









-
LBD genes are transcription factors (TFs) that are peculiar to green plants and may have evolved from charophyte algae. LBD TFs contain a highly conserved lateral organ boundaries (LOB) domain, which is about 100 amino acids. The LOB domain consists of a conserved CX2CX6CX3C zinc finger-like motif at the N-terminal region, a Gly-Ala-Ser (GAS) block in the middle, and leucine zipper-like coiled-coil motif (LX6LX3LX6L) motif[1−3]. Based on the examination of the LOB domain and phylogenetics, the LBD proteins were categorized into two primary classes, namely Class I and Class II.[3]. The Class I proteins encode a complete LOB domain, while Class II lacks a LX6LX3LX6L motif[1,4].
With the release of plant genomic information, the LBD gene family has been investigated gradually in several plants. Fourty two LBD family members were identified in Arabidopsis (Arabidopsis thaliana)[1], 35 in rice (Oryza sativa)[5], 44 in maize (Zea mays)[6], 47 in tomato (Solanum lycopersicum)[7], 40 in grape (Vitis vinifera)[8], 58 in apple (Malus domestica)[9], 57 in poplar (Populus trichocarpa)[10], and 46 in Eucalyptus grandis[11]. LBD family members have only been investigated in plants, indicating their crucial involvement in controlling growth and developmental processes particular to plants. LBDs were formerly believed to play a role in the development of various plant organs, including roots, shoot meristems, leaves, flowers, and embryos[12]. For instance, Arabidopsis AtLBD16 and AtLBD29 can regulate lateral root formation[13], AtLBD6 controls stem meristem[14], leaf adaxial identity, and sepal and petal development[15,16], and AtLBD30 is involved in embryogenesis and floral development[17,18] Moreover, recent research has demonstrated that LBDs also have a function in the process of anthocyanin biosynthesis, nitrogen metabolism, secondary growth, shoot-borne root initiation, plant defenses, hormone response and plant regeneration. For example, AtLBD37/38/39 in Arabidopsis and MdLBD13 in apple can negatively regulate anthocyanin biosynthesis and nitrogen uptake and assimilation[19,20]. PtaLBD1 in poplar (Populus tremula × P. alba) regulates secondary phloem development[21], EgLBD37 and EgLBD29 in E. grandis are involved in secondary xylem differentiation and phloem fiber production[11], respectively. Additionally, Class IIIB members can specifically regulate shoot-borne root initiation in angiosperms[22]. The expression level of MaLBD5, derived from the banana species Musa acuminata, was stimulated by treatment with MeJA and exposure to cold stress. This gene is implicated in the enhancement of cold tolerance mediated by MeJA[23].
The Rosaceae family consists of over 100 genera and can be divided into four subfamilies: Rosoideae, Prunoideae, Spiraeoideae, and Maloideae. Rosaceae plants, including ornamentals, fruit species, aromatic, and medicinal plants, are economically important plant families. In this study, nine representative plants (black raspberry, strawberry, Chinese rose, peach, apricot, mei, hawthorn, pear, and apple) from three traditional subfamilies (Rosoideae, Prunoideae, Maloideae) of Rosaceae were selected to study. The LBD members were first identified in nine plants. Next, we performed phylogenetic analysis, conserved motifs, sequence alignment, chromosome localization, collinearity analysis, and cis-acting element analysis on these genes. Finally, based on transcriptome data and quantitative real-time (qRT)-PCR analysis, we investigated their expression pattern of PmLBDs in different tissues, cold stress, flower bud dormancy release, and plant architecture. Collectively, these investigations will offer fresh perspectives on the evolutionary correlation of the LBD family in Rosaceae and the expression profile of PmLBDs in P. mume.
-
Genome-wide protein data and annotation data of A. thanliana (TAIR 10), P. trichocarpa (v4.0), and nine other Rosaceae species, including R. occidentali s (GDR,v3.0), F. vesca (v4.0.a1), R. chinensis (v1.0), P. persica (v2.0.a1), P. armeniaca (v1.0), P. mume var. tortuous (v1.0), C. pinnatifida (v1.0), P. communis (v2.0), Malus domestica 'HFTH1' (v1.0) were download from the TAIR database (
www.arabidopsis.org , accessed on 12 August 2023)[24], the Phytozome database (https://phytozome-next.jgi.doe.gov/ , accessed on 2 September 2023)[25] and the Genome Database for Rosaceae (www.rosaceae.org , accessed on 15 August 2023)[26], respectively.Identification of LBDs
-
A HMMER search was used to identify the possible LBDs with the LOB domain (PF03195) from the Pfam database 36.0 (
http://pfam.xfam.org/ , accessed on 7 September 2023)[27]. In addition, 43 LBD protein sequences of Arabidopsis were obtained from the TAIR database (www.arabidopsis.org , accessed on 12 August 2023)[24] and used to perform a BLASTp search with an E-value threshold set at e−5. Then, SMART (http://smart.embl-heidelberg.de/ , accessed on 9 September 2023)[28] and CDD (www.ncbi.nlm.nih.gov/cdd , accessed on 9 September 2023)[29] were employed to verify the presence of a LOB domain in putative LBD proteins. Subsequently, the ExPASy-ProtParam tool (https://web.expasy.org/protparam/ , accessed on 10 September 2023)[30] was used to analyze the physical and chemical properties of all identified LBD proteins.Conserved motif, multiple sequence alignment, and phylogenetic analysis
-
The presumed patterns of LBD proteins were examined using the MEME suite (
https://meme-suite.org/meme/tools/meme , accessed on 10 September 2023)[31] with the following parameters: a motif number of 20, minimum width of six, maximum width of 50. CltustalW software was used for multiple sequence alignment of LBD proteins. Then, the WebLogo3 website (https://weblogo.threeplusone.com/create.cgi , accessed on 12 September 2023) was used to generate the conserved motif logos. The alignment of all LBD proteins from Arabidopsis, poplar, and Rosaceae species was constructed by a Muscle method. Following the alignment results, phylogenetic trees were created using the maximum likelihood (ML) method, and the bootstrap was set to 5,000. Using TBtools (v. 2.003) software[32], phylogenetic trees were created.Chromosome location, duplication, and syntenic analysis
-
The chromosomal lengths and locations of LBDs were extracted from the genome database. Then, the chromosomal location figures were created by TBtools software[32]. The segment and tandem duplication events of LBDs were analyzed by McscanX with default settings[33]. The intra-species synteny relationships of nine Rosaceae genomes LBDs and the inter-species synteny relationships among Arabidopsis, poplar, and nine Rosaceae genomes were identified by MCScanX[33], and the collinearity results were visualized using TBtools (v. 2.003)[32]. The Ks, Ka, and Ka/Ks values of gene pairs were calculated using the Ka/Ks calculator in TBtools (v. 2.003)[32].
GO annotation and cis-acting element analysis
-
To further explore the biological processes involved with LBD proteins, the GO annotation of nine Rosaceae LBDs was analyzed using GO Enrichment in TBtools (v. 2.003)[32]. We extracted the 2,000-bp promoter sequences upstream of each identified LBD member and submitted them to the PlantCARE database (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ , accessed on 19 September 2023)[34] for cis-acting element analysis. TBtools (v. 2.003) was used for visualization[32].Expression profiles of PmLBDs
-
To determine the tissue-specificity of the PmLBDs, we analyzed the expression patterns of PmLBDs using RNA-seq data obtained from five distinct tissues of the mei plant (bud, root, fruit, leaf, and stem) (Accession No. GSE40162)[35]. The responses of PmLBDs to natural cold were examined using RNA-seq data from three locations (Beijing [N39°54′, E116°28′], Chifeng [N42°17′, E118°58′], and Gongzhuling [N43°42′, E124°47′], China) and phenological stages (autumn, winter, and spring). To identify the responses of PmLBDs to the regulation of flowering, RNA-seq data (accession numbers: PRJNA833165, PRJNA832606, and PRJNA832060) were downloaded from the NCBI website (
www.ncbi.nlm.nih.gov , accessed on 22 September 2023)[36]. Additionally, we also examined PmLBDs expression at eight developmental stages of upright and weeping branches in the mei F1 population. The heatmaps of PmLBDs expression levels were created using TBtools (v. 2.003)[32].In the qRT-PCR procedure, total RNA was extracted from young stems using the RNAprep Pure Plant Plus Kit (DP441, TIANGEN). Subsequently, first-strand cDNAs were generated from 1 μg of total RNA using the PrimeScript™ RT reagent Kit with gDNA Eraser (RR047, TaKaRa, Bejing, China). To verify the accuracy of the results, nine PmLBDs were selected for qRT-PCR, and gene-specific primers used in qRT-PCR were designed by the NCBI primer tool (Supplemental Tables S1 & S2). The qRT-PCR was performed as described previously for the reaction system and conditions using the SYBR Premix Ex Taq II kit (RR820, TaKaRa) on a PikoReal real-time PCR system (Thermo Fisher Scientific, Waltham, MA, USA)[36]. The gene expression levels were determined by applying the 2−ΔΔCᴛ method, with PmPP2A serving as the internal reference gene[36].
-
Based on BLASTP and HMMER, a total of 406 LBDs were identified in the Rosaceae family. For Rosoideae, black raspberry (Rubus occidentalis), strawberry (Fragaria vesca), and Chinese rose (Rosa chinensis), 39, 34, and 39 LBDs were detected, respectively. The number of LBDs in the Amygdaloideae was comparable, with 42 in peach (Prunus persica) and 41 in the other three plants (Table 1). For Maloideae, the maximum number of LBDs was 69 in apple (Malus domestica) (Table 1). The proportion of LBDs was the highest in mei(P. mume var. tortuosa), followed by apple and hawthorn (Crataegus pinnatifida), and peach was the least (Table 1). The 406 LBD proteins of Rosaceae encoded 80 to 1,099 aa (amino acid), with molecular weights ranging from 8.90 kDa to 125.60 kDa and theoretical pI from 4.64 to 10.72. The mean hydropathicity value of just 12 proteins exceeded 0, suggesting that the majority of proteins exhibited hydrophilic properties (Supplemental Table S3). Subcellular localization prediction of all LBDs was localized in the nucleus (Supplemental Table S3).
Table 1. Number of LBDs in nine Rosaceae species.
Traditional subfamily Genus name Common name Species name Chromosome number Genome gene number Identified LBDs Proportion
of LBDsRosoideae Rubus Black raspberry Rubus occidentalis 8 33,286 39 0.12% Fragaria Strawberry Fragaria vesca 7 28,588 34 0.12% Rosa Chinese rose Rosa chinensis 7 39,669 39 0.10% Amygdaloideae Prunus Peach Prunus persica 8 47,089 42 0.09% Prunus Apricot Prunus armeniaca 8 30,436 41 0.13% Prunus Mei Prunus mume var. tortuosa 8 26,015 41 0.16% Maloideae Crataegus Hawthorn Crataegus pinnatifida 17 40,571 60 0.15% Pyrus European pear Pyrus communis 17 37,445 41 0.11% Malus Apple Malus domestica 'HFTH1' 17 44,677 69 0.15% Phylogenetic analysis, conserved motifs, and sequence alignment of LBD protein
-
To better analyze the evolutionary trajectory of LBD proteins in nine Rosaceae species, a maximum likelihood (ML) tree was constructed with LBDs from the Rosaceae family (406), A. thaliana (43), and P. trichocarpa (80). Based on the classification of Arabidopsis and poplar, 406 LBD proteins were divided into two major groups, Class I and Class II (Fig. 1a & Supplemental Tables S4, S5). Most proteins belong to Class I, which contained 349 (85.96%) members in nine species, while Class II had 57 (14.04%) LBD members (Fig. 1a, 1b & Supplemental Table S4). Subsequent studies revealed that Class I could be categorized into five subclasses (Class Ia-Ie), while Class II could be further separated into subclass IIa and subclass IIb. Each subclass included the LBDs of these 11 species, but there were differences in the distribution of members among different species (Fig. 1a, 1b & Supplemental Table S4). Subclass Ia had the largest number of LBDs (11) in apple, and subclass Ic contained the most members in hawthorn. Interestingly, subclass Ie had the highest number of members in most Rosaceae plants, such as strawberry, peach, mei, and apple (Fig. 1b & Supplemental Table S4). In addition, we found that the number of subclass Ia, Ic, and IIa in Rosoideae was less than that in Maloideae. In the Rosoideae and Amygdaloideae, the number of subclass IIa and IIb was consistent (Fig. 1b & Supplemental Table S4).
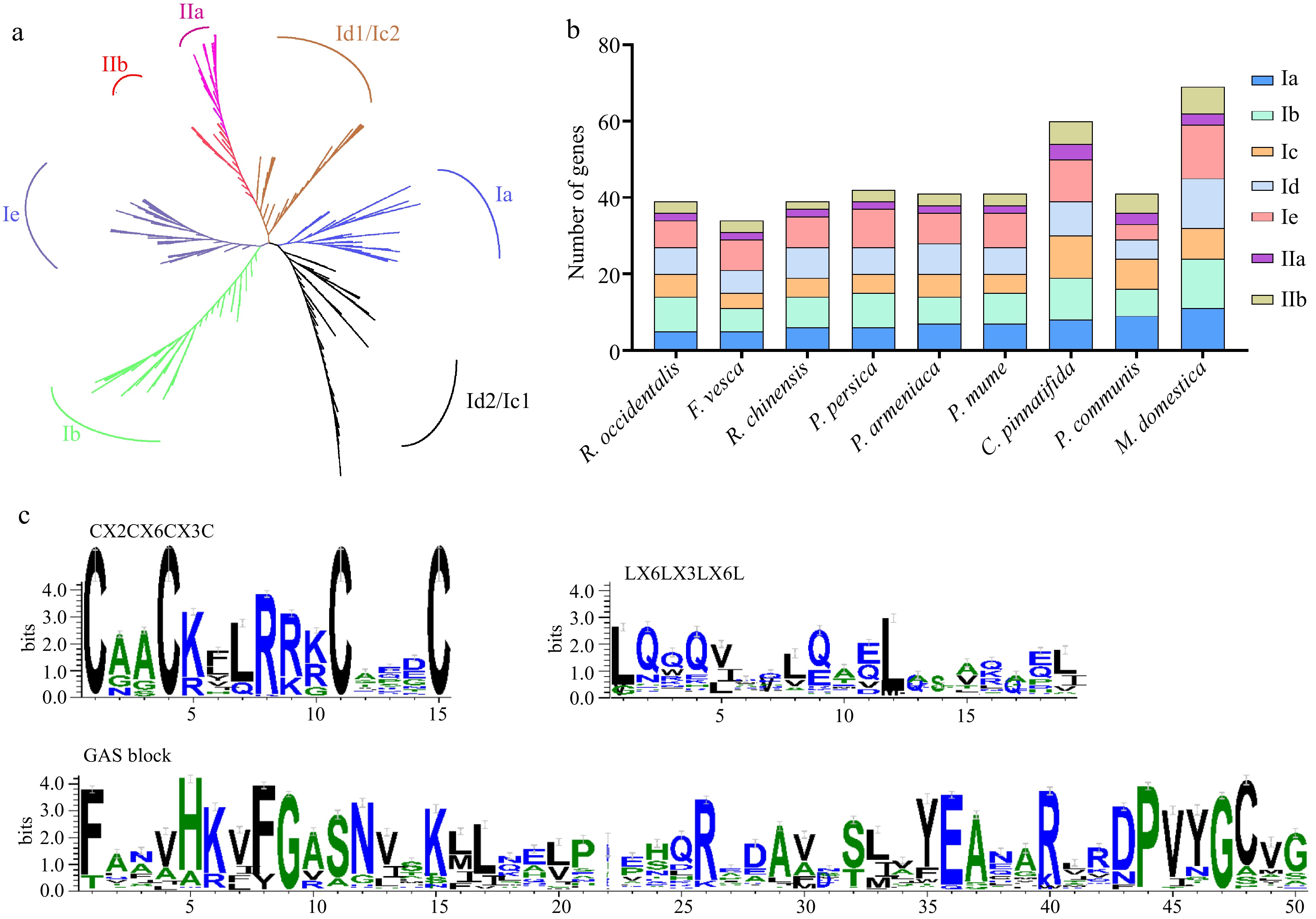
Figure 1.
Phylogenetic tree, conserved domains, and the gene numbers of the subfamily in nine Rosaceae species. (a) ML phylogenetic tree of LBD proteins in 11 plant genomes. (b) The number of genes identified in different classes of the LBD family. (c) Analysis of three conserved domains of LBD proteins in nine Rosaceae genomes.
The investigation of protein domain positioning and structure involved the utilization of ClustalW for carrying out multiple sequence alignment. Additionally, conserved motif logos were developed using the WebLogo3 website. Consequently, nearly all LBDs exhibited the zinc finger-like domain (CX2CX6CX3C) and GAS blocks, whereas Class II LBDs did not include the leucine zipper-like motif (LX6LX3LX6L) (Fig. 1c).
To delve more into the functional variety and evolutionary relationship of LBDs in species belonging to the Rosaceae family, we constructed an independent phylogenetic tree for each subclass and analyzed motifs and domains within these proteins. The subclass exhibited significant variation in both the quantity and diversity of motifs (Fig. 2, Supplemental Figs S1−S5). For example, subclass IIa possessed the lowest number of motifs and only nine types of motifs, while subclass Ie possessed the highest number of motifs with 15 types. Besides, the conserved motifs 1, 2, 10, and 13 were shared by each subclass. Class II did not contain motifs 4 and 6, but these only contained motifs 5 and 12. Subclass Id and Ie were the only subclasses that included motif 6, while motif 16 was exclusively found in subclass IIb. The presence of specific motifs in the LBD subclass indicated that they also had specific roles.

Figure 2.
Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass Ia subfamily members.
Chromosomal localization and evolutionary analysis of the LBDs
-
To gain a deeper understanding of the evolutionary connection between LBDs in Rosaceae species, we conducted a study on the chromosome localization, collinearity analysis, and gene duplication relationship of LBDs in the nine Rosaceae genomes. Through chromosome localization, 406 LBDs in nine Rosaceae genomes were unevenly distributed across the chromosome (Fig. 3 & Supplemental Figs S6, S7). We also found that except for Maloideae, most species had LBDs distribution on each chromosome (Fig. 3, Supplemental Figs S6, S7). Specifically, no LBDs were located on chromosome 4 and 13 in pear and apple (Supplemental Fig. S7b, S7c). Chromosome 7 (Chr7) had the maximum number of LBDs in black raspberry, strawberry, pear, and apple, with 12, 9, 10, and 7, respectively (Supplemental Figs S6a, S6b, S7b & S7c). Nine and 11 LBDs were located in Chr5, which was the largest number in apricot and mei, respectively (Fig. 3b, 3c).
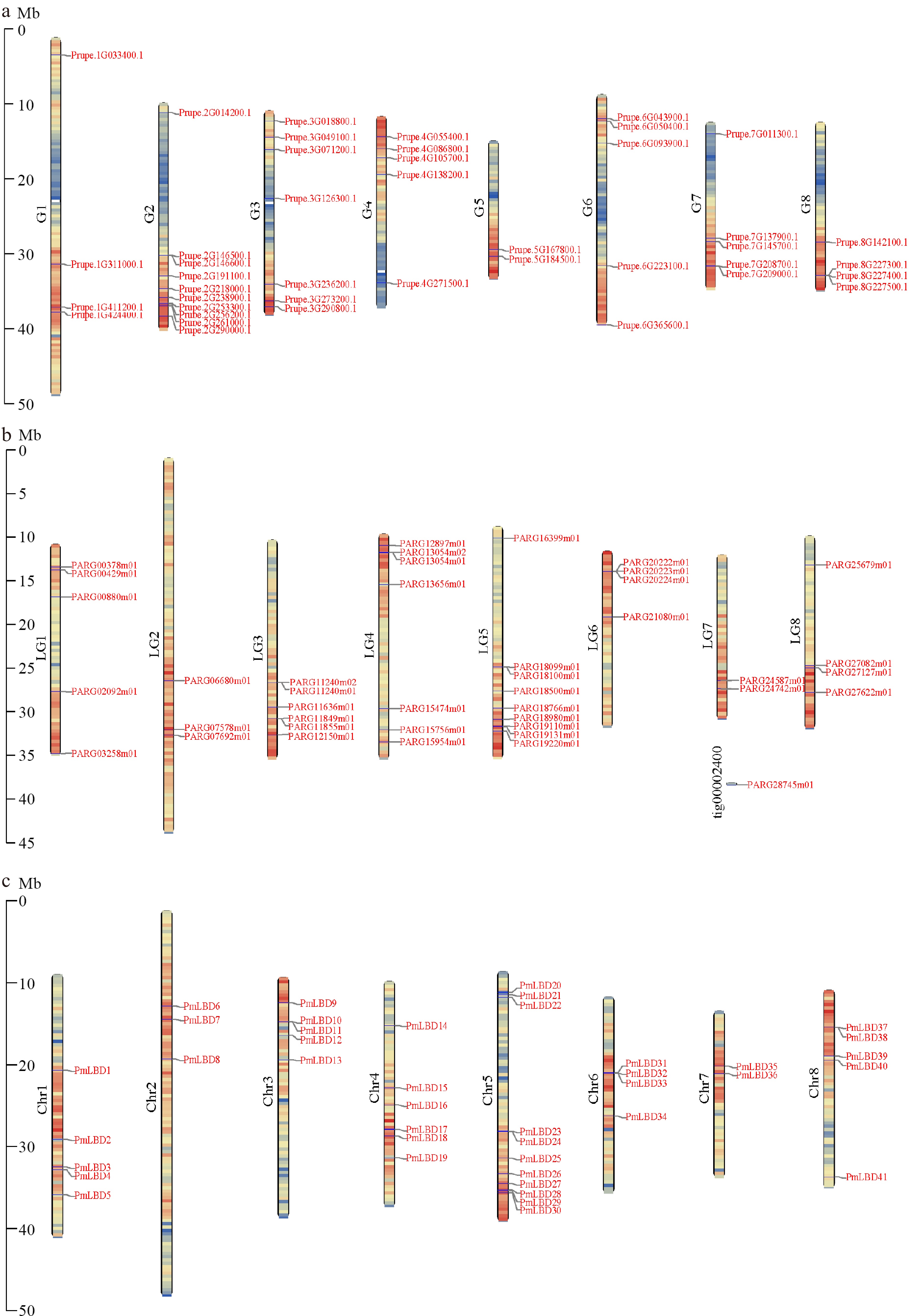
Figure 3.
Chromosome distribution of LBDs in the Amygdaloideae. (a) Chromosome distribution of LBDs in P. persica. (b) Chromosome distribution of LBDs in P. armeniaca. (c) Chromosome distribution of LBDs in P. mume.
In addition, collinearity analysis showed that the most segment duplication gene pairs occurred in Maloideae, such as 87 gene pairs in hawthorn, followed by Amygdaloideae, and the least in Rosoideae, such as only six in Chinese rose (Fig. 4, Supplemental Fig. S8 & Supplemental Table S6). Notably, the tandem duplication gene pairs showed significant similarity among the species of the Rosaceae family. In apple, the highest number of tandem duplication gene pairs observed was five, whereas in pear, the lowest number was two (Supplemental Fig. S3b, 3c & Supplemental Table S7). The prevalence of segment duplication genes, as opposed to tandem duplication genes, indicates that segment duplication is the primary factor responsible for the expansion of LBDs in the Rosoideae and Amygdaloideae. Subsequently, genome collinearity of LBDs among the Rosaceae family, A. thaliana, and P. trichocarpa was conducted on account of species’ evolutionary relationships. The findings indicated a significant collinearity relationship across Rosaceae plants, as depicted in Fig. 5. In Rosoideae, 46 and 42 pairs of orthologous LBDs were detected between black raspberry and strawberry and strawberry and Chinese rose, respectively (Fig. 5). A total of 53 and 49 homologous gene pairs were found in Amygdaloideae (peach vs apricot, apricot vs mei) (Fig. 5). For Malodieae, there were the most gene pairs, with 103 pairs between hawthorn and pear (Fig. 5).

Figure 4.
Collinearity of segmental duplication gene pairs of LBDs in six Rosaceae species. (a) Collinearity of segmental duplication gene pairs of LBDs in R. occidentalis. (b) Collinearity of segmental duplication gene pairs of LBDs in F. vesca. (c) Collinearity of segmental duplication gene pairs of LBDs in R. chinensis. (d) Collinearity of segmental duplication gene pairs of LBDs in P. persica. (e) Collinearity of segmental duplication gene pairs of LBDs in P. armeniaca. (f) Collinearity of segmental duplication gene pairs of LBDs in P. mume. The red lines represent the segment duplication (SD) gene pairs of the LBDs.

Figure 5.
Collinearity analysis of LBDs in different genomes. Colored circular rectangles denote the chromosomes of different plants. The green lines represent gene pairs with a collinear relationship. The grey lines represent other collinear gene pairs of non-LBD gene family members across genomes.
To conduct a more in-depth examination of the rate at which LBDs have evolved in nine Rosaceae species, we computed the Ka (non-synonymous substitution) to Ks (synonymous substitution) ratio for each pair of genes. In our study, the Ks value of gene pairs was mainly distributed at 1.0 to 2.5 in black raspberry, strawberry, apricot, mei, and pear (Fig. 6a & Supplemental Data S1). The main distribution of Ks in other Rosaceae species was 2.0 to 2.5 (Fig. 6a & Supplemental Data S1). In addition, the value of Ks peaked at 2.0-2.5 in strawberry, apricot, mei, and pear, while the peak value was 1.5-2.0 in the other six plants (Fig. 6a & Supplemental Data S1). The majority of the LBD gene pairs exhibited Ka/Ks ratios below 1 (Fig. 6b & Supplemental Data S2), indicating that these genes are likely subject to purifying selection. However, it is worth mentioning that there was one gene pair in peach and two gene pairs in black raspberry with a Ka/Ks value greater than 1 (Fig. 6b & Supplemental Data S2), implying that these genes may undergo functional divergence owing to positive selection.
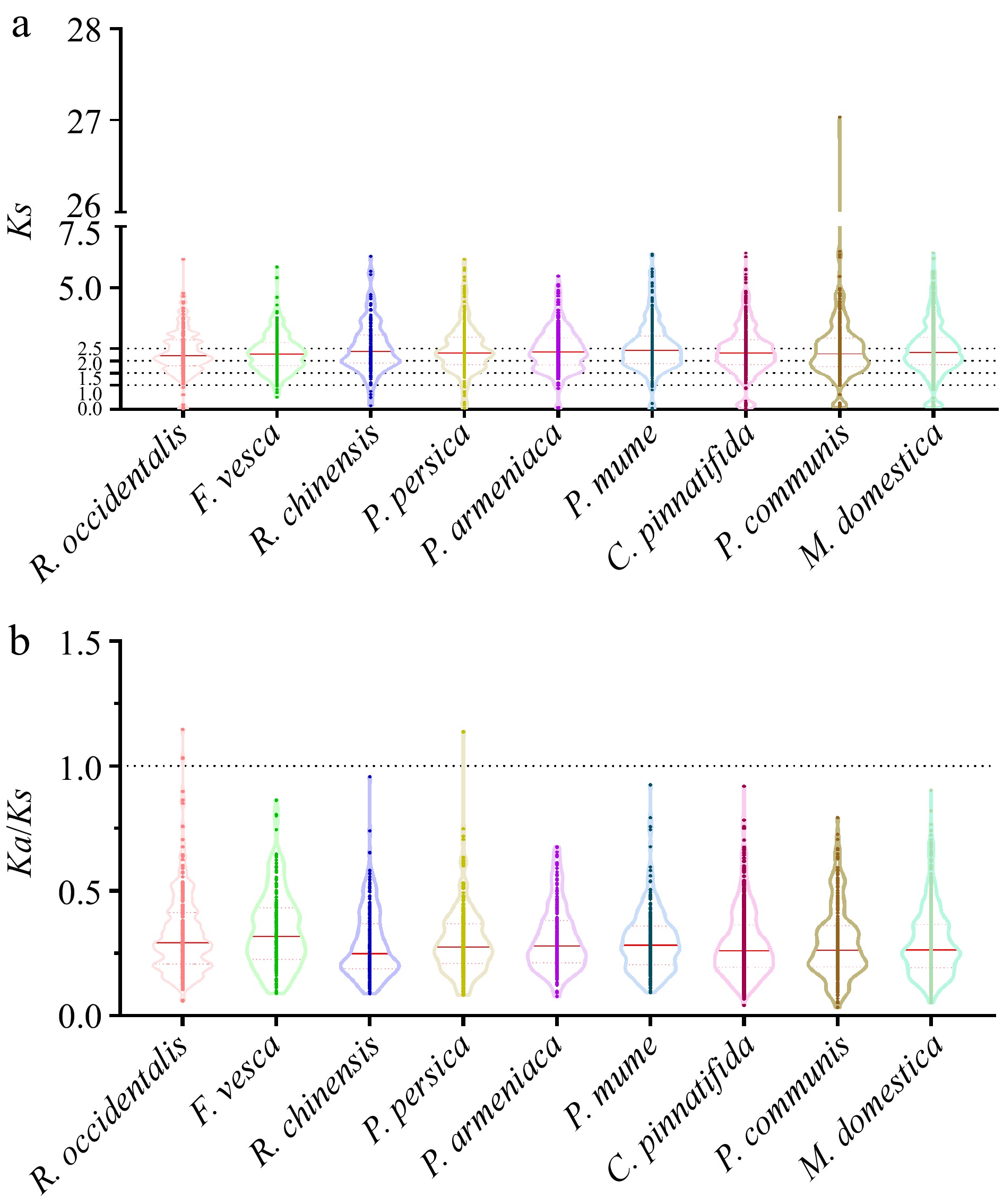
Figure 6.
The Ks and Ka/Ks values of LBDs in nine Rosaceae genomes. (a) The distribution of Ks values among LBDs in nine Rosaceae genomes. (b) The distribution of Ka/Ks values among LBDs in nine Rosaceae genomes.
Functional prediction of the LBDs
-
To further explore the biological processes involved in LBDs, we performed a gene ontology (GO) analysis of Rosaceae LBDs. According to the cellular component results, LBDs were involved in the nucleus, membrane-bounded organelle, intracellular membrane-bounded organelle, organelle and intracellular organelle (Fig. 7a). Regarding molecular function, LBDs primarily participate in protein dimerization activity and protein binding. In addition, these genes were implicated in more than 130 biological processes, including post-embryonic, plant organ, root, flower, and other developmental and morphogenetic processes, as well as phloem or xylem histogenesis, cellular response to jasmonic acid stimulus and jasmonic acid mediated signaling pathway (Fig. 7a).
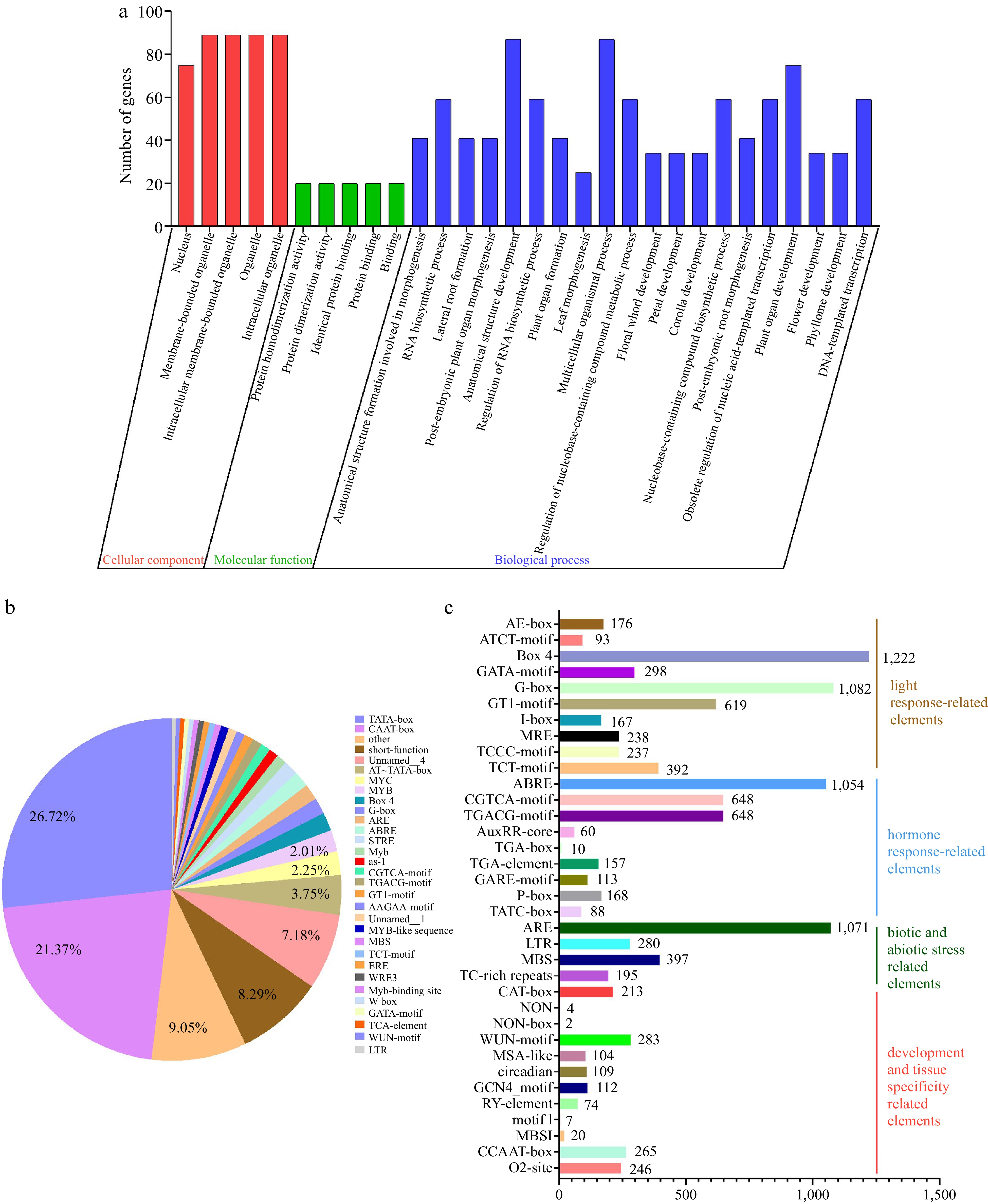
Figure 7.
GO and cis-elements analysis of LBDs in nine Rosaceae species. (a) GO analysis of LBDs in nine Rosaceae species. (b) The proportion of cis-elements predicted in the promoters of LBDs. (c) Numbers of the cis-elements involved in light response, hormone response, biotic and abiotic stress, development, and tissue specificity.
To explore the potential regulatory mechanisms of LBDs, cis-acting element analysis was performed on the region 2,000 bp upstream of 406 LBDs using the PlantCARE database. The findings indicated that a total of 70,479 cis-acting elements were identified, with an average of 173 per gene (Fig. 7b & Supplemental Fig. S9). The promoter region of LBDs exhibited a widespread presence of common regulatory components, namely the CAAT-box and TATA-box, which accounted for 21.48% and 26.61% respectively. Subsequently, 20 major cis-elements were selected for further analysis (Fig. 7b). These cis-elements mainly contained: (1) light response-related elements, with an average of 12 elements per gene; (2) hormone response-related elements, such as abscisic acid, MeJA, auxin, gibberellin; (3) biotic and abiotic stress-related elements, including anaerobic induction, low-temperature, drought-inducibility, defense, and stress responsiveness; (4) development and tissue specificity related elements, such as meristem expression, wound-responsive, cell cycle regulation, circadian control, endosperm expression, seed-specific, root-specific (Fig. 7b, c & Supplemental Fig. S9). Furthermore, despite the distribution of various cis-elements throughout the promoter, the presence of similarly organized cis-acting elements on related gene promoters implies that these genes may have comparable roles (Supplemental Fig. S9). Overall, these results indicated that LBDs may play important roles as transcription factors in many biological processes and could respond to hormone response and stress.
Expression pattern analysis of PmLBDs in P. mume
-
To characterize the tissue-specific PmLBD gene in P. mume, the expression patterns of PmLBD family members were based on RNA-seq data. Among 41 PmLBDs, 26 PmLBDs were expressed in at least tissues (bud, root, fruit, leaf, and stem) (Fig. 8a). The PmLBDs that were considered to be tissue-specifically expressed with RPKM > 2-fold over other tissues were as follows: PmLBD3/6/13/27/29/31/34, PmLBD7/15/17/25/36/40, PmLBD8/23/24, PmLBD28 were expressed in the roots, buds, fruit and stems, respectively (Fig. 8a). The other genes were expressed in two or more tissues, among which PmLBD1 was highly expressed in all five tissues (RPKM > 40) (Fig. 8a). These findings implied that the growth and development process of tissues were regulated by these PmLBDs.

Figure 8.
Expression pattern of PmLBDs in different tissues and different developmental stages of flower buds. (a) Hierarchical clustering of expression profiles of PmLBDs in different tissues (bud, fruit, leaf, root, and stem). (b) Expression profiles of PmLBDs in the flower bud during dormancy release.
To look into the potential role of PmLBDs in the regulation of blooming, particularly in the process of floral bud break, we assessed the expression levels of PmLBDs at four different stages of floral bud dormancy release in P. mume. As shown in Fig. 8b & Supplemental Fig. S10, PmLBD2 exhibited a continuous upregulation with floral bud exit dormancy, while PmLBD12/35 showed a downregulation trend. PmLBD19 expression was suppressed in the endodormancy process, increased during ecological dormancy, and decreased sharply at bud flush, while PmLBD6 was up-regulated during endodormancy and decreased after ecological dormancy. These results demonstrate that these PmLBDs function in floral bud dormancy release.
To examine how PmLBDs react to cold stress in mei, we analyzed the expression patterns in the stem at three different locations throughout three time periods. The expression level of PmLBDs varied greatly at different geographic locations and in different periods (Fig. 9 & Supplemental Fig. S11). For example, PmLBD1/6/13/17 showed large expression levels under all three locations at the same time. In addition, PmLBD13 exhibited an initial downregulation followed by an upregulation trend at three test sites, while PmLBD26 showed an opposite trend (Fig. 9 & Supplemental Fig. S11). Notably, some genes showed inconsistent expression at three test sites. For example, the expression of PmLBD1 exhibited down-regulation in winter and up-regulation in spring in Beijing, a continuous down-regulation trend in Chifeng, and upregulation followed by downregulation in Gongzhuling (Fig. 9a). PmLBD19 showed a continuous up-regulation in Beijing, down-regulation in winter, and up-regulation in spring in Chifeng, and an increase followed by a downregulation in Gongzhuling (Fig. 9b). These results suggest that PmLBDs were involved in the response to cold stress in P. mume.

Figure 9.
Expression pattern of PmLBDs in different locations and seasons. (a) Hierarchical clustering of expression profiles of PmLBDs in different locations. (b) Hierarchical clustering of expression profiles of PmLBDs in different seasons. BJ, Beijing; CF, Chifeng; GZL, Gongzhuling.
We also examined the expression of PmLBDs at eight developmental stages of upright and weeping branches in the mei F1 population. The PmLBDs showed a large variation in expression patterns during branch development (Fig. 10). The expression of PmLBD5 showed a continuous upregulation at eight developmental stages, and PmLBD28/30/40 exhibited an up-regulation followed by a decrease. Notably, PmLBD6 was consistently higher in weeping branches than in upright branches, while PmLBD20 showed an opposite trend (Fig. 10). To verify the accuracy of the transcript levels of PmLBDs in transcriptome data, nine candidate genes were selected based on the subfamily classification and differential gene clustering. Their expression level in upright and weeping branches was investigated using qRT-PCR with PmPP2A as a reference gene. Finally, RNA-seq data were consistent with the qRT-PCR results (Fig. 11).

Figure 10.
Expression pattern of PmLBDs in upright and weeping branches. U1−U8, eight developmental stages of upright branches in the mei F1 population; W1−W8, eight developmental stages of weeping branches in the mei F1 population.
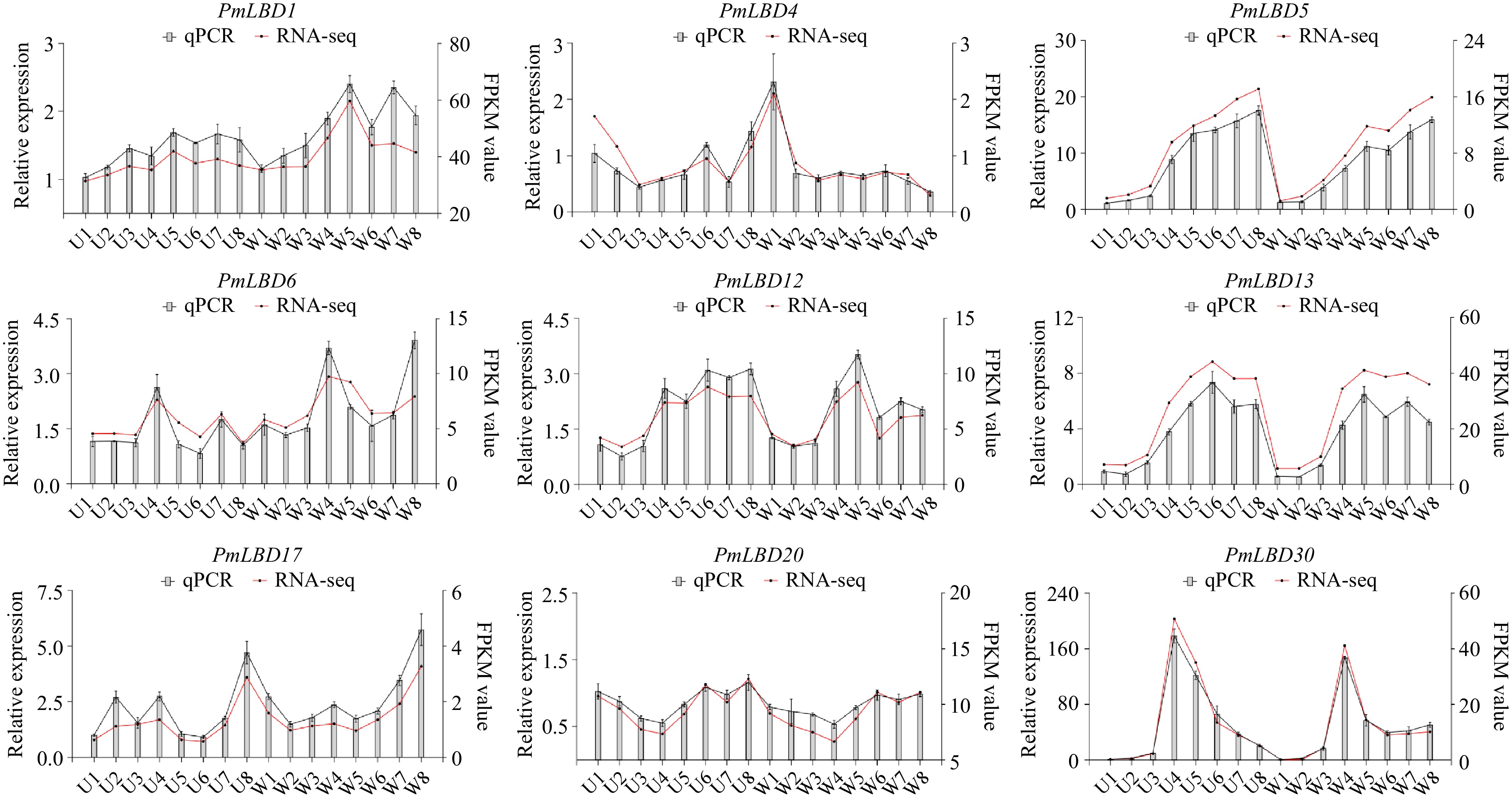
Figure 11.
qRT-PCR analysis of nine PmLBDs in upright and weeping branches. U1−U8, eight developmental stages of upright branches in the mei F1 population; W1−W8, eight developmental stages of weeping branches in the mei F1 population. The relative quantification method (2−ΔΔCᴛ) was used to evaluate quantitative variation. Error bars represent standard error for three replicates.
-
The LBDs, which are exclusive to plants, play a crucial role in regulating a wide range of biological activities such as plant secondary metabolism, growth and development, and response to different types of stress[11,13,22]. Due to their crucial function in plant development, LBDs have been extensively researched in several plant species. Rosaceae is one of the important plant families, however, comparative studies on LBDs in Rosaceae remain unknown. In this study, 42 AtLBDs from Arabidopsis were utilized to identify LBD proteins in nine representative Rosaceae plants, with the number of LBDs ranging from 34 to 69, independent of genome size. The number of LBDs was similar in most selected plants, but was far greater in hawthorn and apple with 60 and 69 LBDs, respectively. The presence of a large number of LBDs in hawthorn and apple may be related to the widespread occurrence of duplication events in their genomes[37,38].
Four hundred and six LBD proteins could be categorized into two classes: Class I (349, 85.96%) and Class II (57, 14.04%), and LBDs in Class I was significantly higher than in Class II among all of the selected Rosaceae plants, which is consistent with previous results[11]. Similar to the previous phylogenetic tree results[11,39], those proteins were divided into seven major subclasses, and the LBDs of these 12 species were distributed in each subclass. The classification was further supported by gene motif analysis and structural domains, indicating that genes in the same subclass usually have similar biological functions. Recent studies have reported that root-type-specific regulation by subclass IB LBDs is deeply conserved[22]. For example, Solyc09g066270, a subclass IB LBD, could specifically regulate the earliest stage of root initiation[22]. Therefore, we hypothesize that subclass IB LBDs may play a deeply conservative role in lateral root initiation and provide reference genes for difficult-to-root plants in Rosaceae, especially in the Prunus genus.
Gene duplication is a crucial factor in the process of evolution and the growth of gene families. Segmental and tandem duplication are the primary mechanisms for gene family growth[40]. A total of 242 pairs of LBD duplication genes were discovered in the nine Rosaceae genomes under analysis. Out of these, 215 pairs were recognized as segment duplication genes, while only 27 pairs were classified as tandem duplication. This finding demonstrates that segmental duplication events play a crucial role in driving the proliferation of the LBD family in the Rosoideae and Amygdaloideae. In addition, synteny analysis revealed a great collinearity relationship among Rosaceae plants, especially those in Malodieae, which proved that the LBDs are relatively conserved in Rosaceae, suggesting that the functions of these homologous genes may be consistent. The Ka/Ks ratio was used to measure the selection pressure experienced by the gene pairs. Previous studies have shown that LBDs proceed with a purifying selection in moso bamboo (Phyllostachys edulis) and ginkgo (Ginkgo biloba)[41,42]. Similarly, almost all LBDs in selected Rosaceae in this study have undergone purifying selection. The presence of a few LBDs may undergo functional divergence owing to positive selection in black raspberry and peach, indicating that functional differentiation of these genes may occur, which is worthy of further research.
GO analysis and promoter cis-acting element prediction can determine the possible functions of LBDs. AtLBD3 and AtLBD4 have been reported to activate secondary growth through cytokinin signaling[43]. AtLBD16/17/18/29 plays key roles in plant regeneration programs mediated by the auxin signals[12]. Our study revealed that the LBD promoters contain many kinds of hormone response-related elements (abscisic acid, MeJA, auxin, and gibberellin). Thus, we postulated that LBDs might play a role in plant development through their reaction to hormones. Furthermore, the LBD promoters were shown to contain components associated with light, low temperature, drought, defense and stress responses, meristematic organization, and tissue specialization. These findings indicate that LBDs have a significant impact on various biological processes and may be involved in hormone response and stress. This is consistent with the results of the PmLBDs expression analysis under low temperature and tree architecture in this study.
The LBDs have been extensively reported to play a crucial role in controlling the development of many plant parts, including roots, flowers, leaves, and stems. This finding is congruent with the results obtained from the GO enrichment study. GO annotations of 470 LBDs contained a variety of plant organ development and formation, including post-embryonic, plant organ, root, and flower. In Arabidopsis, AtLBD13[44], AtLBD16[45], and AtLBD33[46] were shown to play key roles in controlling lateral root development. PmLBD3 with AtLBD13, PmLBD27 with AtLBD33, and PmLBD31 with AtLBD16 were respectively in the same subclades. Notably, the three genes were specifically expressed in the root. Therefore, we hypothesized that PmLBD3/27/31 may be involved in lateral root development.
Previous studies showed that LBDs respond positively to various abiotic stresses. For example, in Ginkgo, GbLBD31, a pleiotropic regulator, was significantly expressed under drought and cold stress[42]. In banana, MaLBD5 may be associated with MeJA-induced cold tolerance and activated jasmonate biosynthesis gene[23]. In our study, the response of PmLBDs to cold stress was revealed at three sites for three periods by analyzing previous transcriptome data. Our investigation revealed significant variation in the expression of PmLBDs across different geographic locations and periods. Additionally, we observed inconsistent expression of certain genes among the three investigated loci. Notably, four differentially expressed genes (DEGs) were predicted to contain low-temperature response elements, eight DEGs contained MeJA response elements, and three DEGs contained both low-temperature and MeJA response elements, indicating that these genes potentially have a role in the development of cold tolerance in mei driven by MeJA and provide potential candidate genes for future research on cold tolerant molecular breeding in mei.
It is known that flowering transition is controlled by the gene regulatory network. In rice, OsLBD37 and OsLBD38 were found to delay flowering by down-regulating Hd3a and RFT1 expression[47]. In transgenic Arabidopsis, CsLBD37 overexpression affects nitrogen-responsive gene expression and nitrate content, which may regulate early flowering in plants through nitrogen signaling[48]. The study observed significant alterations in the expression levels of many LBDs, suggesting their potential involvement in the regulation of blooming.
Prior research has demonstrated that the abnormal growth of phloem, namely the lack of phloem fibers, had a significant role in the development of weeping traits in mei[49]. PtaLBD1 and PtaLBD4 regulate the development of the secondary phloem by inhibiting the expression of identity genes in the meristem[11]. In overexpression poplar, EgLBD29 controls secondary growth especially the development of phloem fiber[11]. This suggests that LBDs may regulate secondary growth in plants, especially phloem fiber. In this study, PmLBD6 was consistently higher in weeping branches than in upright branches, while PmLBD20 showed an opposite trend. Notably, in mei, PmLBD30, the ortholog of EgLBD29, exhibited an up-regulation followed by a decrease, which is hypothesized to possibly play a role in branch development and can be studied as a candidate gene for the formation of a weeping trait in mei.
-
In Rosaceae, 39, 34, 39, 42, 41, 41, 41, 60, 41, and 69 LBDs were identified in black raspberry, strawberry, Chinese rose, peach, apricot, mei, pear, hawthorn, and apple, respectively. Among them, the LBDs were classified into seven major subclasses. The primary factor responsible for the amplification of LBDs in Rosaceae plants was the duplication of segments. Phylogenetic tree and RNA-seq data showed that PmLBD27/31 were tissue-specifically expressed in the roots. Transcription sequencing data from three locations for three periods indicated that PmLBD17/19/41 were induced by low temperature and they all contained both low-temperature and MeJA response elements. Moreover, PmLBD30 exhibited an up-regulation followed by a decrease in the developmental stages of branches. In summary, our studies offer novel perspectives on the evolutionary connection between the LBD family in Rosaceae and the role of LBDs in mei.
-
The authors confirm contribution to the paper as follows: study conception and design: Liu W, Zheng T; conducted experiments and material collection: Zheng T, Liu W, Guo X; data analysis: Liu W, Guo X, Zheng T; conducted fieldwork and material maintenance: Liu W, Guo X, Li X, Wang J, Cheng T, Zhang Q; modified the language modification: Ahmad S; draft the manuscript: Liu W, Guo X, Zheng T; manuscript revision and finalization: Zheng T, Cheng T. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
The research was supported by the National Natural Science Foundation of China (No. 32371947), the Beijing High-Precision Discipline Project, Discipline of Ecological Environment of Urban and Rural Human Settlements, and the Special Fund for Beijing Common Construction Project.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Weichao Liu, Xiaoyu Guo
- Supplemental Tables S1 Primers used in this study.
- Supplemental Tables S2 The reference sequences used for gene-specific primers in Prunus mume.
- Supplemental Tables S3 The physicochemical parameters of all LBD proteins in ten Rosaceae genomes.
- Supplemental Tables S4 The number of LBDs of eleven species in each subclass.
- Supplemental Tables S5 Classification of LBD in each subclass.
- Supplemental Tables S6 Segmental duplication events of nine Rosaceae species.
- Supplemental Tables S7 Tandem duplication events of nine Rosaceae species.
- Supplemental Date S1 Ks values of nine Rosaceae species
- Supplemental Date S2 ka/Ks values of nine Rosaceae species
- Supplemental Fig. S1 Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass Ib subfamily members.
- Supplemental Fig. S2 Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass Ic subfamily members.
- Supplemental Fig. S3 Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass Id subfamily members.
- Supplemental Fig. S4 Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass Ie subfamily members.
- Supplemental Fig. S5 Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass IIa and IIb subfamily members. (A) Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass IIa subfamily members. (B) Phylogenetic evolutionary tree, motifs distributions, and domains of the subclass IIb subfamily members.
- Supplemental Fig. S6 Chromosome distribution of LBDs in the Rosoideae. (A) Chromosome distribution of LBDs in R. occidentalis. (B) Chromosome distribution of LBDs in F. vesca. (C) Chromosome distribution of LBDs in R. chinensis.
- Supplemental Fig. S7 Chromosome distribution of LBDs in the Maloideae. (A) Chromosome distribution of LBDs in C. pinnatifida. (B) Chromosome distribution of LBDs in P. communis. (C) Chromosome distribution of LBDs in M.domestica.
- Supplemental Fig. S8 Collinearity of segmental duplication gene pairs of LBDs in the Maloideae. (A) Collinearity of segmental duplication gene pairs of LBDs in C.pinnatifida. (B) Collinearity of segmental duplication gene pairs of LBDs in P.communis. (C) Collinearity of segmental duplication gene pairs of LBDs in M.domestica. The red lines represent the segment duplication (SD) gene pairs of the LBDs.
- Supplemental Fig. S9 The distribution of the main 20 cis-elements in LBD gene promoters.
- Supplemental Fig. S10 Expression pattern of PmLBDs in different developmental stages of flower buds.
- Supplemental Fig. S11 Expression pattern of PmLBDs in different locations and seasons. BJ, Beijing; CF, Chifeng; GZL, Gongzhuling.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu W, Guo X, Zheng T, Li X, Ahmad S, et al. 2024. Genome-wide identification and characterization of the Lateral Organ Boundaries Domain (LBD) gene family in nine Rosaceae species and expression pattern in Prunus mume. Ornamental Plant Research 4: e007 doi: 10.48130/opr-0024-0005 

Genome-wide identification and characterization of the Lateral Organ Boundaries Domain (LBD) gene family in nine Rosaceae species and expression pattern in Prunus mume
- Received: 13 December 2023
- Revised: 21 January 2024
- Accepted: 25 January 2024
- Published online: 04 March 2024
Abstract: Transcription factors (TFs) encoded by the lateral organ boundaries domain (LBD) gene family are known to control many plant-specific developmental processes. However, the comparative analysis of the LBD gene family in Rosaceae species and its expression pattern in mei remains unclear. Here, we identified a total of 406 LBDs in nine Rosaceae species, including 39 in black raspberry (Rubus occidentalis), 34 in strawberry (Fragaria vesca), 39 in Chinese rose (Rosa chinensis), 42 in peach (Prunus persica), 41 in apricot (Prunus armeniaca), 41 in mei (Prunus mume var. tortuosa), 60 in pear (Pyrus communis), 41 in hawthorn (Crataegus pinnatifida) and 69 in apple (Malus domestica), respectively. The LBDs of nine Rosaceae species were classified into seven major subclasses. The chromosome localization, collinearity analysis, and gene duplication relationship revealed that segment duplication was the main driving force for the amplification of LBDs in the Rosoideae and Amygdaloideae. Ka/Ks analysis suggested most of the LBD gene pairs might be under purification selection. GO and cis-acting elements analysis showed that LBDs may play important roles in many biological processes and could respond to hormones and stresses. RNA-seq data showed that PmLBD17/19/41 genes contained both low-temperature and MeJA response elements and played a significant variation across different geographic locations and periods. PmLBD30, the ortholog of EgLBD29, exhibited an up-regulation followed by a decrease, which is hypothesized to possibly play a role in the formation of a weeping trait in mei. Our studies offer important data about the development of the LBD family in Rosaceae and the subsequent validation of LBDs' functional genes in P. mume.
-
Key words:
- Rosaceae /
- LBD gene family /
- Prunus mume /
- Phylogenetic analysis /
- Collinearity analysis /
- Expression pattern