









-
A. annua is an important natural source of artemisinin, a sesquiterpene lactone that forms the backbone of frontline antimalarial therapies[1]. This compound is both synthesized and accumulated predominantly within the glandular secretory trichomes (GSTs)[2]. While metabolic pathways and key enzymes involved in artemisinin biosynthesis have been extensively characterized[3−5], the mechanisms governing the intracellular transport and secretion of hydrophobic intermediates (e.g., dihydroartemisinic acid) within GSTs remain poorly understood. Non-specific lipid transfer proteins (nsLTPs), which facilitate the binding and transport of lipophilic molecules, represent promising candidates for mediating these processes[6]. However, despite their potential roles in secondary metabolite trafficking and stress adaptation, the nsLTP family in A. annua has not been systematically investigated.
Plant non-specific lipid transfer proteins (nsLTPs) are a group of small secretory proteins in plants that are responsible for transferring phospholipids from liposomes to mitochondria or chloroplasts[6]. All known plant nsLTPs are synthesized as precursors with N-terminal signal peptides. Mature proteins are typically small, with molecular weights usually below 10 kDa[7], and feature a characteristic eight-cysteine motif (ECM) formed by eight cysteine residues arranged as C-Xn-C-Xn-CC-Xn-CXC-Xn-C-Xn-C[8]. The ECM is stabilized by four disulfide bonds, which maintain the three-dimensional structure of the hydrophobic cavity. Various lipids and hydrophobic compounds were shown to bind this structure via in vitro assays[9].
nsLTPs were initially thought to be involved in membrane biosynthesis and regulation of intracellular fatty acid pools due to their ability to transfer lipid molecules and bind acyl chains in vitro[7]. However, the presence of N-terminal signal peptides in all nsLTP precursors suggests their involvement in the secretory pathway. In Arabidopsis, LTPG, a glycosylphosphatidylinositol-anchored lipid transfer protein, is highly expressed in the epidermis of inflorescence stems. It primarily localizes to the plasma membrane, binds lipid probes in vitro, and may play a role in lipid transport in the cuticle. Reduced wax deposition in the stem cuticle was observed in ltpg mutants[10]. Current research has proposed multiple physiological functions for nsLTPs, including plant signaling[11,12], cuticle and wax metabolism[13], cell wall extension[14], pollen development[15−17], somatic embryogenesis[18,19], seed germination[20], response to biotic stresses[21,22] and abiotic stresses[23−26]. For example, nsLTPs have been shown to mediate responses to pathogen attack by interacting with various plant defense pathways[27], while their roles in drought and salinity stress tolerance have been demonstrated through their ability to modulate lipid signaling and cell membrane integrity[28−30]. Nevertheless, a genome-wide analysis of nsLTPs in A. annua is lacking, leaving their roles in artemisinin biosynthesis unresolved.
Previous work in A. annua identified only four nsLTPs (AaLTP1–4), with functional studies limited to heterologous expression systems. AaLTP1, AaLTP2, and AaLTP3 have been shown to enhance the accumulation of (DH)AA in the apoplast of Nicotiana benthamiana leaves, while AaLTP3 and AaLTP4 are involved in sesquiterpene lactone secretion from glandular trichomes in A. annua[31]. However, these studies did not address whether nsLTPs directly influence artemisinin biosynthesis in planta or how their expression is regulated. Moreover, the recent release of the A. annua genome provides an opportunity to comprehensively characterize the nsLTP family and uncover trichome-specific isoforms with specialized roles in artemisinin metabolism[32].
Here, this study addresses these gaps through a genome-wide identification and functional analysis of nsLTPs in A. annua. The objectives were to: (1) systematically classify the nsLTP family; (2) identify isoforms preferentially expressed in glandular trichomes; (3) validate roles in artemisinin biosynthesis; and (4) elucidate upstream regulatory mechanisms. This study performed a genome-wide analysis of the nsLTP gene family, identifying 55 nsLTP genes in A. annua and classifying them into eight types. Protein structures, chemical characteristics, structural motifs, subcellular localization, evolutionary patterns, and expression profiles were examined. Specifically, two AaLTP genes, AaLTP1 and AaLTP2, were cloned and confirmed to exhibit glandular trichome-specific expression. The homeodomain–leucine zipper (HD-Zip) proteins participate in various critical physiological processes, including transcriptional regulation of epidermal and sub-epidermal cell formation, anthocyanin accumulation, wax synthesis, lipid transport, and response to drought signals[33,34]. Interestingly, AaLTP1 and AaLTP2 were discovered to be positively regulated by AaHD8. Moreover, overexpression of AaLTP1 reveals a positive role in artemisinin production. This study provides a comprehensive understanding of the nsLTP gene family in A. annua, highlighting structural and functional diversity, evolutionary aspects, and regulatory mechanisms, particularly in relation to glandular trichome biology and artemisinin biosynthesis.
-
The high artemisinin variety 'Hu-Hao 1' of A. annua L., derived from Chongqing and further developed over several years in Shanghai, is used in all experiments related to A. annua[35,36]. For in vitro culture, A. annua seeds are initially surface-sterilized by immersion in 70% ethanol for 3 min, followed by washing three times with sterile water. Subsequently, the seeds are sterilized in a 20% sodium hypochlorite solution for 10 min and rinsed five times with sterile water. These sterilized seeds are then sown on MS medium containing 3% sucrose and 0.6% agar (pH = 5.7). A. annua plants are grown in pots at a temperature of 23 ± 2 °C under a photoperiod of 16 h light and 8 h darkness[37].
Identification and bioinformatics analysis
-
The protein data of A. annua were obtained from the NCBI database (
www.ncbi.nlm.nih.gov/genome ) (taxonomic ID: 35608). The HMMER-3.2.1 software (www.hmmer.org ) was used to identify proteins containing the LTP domain (accession: PF00234) (plant lipid transfer/seed storage/trypsin-alpha amylase inhibitor). The web tool SignalP 5.0 (https://services.healthtech.dtu.dk/services/SignalP-5.0 ) was employed to predict mature proteins. Sequences longer than 120 amino acids in mature proteins were discarded, and the presence of ECM was manually checked. The remaining candidate proteins were further confirmed for the LTP domain using the batch Web CD-Search tool (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ). The molecular weight and isoelectric point (pI) of AaLTP were calculated using the compute pI/Mw tool (https://web.expasy.org/compute_pi ). The subcellular localization of AaLTP was predicted using DeepLoc-1.0 (www.cbs.dtu.dk/services/DeepLoc-1.0/index.php )[38]. Motif analysis was performed using MEME (https://meme-suite.org/meme/tools/meme ). Promoter analysis was conducted using PlantPAN 3.0 (https://plantpan.itps.ncku.edu.tw/plantpan4/promoter_analysis.php ). The three-dimensional structure of AaLTP was predicted using Phyre2 (www.sbg.bio.ic.ac.uk/phyre2/html )[39]. The coding sequence of the AaLTP gene is shown in Supplementary Table S1.Alignment and phylogenetic analysis
-
Protein sequences of A. annua and A. thaliana were downloaded from NCBI (
www.ncbi.nlm.nih.gov ) and TAIR (www.arabidopsis.org ), respectively. Multiple sequence alignment of the ECM domain was performed using MAFFT and manually adjusted[40]. The results of the multiple sequence alignment were visualized using Jalview software. Using MEGA 7.0 software, maximum likelihood phylogenetic trees were constructed for A. annua under the Complete deletion and LG + G + I model, and for A. annua and A. thaliana under the Complete deletion and WAG + G + I model[41]. To validate reliability, 1,000 bootstrap replicates were performed. The above-mentioned best-fit model was selected using the 'Find Best DNA/Protein Models' function built into MEGA 7.0, with parameters set to 'Complete deletion' for gap treatment and 'Very Strong' for the branch swap filter to ensure the highest accuracy. Further tree beautification was done using iTOL (http://itol.embl.de ) and Figtree software.Expression analysis of nsLTPs
-
RPKM (Reads Per Kilobase of gene model per Million) values of AaLTPs in various organs (root, stem, leaf, seed, bud, and trichome) were extracted from RNA-seq data of A. annua (SRP129502)[42]. TPM (Transcripts Per Kilobase Million) values of all differentially expressed genes (DEGs) from AaHD8-RNAi lines were extracted from RNA-seq data (Supplementary Table S2)[43]. Subsequently, differential expression analysis of different organs and AaHD8-RNAi lines was performed using the online tool imageGP (
www.ehbio.com/ImageGP/index.php ).Cloning procedures and transformation
-
The full-length coding sequences of AaLTP1 and AaLTP2 were amplified from A. annua cDNA using primers LTP1-F/R and LTP2-F/R, respectively, with engineered BamH I and Xba I restriction sites flanking both sequences. Following sequencing verification, the digested PCR products were directionally cloned into the PHB plant expression vector to generate EYFP fusion constructs under the control of the CaMV35S promoter, yielding recombinant plasmids pHB-AaLTP1 and pHB-AaLTP2. For promoter activity analysis, the upstream regulatory regions of AaLTP1 and AaLTP2 were amplified using primers listed in Supplementary Table S3 and cloned into the pCambia 1391Z vector to drive GUS expression.
All constructs (pHB-AaLTP1, pHB-AaLTP2, and promoter-GUS fusions) were introduced into Agrobacterium tumefaciens strain EHA105 via electroporation. A. annua leaf explants were transformed using established Agrobacterium-mediated methods[44]. For overexpression studies, transgenic lines harboring pHB-AaLTP1 or pHB-AaLTP2 were screened by PCR amplification of genomic DNA using primers spanning the vector-AaLTP junctions. For GUS assays, plants transformed with promoter-GUS constructs (or the empty pCambia 1391Z vector as negative controls) were subjected to histochemical staining.
GUS expression in transgenic A. annua plants
-
GUS staining was performed by incubating leaves and stems in staining solution (0.1 M sodium phosphate buffer [pH 7.0], 10 mM EDTA, 0.5 mM potassium ferrocyanide, 0.5 mM potassium ferricyanide, 1 mg/mL X-Gluc) at 37 °C in darkness for 12 h. Chlorophyll was removed by replacing the staining solution with 70% ethanol, followed by repeated ethanol washes until complete tissue decolorization.
Measurement of artemisinin content
-
A. annua leaves were collected for the test. Leaf samples were dried at 50 °C for 48 h and then ground into powder. The leaf powder was extracted twice with 2 ml of methanol using ultrasonication, with each sonication step lasting 30 min[45]. After extraction, the supernatant was centrifuged for 5 min and filtered through a 0.22 μm nitrocellulose membrane filter. Artemisinin content was measured using High Performance Liquid Chromatography (HPLC), following procedures described in previous literature[44].
RNA extraction and qRT-PCR analysis
-
Total RNA was extracted using the RNAprep Pure Plant Kit (Tiangen, Beijing, China) following the manufacturer's instructions. cDNA was synthesized using PrimeScript™ RT Master Mix (Takara, Shiga, Japan). Quantitative RT-PCR analysis was performed following procedures described in previous literature. The transcription levels of the genes were normalized using the A. annua Actin gene as an internal reference. All primers used are listed in Supplementary Table S3.
Yeast one hybrid assays
-
The full-length coding sequences of AaHD8 were amplified using primers listed in Supplementary Table S3 and cloned into the pB42AD vector. Two concatenated L1-box-like motifs from the AaLTP1 and AaLTP2 promoters were inserted into pLacZ. The effector and reporter plasmids were co-transformed into the yeast strain EGY48a. The empty pB42AD vector was used as a control. Transformed yeast cells were cultured on SD/-Trp/-Ura medium. Positive clones were transferred to SD/-Trp/-Ura medium containing X-β-gal and cultured until a blue reaction appeared. All primers used for amplifying promoters and DNA motifs are listed in Supplementary Table S3.
Electrophoretic mobility shift assays
-
The full-length coding sequence of AaHD8 was cloned into the pCold-TF vector (Takara) to generate a fusion protein with a His tag for protein expression and purification purposes. The pCold-AaHD8 construct was transformed into Escherichia coli Rosetta (DE3) strain (TransGen Biotech, China). An empty pCold-TF vector was also introduced into Escherichia coli Rosetta (DE3) as a negative control. Protein expression was induced by adding 0.5 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) to the culture medium, and the induction was carried out at 16 °C for 14 h. The fusion protein was purified using Ni-NTA (Nickel-nitrilotriacetic acid) agarose (Invitrogen, USA) according to the manufacturer's instructions.
Electrophoretic Mobility Shift Assay (EMSA) was performed using biotin-labeled DNA probes as previously described and following the manufacturer's instructions[35]. Each reaction included 10 μg of recombinant His-TF and His-HD8. DNA probes AaLTP1-L1 and AaLTP2-L1 from the AaLTP1 and AaLTP2 promoters were synthesized by Sangon (Shanghai, China). Primers and probes used in the EMSA are listed in Supplementary Table S3.
-
To identify the complete set of potential nsLTP genes in A. annua, a Hidden Markov Model (HMM) based on the nsLTP domain (PF00234) was utilized to search the A. annua proteome using HMMER-3.2.1 software. Initially, 116 proteins were retrieved. These sequences were then screened using SignalP to predict N-terminal signal peptide sequences (NSS), leading to the exclusion of six proteins lacking NSS. As mature nsLTP proteins typically have low molecular weights, 34 predicted mature proteins with more than 120 amino acids were further removed. Additionally, two proteins lacking the characteristic cysteine residue pattern were discarded. Proteins were then analyzed with the Web CD-Search tool to confirm the conserved domain, excluding 11 proteins lacking the LTP or AAI_LTSS structure. Finally, eight highly redundant sequences were removed through alignment and manual inspection, resulting in a total of 55 confirmed nsLTP genes in A. annua (Table 1).
Table 1. Putative nsLTP genes identified in the genome of A. annua L.
Name Gene ID CDS length (bp) AAa SPb SLc MPd (AA) ECMe MP (MWf) MP (pIg) Type I AaLtpI.1 Super-scaffold_
100123g01462191351 116 25 s 91 C-X9-C-X13-CC-X19-CXC-X22-C-X13-C 9,106.48 9 AaLtpI.2 chr2g00359671 345 114 25 s 89 C-X9-C-X13-CC-X17-CXC-X22-C-X13-C 9,016.44 8.87 AaLtpI.3 chr2g00359701 333 110 25 s 85 C-X9-C-X13-CC-X17-CXC-X22-C-X13-C 8,599.97 8.7 AaLtpI.4 chr3g00499441 345 114 25 s 89 C-X9-C-X13-CC-X19-CXC-X21-C-X12-C 9,076.39 9.05 AaLtpI.5 chr1g00152381 411 136 24 s 112 C-X9-C-X13-CC-X19-CXC-X21-C-X13-C 12,177.13 5.54 AaLtpI.6 chr2g00297041 351 116 25 s 91 C-X9-C-X13-CC-X19-CXC-X22-C-X13-C 9,439.93 9.19 AaLtpI.7 chr2g00297131 351 116 25 s 91 C-X9-C-X13-CC-X19-CXC-X22-C-X13-C 9,198.68 9.57 AaLtpI.8 chr2g00297171 348 115 25 s 90 C-X9-C-X13-CC-X19-CXC-X21-C-X13-C 9,019.42 9.26 AaLtpI.9 chr2g00297151 351 116 25 s 91 C-X9-C-X13-CC-X19-CXC-X22-C-X13-C 9,092.46 9.3 AaLtpI.10 chr2g00297111 354 117 25 s 92 C-X9-C-X13-CC-X19-CXC-X22-C-X13-C 9,763.21 8.66 AaLtpI.11 chr3g00546061 369 122 29 s 93 C-X9-C-X16-CC-X19-CXC-X21-C-X13-C 9,745.02 3.72 AaLtpI.12 chr2g00297121 342 113 19 s 94 C-X9-C-X13-CC-X19-CXC-X22-C-X13-C 10,230.87 8.67 AaLtpI.13 chr1g00017271 360 119 27 s 92 C-X9-C-X14-CC-X19-CXC-X21-C-X13-C 9,043.43 8.89 AaLtpI.14 chr2g00309551 354 117 24 s 93 C-X9-C-X14-CC-X19-CXC-X22-C-X13-C 9,682.74 4.14 AaLtpI.15 chr4g00612411 369 122 29 s 93 C-X9-C-X14-CC-X19-CXC-X22-C-X13-C 10,033.22 4.76 AaLtpI.16 chr6g01084791 363 120 28 s 92 C-X9-C-X14-CC-X20-CXC-X21-C-X13-C 9,795.49 7.69 AaLtpI.17 chr2g00297031 360 119 22 s 97 C-X9-C-X15-CC-X19-CXC-X22-C-X12-C 10,454.11 9.3 AaLtpI.18 unctg_3864g01516351 390 129 24 s 105 C-X9-C-X16-CC-X19-CXC-X23-C-X12-C 11,775.69 8.46 AaLtpI.19 chr2g00297011 414 137 25 s 112 C-X9-C-X14-CC-X19-CXC-X22-C-X12-C 11,964.69 9.09 AaLtpI.20 chr3g00499431 348 115 25 s 90 C-X9-C-X13-CC-X19-CXC-X21-C-X13-C 9,170.5 9.16 AaLtpI.21 chr2g00297081 393 130 25 s 105 C-X9-C-X13-CC-X19-CXC-X22-C-X13-C 11,565.33 8.65 Type II AaLtpII.1 chr1g00100031 294 97 27 s 70 C-X7-C-X13-CC-X8-CXC-X19-C-X3-C 7,771.9 8.47 AaLtpII.2 chr1g00099971 297 98 25 s 73 C-X7-C-X13-CC-X8-CXC-X20-C-X2-C 7,861.2 5.06 AaLtpII.3 chr1g00099961 285 94 26 s 68 C-X7-C-X13-CC-X8-CXC-X23-C-X6-C 7,196.39 8.88 AaLtpII.4 chr5g00892001 288 95 27 s 68 C-X7-C-X13-CC-X8-CXC-X23-C-X6-C 7,424.05 9.96 AaLtpII.5 chr1g00099991 294 97 27 s 70 C-X7-C-X13-CC-X8-CXC-X23-C-X6-C 7,999.08 8.95 AaLtpII.6 chr3g00577871 291 96 25 s 71 C-X7-C-X13-CC-X8-CXC-X23-C-X6-C 7,771.01 8.5 AaLtpII.7 chr6g01008841 363 120 25 s 95 C-X7-C-X16-CC-X9-CXC-X23-C-X9-C 10,504.09 4.75 AaLtpII.8 chr3g00577891 294 97 29 s 68 C-X7-C-X13-CC-X8-CXC-X23-C-X6-C 7,165.38 8.91 AaLtpII.9 chr9g01375721 294 97 28 s 69 C-X7-C-X13-CC-X8-CXC-X23-C-X6-C 7,633.87 8.89 AaLtpII.10 chr5g00891931 294 97 29 s 68 C-X7-C-X13-CC-X8-CXC-X23-C-X6-C 7,245.69 9.41 AaLtpII.11 chr3g00483861 300 99 30 s 69 C-X7-C-X14-CC-X8-CXC-X23-C-X6-C 7,522.63 7.74 Type III AaLtpIII.1 unctg_5142g01560471 276 91 27 s 64 C-X9-C-X16-CC-X9-CXC-X12-C-X5-C 6,821.95 5.43 AaLtpIII.2 chr3g00559411 282 93 27 s 66 C-X9-C-X17-CC-X9-CXC-X12-C-X5-C 7,061.03 5.43 Type IV AaLtpIV.1 chr2g00348561 321 106 27 s 79 C-X9-C-X17-CC-X9-CXC-X17-C-X6-C 8,505.87 6.7 AaLtpIV.2 chr8g01299581 294 97 22 s 75 C-X9-C-X15-CC-X9-CXC-X24-C-X6-C 7,822.12 4.34 Type V AaLtpV.1 chr1g00206751 363 120 29 s 91 C-X14-C-X14-CC-X11-CXC-X16-C-X7-C 9,915.79 9.57 AaLtpV.2 chr1g00206761 378 125 28 s 97 C-X14-C-X14-CC-X11-CXC-X24-C-X10-C 10,313.99 8.62 AaLtpV.3 chr3g00569011 357 118 27 s 91 C-X14-C-X14-CC-X11-CXC-X24-C-X10-C 9,379.82 8.18 AaLtpV.4 chr5g00787181 363 120 28 s 92 C-X14-C-X14-CC-X12-CXC-X24-C-X10-C 9,418.59 10.4 Type VI AaLtpVI.1 chr6g01040821 318 105 22 s 83 C-X10-C-X12-CC-X9-CXC-X22-C-X9-C 9,389.32 10.33 AaLtpVI.2 chr6g01040781 318 105 22 s 83 C-X10-C-X12-CC-X9-CXC-X22-C-X9-C 9,111.7 9.45 AaLtpVI.3 chr1g00008331 336 111 29 s 82 C-X10-C-X16-CC-X9-CXC-X22-C-X9-C 8,667.06 6.71 AaLtpVI.4 chr6g01040811 333 110 25 s 85 C-X10-C-X12-CC-X9-CXC-X23-C-X9-C 9,188.67 8.53 Type VII AaLtpVII.1 chr1g00063881 333 110 27 s 83 C-X9-C-X14-CC-X12-CXC-X18-C-X7-C 8,888.1 4.39 AaLtpVII.2 chr1g00063861 330 109 27 s 82 C-X9-C-X14-CC-X12-CXC-X18-C-X7-C 8,722.86 4.25 AaLtpVII.3 chr2g00408761 423 140 26 s 114 C-X9-C-X14-CC-X12-CXC-X25-C-X9-C 12,044.73 4.51 AaLtpVII.4 chr2g00415681 345 114 23 s 91 C-X9-C-X14-CC-X12-CXC-X26-C-X9-C 9,136.58 8.45 Type IX AaLtpIX.1 chr2g00241771 339 112 24 s 88 C-X13-C-X15-CC-X9-CXC-X22-C-X6-C 9,369.99 8.21 nsLTPy nsLTPy.1 chr4g00736531 270 89 26 s 63 C-X8-C-X13-CC-X8-CXC-X17-C-X6-C 6,862.86 4.59 nsLTPy.2 chr4g00736521 273 90 27 s 63 C-X8-C-X13-CC-X8-CXC-X17-C-X6-C 6,910.89 4.56 nsLTPy.3 chr2g00265921 399 132 26 s 106 C-X9-C-X12-CC-X19-CXC-X19-C-X14-C 12,314.22 8.44 nsLTPy.4 chr2g00265911 405 134 25 s 109 C-X10-C-X13-CC-X19-CXC-X21-C-X14-C 12,409.34 8.98 nsLTPy.5 chr4g00694091 423 140 26 s 114 C-X9-C-X22-CC-X19-CXC-X25-C-X14-C 12,533.12 9.44 nsLTPy.6 chr2g00265901 423 140 25 s 115 C-X9-C-X12-CC-X19-CXC-X21-C-X14-C 13,023.16 8.44 a AA, number of amino acids; b SP, signal peptide; c SL, subcellular location, s = secretory pathway; d MP, mature protein; e ECM, eight cysteine motif; f MW, molecular weight in Dalton; g pI, isoelectric point. The detailed classification basis is listed in Supplementary Table S4. These 55 AaLTPs were classified into eight types (I, II, III, IV, V, VI, VII, and IX) following the criteria established by Boutrot et al. (Table 1)[46], which is completely based on protein sequence similarity. This classification was manually curated without the use of automated tools. Given that LTPs are small proteins with conserved features, the established criteria were largely applicable to A. annua. A minor adjustment was made for A. annua-specific sequences: individuals with 17 or 20 amino acid residues between the fourth and fifth conserved cysteine residues were also categorized as type I, based on their high sequence similarity and phylogenetic analysis. The detailed classification basis is listed in Supplementary Table S4. All AaLTPs possess a signal peptide of 19−30 amino acids. It was found through the calculation tool DeepLoc-1.0 that, with the exception of AaLTPVII.3, which is predicted to localize to the cell membrane, the others are expected to be secreted into the extracellular space (Supplementary Table S5). The predicted molecular weights of the mature proteins range from 6,821 to 13,023 Da. Theoretical isoelectric point (pI) calculations show that 39 proteins have basic pI values (7.69−10.4), while the remaining proteins have acidic pI values (3.72−6.71). Predicted 3D structures are visualized in Supplementary Fig. S1. The ECM in all AaLTPs is highly conserved, forming four disulfide bonds that stabilize the hydrophobic cavity, similar to findings in rice, wheat, rape, and Arabidopsis[46,47]. Multiple sequence alignment of mature AaLTPs using ClustalX and visualizing with Jalview software (Fig. 1) demonstrated highly conserved eight cysteine residues and highlighted the diversity of the ECM domain and adjacent amino acid residues in A. annua (Table 2).
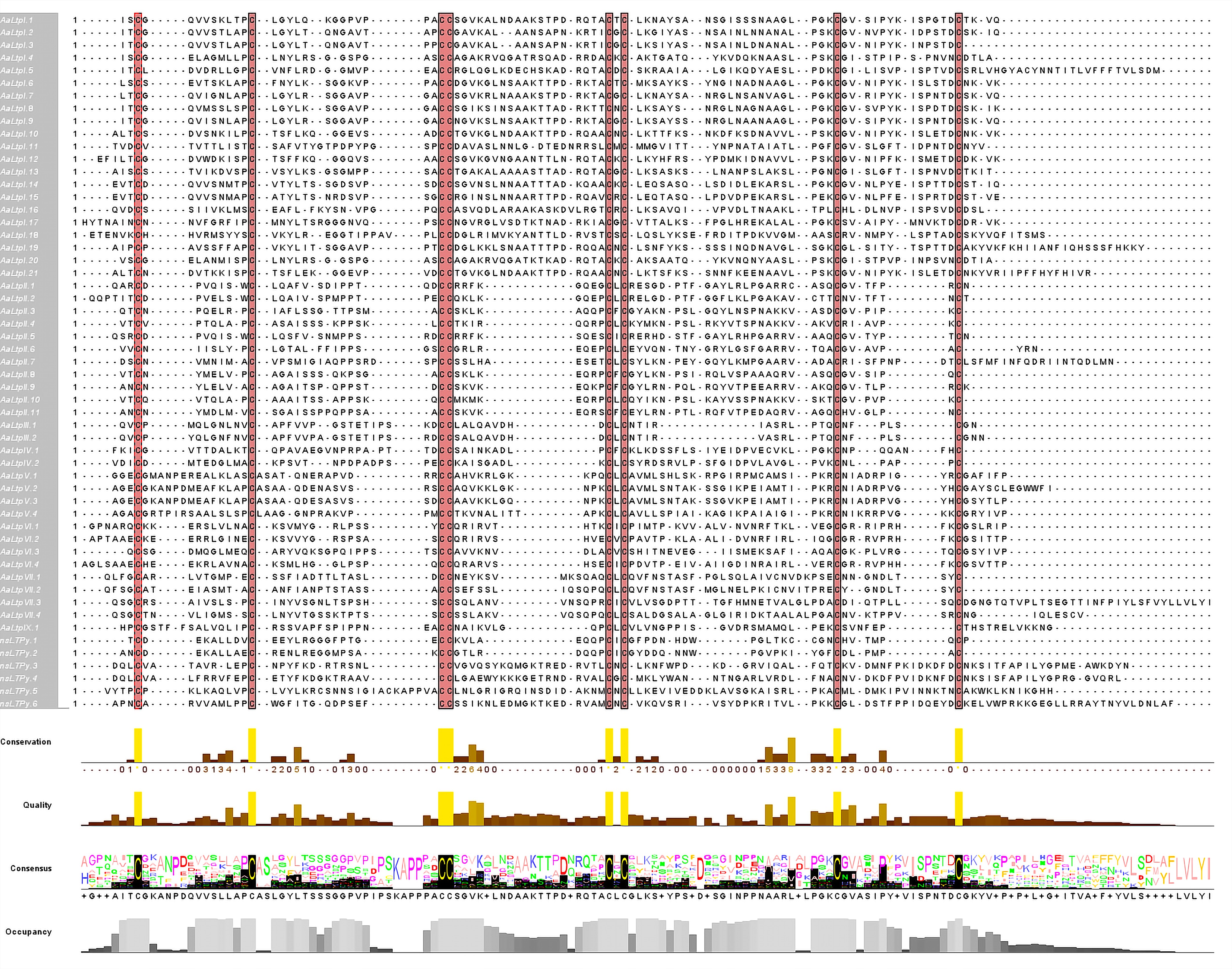
Figure 1.
Alignment of multiple sequences of AaLTP proteins. The conserved cysteine residues are marked against red backgrounds.
Table 2. Diversity of the eight cysteine motif.
nsLTP type ECM and number of flanking amino acid residues 1 2 3, 4 5 6 7 8 Type I X C X9 C X13–16 CC X17, 19, 20 C X C X21−23 C X12, 13 C X Type II X C X7 C X13, 14, 16 CC X8, 9 C X C X19, 20, 23 C X2, 3, 6, 9 C X Type III X C X9 C X16, 17 CC X9 C X C X12 C X5 C X Type IV X C X9 C X15, 17 CC X9 C X C X17, 24 C X6 C X Type V X C X14 C X14 CC X11, 12 C X C X16, 24 C X7, 10 C X Type VI X C X10 C X12, 16 CC X9 C X C X22, 23 C X9 C X Type VII X C X9 C X14 CC X12 C X C X18, 25, 26 C X7, 9 C X Type IX X C X13 C X15 CC X9 C X C X22 C X6 C X Type XI X C X9 C X18−20 CC X13 C X C X24, 25 C X9 C X Phylogenetic analysis of the nsLTP family
-
To analyze the phylogenetic relationships within the AaLTP family, all 55 AaLTP protein sequences were used to construct an unrooted phylogenetic tree using the Maximum likelihood method. This phylogenetic tree, visualized alongside motif analysis, domain analysis, and gene structure analysis (Fig. 2), revealed the evolutionary relationships among the nsLTPs in A. annua. All of the nsLTPs clustered according to their types, forming distinct branches on the evolutionary tree. Motif analysis indicated that nsLTPs of the same type typically contain similar motifs in type and number. Gene structure analysis results were visible for all genes except AaLTPI.1 (mRNA: Super-Scaffold_100123g01462191), AaLTPI.18 (mRNA: unctg_3864g01516351), and AaLTPIII.1 (mRNA: unctg_5142g01560471), which lacked gene annotation files (see domain analysis results for these genes in Supplementary Fig. S2).
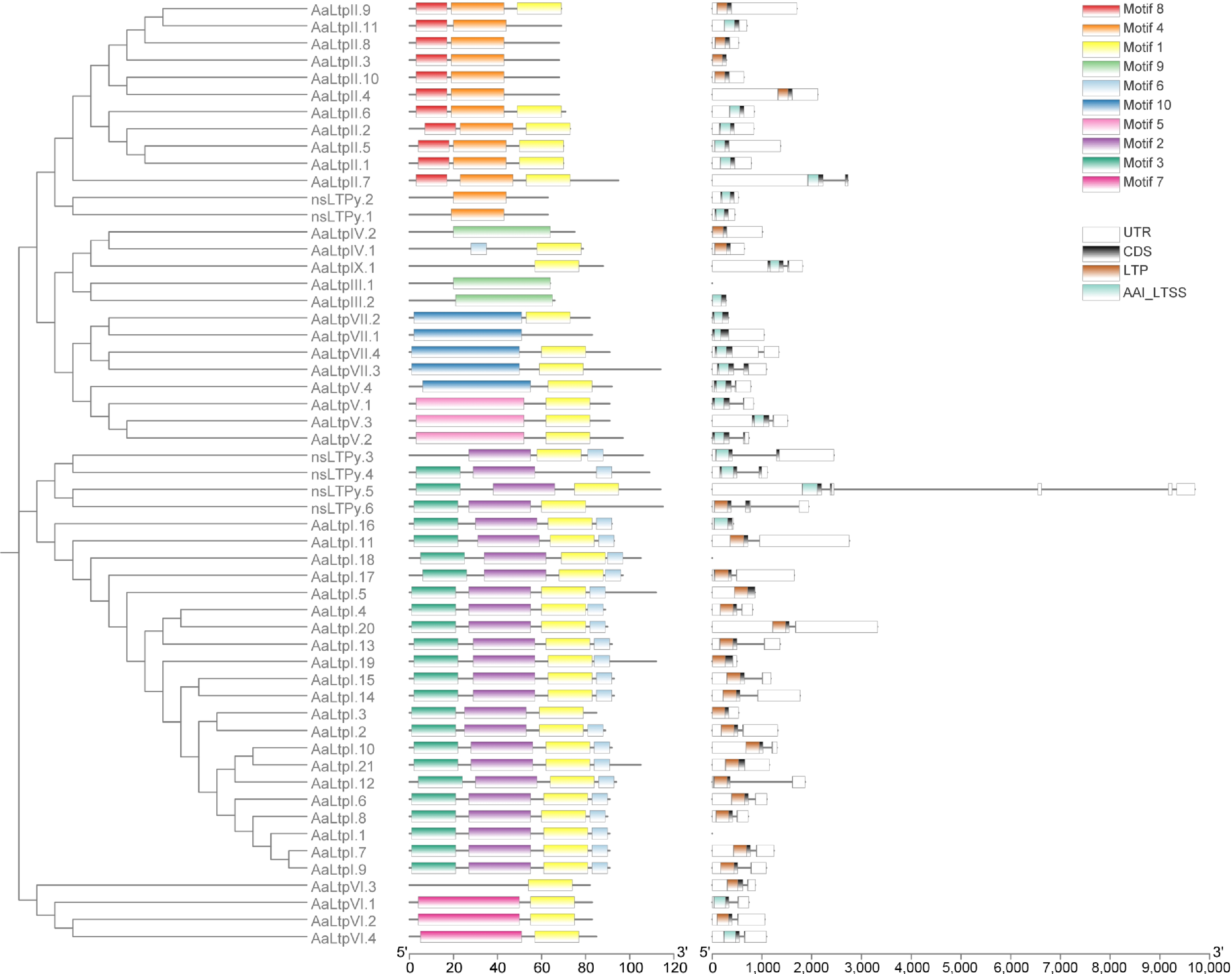
Figure 2.
Phylogenetic tree of AaLTPs together with motif analysis and gene structure analysis. The full length of mature protein sequences of nsLTPs from A. annua was used to construct the phylogenetic tree using a Maximum likelihood method. Motif analysis was performed using MEME, while domain analysis was performed using the batch Web CD-Search tool. All of the above results were visualized together with the gene annotation file.
To further investigate the evolutionary relationship of the nsLTP families between A. thaliana and A. annua, a phylogenetic tree was constructed using the Maximum likelihood method with their 46 and 55 protein sequences, respectively (Fig. 3). The results showed that all nsLTPs could be classified into six clusters (A to F). Type I LTPs were all grouped within cluster A. Type II LTPs were grouped in cluster C, with the exception of one gene (AaLTP II.7). Type III and Type IX LTPs were grouped in cluster E, while cluster D consisted entirely of Type V and Type VII LTPs. Interestingly, AaLTP VI and AtLTP VI were grouped in cluster F and cluster B, respectively, with the exception of AaLTP VI.3 in cluster B. Meanwhile, AtLTP IV.3-5 and AtLTP VI were clustered together in cluster B, whereas an orthologous relationship was observed between AtLTP IV.1/IV.2 and AaLTP IV.1/IV.2, which clustered together in cluster E. Three A. thaliana genes (At3g12545-AtLTPII.8, At3g29105-AtLTPII.10, and At3g52310-AtLTPIX.2) were excluded from the analysis because their predicted mature proteins lacked the conserved ECM structure, which could have potentially interfered with the multiple sequence alignment.
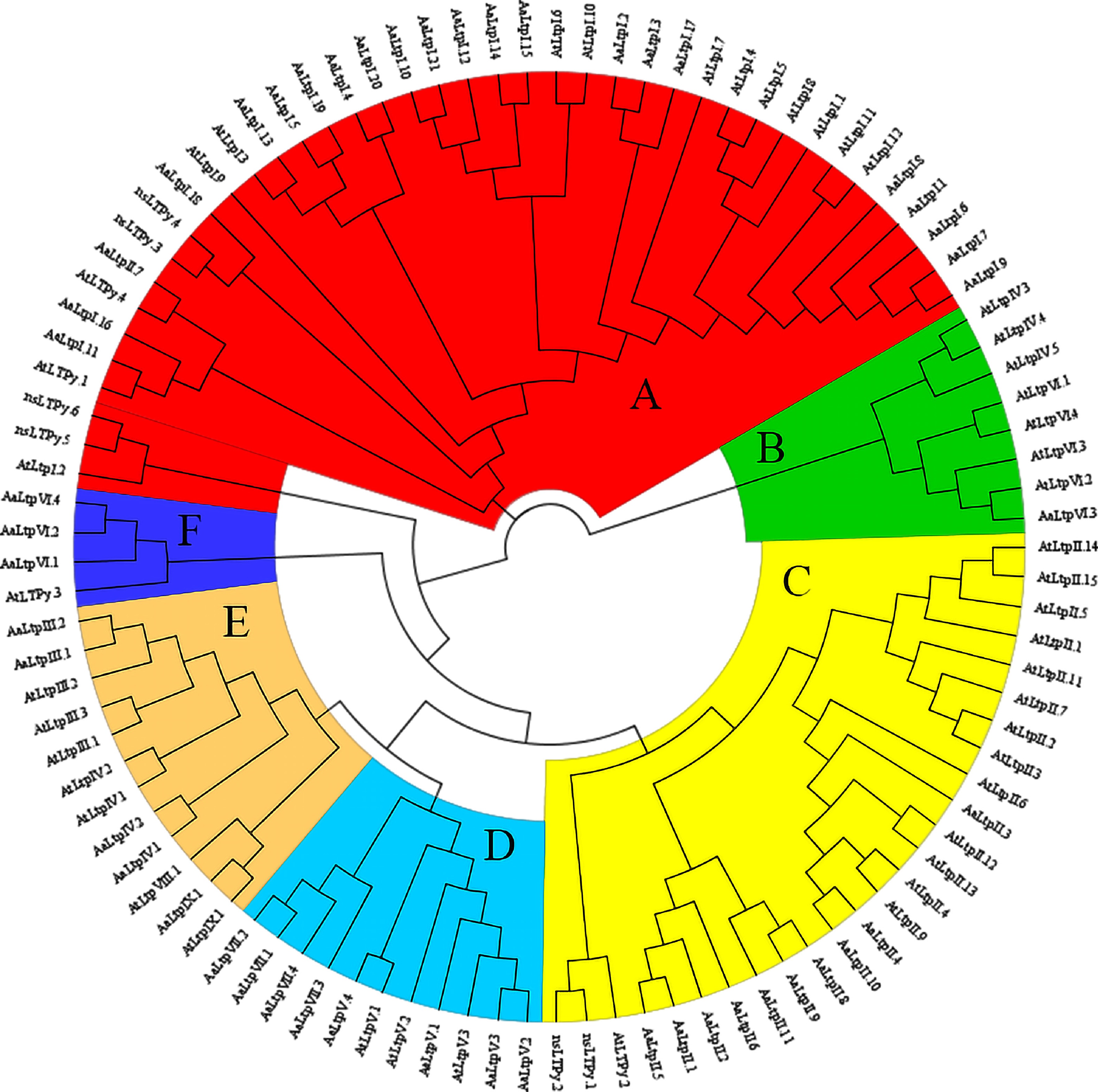
Figure 3.
Phylogenetic tree of nsLTPs in A. thaliana and A. annua. The full length of mature protein sequences of nsLTPs from A. thaliana and A. annua were used to construct the phylogenetic tree using a Maximum likelihood method. Different colors represent different clusters.
Expression analysis of nsLTP genes
-
Analysis of RNA-seq data from six different kinds of organs (roots, stems, leaves, seeds, buds, and trichomes) was used to determine the expression patterns of AaLTP genes[42]. Genes with an RPKM value ≥ 1 were considered positively expressed. As a result, 39 (71%) AaLTP genes were found to be expressed in at least one organ (Fig. 4; Supplementary Table S6). Some genes exhibited specific expression patterns in different organs. For example, chr2g00297041-AaLTPI.6, chr2g00309551-AaLTPI.14, chr4g00612411-AaLTPI.15, and chr1g00206751-AaLTPV.1 were specifically expressed in trichomes. chr3g00577891-AaLTPII.8 and chr5g00891931-AaLTPII.10 showed specific expression in leaves, while chr1g00152381-AaLTPI.5 and chr3g00499431-AaLTPI.20 exhibited specific expression in the seeds.
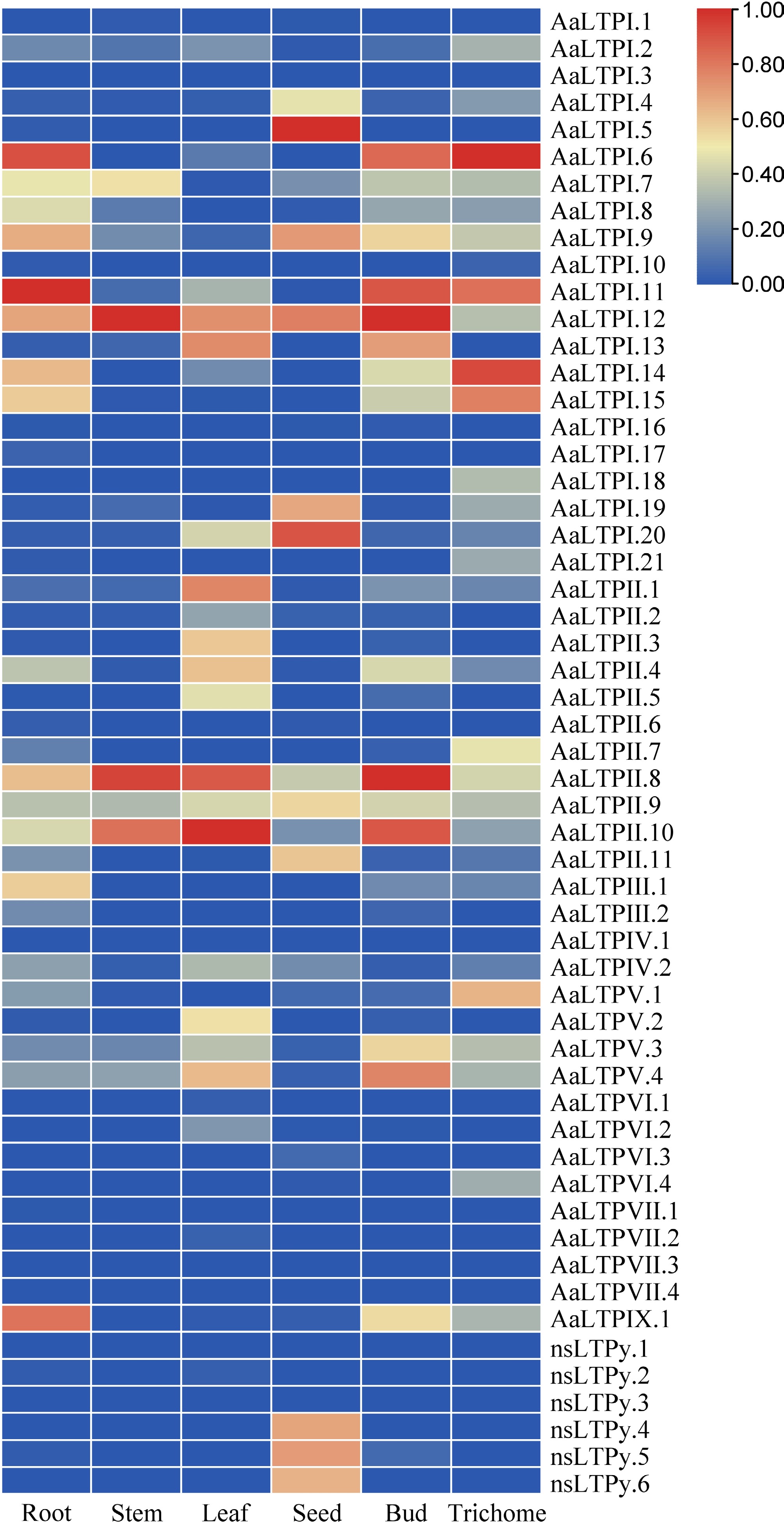
Figure 4.
Expression analysis of AaLTPs. The heatmap shows expression comparison between six organs of AaLTPs. The data shown were the results of column scaling each gene by RNA-seq data using the Zero To One Scale Method. The color bar represents the relative expression level.
Since AaHD8, an HD-Zip IV transcription factor in A. annua, regulates the development of trichome and leaf cuticle, and the LTP gene is mostly related to cuticle, a potential relationship between AaHD8 and AaLTP genes was investigated. Based on RNA-seq data from AaHD8-RNAi lines[43], four AaLTP genes showed significant expression differences (adjusted p-value < 0.05) between 0.038 and 9.19 FC (fold change) (Fig. 5). Specifically, two type I genes, chr2g00297131-AaLTPI.7 and chr2g00297171-AaLTPI.8, exhibited significant upregulation, while two type I genes, chr2g00309551-AaLTPI.14 and chr2g00297041-AaLTPI.6, showed significant downregulation. Expression of chr2g00309551-AaLTPI.14 and chr2g00297041-AaLTPI.6 decreased significantly by 114-fold and 86-fold, respectively, compared to the wild-type (WT), indicating direct regulation of these AaLTP genes by AaHD8, which activated their expression (Supplementary Fig. S3). These findings suggest a regulatory role of AaHD8 in the expression of these AaLTP genes.
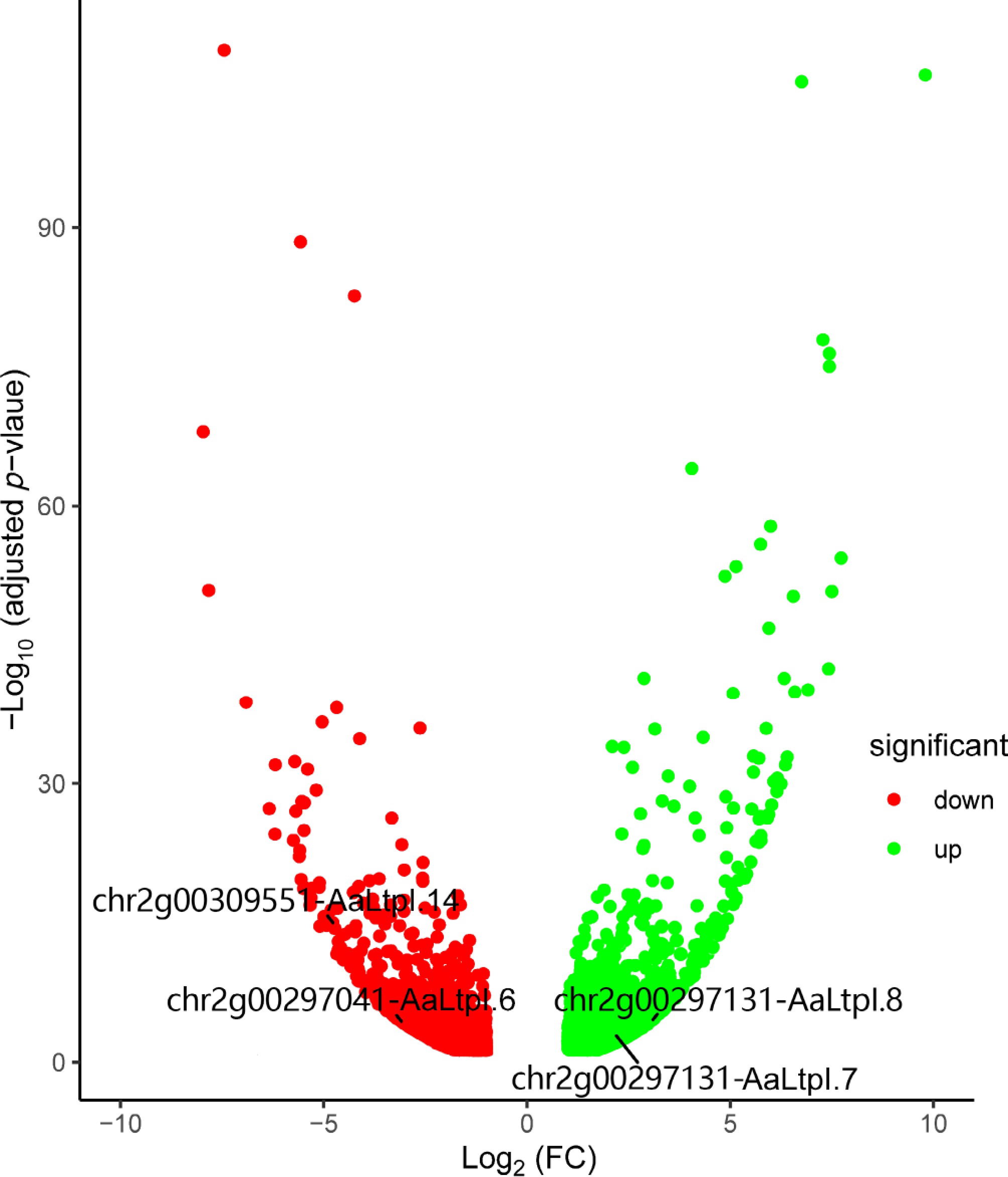
Figure 5.
Volcano plot shows differentially expressed genes in AaHD8-RNAi. The red points indicate down-regulated genes, while the green ones indicate up-regulated genes. FC stands for fold change.
Cloning and expression pattern analysis of AaLTP1 and AaLTP2
-
AaLTPI.14 and AaLTPI.6 exhibited specific and high expression in glandular trichomes (Fig. 4) and were significantly down-regulated in AaHD8-RNAi plants (Fig. 5). Based on their predominant expression and importance, these two genes were cloned and named AaLTP1 and AaLTP2, respectively, following the nomenclature convention for key nsLTPs established in model plants like O. sativa and A. thaliana. To investigate their spatial expression patterns, promoter regions—1,967 bp for AaLTP1 and 1,171 bp for AaLTP2—were cloned into the 1391Z-proAaLTP1-GUS and 1391Z-proAaLTP2-GUS vectors, respectively, and these constructs were transformed into A. annua plants. GUS staining of transgenic plants revealed that both genes are specifically expressed in young leaves, particularly in GSTs, as opposed to T-shaped non-GSTs (TSTs) (Fig. 6). This expression pattern aligns with the RNA-seq data previously determined (Fig. 4), confirming the specificity of AaLTP1 and AaLTP2 expression in glandular trichomes.
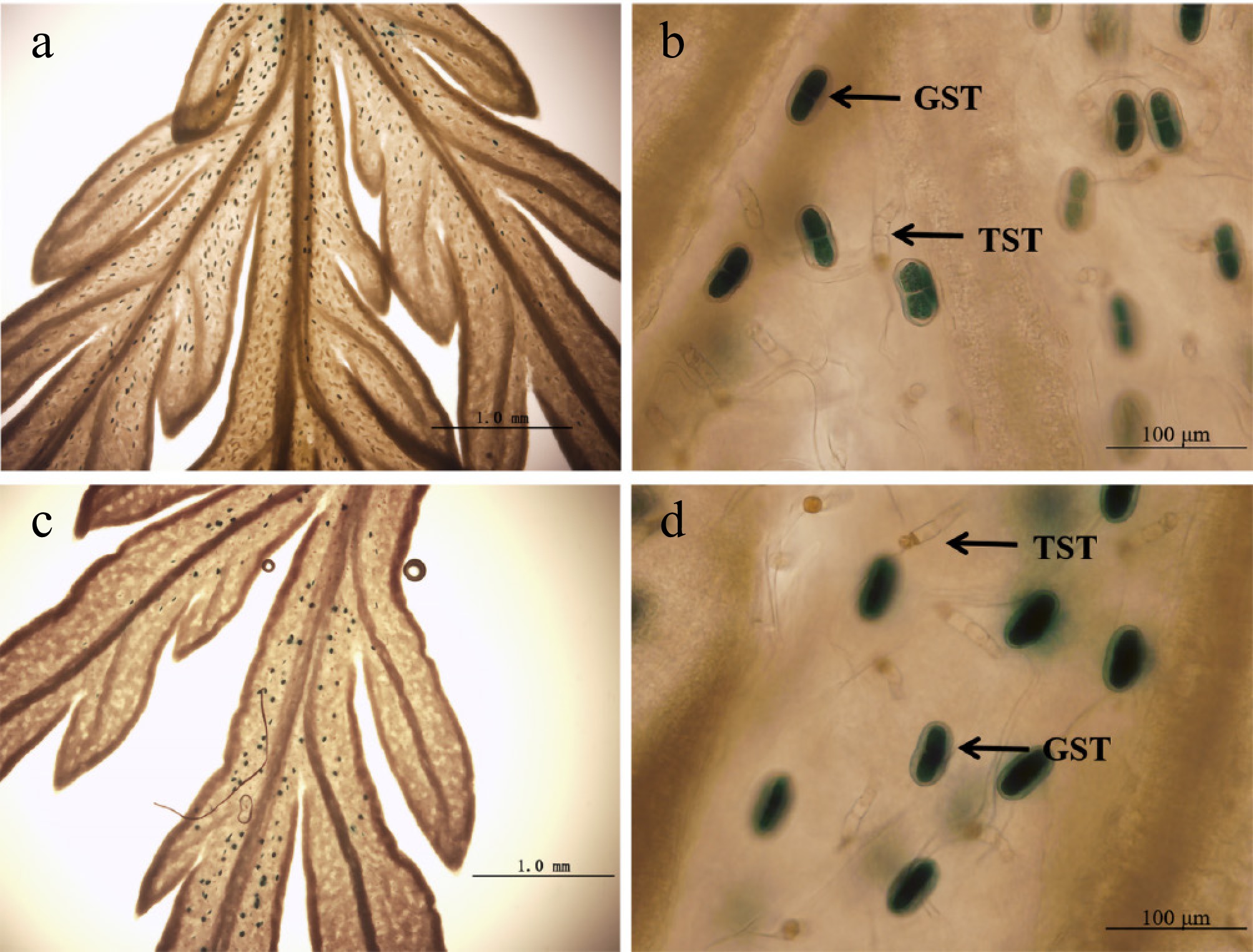
Figure 6.
GUS expression (blue) in A. annua plants transformed with the 1391Z-proLTP1-GUS and 1391Z-proLTP2-GUS vector. (a), (b) Leaves of 1391Z-proLTP1-GUS plants. (c), (d) Leaves of 1391Z-proLTP2-GUS plants.
Enhanced production of artemisinin in transgenic AaLTP1-OE plants
-
Given that AaLTP1 exhibited the most significant downregulation in AaHD8-RNAi plants, to further explore the function of the AaLTP1 gene, transgenic plants overexpressing AaLTP1 (AaLTP1-OE) were generated. Positive transgenic plants were confirmed by PCR, and the expression levels of AaLTP transcripts were validated by qRT-PCR. Subsequently, three independent transgenic lines showing a two to three-fold increase in AaLTP1 mRNA level were selected for further analysis (Fig. 7).
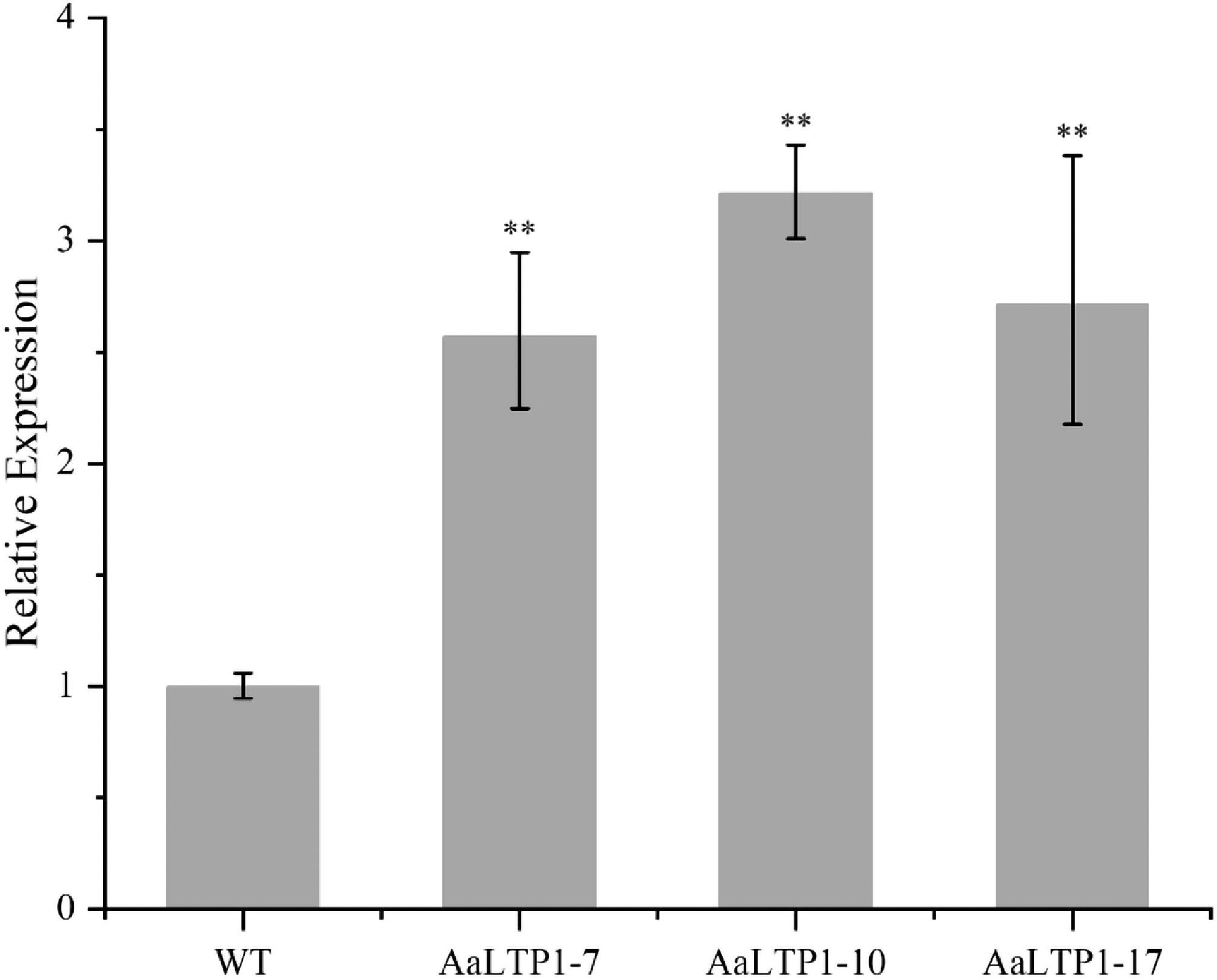
Figure 7.
Expression levels of AaLTP1 in AaLTP1 over-expression plants measured by qRT-PCR. The average expression level of genes in WT was set as 1. Actin was used as internal control; the error bars show means ± SD from three technical repeats; Student's t-test ** p < 0.01, * p < 0.05.
The artemisinin content in the AaLTP1-OE plants was analyzed using High Performance Liquid Chromatography (HPLC). The results demonstrated that the artemisinin content in AaLTP1-OE plants increased by 1.6 times compared to the WT (Fig. 8), indicating that over-expression of the AaLTP1 gene promotes artemisinin accumulation in A. annua.
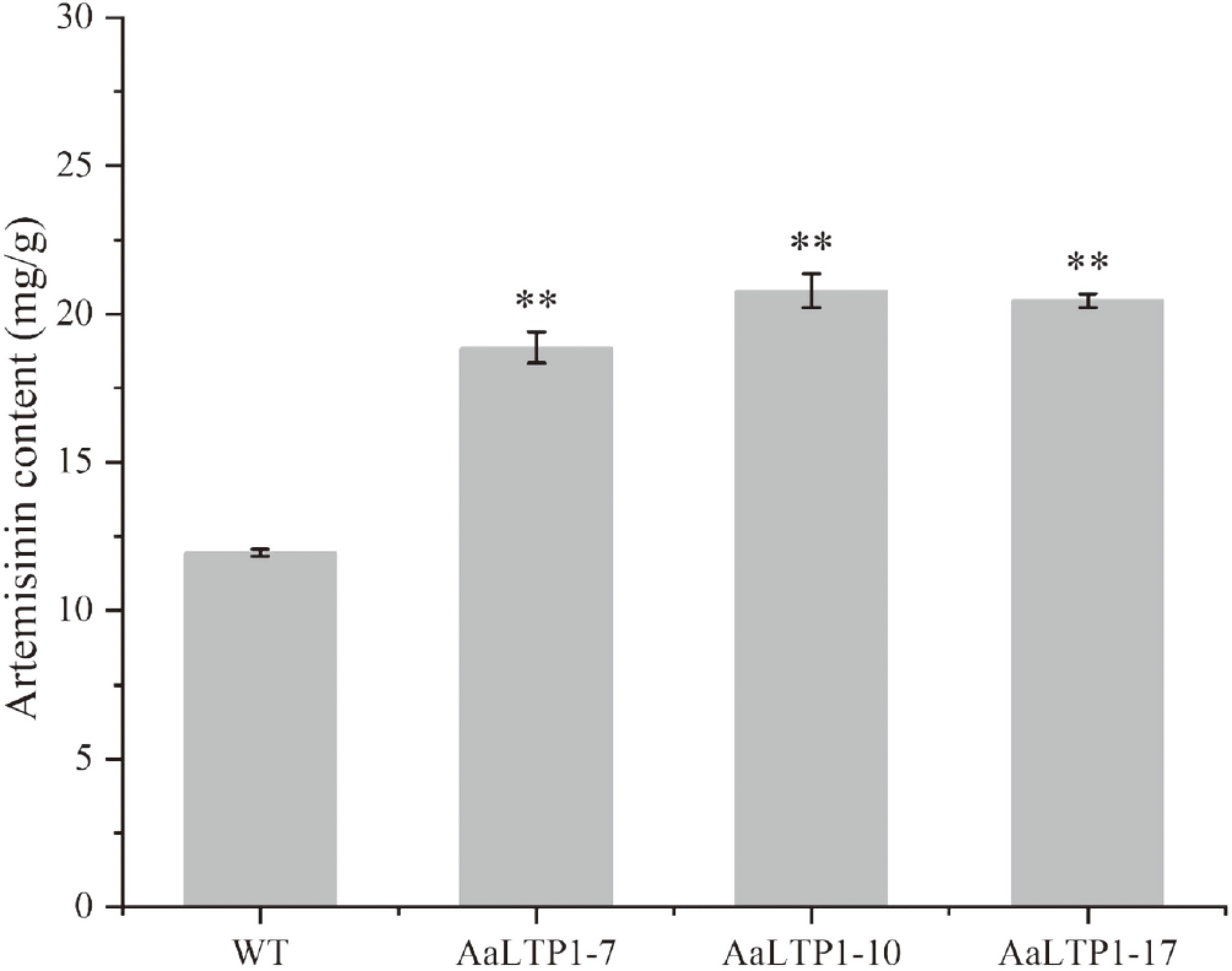
Figure 8.
Artemisinin contents in AaLTP1 over-expression lines were measured by HPLC. Error bars indicate ± SD of three biological replicates. ** p < 0.01, Student's t-test.
AaHD8 positively regulates AaLTP gene expression through direct binding to the promoter
-
AaHD8, a member of the HD-Zip IV gene family, was hypothesized to bind to the L1-box cis-element (5'-TAAATG(C/T) A-3') due to its known affinity for this sequence[48]. Analysis of the AaLTP gene promoters using PlantPAN 3.0 identified L1-box-like sequences, suggesting that AaHD8 may interact with these regions. This hypothesis was tested through yeast one-hybrid assays, which confirmed that AaHD8 binds to the L1-box-like sequences in the promoters of AaLTP1 and AaLTP2 (Fig. 9). Electrophoretic mobility shift assays (EMSA) using recombinant His-tagged AaHD8 and His-tagged trigger factor (TF) purified from Escherichia coli further validated this interaction. A distinct shifted band was observed with His-AaHD8 and a biotin-labeled L1 probe, with no binding seen with His-TF or a mutated probe (Fig. 9). These results demonstrate that AaHD8 regulates the expression of AaLTP1 and AaLTP2 by directly binding to the L1-box-like motifs in their promoters.
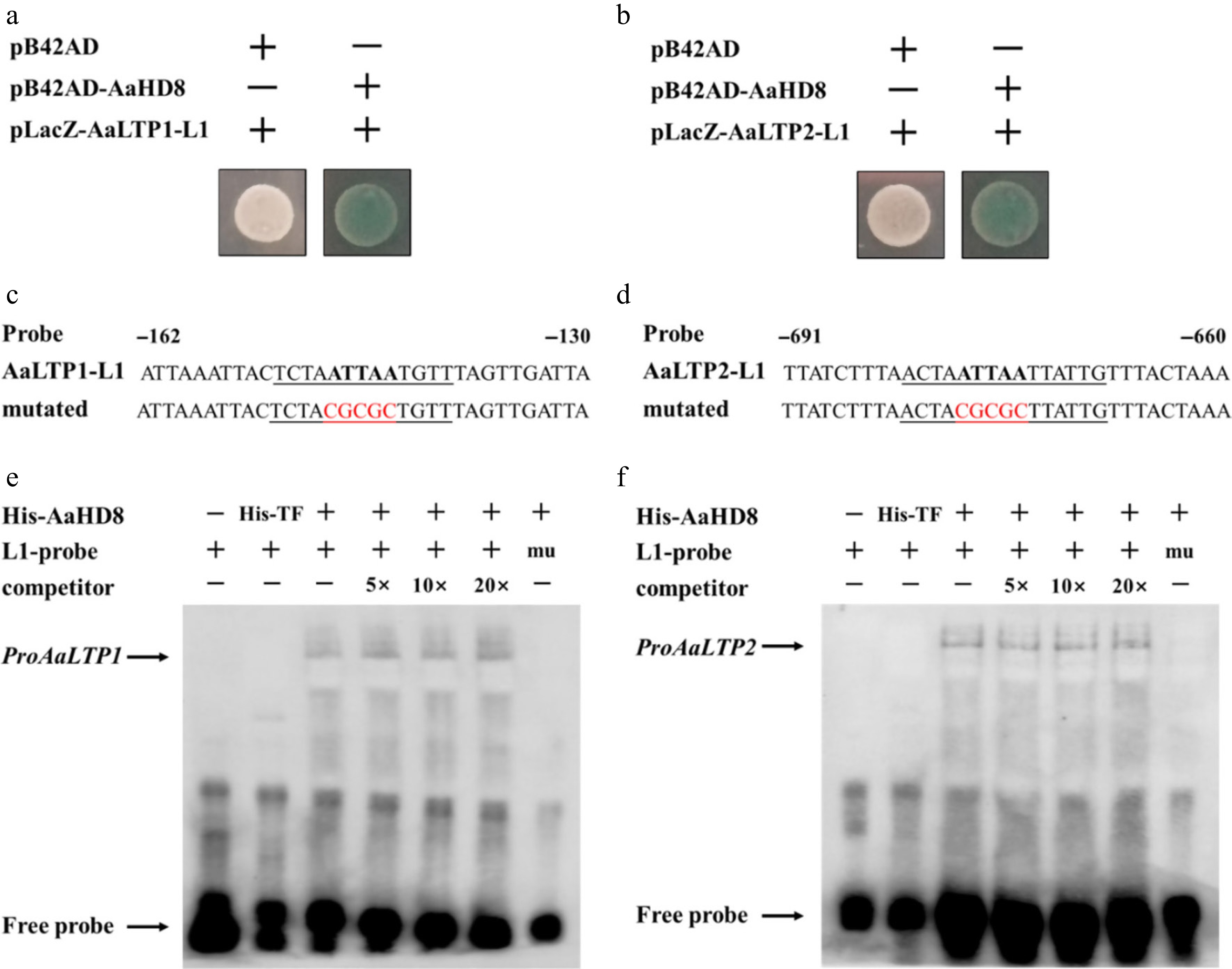
Figure 9.
AaHD8 directly binds to the promoter of AaLTP1 and AaLTP2. (a), (b) Yeast one-hybrid assay of protein–DNA interaction. pB42AD is an empty vector used as a negative control. (c), (d) Probe and mutant probe. (e), (f) AaHD8 binds to the L1-box-like motif from AaLTP; 5×, 10×, and 20× indicate the dilution multiple of cold competitors relative to that of the labeled probe; mu: labeled mutated L1-probes were tested as negative controls; His-TF protein was used as a negative control.
To validate the regulatory role of AaHD8 on AaLTP expression in vivo, qRT-PCR analysis was performed using WT A. annua and transgenic plants overexpressing AaHD8 (Fig. 10). Compared to WT, AaHD8 overexpression increased the expression of AaLTP1 and AaLTP2 genes by 2.2 to 4.9 times and 2.3 to 4.5 times, respectively, in five independent AaHD8 overexpressing plants. These results indicated that AaHD8 overexpression can directly activate the expression of AaLTP1 and AaLTP2 in A. annua plants.
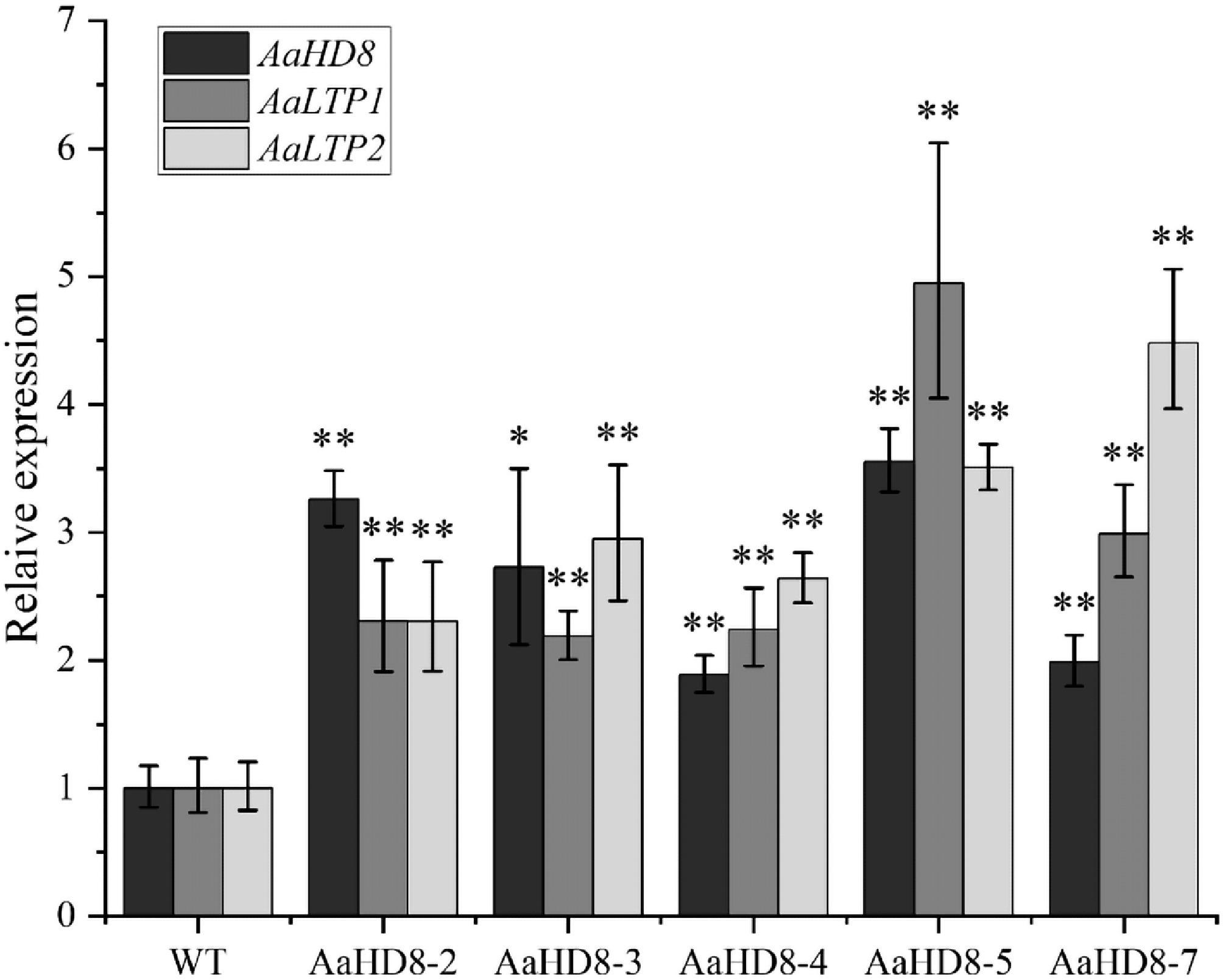
Figure 10.
Relative expression of AaHD8 and AaLTP in AaHD8 over-expression plants. The average expression level of genes in WT was set as 1. Actin was used as internal control; the error bars show means ± SD from three technical repeats; Student's t-test ** p < 0.01, * p < 0.05.
-
This study presents the first comprehensive genome-wide analysis of nsLTPs in A. annua and their involvement in artemisinin biosynthesis. The identification of AaLTP1 and AaLTP2 as glandular trichome-specific nsLTPs highlights their potential role in facilitating the intracellular trafficking of metabolites in these specialized tissues. Indeed, the 1.5-fold increase in artemisinin content observed in AaLTP1-overexpressing lines provides strong evidence that these proteins directly contribute to artemisinin production. This finding contrasts with earlier reports where AaLTP1 and AaLTP2 showed minimal effects on dihydroartemisinic acid (DHAA) accumulation[49], suggesting functional divergence or context-dependent regulation within the nsLTP family. While prior studies implicated AaLTP3 and AaLTP4 in sesquiterpene secretion[31], results establish AaLTP1 as a key regulator of artemisinin biosynthesis, underscoring the need to dissect isoform-specific roles in lipid trafficking.
Comprehensive analysis of the nsLTP gene family in A. annua reveals a significant number of 55 nsLTP genes, encompassing diverse types, including 21 type I, 11 type II, two type III, two type IV, four type V, four type VI, four type VII, one type IX, and six nsLTPy (Table 1). This study provides a detailed characterization of these genes, in contrast to other plant species such as Arabidopsis, rice, rape, and maize, which have been reported to contain 49, 52, 63, and 63 nsLTPs, respectively. Notably, A. annua lacks nsLTP types VIII, X, and XI (Table 2), a loss that is consistent with the absence of type XI nsLTPs in monocotyledonous plants and suggests an evolutionary divergence between monocots and dicots. Despite the identification of proteins resembling type XI nsLTPs in A. annua, domain analysis confirmed that these do not possess the requisite LTP or AAI_LTSS structures, thus excluding them from classification as nsLTPs.
The glandular trichome-specific expression of AaLTP1 and AaLTP2 strongly implies their involvement in the transport of hydrophobic precursors essential for artemisinin biosynthesis. A key question is how extracellularly localized nsLTPs might access cytosolic substrates like FPP or potentially organellar DHAA. It is hypothesized that this may involve an intermediate step: these precursors are first actively transported across the plasma membrane or organellar membranes by specific transporters, thereby becoming accessible to the apoplastic nsLTPs. AaLTP1 and AaLTP2 could then function as extracellular shuttles, facilitating the subsequent movement of these precursors through the glandular trichome secretory space or towards the site of final artemisinin assembly and storage. Structural studies across plant species provide mechanistic clues: nsLTPs universally feature a hydrophobic cavity stabilized by disulfide bonds[50,51], enabling lipid binding. For instance, in Solanum melongena, nsLTP binds lipids through a dynamic process where surface-bound intermediates transiently interact before internalization into the cavity, mediated by conformational changes in 'gating' residues[52]. Similarly, the crystal structure of wheat nsLTP revealed dual lipid binding—one in the main cavity and another in an adjacent pocket[50]—suggesting a capacity for simultaneous transport of distinct lipophilic molecules. In A. annua, AaLTP1 and AaLTP2 may utilize this conserved mechanism to sequester artemisinin precursors such as dihydroartemisinic acid (DHAA) or farnesyl pyrophosphate (FPP) within their hydrophobic cavities, shielding these intermediates from degradation while facilitating their movement across cellular compartments and surrounding tissues, including vascular systems, which may be critical for the efficient delivery of precursors to the artemisinin biosynthetic pathway[53−55]. The observed 1.5-fold increase in artemisinin content in AaLTP1-OE lines supports this hypothesis. Structural analysis of Kalanchoe fedtschenkoi nsLTP[56] demonstrated direct binding of lauric acid within the hydrophobic cavity, a feature likely conserved in AaLTPs. Additionally, the hydrogen bond-mediated closure of the binding pocket observed in eggplant nsLTP[51] suggests that environmental cues (e.g., pH shifts in glandular trichomes) or interactions with partner proteins could regulate lipid release from AaLTPs, thereby synchronizing precursor availability with biosynthetic demands. To rigorously test these hypotheses and clarify the trafficking route, future studies must include molecular docking simulations to predict their affinity for artemisinin precursors like DHAA and FPP. Furthermore, in vitro lipid-binding assays using purified proteins and isotopic tracing of precursor trafficking in transgenic lines will be essential.
The localization of AaLTP1 and AaLTP2 in glandular trichomes aligns with the conserved role of nsLTPs in specialized metabolite transport across plant species, yet highlights species-specific adaptations for distinct secondary metabolites. In Mentha canadensis, nsLTPs such as McLTPII.9 enhance peltate glandular trichome density and monoterpene biosynthesis[57], while in Tanacetum parthenium, trichome-specific TpLTP1 and TpLTP2 mediate extracellular accumulation of sesquiterpene lactones like costunolide and parthenolide[58]. Similarly, AaLTP1 and AaLTP2 likely evolved to facilitate artemisinin precursor transport in A. annua glandular trichomes. This role is not unique to A. annua: the involvement of nsLTPs in trafficking precursors or cofactors essential for secondary metabolite biosynthesis in glandular trichomes is well-documented in plants such as Salvia fruticosa[59], hop[60], Cistus creticus[61], A. annua[62], alfalfa[63], basil[64], and peppermint[65], where lipid transfer proteins mediate transport across membranes in specialized cells. However, substrate specificity varies significantly among species. For instance, TpLTP3 in feverfew exhibits strict selectivity for parthenolide export[58], whereas AaLTP1 may preferentially bind artemisinin precursors such as DHAA, analogous to the lauric acid-binding activity observed in Kalanchoe fedtschenkoi nsLTP[56]. This functional divergence underscores how nsLTPs are tailored to the unique metabolic demands of their host plants. Notably, nsLTPs in non-glandular tissues also play conserved roles in lipid barrier formation. For example, castor bean nsLTP regulates fatty acid β-oxidation in glyoxysomes[66], while Arabidopsis LTPs contribute to cuticular wax synthesis[67]. In contrast, AaLTP1 and AaLTP2 appear specialized for glandular trichome biology, mirroring the trichome-specific roles of McLTPII.9[57] and TpLTP1/2[58]. This tissue-specific specialization suggests that while nsLTPs broadly mediate lipid trafficking, their recruitment to glandular trichomes represents an evolutionary adaptation to enhance the production and secretion of high-value metabolites like artemisinin.
The identification of AaHD8, an HD-ZIP IV transcription factor involved in cuticle biosynthesis and trichome initiation[43], as a direct regulator of AaLTP1 and AaLTP2 expands understanding of nsLTP regulation in A. annua. HD-ZIP transcription factors are known integrators of developmental and environmental signals—such as light, temperature, and biotic stress—that modulate secondary metabolism[68,69]. Findings suggest that AaHD8 may orchestrate artemisinin biosynthesis by linking lipid transport (AaLTP1/2 expression) with trichome development and stress adaptation; for example, upregulation of AaLTP1 and AaLTP2 via AaHD8 might be part of a broader adaptive mechanism to optimize artemisinin production under fluctuating environmental conditions. This regulatory nexus aligns with observations in other medicinal plants, where HD-ZIPs coordinate metabolite production and abiotic stress responses[70]. Future studies should investigate whether AaHD8 interacts with MYB or WRKY transcription factors to fine-tune AaLTP expression and explore its broader role in artemisinin pathway regulation via ChIP-seq or promoter mutagenesis.
The identification and functional characterization of AaLTP1 and AaLTP2 provide a potential route for biotechnological enhancement of artemisinin production. Overexpression of these genes in engineered A. annua could be an effective strategy for increasing the yield of artemisinin. Furthermore, the regulation of AaLTP1 and AaLTP2 by AaHD8 suggests that manipulating this transcription factor could enhance artemisinin production under controlled environmental conditions. Understanding the environmental and genetic factors that influence AaHD8 activity could help in designing plants with enhanced resistance to abiotic stresses while simultaneously boosting the production of this important pharmaceutical compound.
While this study has provided valuable insights into the role of nsLTPs in A. annua, several questions remain unanswered. For instance, the precise mechanism by which AaLTP1 and AaLTP2 influence lipid and artemisinin metabolism at the molecular level is still unclear. Notably, RNA-seq data revealed additional nsLTP candidates potentially involved in glandular trichome biology: AaLTPI.11 is highly expressed in both glandular trichomes and roots, suggesting dual roles in metabolite transport across tissues; AaLTPI.15 and AaLTPV.1, though trichome-specific, exhibit lower expression levels compared to AaLTP1/2, implying subsidiary or stage-specific functions. Intriguingly, in AaHD8-RNAi plants, AaLTPI.7 and AaLTPI.8 were upregulated—contrary to the downregulation of AaLTP1/2—hinting at compensatory mechanisms or divergent regulatory networks among nsLTP isoforms. It is worth noting that the observed phenotypic changes are unlikely to be attributed to alterations in trichome development, as no significant differences in trichome density or morphology were detected, which reinforces the proposed direct role of AaLTP1/2 in lipid transport. Future research should therefore focus on identifying the specific lipid ligands bound by these proteins, their interactions with artemisinin biosynthetic enzymes, and the regulatory logic underlying their opposing expression patterns. Additionally, exploring the roles of other trichome-enriched nsLTPs in stress responses could provide a holistic understanding of lipid metabolism in A. annua.
In summary, these findings represent an important step in elucidating the role of lipid transfer proteins in A. annua and their contribution to artemisinin biosynthesis. The functional validation of AaLTP1 and AaLTP2, along with the identification of AaHD8 as a key regulator, opens new avenues for both basic and applied research. Future studies should focus on resolving the molecular mechanisms by which AaLTP1 facilitates artemisinin precursor transport, including in vitro ligand-binding assays and spatial tracking of lipid trafficking within glandular trichomes. The broader regulatory network involving AaHD8 and its potential crosstalk with other trichome-specific transcription factors (e.g., MYB or WRKY families) warrants systematic exploration through chromatin immunoprecipitation and multi-omics approaches. By bridging mechanistic insights with metabolic engineering strategies, this work provides a new strategy for increasing the yield of artemisinin through genetic engineering methods while advancing understanding of nsLTP functionality in plant metabolism.
-
In this study, the first genome-wide identification and functional characterization of the nsLTP gene family in A. annua, a medicinal plant renowned for its production of the antimalarial compound artemisinin, was conducted. A total of 55 nsLTP genes were identified and classified into eight distinct types, revealing both conserved and species-specific features in their gene structure, motif composition, and evolutionary relationships. Notably, expression profiling and promoter-GUS assays demonstrated that AaLTP1 and AaLTP2 are specifically expressed in glandular secretory trichomes—the primary sites of artemisinin biosynthesis.
Functional analyses revealed that overexpression of AaLTP1 significantly enhances artemisinin accumulation, underscoring its direct role in the artemisinin biosynthetic pathway. Furthermore, the HD-Zip IV transcription factor AaHD8 was established as a positive regulator of AaLTP1 and AaLTP2 by directly binding to L1-box-like motifs in their promoters. This regulatory link integrates trichome development with lipid transport mechanisms, highlighting a coordinated network that supports artemisinin production.
These findings not only expand the understanding of nsLTP functions in specialized metabolism but also provide a genetic toolkit for metabolic engineering aimed at increasing artemisinin yield. Future studies should focus on elucidating the specific lipid substrates of AaLTP1/2, their interaction with artemisinin biosynthetic enzymes, and the broader regulatory circuitry involving AaHD8 and other trichome-specific transcription factors. This work lays a foundation for leveraging lipid transfer proteins in biotechnological strategies to enhance the production of high-value plant metabolites.
This work was supported by the National Natural Science Foundation of China (Grant Nos. 32070329, 31770327), the Natural Science Foundation of Shanghai (Grant No. 25ZR1401180), the National Key R&D Program of China (Grant No. 2018YFA0900600), and the Bill & Melinda Gates Foundation (Grant No. OPP1199872).
-
The authors confirm their contributions to the paper as follows: study conception and design: Wen H, Wang W, Yan S, Li L; data collection: Wang W, Liu H, Wu E; analysis and interpretation of results: Wen H, Wang W, He W, Liu H, Su L, Hu X, Yan X; investigation: Wen H, Wang W, Liu H, Su L, Hu X, Yan X, Peng B, Zhang Y, Liu P; methodology: Wen H, Wang W; software: Wen H; visualization: Wen H; draft manuscript preparation: Wen H, Wang W; writing – review & editing: He W, Yan S, Li L; supervision: Tang K, Li L; project administration: Fu X, Ren L, Tang K, Li L; resources: Tang K; funding acquisition: Tang K, Li L. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Hangyu Wen, Wei Wang
- Supplementary Table S1 The coding sequences of AaLTP genes in A. annua.
- Supplementary Table S2 AaLTPs with significantly changed expression levels (FPKM) in AaHD8-RANi line.
- Supplementary Table S3 Primer used in this study.
- Supplementary Table S4 Detailed classification criteria of AaLTPs.
- Supplementary Table S5 Subcellular location prediction of AaLTPs.
- Supplementary Table S6 RPKM of AaLTP genes in different organs.
- Supplementary Fig. S1 Three-dimensional structures of the mature AaLTPs predicted by Phyre2.
- Supplementary Fig. S2 Domain analysis results using batch Web CD-Search tool. Genes whose structure is not shown in Figure 2 can see domain information in this figure.
- Supplementary Fig. S3 TPM (Transcripts Per kilobase Million) value of AaLTP in AaHD8-RNAi lines.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wen H, Wang W, He W, Liu H, Su L, et al. 2026. Comprehensive genome-wide identification and functional characterization of non-specific lipid transfer proteins in Artemisia annua L.. Medicinal Plant Biology 5: e002 doi: 10.48130/mpb-0025-0040 

Comprehensive genome-wide identification and functional characterization of non-specific lipid transfer proteins in Artemisia annua L.
- Received: 10 August 2025
- Revised: 15 October 2025
- Accepted: 05 November 2025
- Published online: 19 January 2026
Abstract: Plant non-specific lipid transfer proteins (nsLTPs) are small secretory proteins that bind and transport hydrophobic molecules. nsLTPs are crucial players in various plant physiological processes, including lipid metabolism and defense. In Artemisia annua, glandular trichomes serve as the primary sites for artemisinin biosynthesis and storage. Although several nsLTPs have been reported in Artemisia annua, a comprehensive genome-wide analysis was previously lacking. In this study, 55 nsLTP genes were identified in the A. annua genome and classified into eight types (I, II, III, IV, V, VI, VII, and IX) through phylogenetic analysis. Notably, expression profiling revealed that certain AaLTP genes, especially AaLTP1 and AaLTP2, exhibit glandular trichome-specific expression, as confirmed by promoter::GUS assays. Overexpression of AaLTP1 significantly increased artemisinin content by 1.5-fold (p < 0.05), suggesting its direct role in facilitating artemisinin accumulation. Furthermore, an HD-ZIP transcription factor, AaHD8, was confirmed as a positive regulator of AaLTP1 and AaLTP2 through direct promoter binding. Overall, this study provides the first comprehensive characterization of the nsLTP gene family in A. annua and establishes a functional link between nsLTPs, glandular trichome biology, and artemisinin biosynthesis, offering new insights for metabolic engineering to enhance artemisinin production.
-
Key words:
- Glandular trichomes /
- Artemisinin /
- Artemisinin biosynthesis