









-
MicroRNAs (miRNAs) are a class of endogenous non-coding RNAs of approximately 20–22 nucleotides in length that are widely distributed in animals and plants[1]. They negatively regulate gene expression via mRNA cleavage, translational repression, or both. MicroRNA156 (miR156), targeting a subset of SQUAMOSA PROMOTOR BINDING PROTEIN-LIKE (SPL) genes, is one of the most evolutionarily conserved miRNAs in plants[2]. It was initially found in Arabidopsis thaliana[3]. Since then, a large number of miR156 family members have been identified and functionally verified in various horticulture plants. Based on the species registered in the miRBase database (
www.mirbase.org )[4], the miR156 sequences of at least 16 horticultural plants have been revealed to have different numbers of miR156 precursors and mature miR156 (Fig. 1). As a key player in balancing plant age and other biological functions, miR156 is involved in the regulation of a wide range of horticultural traits. Moreover, miR156 plays a remarkably conserved role throughout the angiosperms[5]. Hence, miR156 could potentially be an emerging means suited for biotechnological modification in horticultural plants. In this review, we mainly summarize the multifunctional regulation of miR156 in horticultural plants and also discuss perspectives for future studies.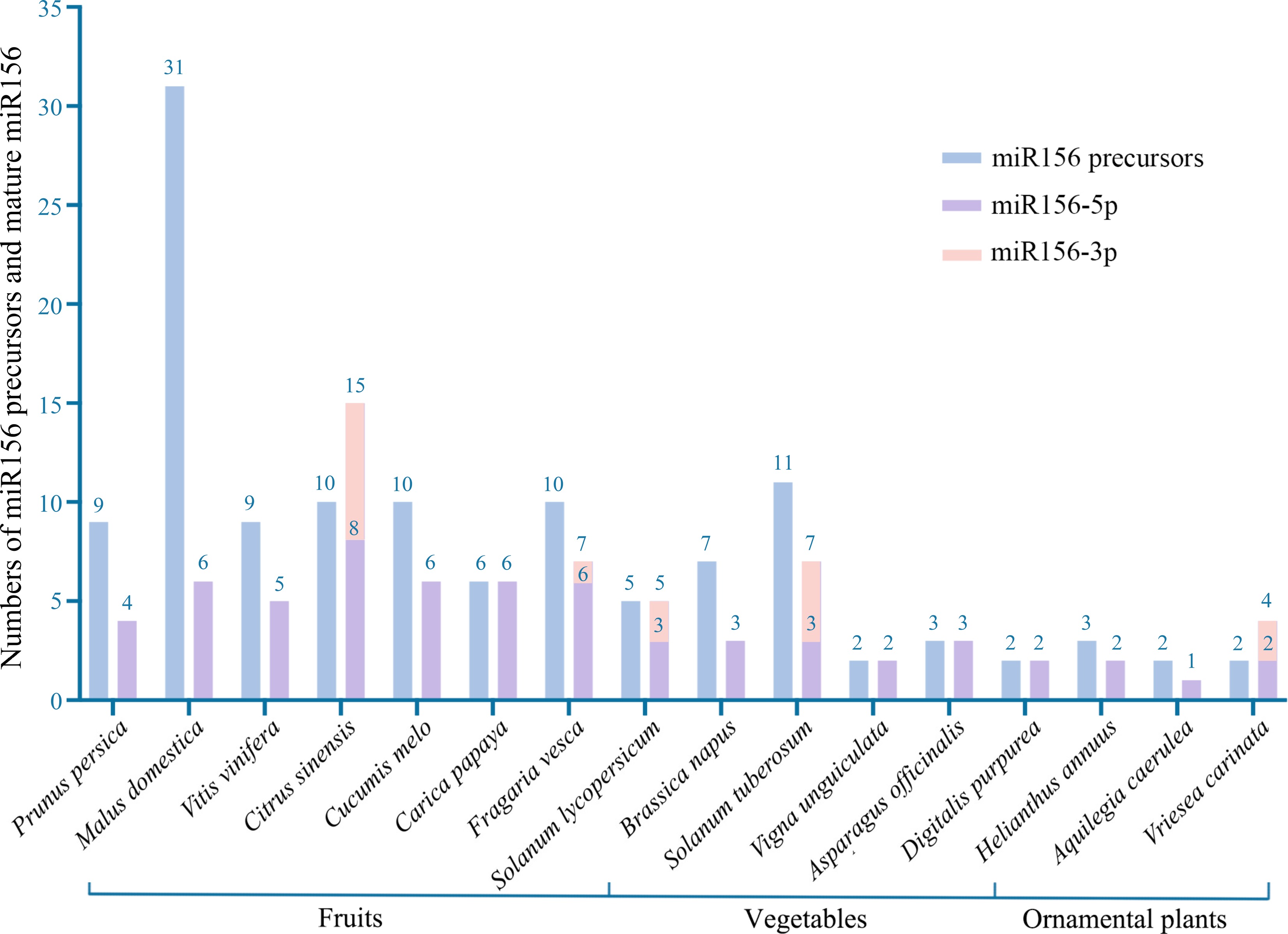
Figure 1.
Number of miR156 precursors and unique mature sequences in horticultural plants published in the miRbase database. Identical mature sequences from different precursors were considered as one sequence.
-
Based on 141 sequences of horticultural plant species released from miRBase 21.0, the miR156 sequences from 16 horticultural plants belong to 26 subfamilies (miR156a-z) (Fig. 2). A study using a total of 310 miR156 sequences from 51 plant species for phylogenetic analysis found a significant expansion in the number of miR156 family members from lower to higher plants[6]. In apple (Malus domestica), 15 different miR156 mature sequences are derived from 31 miR156 precursors; while there is only one unique miR156 sequence in Aquilegia caerulea from two precursors. Though various horticultural plant species exhibit a divergence in terms of the number of the identified mature miR156s, ranging from 1 to 15 (Fig. 1), their sequences show great conservation, according to the alignment of miR156 subfamilies from different horticultural plants (Fig. 2). All the base variations occur at the miR156 skeleton sequence 'TGACAGAAGAGAGTGAGCAC'. Additionally, the variation sites are primarily found in the 11th and 14th base (near the cleavage site) at the 5' end of miR156[6]. These two sites are nearly consistent with the cleavage sites of miRNAs on their target genes, which implies that different members of SPL were targeted by miR156s with corresponding base variation, and mismatch caused by point mutation could perturb the miR156-directed transcriptional cleavage and translation repression[7,8]. miR156-3p is produced from the other arm of miR156 precursors. Unlike common miR156 (miR156-5p) sequences, the strands of miR156-3p show a large divergence and are only found in five horticultural plant species, i.e., Citrus sinensis, Fragaria vesca, Solanum lycopersicum, Solanum tuberosum and Vriesea carinata (Fig. 1). Due to the highly conserved nature, 5p sequences of miR156 have been fairly well-studied compared with 3p sequences. However, it is important to note that evidence has emerged that 3p sequences also serve specific biological functions[9,10].

Figure 2.
Alignment of orthologous miR156 subfamilies from Arabidopsis and horticultural plants. A total of 138 miR156 mature sequences from 16 different horticultural plants were aligned. The sequences boxed in black are miR156 seed sequences. ath: Arabidopsis thaliana; mdm: Malus domestica; csi: Citrus sinensis; cme: Cucumis melo; cpa: Carica papaya; vun: Vigna unguiculata; aof: Asparagus officinalis; han: Helianthus annuus; vca: Vriesea carinata; ppe: Prunus persica; vvi: Vitis vinifera; fve: Fragaria vesca; sly: Solanum lycopersicum; bna: Brassica napus; stu: Solanum tuberosum; dpr: Digitalis purpurea; aqc: Aquilegia caerulea. The miR156 sequences were downloaded from the miRbase database. Sequences are listed in Supplemental Table S1.
Despite the conservation of mature miR156, its precursors vary notably in the number and length of members in different horticultural plants and show more significant discrepancies in sequences. Compared with the mature sequences of miR156, the phylogenetic analysis of its precursor sequences can more clearly reflect the evolutionary history of miR156 (Fig. 3). The gene family varies significantly in size and genomic organization in different plant species, which may cause dosage effects and spatial and temporal differences in target gene regulations[11]. Moreover, despite producing the same mature sequences, research has suggested that different MICRORNAs (MIRNAs) play certain functional roles[12,13]. Studies of the MIR156 family in horticultural plants have primarily concentrated on MIR156A, and it is yet unclear how MIR156s function synergistically or redundantly to regulate plant development.
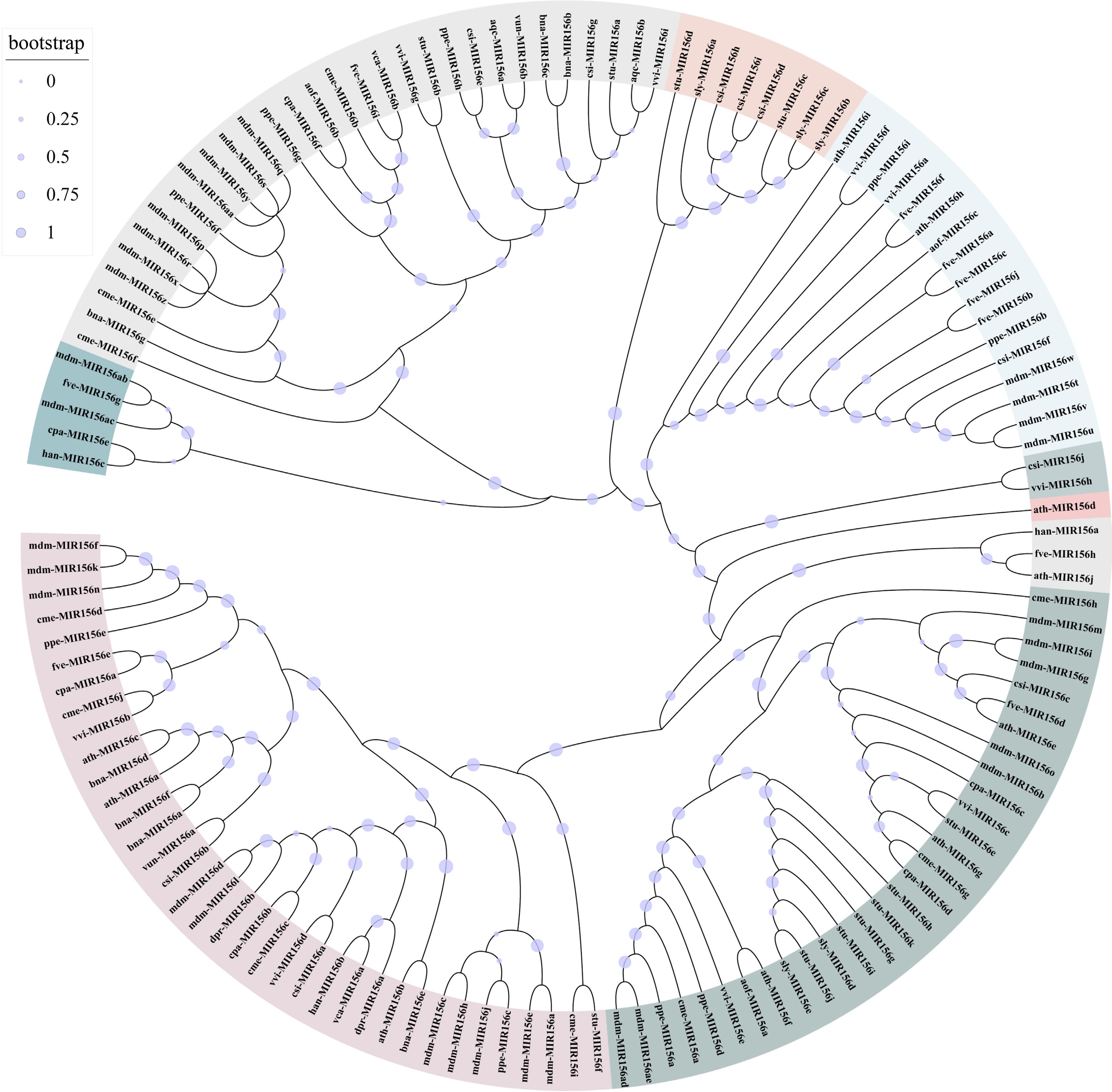
Figure 3.
Phylogenetic relationships between MIR156 from different plants. A total of 122 MIR156 sequences were phylogenetically analyzed. ath: Arabidopsis thaliana; mdm: Malus domestica; csi: Citrus sinensis; cme: Cucumis melo; cpa: Carica papaya; vun: Vigna unguiculata; aof: Asparagus officinalis; han: Helianthus annuus; vca: Vriesea carinata; ppe: Prunus persica; vvi: Vitis vinifera; fve: Fragaria vesca; sly: Solanum lycopersicum; bna: Brassica napus; stu: Solanum tuberosum; dpr: Digitalis purpurea; aqc: Aquilegia caerulea. The MIR156 sequences were downloaded from the miRbase database. Sequences are listed in Supplemental Table S2.
Targets of miR156
-
The targets of miR156 belong to SPL transcription factors containing a highly conserved DNA binding domain (SQUAMOSA promoter binding protein (SBP) domain) with two separate zinc binding sites, while they bear limited conservation outside the SBP domain[5]. SPL genes were first identified in Antirrhinum majus, where the closely related SBP proteins regulate flower development by binding to the promoter of the MADS-box gene SQUAMOSA (SQUA)[14]. SPL transcription factors, as a class of plant-specific proteins, have been widely identified in horticultural plants thereafter. In the genomes of apple, grape (Vitis vinifera), tomato (Solanum lycopersicum), pear (Pyrus pyrifolia), and petunia (Petunia hybrida), 33, 18, 17, 9, 19, and 21 members, respectively, of the SPL gene family were identified. Among them, 20, 12, 7, 7 and 14 SPLs contained miR156 binding sites. Notably, the existence of binding sites does not necessarily imply a negative regulatory role of miR156, as the studies on cucumber suggested[15,16]. The SPL genes targeted by miR156 can be divided into two major groups, represented by SPL3 (SPL3, SPL4, and SPL5) and SPL9 (SPL2, SPL6, SPL9, SPL10, SPL11, SPL13, SPL13-like, and SPL15)[17], which generally serve different roles in plant development[18]. To date, studies have revealed that the miR156-SPL module could engage in multiple important biological processes in horticultural plants, including but not limited to vegetative phase change[19], flower development[20], fruit ripening[21] and stress responses[9]. Functional conservation of the miR156-SPL module has been found in various horticultural plants, while many new functions are also progressively being discovered.
-
Plants go through a vegetative phase and a reproductive phase in their post-embryonic development, and the vegetative phase could be further divided into a juvenile phase and an adult phase[5]. The transition from the juvenile to the adult stage is defined by the acquisition of reproductive competence, while the transition from the vegetative to the reproductive stage is marked by the production of novel reproductive structures, such as flowers or cones[22]. miR156 is a key intermediary connecting plant age and development. This is demonstrated by the fact that the constitutive expression of miR156 prolonged the expression of juvenile vegetative traits and delayed flowering[23, 24]. The expression of miR156 is typically highest in the juvenile stage and declines as plants reach the adult stage. This has been observed in a variety of horticultural plants, such as apple[19], mango (Mangifera indica)[24], mulberry (Morus atropurpurea)[22], cucumber[13], tree peony (Paeonia delavayi)[26], orchid (Dendrobium catenatum)[27] and lily (Lilium × formolongi)[28]. Notably, unlike Arabidopsis, whose life cycle can be completed in less than three months, many horticultural plants, such as lilies, have a protracted juvenile stage that can last for years. In this regard, the miR156-mediated age pathway is a promising tool for horticultural plants with a prolonged juvenile period, as it provides the possibility of accelerating the plant vegetative growth process via a shortened juvenile stage. Similarly, the juvenile stage and the vegetative phase transition are closely related to biomass accumulation in horticultural plants. Overexpression of miR156 delayed the heading time of Chinese cabbage (Brassica rapa), concomitant with prolongation of the seedling and rosette stages; while the overexpression of SPL9 caused significant heading earliness, which could lead to high yield in Chinese cabbage[29]. Thus, manipulating the vegetative phase transition through an age-dependent pathway is a potential approach to improve the productivity of horticultural plants.
Plants show morphological differences under the regulation of the age pathway[30]. The term heteroblasty refers to the phenomenon in which the same plant exhibits abrupt morphological variations during the transition from the juvenile to the adult stage[31]. The term is also increasingly used to encompass the gradual transitions in a variety of traits of most flowering plant species, particularly leaf size and shape, as well as internode length and trichome distribution[32]. miR156 is both necessary and sufficient for the expression of juvenile traits, while an increase in the level of miR156-targeted SPLs is required to trigger heteroblasty[30] by promoting cell growth anisotropy[33]. Many horticultural plants exhibit heteroblastic traits[26,34]. During the seedling stage of cucumber, short and nonfunctional tendrils are formed, while elongated and functional tendrils are formed in the climbing stage. The production of functional tendrils is reported to be age-dependent and regulated by miR156[16]. Passiflora edulis undergoes remarkable changes in leaf morphology with a decrease in the level of miR156[35]. The expression of miR156 and its targets could also be linked to glandular trichome development, which provides resistance to insect pests in tomato[36]. Therefore, these heteroblastic traits deserve to be focused on as they are closely linked to the agronomic performance of horticultural plants.
Flowering time and flower pattern
-
The age pathway is one of the main flowering pathways, together with the photoperiod pathway, the vernalization pathway, the GA pathway, and the autonomous pathway[37,38]. Through the differential expression pattern of the miR156-SPL module, the age pathway regulates flowering time in two ways. One is to suppress the activity of flowering repressors derived from the APETALA2-like (AP2-like) family through microRNA172 (miR172), and the other is to directly induce the expression of floral integrators, such as SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 (SOC1), to induce flowering (Fig. 4)[37,39,40].
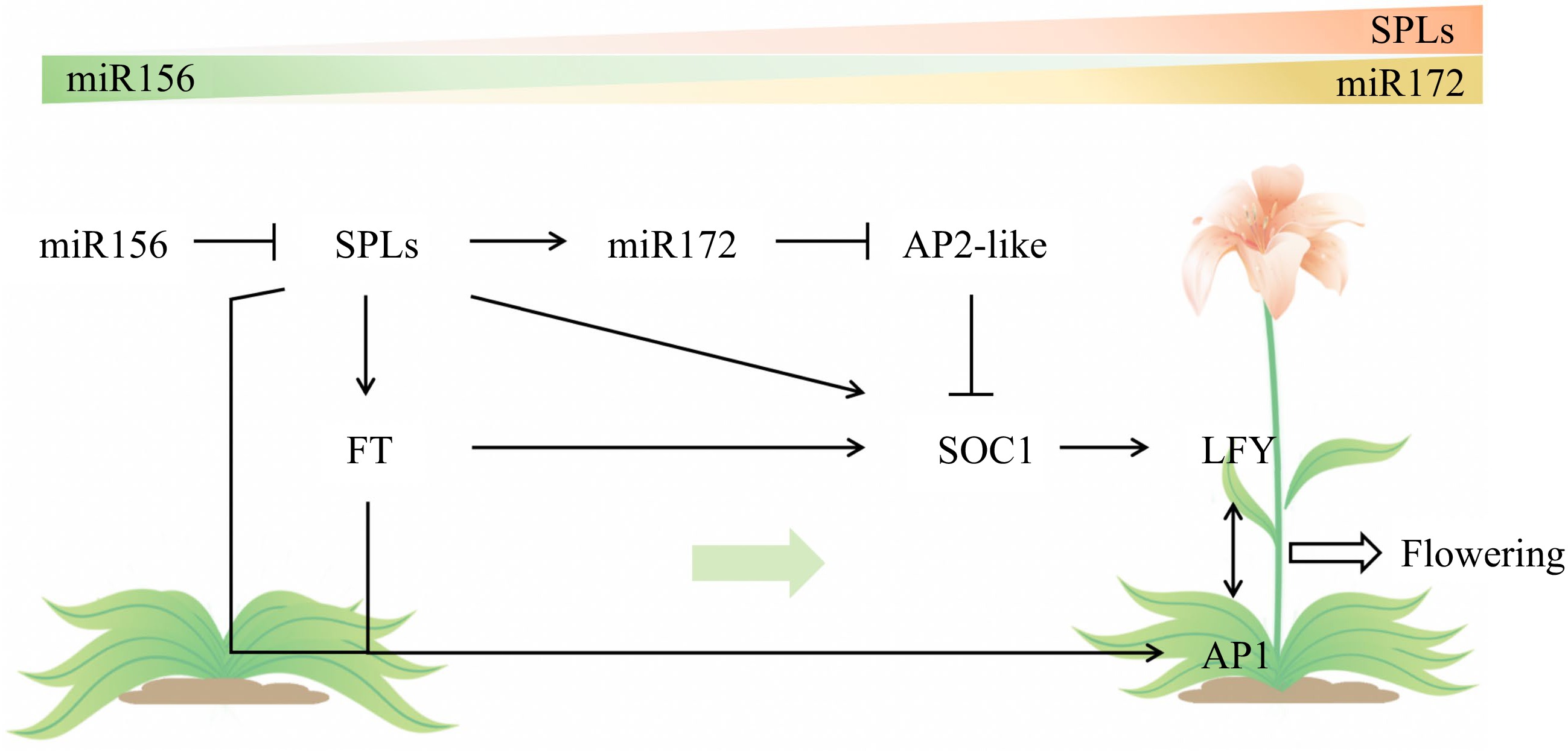
Figure 4.
Age pathway for flowering defined by the miR156-SPL module.
The constitutive expression of miR156 tends to delay flowering, while the miR156-targeted SPLs redundantly promote flowering[23,39,41−43]. miR172, a target of SPLs, acts downstream of miR156 in an antagonistic expression pattern. In contrast to miR156, the expression of miR172 is low during the juvenile phase and exhibits a subsequent increase toward progression to flowering[7,23,30,44]. Early or late flowering of ornamental gloxinia (Sinningia speciosa) was observed in transgenic plants in which miR172 was overexpressed or suppressed, respectively, accompanied by corresponding changes in AP2-like[45]. In addition, several floral integrators work directly downstream of SPL, including SOC1, FLOWERING LOCUS T (FT), LEAFY (LFY) and APETALA1 (AP1)[39,46,47]. SPL transcription factors promote flowering in tomato by positively regulating the expression of the tomato inflorescence-associated gene SINGLE FLOWER TRUSS (SFT) in leaves and the MADS-Box gene AP1/MC in the shoot apex[20,48]. Research on litchi (Litchi chinensis) plants also suggests that SPL transcription factors promote age-dependent flowering by directly binding to the FT promoter[49]. The above findings indicate an effective strategy for regulating flowering time in horticultural plants through the age pathway.
Notably, the miR156-mediated age pathway may define an endogenous mechanism that ensures plants flower in the absence of exogenous inductive cues[39]. This opens the possibility for ornamental flowers to bloom regardless of environmental restrictions. Furthermore, a study on chrysanthemum shows that the expression of the nuclear factor gene CmNF-YB8 is regulated by age, but not by day length, low temperature, or GA, and CmNF-YB8 is shown to bind to the promoter of the cmo-MIR156 and influence flowering time through directly regulating the expression of cmo-MIR156 in the age pathway[42]. The unveiling of the specific upstream transcription effectors of miR156 reveals the significance and independence of the age pathway in flower formation.
miR156 also plays a role in petal coloration and floral pattern formation[50,51]. Increased miR156 expression levels promote accumulation of anthocyanins, whereas the target SPL9 negatively regulates anthocyanin accumulation by directly inhibiting the expression of anthocyanin biosynthetic genes through destabilization of a MYB-bHLH-WD40 (MBW) transcription complex[52]. Similarly, miR156-3p was reported to be involved in yellow flower formation by improving anthocyanin accumulation in herbaceous peony[10,53]. Furthermore, the miR156-SPL module is associated with petal spot rearrangement in Gorteria diffusa, resulting in better mimicry of female flies resting on the flower[54]. Notably, analysis of differentially expressed genes also reveals the potential multifunctional role of miR156 in the development of sterile and fertile flowers[55], double flower forms[51] and multi-tepal patterning[56] in various ornamental plants, which suggests a regulatory node contributing to the diversity of floral morphogenesis.
Plant yield and quality
-
Ample evidence shows the significant role of miR156-SPL in fruit development[24,57,58]. miR156 transgenic tomato exhibited abnormal fruit morphology with extra carpels and ectopic structures as well as small fruit size and reduced fruit numbers[59,60]. miR156 was also found to be an adverse regulator of tuberization in potato[61]. A decrease in miR172 may partially account for this as it was reported to contribute to tuberization[62]. In addition, some evidence has shown that miR156 is instrumental in secondary metabolism, which is closely associated with the quality and nutritional value of horticultural products. As it does in flower petals, the miR156-SPL module serves as an important regulator by participating in anthocyanin biosynthesis in peel coloration during fruit ripening. SPLs in Chinese sand pear interact with MYB10, presumably disrupting the formation of the MBW protein complex[63]. Similar results were also found in horticultural plants such as grape[64,65], litchi[66] and peach[67]. Apart from the involvement in anthocyanin accumulation, miR156 is also engaged in catechin accumulation in tea plants[68,69]. Furthermore, the synthesis of sesquiterpenes, important floral volatiles, has also been reported to be age-regulated. Specifically, the miR156-targeted SPL promotes accumulation of sesquiterpenes in elder plants by upregulating the expression of the patchoulol synthase (PatPTS) gene[70]. These findings provide support for miR156-SPL as a feasible approach to enhance yield and product quality in horticultural plants.
Resistance to biotic stress
-
During the development process, plants are constantly exposed to a variety of biotic stresses, including pest infestation and pathogen invasion. The plant vigor hypothesis suggests greater herbivore abundance on young and vigorous plants[71] and that older plants may display increased resistance to pathogens[72]. Such studies show a tradeoff between plant defense ability and age. This has been partly explained by a study on the age-regulated dynamics of plant insect resistance: the miR156-SPL9 module is responsible for the age-dependent decay of phytohormone jasmonate (JA), a key regulator of plant insect defense, whereas constitutive accumulation of defense compounds such as glucosinolates plays a role in compensating for JA-response attenuation during plant maturation[73].
Research shows that miR156 is involved in the defense against invading pathogens in horticultural plants. High-throughput sequencing shows that miR156 functions in the defense against apple stem pitting virus in pear likely through the regulation of its target gene pbRPS6[74]. In a study on the response to Botrytis cinerea in tomato, bioinformatics analysis on JA-deficient mutants and wild type plants revealed that miR156e-3p is likely involved in pathogen resistance through the JA pathway[75]. Several studies demonstrate the role of miR156 in the resistance to anthracnose disease, which is caused by Colletotrichum gloeosporioides. The expression of miR156 was significantly altered in tea plants after infection[76,77]. Additionally, research on walnut (Juglans regia) revealed that miR156 enhanced plant resistance to anthracnose likely through the regulation of WRKY[78]. Furthermore, miR156s were found to move across the kingdom boundary to the invading C. gloeosporioides cells in cassava, showing a distinct cross-species defense mechanism against the fungal pathogen[79]. Clearly, miR156 is involved in diverse pathogen invasion responses to ensure successful growth and development of horticultural plants.
The miR156-SPL module is also engaged in the regulation of insect resistance[73,80,81], yet it has been reported only sparingly in horticultural plants. A very specific defense mechanism against herbivores exists in swollen thorn acacias (genus Vachellia). In exchange for protection, acacias provide ants with refuge and food in the form of swollen stipular spines, protein-lipid-rich Beltian bodies, and sugar-secreting extrafloral nectaries ('swollen thorn syndrome'). These defense traits are reported to be an age-dependent development correlated with the miR156-SPL pathway rather than a passive developmental pattern arising from developmental constraints[82]. These findings, together with the research on the development of glandular trichome[36] and age-regulated jasmonate response decay and defense metabolite accumulation[73], unveiled the complex mechanisms of miR156-regulated insect resistance during vegetative development (Fig. 5).
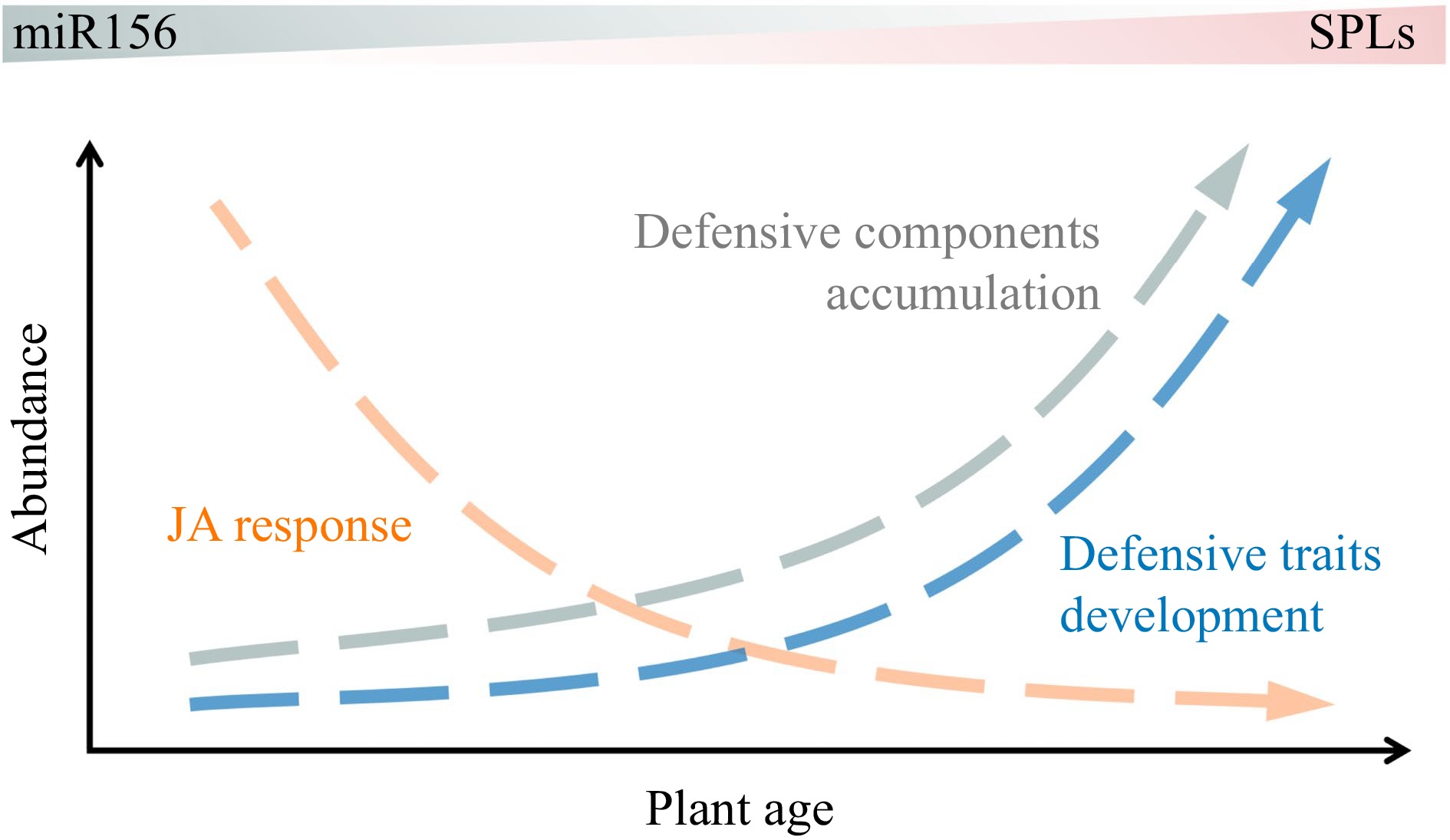
Figure 5.
Age-dependent change in plant resistance to biotic stress.
Resistance to abiotic stress
-
Abiotic stresses strongly affect the growth of horticultural plants. When plants encounter adversity, their development is likely to be delayed. miR156s were reported to play an important role in environmental adaption. This is demonstrated by the fact that miR156 showed a larger differential expression in sugarcane (Saccharum officinarum) under cold, salt and drought stress treatments compared with other implicated miRNAs[83]. However, the underlying mechanism by which miR156 regulates different abiotic stress responses is still unclear.
The expression level of miR156 is dramatically altered under different abiotic stresses in many horticultural plants. For instance, temperature stress resulted in high expression of miR156 in sugarcane[83], banana[84] and cabbage[85]. In addition to this, ample evidence demonstrated the role of miR156 in drought stress. Drought-induced accumulation of strigolactones in tomato leaves leads to the expression of miR156, which may in turn increase guard cell sensitivity to ABA, resulting in stomatal closure. Moreover, the after-effect of drought, by which stomata do not completely re-open after rewatering, was enhanced by miR156[86]. Additionally, the expression of miR156 was much higher in drought-tolerant apple plants than in drought-sensitive plants under control or drought conditions[89]. Notably, the functional role of miR156 against abiotic stresses in horticultural plants has generally not been experimentally verified. Therefore, performing functional validation and further exploring the miR156-related defense mechanism are crucial in future studies.
Generally, expression of miR156 is induced under various abiotic stress conditions to improve stress tolerance[88,89], whereas it is suppressed to accelerate the developmental transition under favorable conditions[90]. This is achieved in part by influencing the anthocyanin biosynthesis through the regulation of the downstream genes DIHYDROFLAVONOL-4-REDUCTASE (DFR) and WD40[90−92]. However, some studies on horticultural plants have reported exceptions. Drought treatment did not regulate miR156 in apple[87,93]. Moreover, expression of miR156 led to weakened salt tolerance in apple, and the overexpression of the miR156-targeted gene MdSPL13 strengthened salt tolerance by activating the MdWRKY100 promoter[93]. These adverse roles of miR156 might be partially explained by the differences in the life cycles of woody and herbaceous plants[93]. The findings above suggest that the multiple functional roles of miR156 in stress response in diverse species is worth exploring in horticultural plants.
Other functions
-
Recent studies have demonstrated a number of novel functions of miR156. Among them, influenced by DELAY OF GERMINATION1 (DOG1), miR156 and miR172 were involved in the regulation of seed dormancy in lettuce (Lactuca sativa). This has been verified in Arabidopsis, in which higher miR156 expression levels enhanced seed dormancy and delayed flowering while overexpression of miR172 exerted the opposite effect[94]. According to the results, a novel linkage, DOG1-miR156-miR172 interaction, has been revealed between dormancy release and flowering, critical developmental transitions in plant life cycles, and it has subsequently been studied in a variety of plants[95,96]. These studies open the possibility for new pathways in dormancy research in horticultural plants, and study of the integrated mechanism for coordinating these two life cycle transitions would be of great significance to optimize both.
miR156 also comes into play during various stages of somatic embryogenesis, including embryogenic callus differentiation and cotyledon and globular embryo development[97]. Overexpression of csi-miR156a significantly enhanced the capability of somatic embryos in preserved citrus embryogenic callus, with more abundant amyloplasts accumulating in the embryonic cells, leading to a significant increase in starch content[98,99]. In contrast, suppression of csi-miR156a reduced somatic embryo capability and the number of amyloplasts and starch content through the downregulation of the expression of both csi-miR172d and some starch biosynthesis genes[100]. These studies suggest a positive role of miR156 in somatic embryogenesis and provide new insights into enhancing somatic embryo capability in horticultural plants.
miR156 is broadly involved in the regulation of many biological processes in horticultural plants to a remarkable extent (Table 1) and is recognized to be a regulatory hub toward various horticultural traits. Notably, rather than operating independently, miR156-SPL functions in collaboration with other pathways. Hence, cross-talk between age and other signaling pathways is discussed in the next section. Furthermore, studies on miR156 have focused on age-regulated plant growth and development, while the involvement of miR156 in other biological functions such as stress response have been addressed, but mostly not functionally verified. Therefore, in the final section, we summarize the methods used for functional validation of miR156 with a view to providing useful information for the in-depth study of the function of miR156.
Table 1. Biological functions of miR156 in horticultural plants.
Classification Species Growth and development Stress response Others Ref. Vegetables Solanum tuberosum Tuberization; development of leaves, trichomes, branches, lateral roots and inflorescences Cytokinin and strigolactone level [61, 101] Solanum lycopersicum Development of leaves, branches and trichomes; floral induction and flower development; ovary and fruit development Drought, cold; Botrytis cinerea infection [9, 20, 36, 48 ,
59, 60, 75, 86]Brassica campestris Vegetative phase change; heading Heat [29, 85] Cucumis sativus Tendril formation; vegetative phase change [15,16] Brassica alboglabra Seed development [102] Brassica oleracea Xanthomonas campestris infection [103] Lactuca sativa Seed dormancy [94] Fruits Malus spp. Vegetative phase change; adventitious root formation Salt, drought Browning inhibition [19, 92, 93,
104, 105]Citrus sinensis Vegetative phase change Somatic embryogenesis [99, 100] Vitis vinifera Vegetative phase change; flower development in winter; grape berry development and ripening [65, 106, 107] Citrus × paradise Vegetative phase change [108] Morus atropurpurea Vegetative phase change Silkworm herbivory [25, 81] Passiflora edulis Vegetative phase change [48] Saccharum officinarum Cold, drought and salt [83] Musa acuminata Fruit ripening Heat [58, 84] Pyrus spp. Red peel coloration Apple stem pitting virus infection Anthocyanin biosynthesis [63, 74] Vaccinium corymbosum Fruit coloration Anthocyanin biosynthesis and chlorophyll degradation [64] Litchi chinensis Flowering Anthocyanin biosynthesis [49, 66] Ornamental plants Paeonia lactiflora Flower coloration Anthocyanin biosynthesis [10, 53] Paeonia delavayi Vegetative phase change [26] Chrysanthemum morifolium Flowering time [42] Rosa chinensis Vegetative phase change; flowering time [109, 110] Vachellia cornigera Herbivore defense [82] Paulownia fortunei Paulownia witches' broom (PaWB) infection [111] Pogostemon cablin Sesquiterpene biosynthesis [70] Gorteria diffusa Petal spot placement [54] Lilium Oriental Trumpet Vegetative phase change [28] Viburnum macrocephalum Differentiation of fertile and sterile flowers [55] Cymbidium goeringii Reproductive organ development [56] Dendrobium catenatum Vegetative phase change [27] Petunia spp. Development of branches and internodes; flowering time [43] Tea plants Camellia sinensis Double flower domestication Colletotrichum gloeosporioides infection Taste compound (catechin, caffeine, and theanine) biosynthesis [51, 68,
69, 76, 77] -
Despite showing great functional diversity, the miR156-SPL module does not work in isolation, but rather in concert with other signaling pathways. The juvenile-to-adult phase transition triggered by miR156 is reported to occur following an increase in sugar[112,113]. The glucose-induced repression of miR156 is thought to be conserved in many plants[112] and is partly dependent on the signaling activity of HEXOKINASE1 (HXK1)[113], which, according to a recent study, is required for CURLY LEAF (CLF)- and SWINGER (SWN)-mediated histone H3 lysine 27 (H3K27me3) deposition and glucose-mediated gene repression[114]. Previous studies suggest that CLF and SWN are responsible for the increased level of H3K27me3 at MIR156A, which thereby leads to the repressed expression of miR156[115,116]. The above studies indicate that glucose signaling likely regulates miR156 expression through epigenetic modification. In addition, several studies demonstrate that the trehalose 6-phosphate (T6P) pathway acts downstream of sucrose to impact vegetative phase transition. To be specific, T6P, a disaccharide signaling molecule that conveys sucrose status to downstream signaling pathways, is able to regulate the expression level of SPL3/4/5 through miR156[117]. Additionally, the T6P pathway strongly suppresses the miR156-SPL-mediated age pathway to promote the vegetative phase change[118]. These findings demonstrate that the T6P pathway is identified as signals associated with age to control the miR156/SPL module. Recent research has reported the effect of sucrose treatment on lily juvenile vegetative development. Exogenous sucrose treatment enhanced the growth of seedlings, especially lily bulbs, with a significant decline in miR156 expression and an increasing level of trehalose-6-phosphate synthase (TPS)-coding genes and LfTPSs[119]. Bulb flowers, typified by lilies, generally encounter the problem of prolonged breeding cycles due to long juvenile stages and bulb size (and likely carbohydrate accumulation), and miR156 is shown to be pivotal for vegetative phase change in bulb flowers[120]. Uncovering the molecular link of sugar signaling with the miR156-mediated age pathway in bulb flowers will certainly help to address the problem of long juvenile stage in bulb plants.
Strigolactones (SLs), a newly identified class of phytohormones, have also been shown to regulate biological process in horticultural plants in coordination with miR156. A typical example is that exogenous SLs are sufficient for miR156 accumulation in tomato leaves and endogenous SLs act as a molecular link between drought stress and miR156 in tomato[86]. Moreover, several studies have reported the integration of SL signaling and the miR156/SPL module in the regulation of plant architecture[121,122].
The age-dependent pathway also interacts with other flowering pathways at multiple levels to ensure the optimal timing of flowering[123]. The molecular mechanism by which the age pathway is integrated into other flowering time pathways, including the autonomous, GA, photoperiod, and vernalization pathways, has been summarized by a previous review[124]. For instance, TAF15b, an autonomous pathway protein, is revealed to interact with miR156-targeted SPL1 and both act synergistically to promote early flowering of rose (Rosa chinensis)[110]. Notably, the mode of integration between age and other pathways may vary in different species[124].
-
The techniques for functional validation of miR156 include overexpression and expression suppression. Specifically, overexpression of miR156 is achieved by overexpressing miR156 precursors, and expression suppression could be achieved by two technologies, namely target mimicry (TM) and short tandem target mimicry (STTM). An endogenous long noncoding RNA (lncRNA), Induced by Phosphate Starvation1 (IPS1), was shown to bind to miR399 and cause the abolishment of the cleavage effect of miR399 on IPS1. This mechanism of miRNA activity inhibition was termed 'target mimicry'[125]. Artificial target mimics have been shown to be similarly functional in crippling other corresponding miRNAs including miR156[30]. However, IPS1-based TM was also reported to be responsible for the incomplete inactivation of miRNA and work effectively only on a small set of miRNAs[126]. Hence, STTM, which harbors two copies of partially complementary miRNA sequences linked by a short spacer, was developed and demonstrated to be more effective in reducing the levels of targeted miRNAs[127]. As a powerful approach to interrogate the function of miRNAs, STTM has been successfully applied in various horticultural plants to validate the functional role of miR156[92,100]. Despite its effectiveness, STTM still has limitations, exemplified by the fact that STTM targets mature miRNAs and therefore fails to distinguish the functional diversity of each MIRNA gene[40]. The precise editing of plant genome sequences has been achieved thanks to the development of CRISPR/Cas9 technology in the past few years and it has been successfully applied to the study of miRNA functions in horticultural plants to create loss-of-function mutagenesis[13,128]. Since it has been recognized that MIRNAs coding for the same mature miRNA may still carry out specific functions[13], the application of CRISPR/Cas9 technology has made it possible to examine the functional role of individual MIR156 genes, which enables the unraveling of a more complex regulatory network of miR156.
-
Due to its highly conserved nature and significant activity in numerous biological processes, miR156 has the potential to serve as a versatile tool to regulate multiple horticultural traits. Since miR156 is a key regulator of the age pathway, how to shorten the juvenile stage of fruit trees and perennial flowers and achieve the enhancement of quality and yield of horticultural plants through the age-regulated pathway remains a priority for future studies. To achieve the biotechnological modification of horticultural traits, in-depth research is required on several specific issues. First, despite the many important studies reviewed herein, the complex regulatory mechanisms of miR156 in horticultural plants remain largely unknown. Horticultural plants often have characteristics such as special organs and developmental stages that are not present in Arabidopsis, such as fruit development in tomato, tuberization in potato, petal spot arrangement in ornamental flowers, tendril formation in cucumber, and heading in Chinese cabbage (Fig. 6). The involvement of miR156 in the development of these special horticultural traits, which are often tightly linked to the economic and ornamental value of horticultural plants, has been reported but has not been fully investigated. It would thus be of great significance to focus on these specific traits for the improvement of horticultural plant quality in the future. Second, due to the diversity of functions performed by miR156, there may be a knock-on effect when it is utilized to improve specific horticultural traits. In this regard, the mechanism by which miR156, as a regulatory hub, manages to balance age with other biological functions is an interesting topic of research. This may be achieved in part through precise regulation of individual genes. Additionally, both SPLs and MIR172s were reported to be functionally redundant, and may fail to lead to phenotypic differences. Therefore, having a thorough grasp of the specific roles played by individual genes in the age pathway as well as those found downstream of it would be helpful to enable precise engineering. Third, as described above, the age pathway defined by miR156-SPL is partly tangled with other signaling pathways. To explore the integration mechanism by which the age pathway works synergistically with other signaling molecules in different plant species is a future challenge.
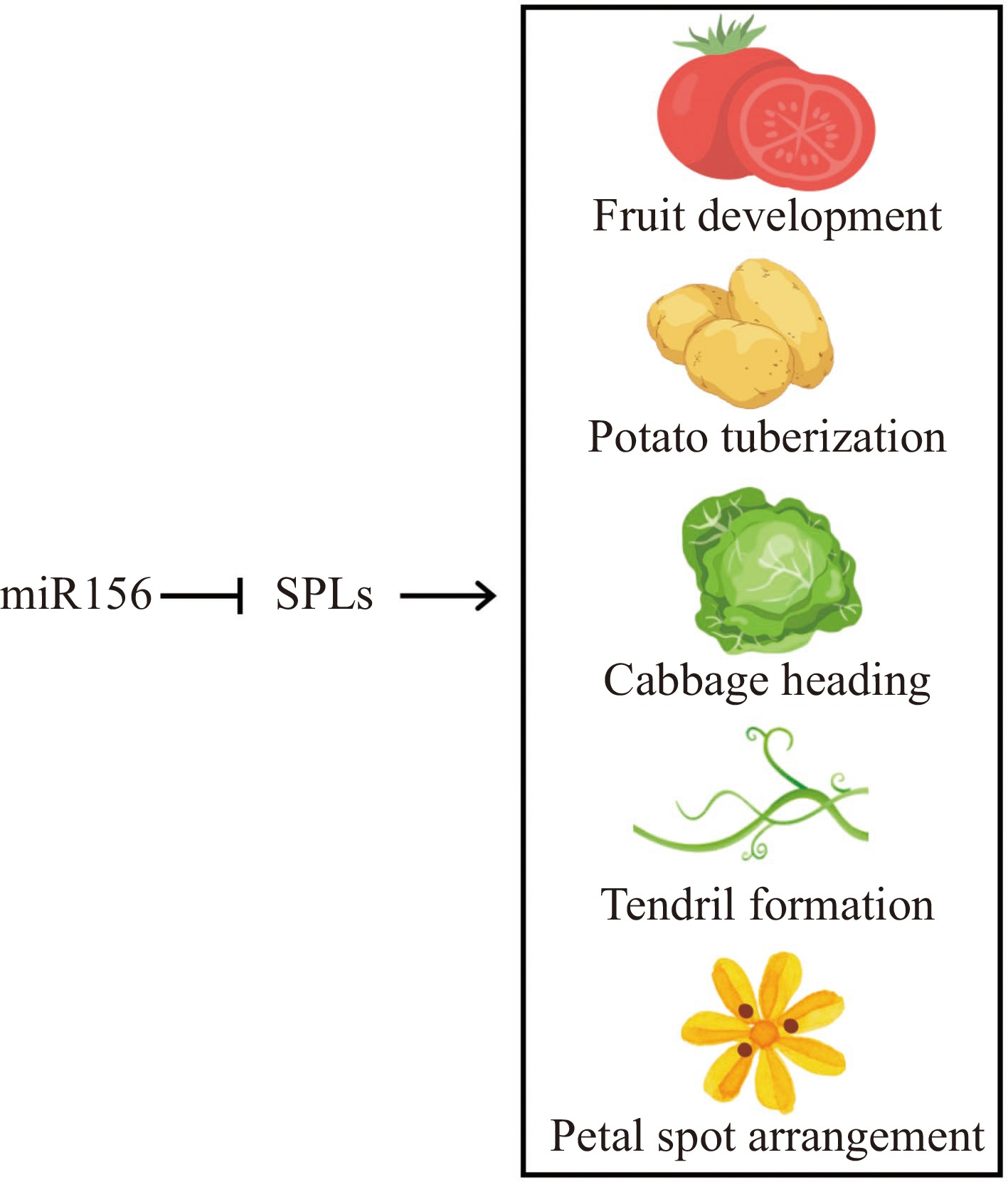
Figure 6.
miR156-SPL module regulates special horticultural traits.
There is considerable variation in the characteristics of different horticultural plants. For example, as mentioned above, fruit trees and certain perennial flowers often undergo a protracted juvenile period, thus it is crucial to speed up the vegetative growth process in order to shorten the breeding cycle and encourage large-scale output. Moreover, given the comparatively short vegetative period of annual vegetables and ornamental plants, extending the vegetative phase may increase production by allowing biomass to accumulate. The coloring of floral and foliar organs, in general, is what confers ornamental plants their desirable characteristics, therefore research into the mechanism of coloration might serve as a foundation for the enhancement of ornamental value and commercial quality. Furthermore, traits such as secondary flowering and petal spots are conducive in increasing the value of ornamental plants further. All of these characteristics are distinct from those of Arabidopsis. Therefore, it is pivotal to understand the functional diversity and regulation mechanism of miR156 to improve the quality and benefits of horticultural plants. In future research, the priority is to unveil the regulatory mechanism of miR156, construct its entire regulatory network, and validate its role in regulating life processes with an emphasis on the special developmental phases and organs in horticulture plants.
-
The authors confirm contribution to the paper as follows: draft manuscript preparation: Xu Y, Gao C; data analysis: Xu Y, Hang Z, Liu J, Ren Z; manuscript revision: Xu Y, Wu Y, Feng S, Xia Y, He Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this article and its supplementary information files.
We thank Dr. Heng Lian (CAS Center for Excellence in Molecular Plant Sciences, China) for the careful revision of this manuscript. This work was funded by National Natural Science Foundation of China (Grant No. 32002071), National Natural Science Foundation of China (Grant No. 32372743), Zhejiang Sci-Tech University Research Program Start-up Funding (Grant No. 21052103-Y), Zhejiang Province first-class discipline (civil engineering) construction project (Grant No. 11141131282001) and 2022 Hangzhou Agricultural and Social Development Scientific Research Guidance Project (Grant No. 20220919Y1).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yunchen Xu, Cong Gao
- Supplemental Table S1 miR156 mature sequences for alignment.
- Supplemental Table S2 MIR156 sequences for phylogenetic analysis.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu Y, Gao C, Huang Z, Liu J, Ren Z, et al. 2024. MicroRNA156: a count up timer with potential to enhance horticultural traits. Ornamental Plant Research 4: e010 doi: 10.48130/opr-0024-0008 

MicroRNA156: a count up timer with potential to enhance horticultural traits
- Received: 31 October 2023
- Revised: 21 January 2024
- Accepted: 18 February 2024
- Published online: 02 April 2024
Abstract: MicroRNA156 (miR156), targeting a subset of SQUAMOSA PROMOTOR BINDING PROTEIN-LIKE (SPL) genes, is one of the most evolutionarily conserved microRNAs (miRNAs) in plants. miR156 is a recognized molecular marker of plant age. In addition to being an age indicator, miR156 has demonstrated strong multifunctionality in the regulation of a broad range of biological processes in horticultural plants, including vegetative growth, floral induction, fruit ripening, stress responses, and secondary metabolism. Due to its versatility and high level of conservation among phylogenetically distinct plant species, miR156 may be a powerful tool for biotechnological modification in horticultural plants. Here, we summarize the functional diversity of miR156 in horticultural plants to provide new insights for further research on the biological function and regulatory mechanism of miR156 and on how to employ miR156 to achieve the enhancement of horticultural traits.
-
Key words:
- miR156 /
- SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) /
- Age /
- Horticultural traits