conductivity, and mortality. Additionally, in the Luli/Qingzhen-1 combination, genes associated with antioxidant enzyme activity and stress signal transduction were significantly upregulated. In conclusion, Qingzhen-1 demonstrates greater salt tolerance than M9-T337 and PYTC, making it a promising rootstock for cultivating 'Luli' scions in saline-alkali soils.
-
Key words:
- Apple rootstock /
- Salt stress /
- Growth change /
- Physiological mechanism /
- Comprehensive evaluation