









-
Clustered regularly interspaced palindromic repeats/CRISPR-associated protein (CRISPR/Cas)-mediated genomic editing is a low-cost, convenient, highly efficient technique that can produce mutants in which target gene functions are completely knocked out. CRISPR/Cas systems, which are widely used in model organisms, comprise three major types based on the nuclease involved. Among these, type II CRISPR/Cas with the Cas9 enzyme derived from Streptococcus pyogenes is the best-developed system[1]. CRISPR/Cas9 has important advantages over other gene-editing technologies: the user only needs to design an RNA sequence corresponding to the target nucleic acid, and the technique is simple, highly efficient, and inexpensive, providing an excellent platform for genome-editing[2]. CRISPR/Cas9 has been successfully applied to Arabidopsis thaliana[3], rice (Oryza sativa)[4,5], wheat (Triticum aestivum)[6], tomato (Solanum lycopersicum)[7], Brassica napu and poplar (Populus L.)[8,9]. However, the efficiency of CRISPR/Cas9-mediated gene-editing systems varies greatly in different experiments[10], and there is often a high miss rate.
Golden Gate Assembly is an efficient method for inserting plasmids into vectors, allowing the assembly of up to nine fragments at a time in a recipient plasmid[11]. This cloning strategy not only makes it easy to construct a single gRNA expression plasmid, but is also suitable for expressing multiple gRNAs. The use of the Golden Gate method makes it easy and efficient to construct multi-target CRISPR/Cas9 vectors[12]. The TCEP method, in which the editing system is tested using transient Cas9/single guide RNA (sgRNA) expression prior to stable transformation, can be used to optimize CRISPR/Cas systems because it allows CRISPR/Cas-mediated gene editing in the absence of a stable transformation system and can be completed within 3–4 d. In theory, this technique can be applied in any plant species that can be transformed by Agrobacterium.
Phytoene Dehydrogenase (PDS), which encodes a key catalytic enzyme in the carotenoid biosynthesis pathway, is often used as a reference gene during CRISPR/Cas9 gene editing in plants[13]. Silencing or deletion of PDS in plants blocks carotenoid biosynthesis and results in the loss of protection of chlorophyll against degradation, causing an easily detectable albino phenotype in tissues and organs that are normally green. For instance, CRISPR/Cas9 gene editing has been used to produce pure white mutants in Arabidopsis thaliana[14], rice[15], poplar[8], tomato[16], banana[17], and apple[18].
Chrysanthemum morifolium is one of the top ten traditional flowers in China and ranks among the top four cut flowers worldwide. Chrysanthemum White Rust (CWR), a worldwide epidemic disease caused by Puccinia horiana. Henn (P. horiana) has been called the 'cancer' of chrysanthemum[19,20]. CWR damages flower quality and results in up to 100% yield loss[21]. Utilizing resistance genes to cultivate resistant varieties is likely the most efficient way to control CWR. P. horiana is an obligate biotrophic fungus, and we previously demonstrated that the application of salicylic acid (SA) increases resistance to CWR[22]. Pthogenesis-related (PR) genes involved in the SA pathway are often regulated by TGA transcription factors (TFs) that enhance plant resistance to biological stress[23]; previous studies have shown that overexpression of MdTGA2.1 GvTGA2 GmTGA8 and GmTGA19 can promote the expression of downstream PRs in different plants and enhance resistance to disease[24,25]. Notably, the transcript levels of CmTGA1 in chrysanthemum leaves are significantly up-regulated upon P. horiana infection[26].
To date, the main methods used for research on gene function in C. morifolium are virus-mediated gene silencing (VIGS) and RNA interference (RNAi). Although these two technologies have many advantages, they cannot achieve complete knockout of a target gene[27]. Therefore, access to CRISPR/Cas9 technology would be a very important asset in efforts to breed disease-resistant chrysanthemum. Here, starting with the C. morifolium resistant cultivar 'C029' as the experimental material, we tested Golden Gate Assembly, as an improved CRISPR/Cas9 multi-target vector system, combined with TCEP to verify the reliability of the gene targeting. We further performed stable genetic transformation and achieved efficient targeted knockout of the endogenous CmPDS gene in C. morifolium, thereby successfully establishing a CRISPR/Cas9 gene-editing system for chrysanthemum. Finally, we tested CmTGA1 as a target to validate the efficiency of our newly designed gene-editing system. Our findings lay the foundation for the use of gene editing in genetic improvement of C. morifolium.
-
C. morifolium resistant cultivar 'C029' was used as the experimental material. The plant materials were provided by the Forestry College of Shenyang Agricultural University, Shenyang, China. Seedlings at the 6- to 8-leaf stage were transferred to a potting soil mixture, placed in a greenhouse, and grown under 16-h light/8-h dark photoperiod for 2 weeks at 25 ± 3 °C. The plant expression vectors pCBC-DT1T2 and pHSE401 were purchased from the MiaoLing Plasmid Sharing Platform (
www.miaolingbio.com ) (Supplemental Fig. S1a & b).Design of the Cas9 target sites and construction of the pHSECRISPR/Cas9-gRNA plant expression vector
-
The online tool CRISPR v2.0 was used to determine the appropriate sequences of the CRISPR/Cas9 target sites based on the C. morifolium gene sequence of CmPDS (Supplemental Table S1): T1, TGCCTGCACCATTGAATGG; T2, ACCTGAGAGACTTTGCAAGCCAA, schematic representation of the target sites is shown in Supplemental Fig. S2a. Primers were designed according to basic design principles for target sites and sgRNA construction (Table 1). Using pCBC-DT1T2 as a template, the sgRNA expression cassette was constructed by overlapping PCR, and the sgRNA expression cassette was connected to pHSE401 by Golden Gate Assembly. The newly constructed CRISPR/Cas9-gRNA recombinant plasmid targeting CmPDS was transferred into Agrobacterium strain GV3101. To verify the editing efficiency of the system, primers were designed based on the target CmTGA1 gene sequence (Supplemental Table S1): T1, AAGCTAACCCTAA-TAGAAG; T2, ATTCGAACAAGCTTTATTC. The CRISPR-CmTGA1 knockout vector was constructed using the same method for verification of gene editing, schematic representation of the target sites of CmTGA1 is shown in Supplemental Fig. S2b.
Table 1. Primers used for mutant identification.
Primer
namePrimer sequence PDST1-BsF ATATATGGTCTCGATTGTGCCTGCACCATTGAATGGGTT PDST1-F0 TGTGCCTGCACCATTGAATGGGTTTTAGAGCTAGAAATAGC PDST2-R0 AACTTGGCTTGCAAAGTCTCTCCAATCTCTTAGTCGACTCTAC PDST2-BsR ATTATTGGTCTCGAAACTTGGCTTGCAAAGTCTCTCCAA TGA1T1-BsF ATATATGGTCTCGATTGAAGCTAACCCTAATAGAAGGTT TGA1T1-F0 TGAAGCTAACCCTAATAGAAGGTTTTAGAGCTAGAAATAGC TGA1T2-R0 AACGAATAAAGCTTGTTCGAATCAATCTCTTAGTCGACTCTAC TGA1T2-BsR ATTATTGGTCTCGAAACGAATAAAGCTTGTTCGAATCAA PDS-TF GTACAGAACCTGTTTGGAGAGTTA PDS-TR CTTCCGGCAAAAGAAGCTTCAGAA TGA1-TF ATGAGCATCCTTGAAACAACATTC TGA1-TR TCATAGTGTTTGAGTTGCCATCAT Transient genetic transformation of C. morifolium with the CRISPR-CmPDS/CmTGA1 vector
-
Agrobacterium tumefaciens cells harboring CRISPR-CmPDS/CmTGA1 were streak inoculated onto LB solid medium containing kanamycin and rifampicin. The plates were inverted, transferred to an incubator at 28 °C, and cultured in the dark for 2 d. A single colony was inoculated into 10 mL LB liquid culture medium containing kanamycin and rifampicin and cultured in the dark with shaking at 200 rpm·min–1 and 28 °C until the Agrobacterium liquid was turbid. One mL sample of the culture was inoculated into 100 mL YEP liquid medium and cultured in an incubator 28 °C with shaking at 200 rpm·min–1 until the concentration reached OD600 = 0.8 – 1.0. Following centrifugation at 5,000 rpm·min–1 for 10 min, the bacteria were resuspended in infection solution and activated at room temperature for 3 h. Untreated wild-type (WT) plants were used as the control, and plants infected with Agrobacterium harboring the CRISPR-CmPDS /CmTGA1 vector were used as the experimental group.
Whole 30-d-old, robust 'C029' in vitro-grown seedlings at the 6- to 8-leaf stage were soaked in Agrobacterium solution as the experimental group. There were three seedlings per group, and a total of three groups were treated. The plants were washed with sterile water until there was no residual Agrobacterium liquid on the surface, plated on MS culture medium, cultured in the dark at 25 °C for 1 d, and placed in a tissue culture room at 25 °C under a 16-h light/8-h dark photoperiod for further culture.
Total RNA was extracted from the leaves of 'C029' plants using an RNA prep Pure Plant Kit and synthesized first-strand cDNA using a Prime Script II 1st Strand cDNA Synthesis Kit following the manufacturer's protocol. These cDNAs were used as templates in designing specific primers based on the CmPDS and CmTGA1 sequences for RT-qPCR detection. The expression of CmPDS and CmTGA1 was measured in control and Agrobacterium-treated plants using C. morifolium acting as an internal reference.
Genetic transformation of chrysanthemum 'C029'
-
Genetic transformation of 'C029' was performed using three rounds of transformation, using 10 leaf discs each time. The leaf discs were pre-cultured on medium (MS + 0.5 mg·L–1 6-BA + 0.3 mg·L–1 NAA) for 2 d. When the concentration of the Agrobacterium solution reached OD600 = 0.5, the solution was diluted 100-fold, and the leaf discs were infected with the diluted Agrobacterium culture for 7 min. The infected leaf discs were transferred to coculture medium (MS + 0.5 mg·L–1 6-BA + 0.3 mg·L–1 NAA) and incubated for 2 d, after which they were incubated in delayed culture medium (MS + 0.5 mg·L–1 6-BA + 0.3 mg·L–1 NAA + 400 mg·L–1 Cef) for 2 d. Finally, the materials were transferred to screening medium (MS + 0.5 mg·L–1 6-BA + 0.3 mg·L–1 NAA + 400 mg·L–1 Cef + 80 mg·L–1Kan), which was replaced every 15 d. After antibiotic-resistant buds formed, they were transferred to rooting medium (MS + 400 mg·L–1 Cef) for culture.
Detection of transgenic C. morifolium seedlings
-
Genomic DNA was extracted from the leaves of control and transgenic C. morifolium seedlings with a kit (New Type Plant DNA Kit of Aidlab). To determine whether foreign DNA was inserted into the C. morifolium genome, specific primers were designed based on the sequence of the pCBC-DT1T2 vector (F: GTTTTAGAGCTA-GAAATAGCAAGT; R: GAATACTCAAGCTATGCAAGGCCC) and used for PCR amplification of a 626-bp fragment. After confirmation was obtained that the foreign DNA fragment was inserted into the C. morifolium genome, PCR primers including the sequences of endogenous CmPDS/CmTGA1 targets were inserted into C. morifolium to determine whether the target genes were mutated. The PCR primers are shown in Table 1. The PCR amplification product was ligated to the T1 vector after gel recovery. Monoclonal colonies were selected for sequencing, and the sequencing results were analyzed.
Quantitative reverse transcription polymerase chain reaction
-
To assess the disease resistance of wild-type plants and transgenic lines after inoculation with P. horiana, three disease-resistance genes (PR1, PR2, and PR5) in the SA pathway were selected for verification by Quantitative Reverse Transcription Polymerase Chain Reaction (RT-PCR). Primers are shown in Table 2. Leaves of the 'C029' WT and CmTGA1-CRISPR/Cas9 lines were harvested at 0 to 72 h after treatment with P. horiana. A PrimeScript™ RT reagent Kit with gDNA Eraser (Perfect Real Time), PrimeScript™ II 1st Strand cDNA Synthesis Kit, and SYBR Premix ExTaq™ II were purchased from Takara. RT-qPCR was performed using an SYBR Premix ExTaq™ II, following the protocols of the manufacturer. The 2−ΔΔCᴛ method was used to calculate the expression levels of these disease-resistance genes. All experiments were performed with three biological replicates.
Table 2. Primers used for RT-PCR.
Primer name Primer sequence PR1-F CCTTAAACCCAATCTCACAATG PR1-R TGAGTGCGTTGGTGGAGTT PR2-F TCGACTGACATAAGGCACATAACGC PR2-R GCTTCCATAACCTCCATCTCCTTCG PR5-F CCAATGGAGTTTAGCCCCGT PR5-R GTCCACAACTACCACGCTCA Actin-F TCCGTTGCCCTGAGGTTCT Actin-R GATTTCCTTGCTCATCCTGTCA -
Using the pCBC-DT1T2 plasmid as the template, a band of approximately 626 bp was obtained by overlapping PCR, which was the expected size (Supplemental Fig. S3a). The PCR product was recovered, and its correct sequence verified. We used the Golden Gate Assembly method to insert this gene into the BsaI site of pHSE401 (Supplemental Fig. S3b). Gel electrophoresis confirmed that the newly constructed plasmid had the correct sequence, indicating that the desired CRISPR-CmPDS knockout vector had been constructed. We then successfully transferred this plasmid, containing a Cas9 and gRNA expression cassette, into Agrobacterium strain GV3101 via the freeze-thaw method (Supplemental Fig. S3c).
Verification of editing efficiency of the CRISPR-CmPDS vector using TCEP
-
To determine the editing efficiency for the target, we performed reverse transcription–quantitative polymerase chain reaction (RT-qPCR) to measure the expression of CmPDS in the leaves of 'C029' WT and experimental group. DNA was isolated from the transgenic plants, and quantitative PCR (qPCR) was performed. Compared to the WT, the expression of CmPDS was significantly reduced in the leaves of the experimental group, with an expression level of approximately 19.1%–52% that of the WT (Fig. 1). These results indicate that the CRISPR-CmPDS vector could be used for efficient gene editing and suggested that the selected knockout target could successfully be used for stable genetic transformation of C. morifolium.
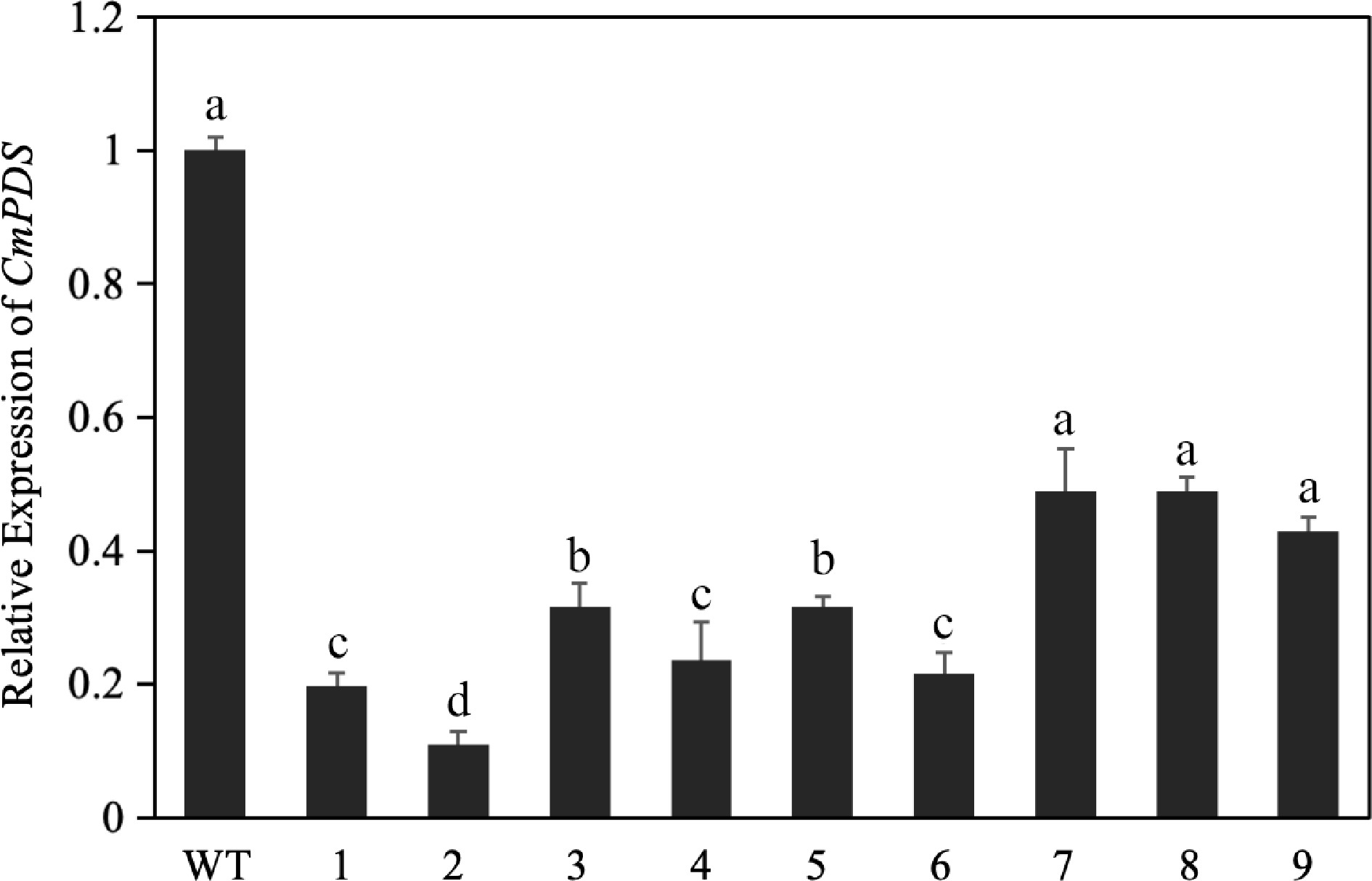
Figure 1.
Relative expression of CmPDS after transient silencing, as determined by RT-qPCR. Different lowercase letters indicate significant differences between different treatments at the same time (p < 0.05).
Obtaining transgenic lines by Agrobacterium mediated stable genetic transformation
-
After verifying the editing efficiency of the target by using the TCEP method, we transfered the recombinant vector, CRISPR-CmPDS, into Agrobacterium competent cells for stable genetic transformation. We obtained 12 antibiotic-resistant buds by using stable transformation.
Identification of CmPDS transgenic C. morifolium plants
-
Then we examined the editing efficiency of the CRISPR-CmPDS vector in 12 antibiotic-resistant buds obtained using stable transformation procedures. We conducted tissue culture and rapid propagation of individual plants of the 12 antibiotic-resistant lines. Among them, eight lines (i.e., 66.7%) showed albino phenotypes, including four totally albino lines (Fig. 2d) and four lines with an interalbino phenotype (Fig. 2e). No albino phenotype was found among the negative control plants (plants infected with Agrobacterium harboring empty vector; Fig. 2f). Plants that were completely albino died shortly after seedling formation, and the plants that survived were all mosaic and grew more slowly than negative control plants. To determine whether foreign genes were integrated into the genome of these plants, we subjected four surviving plants with albino phenotypes, one negative control plant and one 'C029' WT to PCR verification. We extracted genomic DNA from leaves and carried out PCR amplification, using primers targeting the pCBCD-T1T2 vector. We successfully amplified 626-bp bands from the albino lines, but not from the negative control plant and 'C029' WT (Supplemental Fig. S4). Preliminary results showed that the genomes of the antibiotic-resistant seedlings harbored knockout carrier sequences. Therefore, we conclude that the CmPDS knockout vector had been successfully transferred into C. morifolium, and these albino plants were transgenic.

Figure 2.
Process for genetic transformation of C. morifolium. (a) Preculture. (b) Coculture. (c) Screening and cultivation. (d) Albino plant. (e) Interalbino plant. (f) Negative control plant.
We designed specific primers upstream and downstream of the target site CRISPR/Cas9 to detect the editing of CmPDS in the transgenic plants. We used genomic DNA from the four transgenic plants as templates for PCR amplification and sequencing. All four transgenic plants contained edited genes. We identified two types of genome editing, in target sites 1 and 2 of these four plants (see Methods for definitions of target sites). Plant 1 contained a 3-bp substitution at target 1 and a 2-bp deletion and 4-bp substitution at target 2; plant 2 contained a 2-bp substitution at target 1 and a 2-bp deletion and 2-bp substitution at target 2; plant 3 contained a 1-bp substitution and a 3-bp deletion at target 1 and a 3-bp deletion at target 2; and plant 4 contained a 3-bp substitution at target 1 and a 2-bp substitution at target 2 (Fig. 3). All of these mutations cause errors in the translation of the CmPDS protein, resulting in the loss of CmPDS function and an albino appearance. By contrast, the sequences of the CmPDS target sites in the negative control plants were consistent with those of the WT, with no mutations detected.
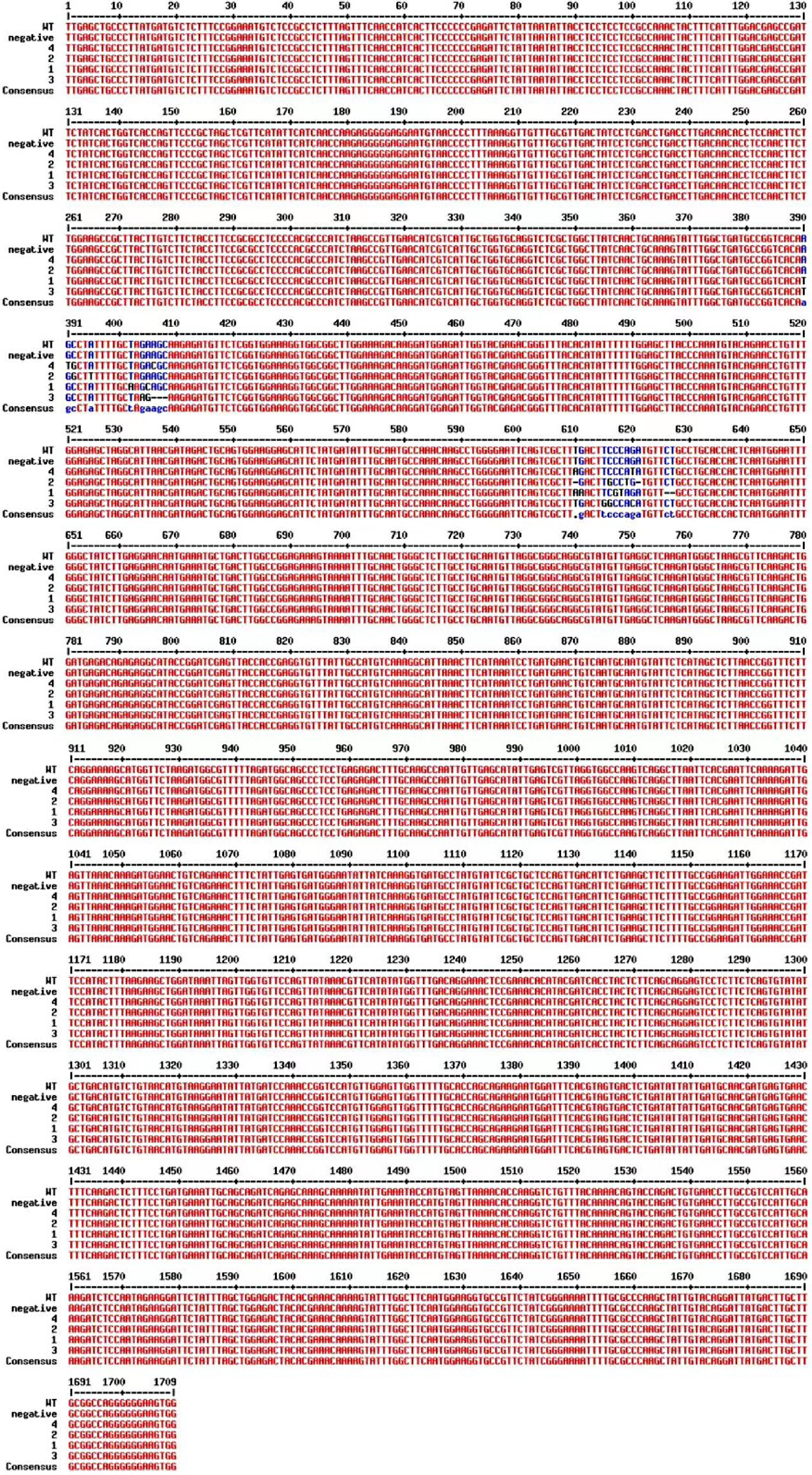
Figure 3.
Sequence alignment of CmPDS from transgenic plants along with the WT and the negative control plants.
Construction of a CRISPR-CmTGA1 knockout vector and analysis of editing efficiency
-
To test the editing efficiency of our system for a different target gene, we generated a CRISPR-CmTGA1 knockout vector and transferred it into Agrobacterium tumefaciens strain GV3101 for transient expression (Supplemental Fig. S5a–c). We performed RT-qPCR to measure the expression level of CmTGA1 in WT and the experimental group. Compared to the WT, CmTGA1 gene expression in the experimental group was significantly lower, approximately 21.1%–70.3% that of the WT (Fig. 4). These results indicate that the CRISPR-CmTGA1 vector successfully edited the target gene and that the selected target was successfully knocked out, meaning that the CRISPR-CmTGA1 knockout vector could be suitable for stable genetic transformation of C. morifolium.
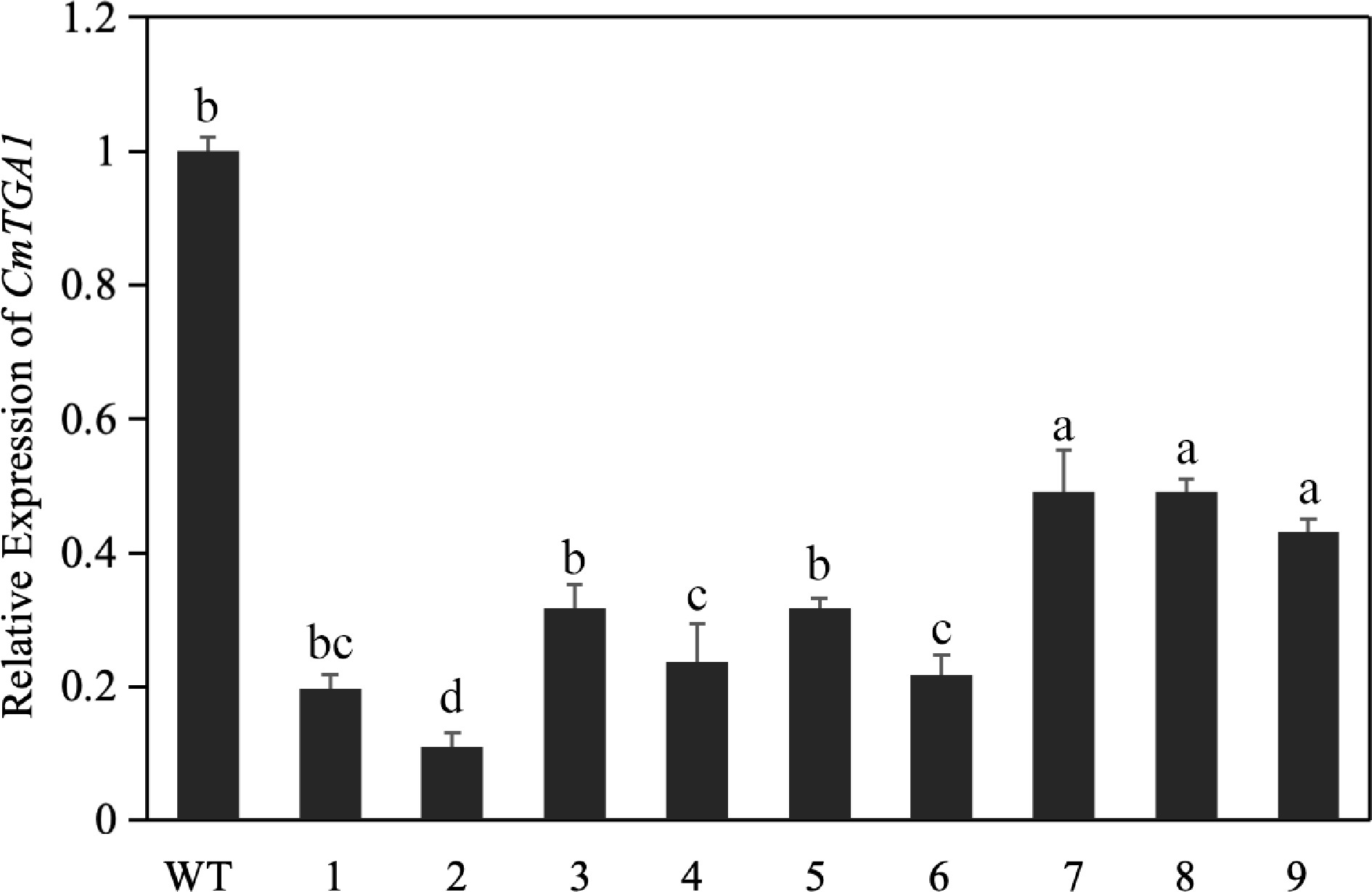
Figure 4.
Relative expression of CmTGA1 after transient silencing, as determined by RT-qPCR. Different lowercase letters indicate significant differences between different treatments at the same time (p < 0.05).
To examine the editing efficiency of the knockout vector in stable transformation, we used primers specific for the pCBCD-T1T2 vector for PCR amplification. As a result, 626-bp bands were amplified from the transgenic plants, indicating that the CRISPR-CmTGA1 knockout vector had been successfully transferred into C. morifolium (Supplemental Fig. S6). We analyzed the CmTGA1 gene sequences of six stably transformed plants, finding that three of them had been successfully gene edited, for a gene-editing frequency of 50% (Fig. 5). In plant 1, target site 1 contained a 4-bp substitution and a 2-bp deletion, whereas target site 2 contained a 2-bp substitution and a 3-bp deletion; in plant 2, target site 1 contained a 4-bp substitution and a 4-bp deletion, whereas target site 2 contained a 5-bp substitution and a 2-bp deletion; in plant 3, target site 1 contained a 3-bp substitution and a 1-bp deletion, whereas target site 2 contained a 2-bp substitution and a 3-bp deletion. These mutations led to the knockout and loss of function of CmTGA1 (Fig. 6). These results validate the efficiency of gene editing in C. morifolium using the double-target genome-editing vector pHSE401.

Figure 5.
Process for genetic transformation of C. morifolium. (a) Preculture. (b) Coculture. (c) Delayed cultivation. (d) Screening and cultivation (the red box highlights a resistant bud). (e) Transgenic plant.
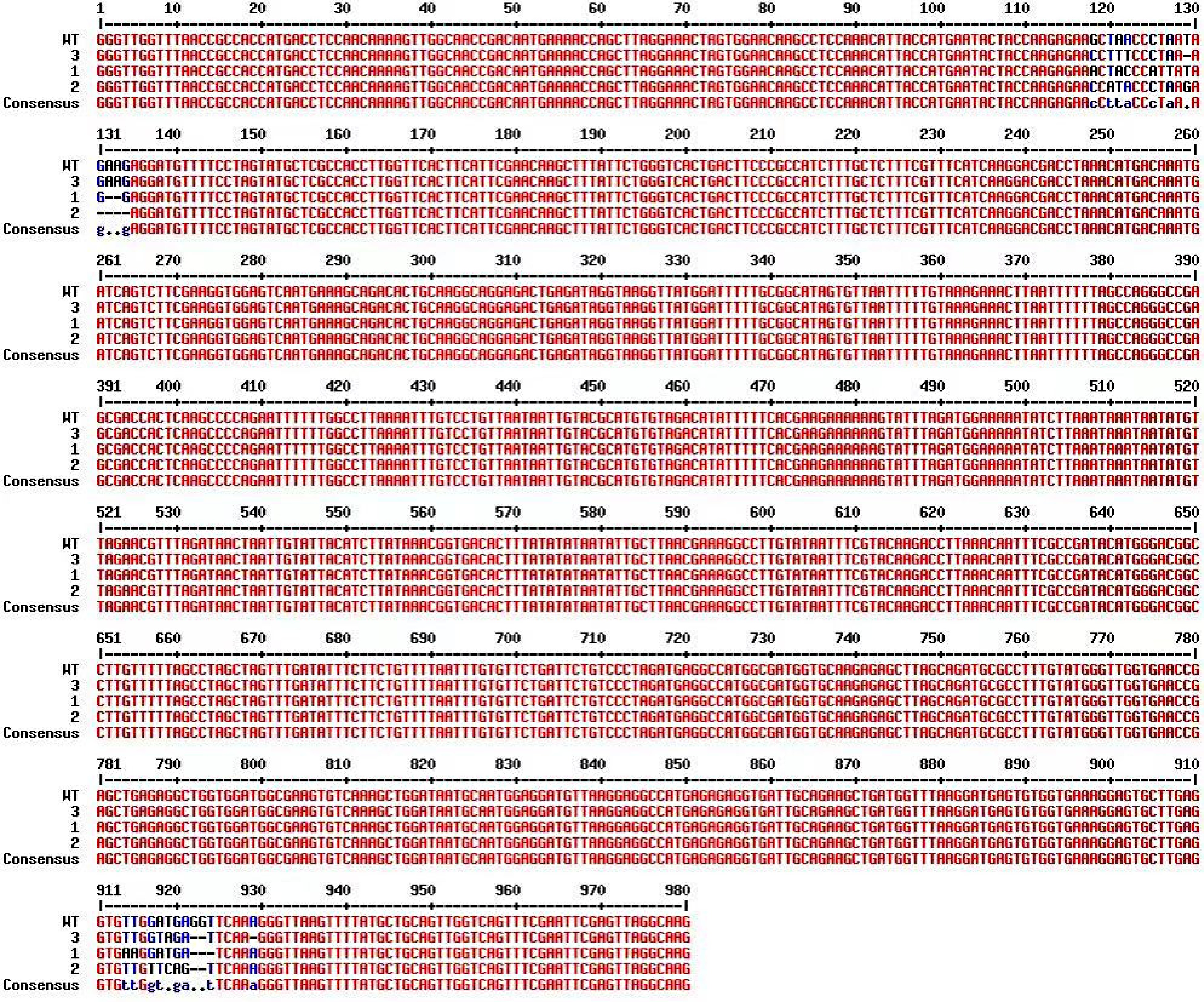
Figure 6.
Sequence alignment of CmTGA1 from transgenic plants compared to the WT.
Knockout of the CmTGA1 reduced resistance to P. horiana
-
We obtained three cmtga1 mutants using our CRISPR/Cas9 platform, and plant 1 was used for the ensuing studies. To explore the disease resistance of 'C029' after CmTGA1 knock out, we measured the expression of PR1, PR2, and PR5 in both WT and cmtga1 mutant by RT-qPCR after inoculation with P. horiana. As shown in Fig. 7, the expression of PRs in cmtga1 mutant were lower than that in WT and showed a trend of increasing first and then decreasing. Although the expression of PRs in WT and cmtga1 mutant reached their maximum value at 48 h, the expression of PRs in cmtga1 mutant were only about 0.11, 0.12 and 0.54 times of those in WT. Overall, our results show that knockout of the CmTGA1 reduced resistance to P. horiana.
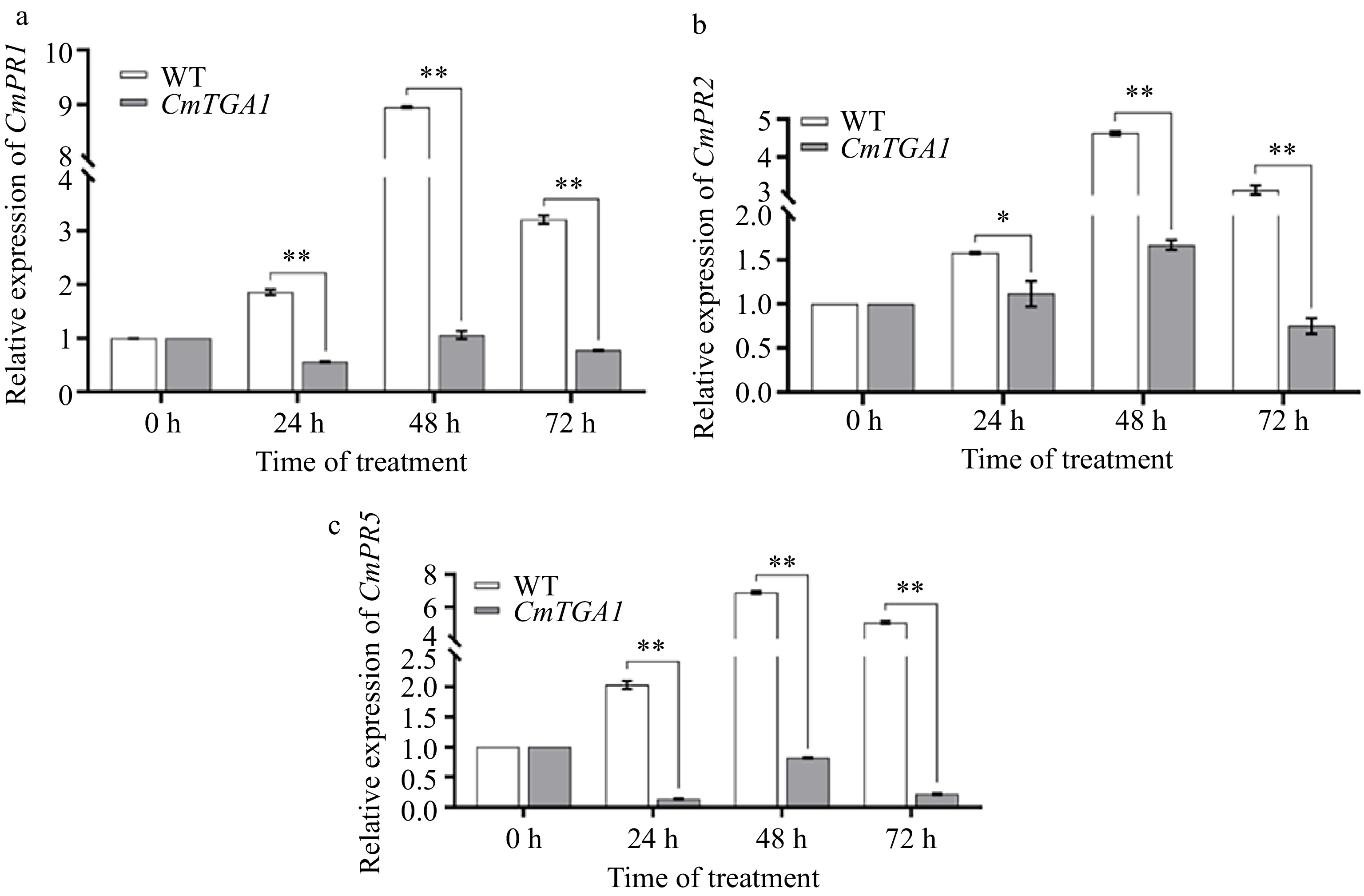
Figure 7.
Expression of CmPR1, CmPR2 and CmPR5 genes in WT and CmTGA1 lines after inoculation with P. horiana. (a) Expression of CmPR1. (b) Expression of CmPR2. (c) Expression of CmPR5. Analysis of significant differences was performed using Student's t-test (**, p < 0.01).
-
The development of CRISPR/Cas9, a powerful gene-editing technology, has greatly advanced for many non-model plants. In Ipomoea, DFR-B was used as the target gene to change the color of the stem[28]. In Lilium, the PDS gene in two species has been knocked out, with completely albino, pale yellow and albino–green chimeric mutants observed[29]. In a previous study, the CRISPR/Cas9 gene-editing technology was used to alter flowering time in Chrysanthemum indicum[30]. However, gene-editing technology has not been reported in the Chrysanthemum cultivar 'C029'. In this study, we chose two optimal target sites in CmPDS according to basic design principles for target sites and sgRNA construction and generated an sgRNA expression box[31]. We connected the sgRNA expression box to the Cas9 vector and successfully constructed the pHSE401-PDS recombinant vector using Golden Gate Assembly. Using a modified CRISPR/Cas9 multi-target vector system, we successfully achieved site-specific knockout of the endogenous C. morifolium CmPDS gene and obtained albino mutant lines.
Although CRISPR/Cas9 technology holds great promise for genome editing, the editing efficiency of CRISPR/Cas9 differs greatly among plant species and is often subject to a high miss rate. This may be caused by the use of different sgRNAs for different target sites, the different binding forces of sgRNAs to target DNA, or differences in chromosome structure, chromosome methylation, or other genetic characteristics[32,33]. Agrobacterium-mediated gene transformation and plant tissue culture can produce gene-edited plants, but induction and regeneration are extremely inefficient and time-consuming, which hinders the application of this technology[34]. We reasoned that identification of target editing efficiency before attempting stable genetic transformation would improve the efficiency of the process. In this study, we developed a CRISPR/Cas9 gene-editing approach in which we combined transient transformation with stable transformation in the commercially important flower C. morifolium. We first verified the feasibility of targeting the intended sites via transient transformation of C. morifolium seedlings, and then we conducted a stable transformation. This combination method not only improves the reliability of the experiment, but also reduces the cost of the experiment.
Analysis of stable genetic transformants showed that the mutations in C. morifolium induced by CRISPR/Cas9 were mainly base deletions or substitutions at the gene target sites, which led to errors in the translation of the target protein and the loss of its function. The mutation rate, as revealed by an albino phenotype, reached 66.7%. Some plants exhibited mosaic phenotypes, similar to observations in other studies of CRISPR/Cas9 editing in plants[35−37]. The main possible causes for these heterozygous albino phenotypes are base deletions or base substitutions leading to the substitution or deletion of individual amino acid residues in CmPDS, or the introduction of premature stop codons leading to early termination of translation. The partial CmPDS protein produced due to a premature stop codon may still be translated properly and perform its function as a catalytic enzyme, sometimes to a lesser degree than the complete protein, which may be one of the main reasons for the emergence of chimeric phenotypes. In addition, during the callus induction stage of leaves infected by Agrobacterium, it is difficult to completely separate mutant and non-mutant cells due to their close aggregation, which may be another important reason for the emergence of chimeric plants. Thus, these chimeric plants do not offer stable inheritance of targeted genes/traits.
CRISPR/Cas9 technology can be used to enhance the tolerance of ornamental plants to certain diseases, pests, and stresses, thereby creating new plant germplasm resources with strong resistance. In Arabidopsis, introducing functional mutations in the WRKY3 and WRKY3 genes using this technology reduced plant tolerance of salt and Me-JA stress. In addition, expression Cas9/sgRNA in Arabidopsis could cause deletion or mutation of nucleotide sequences in key functional regions of plant viruses[37]. Chrysanthemum White Rust (CWR) caused by Puccinia horiana Henn. is a major disease affecting the production of chrysanthemum. In a previous study, the transcript level of CmTGA1 was found to be up-regulated upon P. horiana infection. We chose this gene to test the editing efficiency of our system.
When we used CmTGA1, our date verified the high editing efficiency using the pHSE401 vector. We observed genome editing in three of the six CmTGA1 transgenic plants, for a genome-editing efficiency of 50%, confirming that the pHSE401 vector has extremely high gene-editing efficiency in C. morifolium. At the same time, we confirmed that knockout of the CmTGA1 reduced resistance to P. horiana.
In previous studies, the CRISPR/Cas9 system in chrysanthemum was used to obtain a CRISPR-CpYGFP-chrysanthemum shoot containing a mutation in the CpYGFP sequence[38]. In our study, we first targeted knockout of the CmPDS gene in C. morifolium and verified editing efficiency by knocking out CmTGA1. Knockout of CmPDS gene provides a clear visible marker of transformation. The successful knockout of CmTGA1 further supports the reliability of our system and provides important information for battling CWR disease.
In conclusion, the combination of transient and stable transformation in this study provided efficient and successful genomic site-directed editing, saving both time and money. The CRISPR/Cas9 gene-editing system in C. morifolium developed in this study will facilitate basic research on C. morifolium gene function and the genetic improvement and breeding of this plant.
-
In this study, we firstly established the CRISPR/Cas9 system by targeting CmPDS gene in chrysanthemum and combined the TCEP method with stable transformation to ensure the reliability. We further validated editing efficiency by knocking out CmTGA1 and verified the disease resistance function of CmTGA1.
-
The authors confirm contribution to the paper as follows: study conception and design: Mao H, Chen Q; performing experiment: Chen Q, Zhang X; data analysis and article discussion: Jin R; draft manuscript: Mao H, Chen Q. All authors contributed to the article and approved the final version.
-
All data generated or analyzed during this study are included in this manuscript and its supplementary information files, and are available from the corresponding author on reasonable request.
We thank the National Natural Science Foundation of China (31972447) and Foundation of Liaoning Province Education Administration (LJKMZ20221029) for supporting our study.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Qi Chen, Xin Zhang
- Supplemental Fig. S1 (a) The map of the pCBC-DT1T2 plasmid. (b) The map of the pHSE401 vector. (all the maps are obtained from Miaoling Plasmid Platform).
- Supplemental Fig. S2 (a) The schematic representation of the target sites of CmPDS. (b) The schematic representation of the target sites of CmTGA1.
- Supplemental Fig. S3 Construction of gel electrophoresis maps using the CRISPR-CmPDS vector. (a) PCR amplification of sgRNA-CmPDS. (b) PCR verification of the Agrobacterium solution. (c) PCR validation of transgenic Agrobacterium tumefaciens.
- Supplemental Fig. S4 PCR verification of CRISPR-CmPDS transgenic plants.
- Supplemental Fig. S5 Construction of gel electrophoresis maps using the CRISPR-CmTGA1 vector. (a) Verification of the amplification of sgRNA-CmTGA1. (b) PCR verification of the Agrobacterium solution. (c) PCR validation of transformed Agrobacterium tumefaciens cells.
- Supplemental Fig. S6 PCR verification of CRISPR-CmTGA1 transgenic plants.
- Supplemental Table S1 The gene squences of CmTGA1 and CmPDS.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen Q, Zhang X, Jin R, Mao H. 2024. Establishment of a CRISPR/Cas9 gene-editing system for Chrysanthemum morifolium. Ornamental Plant Research 4: e014 doi: 10.48130/opr-0024-0012 

Establishment of a CRISPR/Cas9 gene-editing system for Chrysanthemum morifolium
- Received: 01 February 2024
- Revised: 21 March 2024
- Accepted: 22 March 2024
- Published online: 16 May 2024
An Author Correction to this article was published on 23 October 2025, http://doi.org/10.48130/opr-0025-0041.
Abstract: Chrysanthemum morifolium ranks among the four highest-selling cut flowers worldwide. Gene editing is an important tool to research gene function, but efficient and precise genome-editing tools are not available for chrysanthemum. Here, we established a CRISPR/Cas9-mediated gene-editing system to explore gene functions and enhance the breeding of chrysanthemum. We used the Golden Gate Assembly system to construct CRISPR/Cas9 vectors for dual targeting of the Phytoene Dehydro (PDS) gene. To test the accuracy of sgRNA design, we initially used the transient CRISPR/Cas9 editing in plants (TCEP) method. Target gene expression in nine plants subjected to transient transfection was 19.1%–52% of the normal level, confirming the feasibility of target gene knockout. We carried out stable transformation; PCR and sequencing of the target sites showed that four of eight albino plants obtained had been stably edited at the target sites. We further assessed the editing efficiency of the system by targeting another gene, CmTGA1, chosen because of its potential importance in Chrysanthemum White Rust (CWR) disease progression. Our data indicates that combining transient and stable transformation improves the efficiency and success rate of genome fixed-point editing. The effective, heritable CRISPR/Cas9-mediated genome-editing system we have established here lays the foundation for functional gene studies and genetic improvement of C. morifolium.
-
Key words:
- CRISPR/Cas9 /
- Chrysanthemum morifolium /
- CmPDS /
- CmTGA1 /
- Gene editing