








-
Floriculture is one of the major and dynamic branches of horticultural sciences, since antiquity ornamentals crops have always dazzled humans. In accordance with a floriculture industry analysis conducted by Coherent Market Insights (
www.coherentmarketinsights.com ): The global floriculture market was valued at 34 billion Euros in 2022 according to reports from the International Association of Horticultural Producers (AIPH). Europe holds the largest market share (40.3%) in global floriculture production, which is 16% of the total floriculture market, while India, China, and the Netherlands are the world's largest producers of cut flowers and potted plants, ornamental plants, and seed balls, respectively. The main flower exporting countries are the Netherlands, Colombia, Italy, Germany, and Ecuador, and the main flower importers are Germany, the Netherlands, the United States, France, and the United Kingdom. The floricultural industry is sustained by novelty, and there has been a quest for the development of new cultivars in floricultural crops. Due to constantly expanding demand, the floricultural farming industry requires new and better varieties with novel characteristics in ornamental plant breeding, such as flower longevity, enhancement of flowering, regulation of flowering time, a novelty in colour range, and perfumes[1,2]. To keep up with demand, the ornamental plant industry wants access to cutting-edge plant varieties with superior characteristics, such as enhanced anatomical qualities, flower colour, pigments, stress tolerance, and disease resistance[3]. This may be with the usage of conventional breeding, like heterosis, hybridization, or by other means like cybridization, somaclonal variations, or with hybridoma techniques. Despite the widespread use of traditional breeding methods, there are clear drawbacks and limitations to these approaches, like low efficiency, time-consuming, and non-directional[4]. Recently, many biotechnological interventions have been carried out to develop new varieties, including the development of transgenics and gene editing. As we have seen in many crops like rice, maize, tomato, etc, there have been huge ethical and religious issues related to the development of transgenics. Many fears were voiced and ideas formed as a result of the increasing population globally and the direct impact these field crops had on human life, such as their use as food. There are no or negligible ethical issues related to the development of transgenic ornamentals, but the researchers were not able to develop the most striking novel traits like colour modification, plant architectural changes, vase period, and flowering time modifications. More than a thousand plant species have been genome sequenced, the post-genomic age necessitated the development of efficient gene function analysis tools by scientists[5,6]. Genetic engineering, genome editing, and similar procedures have gained widespread acceptance over the past decade as practical alternatives to the inherent difficulties that have plagued more traditional approaches[7]. At the beginning of gene editing, to understand gene function, metabolic pathways, and regulatory mechanisms, the scientific community has extensively adopted reverse genetics techniques including gene silencing[8]. However, some of the currently used gene silencing techniques, such as RNA interference, have several inherent flaws, such as instability and insufficient silencing, that can make it difficult to understand the phenotypes that arise. Given that it introduces mutations in the genome that are inherently stable and heritable across many generations, targeted genome editing provides a variety of advantages over other methods[9]. Several methods, including Zinc Finger Nucleases (ZFNs), Transcription Activator Like Effector Nucleases (TALENs), and Clustered Regularly Interspaced Short Palindromic Repeats and their associated protein systems (CRISPR/Cas) have been developed. Within these techniques, several different CRISPR/Cas systems have been developed to achieve target-specific genome editing and are more effective and can edit many target genes at once than their predecessors, ZFNs, and TALENs[10]. For CRISPR/Cas systems, the Cas9 protein system is the most widely used in plant research, although other Cas proteins have also been reported like Cas12 and Cas13. Researchers have been able to accelerate their efforts to modify crops by employing CRISPR/Cas9, a genetic editing technique with great promise and easier application. The utilization of CRISPR/Cas9 genome-editing technology which has actively promoted the efficiency of crop breeding helped agricultural and horticulture crops[11]. Numerous new research avenues have been made possible by the advent of CRISPR/Cas systems. Along with CRISPR interference (CRISPRi) and CRISPR activator (CRISPRa) gene regulators, this involves the development of a base editor, prime editor, and epigenetic editor. Furthermore, it has imaging applications for targeting chromatin and RNA[12]. Many areas of biology, including medicine and agriculture, have been revolutionized as a direct result of its extensive application. Screening and detection of human and plant diseases were aided by CRISPR during the coronavirus (COVID-19) epidemic[13]. Here, we describe the current state of enabling technologies and data on genes underlying features of interest, as well as update our previous evaluation of genome editing applications in ornamental plants (Fig. 1).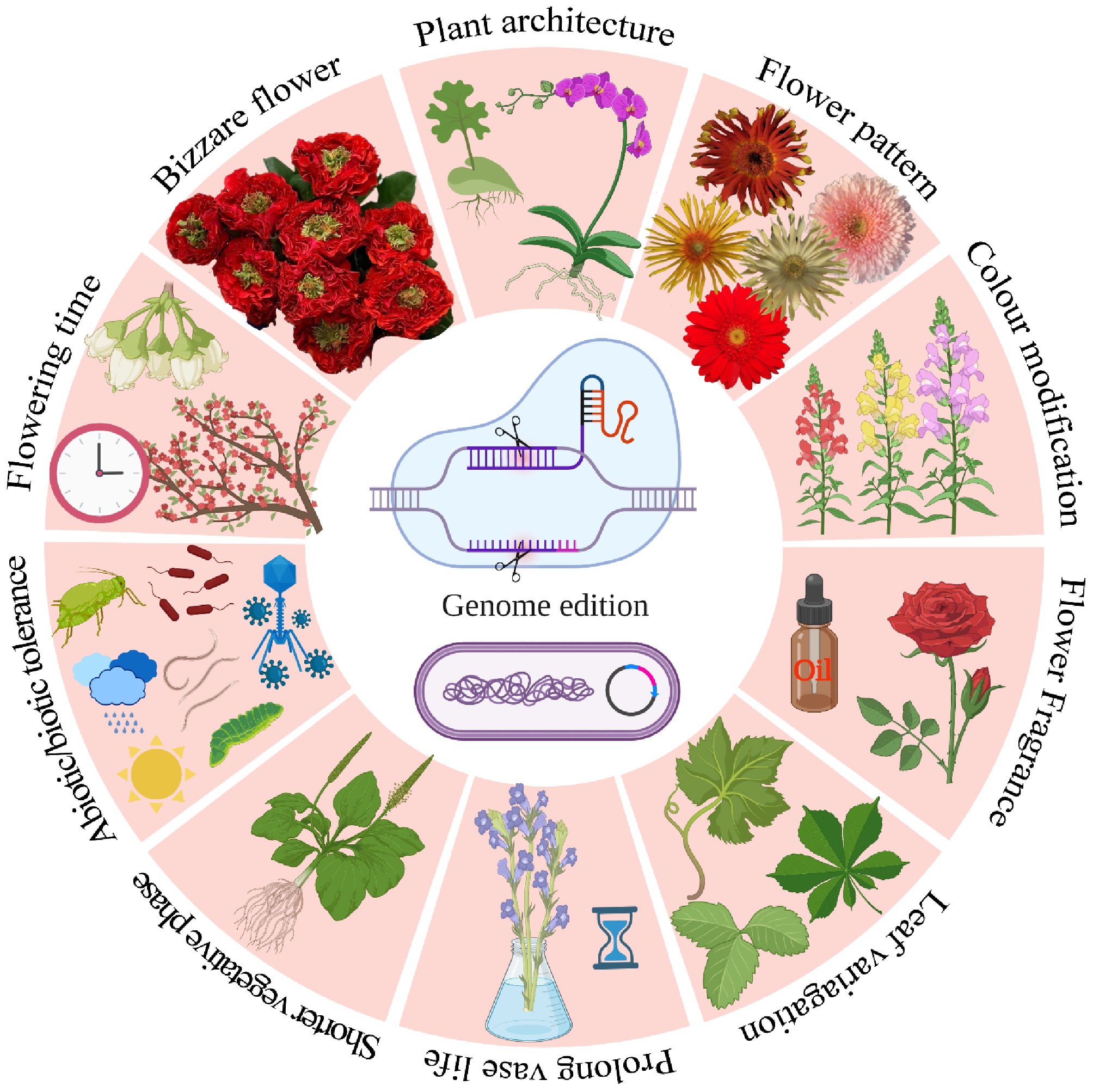
Figure 1.
Applications of genome editing strategies in breeding novel ornamental crops.
-
The use of genome sequencing technology (Table 1) has had a major impact in recent decades on the development of better-blooming crops[14]. While mutation breeding results in a 'subtractive' one-point improvement in floriculture, genetic transformation is vital for the creation and improvement of floricultural crops. Since the first recorded transformation in ornamentals in the eighties[11], genetic engineering technology has been widely regarded as an important strategy for creating new, desirable cultivars. The application of cutting-edge technologies such as CRISPR tools, which require sequence information, has been given a significant boost by the recent completion of the sequencing of the entire genomes and transcriptomes of several important floricultural ornamental crops. It has been clear that knowing the genome sequences of significant ornamental plant species will greatly benefit their breeding and research[15]. So far, the genomes of more than 100 ornamental plants have been sequenced, including 14 annuals, three ornamental aquatics, 19 cut flowers, one cacti, seven ornamental grasses, eight herbaceous perennials, seven orchids, five pot plants, 51 shrubs, and 14 trees[16]. Targeted mutagenesis using CRISPR/Cas9 will improve our understanding of gene function in these crops, which in turn will allow us to develop more effective genome editing tools. Genome editing also necessitates standard operating procedures for cell transformation and regeneration. This method has been standardised for a wide range of crops, but for many ornamental plants, there are no established guidelines for this methodology. As our understanding of tissue culture expands, we will be able to more quickly implement uniform techniques for plant regeneration across all crops.
Table 1. A comprehensive and comparative genomics of ornamental plants, insights into genome size, chromosome number, and sequencing technology.
No. Species Sequencing technology Genome size Chromosome no., and ploidy level Ref. (A) Annuals 1 Amaranthus hypochondriacus Illumina HiSeq 2500 466 Mb 2n = 32 [17] 2 Antirrhinum majus Illumina HiSeq 2000, PacBio RS II 520 Mb 2n = 16 [18] 3 Eschscholzia californica Illumina HiSeq 2500 502 Mb 2n = 12 [19] 4 Gypsophila paniculata MGISEQ-2000 PE 150 5–10 Mb 2n = 2x = 34 [20] 5 Helianthus annuus PacBio RS II 3.6 Gb 2n = 34 [21] 6 Ipomoea nil PacBio, HiSeq 2500 750 Mb 2n = 2x = 30 [22] 7 Mimulus guttatus Illumina HiSeq 2000 531 Mb 2n = 28 [23] 8 Mimosa pudica Illumina HiSeq 2000/4000 896 Mb 2n = 4x = 48 [24] 9 Petunia axillaris HiSeq 2500, PacBio 1.4 Gb 2n = 2x = 14 [25] 10 Petunia inflata HiSeq 2500, PacBio 1.4 Gb 2n = 2x = 14 [25] 11 Salvia splendens PacBio RS II; Illumina HiSeq X Ten 711 Mb 2n = 2x = 44 [26] 12 Tarenaya hassleriana Illumina HiSeq 2000 300 Mb 2n = 20 [27] (B) Aquatic 1 Nelumbo nucifera Illumina HiSeq 2000 879 Mb 2n = 16 [28] 2 Nymphaea colorata PacBio RS II, Hi-C 433 Mb 2n = 28 [29] (C) Cut flowers 1 Chrysanthemum seticuspe Illumina HiSeq 2000 and MiSeq 3.0 Gb 2n = 2x = 18 [30] 2 Chrysanthemum nankingense Oxford Nanopore, HiSeq 2000, PacBio RS II 3.07 Gb 2n = 18 [31] 3 Chrysanthemum lavandulifolium PacBio RS II platform, Oxford Nanopore 2.60 Gb 2n = 18 [32] 4 Dianthus caryophyllus 'Scarlet Queen' contig N50: 14.67 Mb; BUSCO: 97.15% 636 Mb 2n = 2x = 30 [33] 5 Eustoma grandiflorum PacBio HiF 1.71Gb 2n = 6x = 72 [34] 6 Platycodon grandiflorus Illumina HiSeq 2500 platform 680 Mb 2n = 2x = 18 [35] 7 Primulina huaijiensis Illumina HiSeq 2500/HiSeq X Ten platform 478 Mb 2n = 36 [36] 8 Rosa chinensis Illumina HiSeq 2500, PacBio RS-II 532.7 Mb 2n = 2x = 14 [37] 9 Rosa multiflora Illumina MiSeq and HiSeq 2000 platforms 711 Mb 2n = 2x = 14 [38] 10 Rosa rugosa, PacBio HiFi 382.6 Mb 2n = 14 [39] 11 Rosa wichuraiana 'Basye's Thornless' PacBio HiFi, Oxford Nanopore 530.07 Mb 2n = 14 [40] 12 Rosa roxburghii Illumina HiSeq 2500 480.97 Mb 2n = 14 [41] (D) Cacti 1 Carnegiea gigantea Illumina HiSeq 2000, MiSeq 980.3 Mb–1.3 Gb – [42] (E) Grasses 1 Lolium perenne Illumina HiSeq 2000 99 Gb 2n = 14 [43] 2 Trifolium pratense Illumina HiSeq 2000 420 Mb 2n = 14 [44] 3 Trifolium subterranum Illumina HiSeq 2000 552.4 Mb 2n = 16 [45] 4 Zoysia japonica Illumina HiSeq 340 Mb 2n = 40 [46] 5 Zoysia matrella Illumina HiSeq 2000, MiSeq 423 Mb 2n = 40 [46] 6 Zoysia pacifica Illumina HiSeq 2000, MiSeq 302 Mb 2n = 40 [46] 7 Selaginella lepidophylla PacBio RsII platform, Illumina MiSeq platform 122 Mb 2n = 20 [47] (F) Herbaceous perennials 1 Asparagus setaceus HiSeq X Ten, Hi-C 720 Mb 2n = 20 [48] 2 Drosera capensis Illumina HiSeq 2500 264–293 Mb – [49] 3 Dionaea muscipula PacBio RS II 3.19 Gb – [50] 4 Boea hygrometrica Illumina HiSeq 2000, Roche 454 1.69 Gb – [51] 5 Catharanthus roseus Illumina HiSeq 2000 738 Mbp 2n = 16 [52] 6 Primula veris Hiseq 2000, Miseq, Ion Proton, PacBio 479.22 Mb 2n = 2x = 22 [53] 7 Rhodiola crenulata Illumina HiSeq 2000/4000 420.2 Mb 2n = 38 [54] 8 Tanacetum cinerariifolium HiSeq X, HiSeq 4000 7.1 Gb 2n = 18 [55] (G) Orchids 1 Apostasia shenzhenica Illumina HiSeq 2000, PacBio 471 Mb 2n = 68 [56] 2 Dendrobium catenatum HiSeq 2000 1.1 Gb 2n = 2x = 38 [57] 3 Dendrobium officinale Illumina HiSeq 2000, PacBio RS II 1.27 Gb 2n = 2x = 38 [58] 4 Phalaenopsis aphrodite GAIIX, HiSeq 1.20 Gb 2n = 2x = 38 [59] 5 Phalaenopsis equestris Illumina HiSeq 2000 1.16 Gb 2n = 2x = 38 [60] 6 Cymbidium ensifolium PacBio RSII platform 3.62 Gb 2n = 2x = 40 [61] 7 Cymbidium goeringii PacBio Sequel platform 4.10 Gb 2n = 40 [62] (H) Pot plants 1 Begonia fuchsioides Illumina HiSeq 2000/4000 935 Mb 2n = 2x = 22 [24] 2 Kalanchoe fedtschenkoi MiSeq 260 Mb 2n = 34 [63] 3 Primula vulgaris Illumina HiSeq 2500 474 Mb 2n = 22 [64] 4 Rhododendron simsii PacBio RS II, Hi-C 525 Mb 2n = 26 [65] 5 Sedum album PacBio RS II 305 Mb 2n = 48 [66] (I) Shrubs 1 Chimonanthus praecox Illumina HiSeq 2000, PacBio RS II, HiSeq X, Hi-C 778.71 Mb 2n = 22 [67] 2 Chimonanthus salicifolius Illumina HiSeq 2000, PacBio RS II, Hi-C 835.5 Mb 2n = 22 [68] 3 Forsythia suspensa Illumina HiSeq 2500, Oxford Nanopore 701.40 Mb 2n = 28 [69] 4 Gardenia jasminoides Illumina HiSeq 2000, Oxford Nanopore, Hi-C 547.5 Mb 2n = 22 [70] 5 Hibiscus syriacus HiSeq 2000 1.9 Gb 2n = 4x = 80 [71] 6 Jasminum sambac Illumina HiSeq X Ten platform - 537.99 Mb 2n = 2x = 26 [72] 7 Lavandula angustifolia Illumina HiSeq 2000 870 Mb 2n = 50 [73] 8 Osmanthus fragrans HiSeq X ten, Hi-C 733.5 Mb 2n = 46 [74] 9 Paeonia suffruticosa BGISEQ-500, PacBio RS II 13.66–15.76 Gb 2n = 10 [75] 10 Rhododendron delavayi Illumina HiSeq 2000 697.94 Mb 2n = 26 [76] 11 Rhododendron williamsianum Illumina HiSeq 2000, Hic 532.5 Mb 2n = 2x =26 [77] 12 Rhododendron griersonianum Illumina HiSeq X Ten system 677 M 2n = 26 [78] 13 Rhododendron ovatum PacBio Sequel platform 549 Mb 2n = 26 [79] (J) Trees 1 Bombax ceiba Illumina HiSeq 2000, PacBio RS II 809 Mb – [80] 2 Camptotheca acuminata Illumina HiSeq 2000 516 Mb 2n = 44 [81] 3 Casuarina equisetifolia Illumina HiSeq 2000, PacBio RS II 300 Mb – [82] 4 Casuarina glauca Illumina HiSeq 2000/4000 314 Mb 2n = 18 [24] 5 Cerasus serrulata Illumina X ten, Nanopore, Hic-C 256.65 Mb 2n = 16 [83] 6 Cerasus yedoensis Illumina HiSeq 2000, MiSeq, HiSeq X 690.1 Mb 2n = 16 [84] 7 Cercis canadensis Illumina HiSeq 2000/4000 301 Mb 2n = 2x = 14 [24] 8 Fraxinus excelsior Illumina HiSeq 2000, MiSeq, Roche 454 877.24 Mb 2n = 22 [85] 9 Ginkgo biloba Illumina HiSeq 2000/4000 10 Gb 2n = 24 [86] 10 Handroanthus impetiginosus Illumina HiSeq 2000 557 Mb 2n = 40 [87] 11 Prunus mume Illumina GA II 280 Mb 2n = 16 [88] 12 Prunus yedoensis HiSeq X Ten, PacBio RS II 257 Mb 2n = 16 [89] 13 Liriodendron chinense Illumina HiSeq 2000, PacBio RS II, Bionano 1.8 Gb 2n = 38 [90] 14 Jacaranda mimosifolia PacBio Sequel II Platform, Illumina HiSeq 2500 platform 707.32 Mb 2n = 36 [91] -
Agrobacterium-mediated transformation uses Agrobacterium tumefaciens to transfer CRISPR components into plant cells, leveraging its efficiency and ability to integrate foreign DNA into the plant genome. This method has seen significant improvements in transformation efficiency, extending its use to a broader range of ornamental plants[92]. Meanwhile, the biolistic or particle bombardment method propels microscopic particles coated with CRISPR components into plant cells. This technique is particularly useful for plants that are not easily transformed by Agrobacterium, with recent optimizations enhancing delivery efficiency and reducing cellular damage[93]. Polyethylene glycol (PEG)-mediated protoplast transformation facilitates the uptake of CRISPR components by plant protoplasts, although it is technically challenging. Recent research has focused on improving PEG-mediated transformation protocols to increase the efficiency and viability of transformed protoplasts. Among the potential solutions to improve CRISPR delivery, viral vectors such as adenoviral vectors (AdVs), adeno-associated viruses (AAVs), and lentivirus vectors (LVs) are being explored for their high delivery efficiency. These vectors can deliver CRISPR components directly into plant cells, potentially overcoming some of the limitations of traditional methods. Recent advancements have optimized viral vector systems for specific plant species, improving their safety and stability. Additionally, nanoparticle-mediated delivery, which encapsulates CRISPR components in nanoparticles, shows promise despite being experimental. This method targets specific cells and tissues, with recent studies developing novel nanoparticle formulations that enhance delivery efficiency and reduce off-target effects[93]. Another innovative approach is magnetofection, which uses magnetic fields to concentrate CRISPR-loaded magnetic nanoparticles into plant cells, enhancing delivery efficiency and showing potential for targeted delivery in various plant species.
-
In the past, new traits were introduced into ornamental plants through classical breeding methods such as selection, crossing, mutation breeding, polyploidization, heterosis breeding, and chromosome doubling. However, these techniques faced challenges such as high chromosome and ploidy numbers, as well as issues of incompatibility. To overcome these hurdles and improve the gene pool, techniques like tissue culture, embryo rescuing, hybridization, and intra/interspecific crossing were employed. However, to meet the increasing demands for quality and quantity, biotechnological interventions were adopted in ornamental plants[94,95]. In recent decades, biotechnological interventions have gained further momentum with the utilization of genetic engineering to modify specific traits. Genetic engineering has overcome the barriers of cross-breeding and enabled the introduction of genes from distant gene pools. This technology has enhanced the quality and novelty of ornamental plants, including the modification of flower colours (e.g., production of blue chrysanthemum), variegation patterns in petals and leaves, alternatives to growth retardants for compactness, improved tolerance towards ethylene, disease resistance, and flower fragrance (Fig. 1).
Flower colour modification
-
Flower colour modification has always fascinated humans, and it primarily involves three types of bio-pigments: betalains, carotenoids, and flavonoids. There is a constant quest to develop novel colours in ornamental plants, particularly blue flowers. In genetic engineering, the focus has mainly been on modifying the flavonoid pathways in flowers. For example, the expression of the Lc gene (bHLH) under a constitutive promoter (CaMV35S) has been used to enhance anthocyanin production[96]. This approach has also been applied to improve anthocyanin levels in transgenic petunias, which have the potential to become valuable ornamental plants. Researchers have made numerous attempts to engineer blue flowers in popular cut flowers such as roses, carnations, and chrysanthemums, which collectively cover approximately 60% of the global market. However, these flowers only possess pelargonidin and cyanidin derivatives that lack aromatic acyl groups, making them prime targets for engineering the synthesis of delphinidins, which are responsible for blue hues[97]. Expression of the petunia flavonoid 3',5'-hydroxylase (F3'5'H) gene in a carnation line that accumulates cyanidin-based pigments resulted in low levels of delphinidin production and no significant impact on flower colour[98]. However, Florigene Ltd and Suntory Ltd managed to develop transgenic violet carnations (Florigene mauve Moondust™ and Florigene Moonshadow™) by inserting the F3'5'H gene and a petunia DFR gene into a white carnation lacking DFR[99]. These transgenic carnations exhibited higher levels of delphinidin, but they were not completely blue. On the other hand, Torenia flowers with an antisense DFR gene were bluer than those with an antisense CHS gene, as incomplete down-regulation of DFR led to flavone accumulation, resulting in co-pigmentation effects that shifted the flower's colour towards blue[100]. However, down-regulating FNSII genes in blue Torenia resulted in reduced flavone levels and increased precursor flavanones, leading to unexpectedly pale blue flowers[101]. This highlights a major drawback of the transgenic production of blue flowers, as the resulting colours may not be stable or novel over time. An alternative strategy for developing blue-coloured chrysanthemums was developed by Noda et al.[102] called metabolic engineering. They successfully generated the world's first true-blue chrysanthemums by incorporating the butterfly pea UDP-glucose: anthocyanin 3',5'-O-glucosyltransferase gene, and the Canterbury bells F3'5'H gene. This two-step modification of the anthocyanin structure proved to be a simple and effective method to achieve blue flowers in chrysanthemums. In addition to overexpressing specific genes involved in critical enzyme production, selecting an appropriate host with the right genetic makeup is crucial for redirecting pigment biosynthesis and achieving the desired colour change. This approach can help mitigate native route antagonism or allow the downregulation of other antagonistic pathways, ultimately leading to successful flower colour modification.
Flower fragrance and secondary metabolites
-
Floral fragrances play a crucial role in plant reproduction and are also of economic importance due to their enhancement of ornamental plant appearance. Modifying floral perfumes can significantly increase the value of ornamental plants, but a better understanding of the underlying processes is necessary. Flowers produce a wide range of specialized metabolites, including volatiles that attract pollinators, hormones that promote or suppress signalling cascades, and volatiles that protect against herbivores and pathogens[103]. However, flower-specific metabolites are often produced in small quantities, making their identification and characterization challenging[104]. Classical breeding of ornamental plants has led to a reduction in floral fragrance, but a more in-depth understanding of fragrance chemical biosynthetic pathways could help overcome this issue. While few scent genes have been identified, recent research has discovered numerous genes involved in floral aroma biochemistry. However, only a few of these fragrance genes directly encode enzymes that produce volatile chemicals[105]. Genetic engineering has shown promise in enhancing flower fragrance potential. For example, the linalool synthase (LIS) gene, as well as other genes like IEMT, BEAT, and SAMT, have been cloned from Clarkia breweri, a plant species known for its fragrance[106]. Similarly, genes such as (E)-ß-ocimene and myrcene synthases, and BAMT, involved in the synthesis of methyl-benzoate, have been extracted from snapdragon flowers[107]. However, the successful modification of floral scent through genetic engineering has been variable, with some transgenic plants not exhibiting the desired fragrance[108]. Another approach to enhancing floral fragrance is through the transfection of specific genes. For example, lisianthus plants were transfected with the C. breweri BEAT gene, which led to the induction of petal scent through the synthesis of benzyl acetate[109]. Transcriptomic profiling has also provided valuable insights into the development of floral fragrances in plants such as Hedychium coronarium[110]. The regulation of fragrance production pathways involves the action of transcription factors (TFs). While the transcriptional regulation of phenyl propanoid/benzenoid pathway TFs has been extensively studied, the regulation of terpenoid pathway TFs remains unknown. For example, the exclusively expressed ODORNT1 (ODO1) TF regulates the shikimate pathway in petunia petals[111]. R2-R3 type flower-specific TFs, such as Petunia EOBI, have also been identified and found to influence scent gene expression[112]. Despite significant advancements in understanding fragrance production, there is still much to learn. Master regulators that control volatile chemical synthesis and metabolic pathways have yet to be identified. However, the identification of fragrance production genes and enzymes has opened up possibilities for metabolic engineering, leading to improved plant defence and fragrance value. In conclusion, while there is promise in modifying floral fragrances through genetic engineering, our limited knowledge of smell metabolic mechanisms and regulation highlights the need for further research in this field.
-
With the development of biotechnology, the CRISPR/Cas9 system has become a more advanced and useful gene editing technology. Firstly, the CRISPR/Cas9 technique is more efficient than that of Zinc-Finger Nucleases (ZFNs, the first generation, 1993) and Transcription Activator-like Effector Nucleases TALENs (TALENs, the second generation, 2009). Lozano-Juste & Cutler[113] observed higher mutation rates in plants with CRISPR/Cas9 in comparison to ZFNs and TALENs. This is further supported by the facts of Liang et al.[114] who observed that CRISPR was 13.1% more efficient than TALENs (9.1%) while targeting the same sites in Zea mays. Secondly, CRISPR/Cas9 systems are simpler to build than programmable sequence-specific nucleases like ZFNs and TALENs because the target specificity of the nuclease is directly determined by a short sequence in the sgRNA. The specificity of the DNA sequence to which Cas9 will bind is determined by a single-guide RNA (sgRNA) that directs Cas9 to a 20-nucleotide (nt) sequence on the DNA sequence (the so-called protospacer). For Cas9 to bind and cleave the target sequence, an additional 3-nucleotide element (protospacer-adjacent motif; PAM) with the NGG sequence downstream of the target sequence is necessary. Specifically, 3 base pairs (bp) upstream of PAM is where DNA is cleaved. Insertional/deletion mutations of varying sizes can be caused by the repair of Cas9-induced DNA double-stranded breaks (DSBs) within the protospacer. Therefore, CRISPR/Cas9 systems are simpler to build than programmable sequence-specific nucleases like zinc-finger nuclease or transcription activator-like effector nuclease because the target specificity of the nuclease is directly determined by a short sequence in the sgRNA (TALEN). As a result, the CRISPR/Cas9 system has been rapidly adopted for gene editing across a wide range of species[115], such as rice, wheat, potato, rose, orchid, petunia, chrysanthemum, and other ornamentals. All of these genome-edited plants also have genome data from at least one related plant species.
Advantages of the CRISPR/Cas9 system
-
ZFNs or TALENs can theoretically be used to accomplish anything that the CRISPR/Cas9 system can. The fact that so many publications using the CRISPR/Cas9 technology have appeared in such a short period of time, including the first reports of genome editing in some horticultural species[116], nevertheless, emphasizes the technology's undeniable benefits in terms of ease of use, affordability, and adaptability. The CRISPR/Cas9 technology makes it much easier to test various gRNAs for each target gene because it does not involve any protein engineering procedures, in contrast to its predecessors. Additionally, cloning is not necessary because only 20 nucleotides in the gRNA sequence need to be altered to bestow a different target specificity. By employing two complementary annealed oligonucleotides, any number of gRNAs can be generated in vitro[117]. The CRISPR/Cas9 system can now be employed for high-throughput functional genomics applications, making genome editing affordable for any molecular biology laboratory. This enables the quick and low-cost assembly of huge gRNA libraries. Secondly, the CRISPR/Cas9 system enables genomic changes that are not possible with the other nucleases, such as While ZFNs and TALENs cannot cleave methylated DNA in human cells, the CRISPR/Cas9 system enables genomic changes that are not possible with the other nucleases[118]. Although this feature hasn't been extensively studied in plants, it is plausible to believe that the CRISPR/Cas9 system's capacity to cleave methylated DNA is inherent to the system and independent of the target genome. Plants have methylated CpG/CpNpG sites at a rate of about 70%, especially the CpG islands in promoters and proximal exons[119]. Consequently, the CRISPR/Cas9 method is more adaptable for genome editing in plants in general but is especially suitable for monocots with high genomic GC content, such as rice[120]. In contrast, conventional TALENs cannot break DNA containing 5-methylcytosine, since methylated cytosine is identical to thymidine in the main groove. To create TALENs that can cleave methylated DNA, the repeat that identifies cytosine can be swapped out for a repeat that recognises thymidine. Thirdly, the simplicity of multiplexing is CRISPR/primary Cas9's practical advantage over ZFNs and TALENs. It is possible to edit many genes simultaneously by introducing DSBs at various locations[121]. This technique is very effective for eliminating redundant genes or parallel pathways. By focusing on two widely separated cleavage sites on the same chromosome, the same method can also be used to create massive genomic deletions or inversions[121]. The monomeric Cas9 protein and any quantity of distinct sequence-specific gRNAs are all that is needed for multiplex editing with the CRISPR/Cas9 system. For each target site, distinct dimeric proteins are needed when using multiplex editing using ZFNs or TALENs. Finally, the CRISPR research community's open access policy has encouraged the general adoption and application of this technology, as opposed, for instance, to the ZFN platform's exclusive character. The community offers access to plasmids (for example, through the non-profit repository Add gene), web-based tools for choosing gRNA sequences and determining specificity (for example, for plant genomes:
http://cbi.hzau.edu.cn/cgi-bin/CRISPR ;www.genome.arizona.edu/crispr ;www.rgenome.net/cas-offinder etc. These facilities have influenced newcomers to adopt the technology and helped our understanding of the system and its practical applications advance quickly. This technique has the huge potential of avoiding marketing gluts/glitches with reduced supply-demand timing. One of the remarkable aspects of this technology is that it is the most user-friendly among all the genome editing technologies available to date[122].Why do we need to modify floral traits through CRISPR/Cas9 technology?
-
Ornamental plants are remarkably beautiful and eye-catching flowers. Besides their ornamental value, they have pharmaceutical, culinary, confectionery, aromatherapeutic, and essential oil utilisation. For instance, essential oils and secondary metabolites in rose and other crops like jasmine have their utilization in the production of cosmetics, drugs, organic teas, perfumes, hormones, insect pest repellents, etc. Consumers demand distinctive and useful traits within the developed novel cultivars. For instance, people need more colourful flowers and flower sharps, the more novel and exaggerated looks are welcome. What's more, it's even better if add a little scent to some cut flowers that are not floral. However, from the technical means of breeding, it is difficult to achieve the above novel traits, not to mention the specific breeding technical problems such as cross incompatibility. As we know majority of the ornamental plants are not cross-compatible, besides incompatibility issues also arise within and among the many species. Therefore, we agree that conventional plant breeding methods and even conventional biotechnological methods are not sufficient and efficient enough to develop novel and newer traits within ornamental plants. Consequently, there is an urgent need to develop new versatile techniques or tools to overcome the obstacles faced by researchers in developing the novelty within ornamentals. During the last few decades, efforts and achievements have been made in developing the tools for the efficient targeting of plant genomes with CRISPR/Cas9 technology. This is a versatile tool for targeted genome editing in many plants including ornamentals, which brought a new era for the improvement of ornamental plants. However, the technique had not been used in ornamentals to the extent it could be. Because of its high target specificity, the construction of the CRISPR/Cas9 system is easier to construct than other programmable sequence-specific nucleases, like transcription activator-like effector nuclease (TALEN) or zinc-finger nuclease. Consequently, there has been rapid adoption of the CRISPR/Cas9 system for gene editing in any species or a variety of horticultural plants[115]. With the help of another natural, simple, and convenient method, Agrobacterium bacterium the plasmid containing CRISPR/Cas9 can be transferred into most of the plant's cells/organisms to achieve genome editing[123].
-
It's no secret that the global market for ornamental crops like roses, chrysanthemums, orchids, and tulips is worth well over a billion dollars[124]. There is a significant need for novel features linked with acceptable plant architecture, scent, and colour in the ornamental plant industry because these plants are primarily grown for their aesthetic value. Breeding through hybridization and breeding through mutation have long been utilised to create such features[125]. However, the time and effort involved in breeding, the difficulty in achieving homozygous mutation in polyploids, the lack of precision, and the low frequency in producing the desired outcomes call for the implementation of additional technologies, such as transgenic and genome editing technologies. Transgenic plants like the petunia (created by inserting a maize gene expressing dihydro flavanol reductase) and the early flowering chrysanthemum and orchid (created by inserting genes from the MADS-box gene family) have been developed in recent years. The advent of genome editing tools like CRISPR/Cas9 has provided a fresh opportunity for their rapid evolution. Recently sequenced and assembled genomes can benefit from these methods as well, as their annotations can be parsed for insights into their metabolic activity. The first specific genome editing in floricultural crops is been reported in Chrysanthemum, one of the top ten cut flowers on a global level. There have been many hurdles in the breeding of novel flower traits because of the complex ploidy of the genome. The quest for the development of novel varieties in chrysanthemum has surged in the last decade. Conventional biotechnology along with transgenic one has altogether failed to develop novel and sustainable traits in chrysanthemum. An attempt to perform the world's first genome editing with CRISPR/Cas9 system in chrysanthemum was undertaken[126]. First of all, they developed transgenic lines of chrysanthemum expressing a yellowish-green fluorescent protein gene from Chiridius poppei (CpYGFP) and then targeted CpYGFP for gene editing followed by comparing the activity of Cauliflower mosaic virus (CaMV) 35S promoter and parsley ubiquitin calli and derived Cas9 with parsley ubiquitin promoter. In the next step, they choose two sgRNAs for targeting different positions in the CpYGFP gene thus obtaining transgenic calli having mutated CpYGFP genes (CRISPR_CpYGFP-chrysanthemum). After going through DNA sequencing and florescence observations, they finally obtained CRISPR_CpYGFP-chrysanthemum shoot containing a mutation in the CpYGFP sequence. This was the hallmark for exploiting the genome editing with CRISPR/Cas9 system in the floricultural research field (Table 2). The study paved the way for utilising CRISPR/Cas9 system for developing the never-possible varieties or traits in floricultural crops.
Table 2. Comprehensive list of genome-edited ornamentals plants, targets, tools, and phenotypic outcomes.
Species Delivery method Material Cas9 promotor sgRNA promotor Target gene Final produce Phenotype Ref. Campanula portenschlagiana Agrobacterium In vitro grown petioles CaMV 35S AtU6-26 CpEil1a, CpEil1b Plants Increased ethylene tolerance [141] Chrysanthemum morifolium 'Sei-Marin' Agrobacterium Young leaves PcUbi AtU6 CpYGFP Plant Decreased fluorescence [150] Chrysanthemum morifolium Agrobacterium leaf discs CaMV 35S AtU6-26 CmPDS, CmTGA1 Plant Display an albino phenotype [146] Crocus sativus Agrobacterium Corms CaMV 35S − − Plant − [151] Dendrobium officinale Agrobacterium Protocorms CaMV 35S OsU3 C3HC4H, 4CL, CCR, IRX Plant − [127] Dianthus caryophyllus CRISPR/Cas9 ribonucleoprotein (RNP) complex Leaf segments CaMV 35S AtU6-26 ACS1, ACO1 Plant Reduced ethylene production, delayed flower senescence, improved longevity [140] Euphorbia pulcherrima (Poinsettia) Agrobacterium mediated Internode stem F3′H Flower colour changed from vivid red to vivid reddish-orange [152] Fortunella hindsii Agrobacterium Seedlings CaMV 35S AtU6 FhPDS Plant Albino leaves [153] Ipomoea nil 'AK77' Agrobacterium Embryos PcUbi AtU6g InCCD4 Plant Altered flower color [135] Ipomoea nil 'AK77' Agrobacterium Embryos PcUbi AtU6g InEPH1 Plant Extended flower longevity [138] Ipomoea nil 'Violet' Agrobacterium Embryos PcUbi AtU6g InDFR Plant Altered flower color [132] Ipomoea nil 'Violet' Agrobacterium mediated Immature embryo EPH1 Delay in petal senescence [138] Japanese Gentian (Albireo) Agrobacterium mediated Leaf Gt5GT, Gt3'GT, Gt5/3'AT Pale red violet, dull pink, and pale mauve flowers [154] Japanese Gentian (Albireo) Agrobacterium mediated Leaf GST1 Reduced anthocyanin in petals, white and pale blue flower [136] Lilium longiflorum 'White Heaven' Agrobacterium Seedling scales Ubi OsU3 LlPDS Plant Albino leaves [129] Lilium pumilum Agrobacterium Embryos Ubi OsU3 LpPDS Plant Albino leaves [129] Lotus japonicus Agrobacterium mediated LjLb1, LjLb2, LjLb3 Nodule formation in the presence of rhizobia and establishing low free oxygen concentration in nodule [142] Petunia hybrida 'Madness Midnight' PEG Protoplasts − − PhNR Protoplasts − [143] Petunia hybrida 'Madness Midnight' PEG Protoplasts − − PhF3H Protoplasts Flower color [134] Petunia hybrida 'Mirage Rose' Agrobacterium Young leaves CaMV 35S AtU6 PhACO1 Plant Extended flower longevity [137] Petunia hybrid 'Mitchell Diploid' Agrobacterium mediated Leaf DPL Absence of the Vein associated anthocyanin pattern [155] Petunia hybrida 'Mitchell' Agrobacterium Young leaves CaMV 35S AtU6 PhPDS Plant Albino leaves [144] Petunia × hybrida 'Mitchell Diploid' Agrobacterium Young leaves CaMV 35S 6 (AtU6) PhFT1, PhTFL1a, etc. Plant Early Flowering, Increased Branching [130] Petunia inflata Agrobacterium Young leaves CaMV 35S AtU6 PiSSK1 Plant Altered compatibility of pollens [145] Phalaenopsis amabilis Agrobacterium mediated PDS3 Protocorms Albino leaf [156] Phalaenopsis equestris Agrobacterium mediated MADS44, MADS36 and MADS8 Long juvenile period [128] Populus tomentosa Carr. (clone 741) Agrobacterium mediated Leaf PtoPDS Albino phenotype was observed [157] Festuca arundinacea. Transient Agrobacterium mediated transformation Leaf SGR Chlorophyll degradation [158] Salvia miltiorrhiza (SmCPS1) Tanshinone biosynthesis [133] Torenia fournieri 'Crown Violet' Agrobacterium Young leaves CaMV 35S AtU6 TfF3H Plant Altered flower color [131] Torenia fournieri TfRAD1 Leaf sections Flowers with abnormal shape & pigmentation [159] Flower shape
-
Recent work has shown CRISPR/Cas9 as a viable gene-editing tool in chrysanthemum by targeting transgenic plants with a multicopy GFP gene utilising a variety of promoters. Compared to CaMV35s, the ubiquitin promoter from Petriselinum crispum was found to be more successful at inducing mutations in chrysanthemum and, consequently, producing more severe decreases in fluorescence[126]. Dendrobium officinale, a type of orchid, had its genome edited using CRISPR/Cas9 in a separate study. Because of its high lignocellulose content, D. officinale is rarely used in natural medicine. Mutation rates of 10%−100% were achieved in different loci of five genes involved in lignocellulose biosynthesis when multiple gRNAs were used to target these genes[127]. These genes are coumarate 3-hydroxylase, cinnamate 4-hydroxylase, coumarate-coenzyme A ligase, cinnamoyl coenzyme A reductase, and irregular xylem 5. CRISPR/Cas9 has been used to successfully create numerous MADS gene mutants (MADS8, MADS36, and MADS44) in the orchid Phalaenopsis equestris. Successful generation of triple mutants (at a rate of 97.9%) was accomplished, and single and double mutants were also created for the study of the role of individual genes in flowering plant growth[128]. Using Agrobacterium-mediated gene editing and protoplast transfection, the CiPDS (chicory phytoene desaturase) gene was modified under the U6 promotor.
Flowering
-
CRISPR/Cas9 was used to initiate flowering and advance flower growth by targeting MADS genes (MADS, MADS44, MADS36, and MADS8) from the orchid species Phalaenopsis[128]. There are over a hundred different species of the genus Lilium, all of which bloom after emerging from their underground bulbs. In three species of Lilium, PDS has been used as an editing target to show that CRISPR/Cas9 can be used to modify the genome. Fisch and White Heaven cultivars showed mutation rates of 69.57% and 63.64%, respectively[129]. Abdulla et al.[130] successfully utilized CRISPR/Cas9-mediated mutagenesis to target FT and TFL1 genes in petunia, resulting in early flowering and compact growth. The edited plants exhibited significant phenotypic changes, including shorter internodes and increased branching. This study demonstrates the potential of genome editing to enhance ornamental traits in petunia, paving the way for advancements in ornamental plant breeding.
Flower colour
-
The colour of flowers is an important trait in ornamental crops. Recent studies have focused on gene targeting to modify flower colour in various plant species. In Torenia fournieri, the gene for flavanone-3-hydroxylase, a key enzyme in the flavonoid production pathway, was edited. This resulted in a light purple to nearly white bloom colour in 80% of regenerated transgenic lines[131]. Knocking out the InCCD4 gene using CRISPR/Cas9 in an undisclosed plant species led to the development of yellow midribs in the flower petals. In this case, the disruption of the dihydroflavonol-4-reductase-B (DFR-B) gene, which affects anthocyanin production, resulted in flowers with various colours such as white, violet, and pale-violet, as well as stems and leaves devoid of anthocyanins[132]. In Salvia miltiorrhiza, a Chinese medicinal plant, the diterpene synthase gene (SmCPS1) involved in tanshinone production was knocked out using CRISPR/Cas9 to create tanshinone-free plants while leaving other phenolic compounds unaffected[133]. A gRNA was designed to simultaneously target two F3H genes (F3HA, F3HB) in the petunia cultivar 'Madness Midnight'. After transformation into protoplasts, the gRNA induced indel mutations ranging from 9.99% to 26.27%. Among the 67 protoplast-regenerated plants, only one double-knockout mutant exhibited a noticeable flower color change, transitioning from purple to pale purplish pink[134]. In Ipomoea nil cultivar AK77, the carotenoid cleavage dioxygenase 4 (CCD4) gene associated with carotenoid degradation was knocked down using CRISPR/Cas9. The resulting knockout mutant plants displayed pale yellow petals and a 20-fold increase in carotenoid content compared to the wild type[135]. In Japanese gentian flowers, the glutathione S-transferase 1 (GST1) gene was targeted for gene editing by Tasaki et al.[136]. Only 7.5% of the plants underwent editing at the target site, and two mutants with different phenotypes were generated: severe (almost white) and mild (pale blue). Another study focused on the DFR-B gene, which encodes an anthocyanin biosynthetic enzyme. Using CRISPR/Cas9 gene editing in the secondary embryos of Ipomoea nil, a mutation rate of 75% was achieved. Biallelic mutants exhibited green stems and white flowers, while a few monoallelic mutants displayed green stems with violet flowers or violet stems with pale flowers[132]. These studies demonstrate the successful use of CRISPR/Cas9 gene editing to modify flower colour in various plant species, offering potential applications in crop improvement and ornamental plant breeding.
Vase life
-
Reduced activity of 1-aminocyclopropane-1-carboxylate oxidase 1 (ACO) in petunias has been found to result in longer-lasting blooms[137]. ACO is the terminal enzyme in ethylene biosynthesis, and partial knock-out of this gene led to a significant decrease in ethylene production in flower organs. As a result, the mutant flowers exhibited an extended vase life of four additional days, totalling ten days. In another study focused on Ipomoea nil (Japanese morning glory), the primary targets of CRISPR/Cas9 technology were flower longevity and colour[138]. The NAC transcription factor family, which plays a vital role in hormone signalling in plants, was found to influence disease resistance. Specifically, the deletion of the Ephemeral 1 (EPH1) transcription factor using CRISPR/Cas9 resulted in an approximately 84% increase in flower longevity compared to the wild type. This increase was attributed to mutations occurring at single or multiple target sites, involving insertions and/or deletions of one or more bases, ultimately leading to an extension of senescence by up to 10.8 h[132]. Additionally, the cleavage of carotenoids in mature Ipomoea nil petals is mediated by a dioxygenase called indole-3-carboxylate dioxygenase 4 (InCCD4)[135]. Furthermore, the knockout of the RhEIN2 gene (ETHYLENE INSENSITIVE2) in roses was found to significantly impact the response of the flowers to ethylene, resulting in almost complete inhibition of flower opening in the mutant[139]. Lastly, the CRISPR/Cas9 editing of the ACO1 gene in petunias led to reduced ethylene production and increased flower vase life in the T0 mutant[137]. Adedeji et al.[140] successfully edited ethylene biosynthesis genes (ACS1 and ACO1) in carnation using the CRISPR/Cas9 ribonucleoprotein complex system, achieving mutation frequencies ranging from 8.8% to 58.5%. The edited plants exhibited reduced ethylene production, leading to delayed flower senescence and extended vase life. This study demonstrates the potential of CRISPR/Cas9 technology for improving ornamental plant longevity and reducing reliance on chemical inhibitors. Holme et al.[141] utilized CRISPR/Cas9 to knockout CpEil1a and CpEil1b genes in Campanula portenschlagiana, enhancing ethylene tolerance without affecting flower longevity. The study demonstrated an allele dosage effect, where increased mutations led to higher ethylene tolerance.
Nitrogen fixation
-
CRISPR/Cas9 has recently been employed to develop desired features in ornamental crops. Symbiosis receptor-like kinase and three haemoglobin genes in Lotus japonicas were targeted to promote nodule formation and decrease free oxygen content in nodules while in the presence of rhizobia, hence improving symbiotic nitrogen fixation. The mutation rate for symbiosis receptor-like kinase-targeting gRNA was 35%, while that for leghaemoglobin-targeting gRNA was 28.5%[142]. To alter the endogenous nitrate reductase gene in petunia, a model ornamental crop, protoplasts were used[143]. Loss-of-function mutants of the nitrate reductase enzyme are unable to absorb nitrate effectively because they are not able to take in and use the nutrients. As with nitrate reductase, petunia has shown to be capable of a knock-out of PDS with efficiencies ranging from 55.6% to 87.5%[144].
Other traits
-
In a study conducted by Kui et al.[127], five genes involved in the lignin synthesis pathway were edited using the CRISPR/Cas9 system. These genes included coumarin acid 3-hydroxylase (C3H), cassia bark acid 4-hydroxylase (C4H), 4-coumarin acid: coenzyme A ligase (4CL), cassia bark acyl-coenzyme A reductase (CCR), and IRREGULAR XYLEM5 (IRX). After transforming the edited genes into the original stem, the researchers detected base insertions, deletions, and substitutions, which had an impact on the lignin biosynthesis of Dendrobium officinale. In another study by Sun & Kao[145], the piSSK1 gene of the Skp1 subunit in the Petunia SCF-SLF complex was edited using the CRISPR/Cas9 system. The edited gene was then transformed into leaves to investigate the effect of piSSK1 on self-incompatibility. The researchers found that the loss of piSSK1 in transgenic pollen grains inhibited the growth of pollen tubes. Self-incompatibility in Petunia is regulated by a polymorphic S-locus, which includes several genes (SLF) expressed only in the late stages of pollen development, and a single gene expressed only in the pistil during fruit set. For these SLF proteins to function, they must form an E3 ubiquitin ligase complex with another protein containing an F-box sequence (Skp1, Cullin1, and F-box; SCF). In SCFSLF complexes, the Cullin1 (PiCUL1-P) and Skp1 (PiSSK1) subunits are restricted to pollen. While the role of PiSSK1 in self-incompatibility is known, its specific function is still unclear. Crosses between wild-type plants and knock-out mutant lines resulted in no fruit set, demonstrating the essential role of the Skp1 component PiSSK1 in proper SCFSLF complex assembly[145]. Chen et al.[146] established a CRISPR/Cas9 gene-editing system for Chrysanthemum morifolium, targeting the CmPDS and CmTGA1 genes to enhance disease resistance and improve breeding efficiency. The study demonstrated high editing efficiency and successful gene knockout, resulting in albino phenotypes and reduced resistance to Chrysanthemum White Rust.
Aspects of the future genome editing in ornamental species
-
In the floriculture market, one of the most differentiating aspects is the availability of appealing colour combinations. Grape hyacinth was used by Chen et al.[147] to identify MaAN2, an R2R3 transcription factor. In tobacco, transformation, and overexpression of MaAN2 led to the upregulation of genes involved in anthocyanin production, resulting in transgenic tobacco with a noticeable dark hue. Consumers are also drawn to vases because of their long shelf lives. Ethylene is a hormone in plants that has a role in growth and development. When both exogenous and endogenous ethylene levels are high, they trigger ethylene responses that hasten the ageing process. By using virus-induced gene silencing, Liu et al.[148] were able to knock out EIL1 and EIL2 in petunia, two probable downstream components of the ethylene signalling pathway. By keeping an eye on things, they discovered that down-regulating the two EIL genes affects the lifetime of petunia blooms. To test this, we propose that by using CRISPR knock-in to replace one of the EIL genes[148] with MaAN2[147], and then increasing pigmentation through CRISPRa-facilitated upregulation of the transgene, we can improve two types of aesthetic quality in mutant lines at once (beauty and consistency of the beauty per se). The resulting plants would have flowers with intense colour and lengthy vase life, and the consequences would be minimal. The inability of plants to detect ethylene typically results in profound malformation. However, the functional redundancy of EIL genes prevented the observable phenotypic alterations seen in transfected plants after EIL gene silencing, save for the increased bloom lifetime[148]. Colour changes in plants are a typical occurrence, and this is true regardless of customer taste or the ease of identification. High mutation rates are found in enzymes involved in complex biosynthesis, such as anthocyanin and carotenoid synthesis because these pathways involve a wide variety of metabolites catalysed by a large number of enzymes. Point mutations, which occur when there is only one change in a base sequence, are a common cause of albinism and other colourless phenotypes in flowers. Base editing, prime editing, and CRISPR knock-in via HDR are promising CRISPR/Cas9-derived approaches that can be used to alter the altered gene's function[149]. The CRISPR/Cas system has superseded several older methods of genetic engineering. Validation of the safety, stability, and practicability of these newly proposed uses of the CRISPR system is underway, but they are being actively and immediately used. It will be up to society to judge the system's superior performance among other genome-editing techniques, and new findings are expected to speed up plant breeding.
-
CRISPR/Cas9 is poised to revolutionize plant biology research by enabling routine targeted mutagenesis in plants. Unlike traditional random mutagenesis methods, which introduce genes at random locations, CRISPR-Cas9 offers precise editing, allowing researchers to target specific genes for inactivation. This technology holds significant promise for developing mutants of genes that were previously difficult to access, enabling the mutation of multiple loci and the creation of widespread deletions. As a result, plant breeding can progress without the need to introduce transgenes. Genetically modified (GM) plants, if not for concerns about potential environmental and health risks, could have significantly advanced agricultural practices[160]. In traditional genetic engineering, a DNA construct can be introduced into multiple chromosomes, resulting in random genome alterations. However, these random insertions can have unintended effects and are unsuitable for large-scale modifications, such as the incorporation of an entire metabolic pathway into a plant. By contrast, plant breeding has traditionally relied on natural genetic diversity and random mutagenesis to generate novel traits. CRISPR/Cas9 technology, however, enables precise and stable genetic changes in plants that can be inherited across generations. Furthermore, it can be easily erased after gene editing, resulting in plants free from CRISPR modification by the time they are cultivated[161]. Alongside other gene-editing techniques such as zinc finger nucleases (ZFNs) and transcription activator-like effector nucleases (TALENs), the Cas9/sgRNA system has emerged as a cornerstone of new plant breeding technologies (NPBTs). These techniques can rapidly produce null segregant lines that lack transgenic inserts, a process much faster than conventional breeding methods. Importantly, NPBT plants resemble conventionally bred plants and should be evaluated based on their final characteristics, not the method by which they were produced. As a result, CRISPR-edited plants may fall outside the scope of current GMO regulations, raising new challenges for the regulation and acceptance of genetically engineered (GE) crops[162]. Currently, advisory and regulatory bodies are exploring how to classify genome-edited crops under existing GMO legislation, with some advocating for the classification of GE crops created by genome editing as non-GM, to promote social acceptance[162]. Araki & Ishii[163] propose regulatory concepts for GE crops, suggesting that plants modified using CRISPR-Cas9 technology should not be classified as GMOs under product-based GMO regulations. This is because the end product, despite having been genome-edited, does not consist of a genetically modified organism in the traditional sense. However, more research is needed to refine CRISPR/Cas9 systems for plant genome editing, as this powerful tool has the potential to generate valuable agronomic traits in plant species. This approach could complement classical breeding techniques in the study of complex quantitative traits determined through traditional breeding methods. GE plants with regulatory frameworks in place may be more acceptable than those carrying foreign DNA, as they could bypass the common concerns associated with transgenics[113]. In the US, the Department of Agriculture has proposed that GE plants not containing foreign DNA should not be considered GMOs, whereas the European Commission is expected to address the regulatory uncertainties surrounding genome editing in the near future[163]. The success and public perception of CRISPR technology in agriculture will depend on its ability to deliver results in terms of food security and crop improvements[163]. In the European Union (EU), the acceptance of transgenic and NPBTs is shaped by a strict regulatory framework, as evidenced by the 2018 ruling by the Court of Justice of the European Union (CJEU). This ruling classified organisms derived from new mutagenesis techniques as GMOs, subjecting them to the EU's GMO directive while exempting organisms produced through traditional mutagenesis methods with a long safety record (European Union, 2018b, paragraph 54). This regulatory approach reflects the EU's cautious stance toward transgenic crops and NPBTs, resulting in limited acceptance compared to other global regions[164]. The ruling has significant economic implications for the EU's agricultural and food sectors, particularly concerning the importation of goods created using NPBTs. Moreover, it highlights a disconnect between scientific progress in plant breeding and existing regulatory frameworks. Many argue that newer, more targeted techniques like CRISPR may be safer than older methods that remain exempt from regulation. These authors suggest that the CJEU ruling underscores the need for regulatory reform to better align with scientific developments and global agricultural practices[164]. One challenge stemming from the CJEU ruling is that many mutagenic NPBTs are not detectable in the final product and cannot be distinguished from plants altered through traditional breeding methods. While this ruling primarily addresses the EU's regulatory approach, it highlights the more restrictive stance taken in Europe compared to other regions, likely reflecting public concerns about GMOs and new breeding technologies. The tension between scientific progress, regulatory frameworks, public perception, and international trade continues to complicate the widespread acceptance of transgenics and NPBTs in the EU.
-
CRISPR/Cas9 has rapidly gained prominence in molecular biology due to its transformative technological capabilities and unique attributes, including ease of manipulation, high efficiency, and broad applicability. With the complete sequencing of several ornamental plant genomes, the use of CRISPR/Cas9 to modify target genes in these plants is now a tangible reality. This enables researchers to investigate the molecular mechanisms regulating important ornamental traits and to enhance either the aesthetic value or stress resistance of these plants. The integration of advanced technologies like CRISPR/Cas9 is becoming a cornerstone of modern ornamental plant breeding, particularly for the study and modification of traits such as flower colour, type, fragrance, and resistance. These efforts aim to accelerate the development of novel and exotic ornamental plant varieties. Currently, CRISPR/Cas9 is primarily applied for genome editing and gene expression regulation. In addition, its potential extends to DNA labelling and epigenome editing, as demonstrated in studies[115]. While these applications have not yet been explored extensively in plants, CRISPR/Cas9 shows great promise for epigenome editing through modifications such as DNA methylation and histone changes. Similarly, it offers exciting possibilities for DNA tagging using fluorescent-labelled Cas9 proteins and custom guide RNAs (gRNAs). Despite its potential, the use of CRISPR/Cas9 in ornamental plants is not without challenges. Key obstacles include minimizing off-target effects and enhancing the efficiency of Cas9-mediated edits. Tailoring CRISPR vector promoters to suit the unique genomes of different ornamental species is another significant hurdle. Each species' genetic makeup requires customized promoter designs, making the development of universal promoters impractical. This necessitates extensive research to optimize promoter efficiency and precision for each species. Additionally, improving homology-directed repair (HDR)-a critical pathway for precise gene editing—remains a priority in overcoming technical limitations. The floriculture industry stands to benefit immensely from the advancements in CRISPR/Cas9 technology. For instance, it can be leveraged to enhance ornamental traits such as flower colour, shape, and fragrance, creating unique varieties that appeal to consumers. CRISPR also has the potential to develop disease-resistant plants, reducing dependency on chemical treatments and promoting sustainable cultivation practices. Moreover, extending the vase life of cut flowers is a major industry goal, and CRISPR can be employed to modify genes that regulate flower senescence, ensuring prolonged freshness. Additionally, CRISPR can improve the tolerance of ornamental plants to abiotic stresses such as drought, heat, and salinity, enhancing their resilience and ease of cultivation. Looking ahead, further research is essential to refine CRISPR/Cas9 applications and expand its utility in ornamental horticulture. Advancements in these areas will not only address the current challenges but also unlock new opportunities for innovation, transforming the ornamental plant industry.
This work was supported by Yunnan Xingdian Talents—Special Selection Project for High-level Scientific and Technological Talents and Innovation Teams-Team Specific Project, Xingdian Talent Support Project (CYRC2020004), and Yunnan Xingdian Talents—Youth Special Project (XDYC-QNRC-2022-0731).
-
The authors confirm contribution to the paper as follows: writing – original draft, reviewing, and editing: Din A, Wani MA, Nazki IT; supervision and funding acquisition: Jin C, Ma J, Li F. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Din A, Wani MA, Jin C, Nazki IT, Ma J, et al. 2025. Post-genomic era of CRISPR/Cas technology in ornamental plants: advantages, limitations, and prospects. Ornamental Plant Research 5: e010 doi: 10.48130/opr-0025-0007 

Post-genomic era of CRISPR/Cas technology in ornamental plants: advantages, limitations, and prospects
- Received: 12 July 2024
- Revised: 03 January 2025
- Accepted: 15 January 2025
- Published online: 07 March 2025
Abstract: The global ornamental plant industry is still booming, and the demand for new combinations of plant quality, resistance, and germplasm resources is increasing. Traditional hybridization, mutagenesis, and mutation breeding strategies have made outstanding contributions to cultivating new varieties. However, due to its long cultivation cycle, there are limitations in adapting to the rapidly changing quality requirements of the market. In the past 20 years of the genome era, especially the emergence and development of transgenic and gene editing technologies, molecular biologists have recognized and verified the intrinsic molecular mechanism of many plant traits, providing an operational blueprint for efficient and directional molecular breeding of ornamental plants. The latest development of CRISPR/Cas technology, knockout, and knock-in technology, provides a powerful tool to meet the simultaneous increase of beneficial traits and knockout of adverse traits. Its faster breeding cycle, lower breeding cost, and higher accuracy show a broader prospect for molecular breeding in the post-genome era. In this paper, the application of CRISPR/Cas technology in ornamental plants is reviewed. In addition, we also explore the limitations of its use in ornamental plants. Although CRISPR/Cas technology has many advantages, in some applications, the argument against the use of CRISPR/Cas technology is valid. We think it is important to carefully assess the exact benefits and disadvantages of using this technique in floral crops.
-
Key words:
- CRISPR /
- Cas /
- Ornamentals /
- Genome editing /
- Sequencing