









-
Roses are one of the most popular cut flowers and have been cultivated all over the world for more than 2,000 years, containing 200 species and 35,000 cultivars[1], being both an important ornamental plant and a crucial oil crop. For centuries, roses have been used as a raw material for natural fragrances and oil[2], also known as 'liquid gold'. In addition, roses have other economic benefits for humans, such as being used as food, pies, tea, medicine, creams, and crafts[3]. Rosa gigantea, R. chinensis 'Hume's Blush Tea-scented China', 'Parks' Yellow Tea scented China', 'Old Blush', and 'Chilong Hanzhu' are among the most important progenitors in the cultivation of modern roses[4,5]. Previous studies have found that the Chinese rose is rich in distinctive fragrance, while various modern roses lack fragrance diversity due to breeders' emphasis on the appearance of roses rather than fragrance during the long history of traditional cross-breeding[1,6]. Hence, increasing the fragrance diversity in roses can not only improve their performance as an ornamental flower but also promote their economic benefits.
Plants emit a variety of volatile organic compounds (VOCs) from different tissues (such as roots, leaves, and fruits), and flowers were found to be an important apparatus that releases the highest amounts and diversity of VOCs[7−9]. Based on their biosynthetic origin, floral VOCs can be divided into three major classes, terpenoids, phenylpropanoids/benzenoids, and fatty acid derivatives[8,10]. Recently, there has been a growing emphasis on the study of genes related to the VOCs' biosynthesis in Clarkia breweri (C. breweri), Antirrhinum majus (A. majus), Arabidopsis thaliana (A. thaliana), Petunia hybrida (P. hybrida), and Rosa. For example, terpene synthases (TPSs) have been confirmed to participate in the formation of monoterpenes such as linalool, (E)-β-ocimene, germacrene D, and other terpenoids in C. breweri, A. majus, A. thaliana, and R. hybrida[11−15]. In petunia, phenylacetaldehyde synthase (PAAS), as a member of the aromatic amino acid decarboxylase (AADC) family[16−18], catalyzes the formation of phenylacetaldehyde (PAld), which is a precursor for the synthesis of 2-phenylethanol (2-PE)[16]. In the biosynthesis of phenylpropanoid compounds, O-methyltransferases play an essential role in important phenylpropanoid VOCs, such as 3,5-dimethoxytoluene (DMT) and 1,3,5-trimethoxybenzene (TMB) in rose[19,20], and methyleugenol and isomethyleugenol in C. breweri[21].
The fragrance of rose is primarily composed of monoterpene alcohols, including 2-PE, citronellol, geraniol, nerol, and acetate derivatives[22]. The 2-PE, a significant floral compound in roses, plays a crucial role in the production of perfumery and essential oils[20]. The concentration of 2-PE showed significant variability among different rose species. R. multiflora and R. damascena were demonstrated with elevated levels of 2-PE, whereas cultivars in R. chinensis ('Hermosa', 'Miss Lowe', and 'Semperflorens') and modern rose ('Papa Meiland') were observed with trace amounts of 2-PE emitted from petals[23]. In the biosynthesis of 2-PE, two key enzymes, aromatic amino acid aminotransferase (AAAT) and phenylpyruvate decarboxylase (PPDC), are shown to be responsible for 2-PE production based on L-Phe via phenylpyruvic acid (PPA)[24,25]. In rose, the key enzymes involved in the biosynthesis of 2-PE were identified, including the enzymatic actions of AADC and phenylacetaldehyde reductase (PAR)[17,26]. The overexpression of AADC, AAAT, and PAR genes resulted in an elevation of 2-PE content in Solanum lycopersicum (S. lycopersicum)[27,28], and AADC, AAAT in R. rugosa 'Tanghong'[29]. Hence, investigating the control of 2-PE biosynthesis and release in rose is crucial for understanding this process.
In this study, we selected R. chinensis 'Old blush' ('OB') which has a light fragrance and pink petals, and R. chinensis 'Chilong Hanzhu' ('CH') which has intensely fragrant and red petals as materials. We identified differentially abundant volatiles in the petals of 'OB' and 'CH' by gas chromatography-mass spectrometry (GC-MS). Combining transcriptomics analysis, we identified key genes involved in the floral fragrance biosynthesis pathway of R. chinensis. Further confirmation by performing transient transformation of key genes in tobacco leaves uncovered their contribution to the accumulation of 2-PE. These new findings provide a theoretical basis for molecular breeding of Chinese rose fragrance.
-
Two rose germplasm resources, R. chinensis 'OB' and R. chinensis 'CH', were cultivated in the same greenhouse under natural conditions in Shenzhen Comprehensive Experimental Base, Chinese Academy of Agricultural Sciences, Guangdong Province, China (22.60° N, 114.51° E). The flowers were divided into three developmental stages: green flower buds (S1), flower buds with petals beginning to change color (S2), and full blooming flowers (S3), according to previous research[30,31]. For in vivo bioassay of RcCH_AADC1, RcCH_PAR1, and RcCH_PAR1_Like recombinant proteins, Nicotiana benthamiana (N. benthamiana) plants were grown in a greenhouse at 22 °C under a light/dark period of 16 h/8 h, light intensity of 200 μmol·m−2·s−1 and relative humidity of 60%. The young leaves of 4-week-old N. benthamiana were used for Agrobacterium tumefaciens (A. tumefaciens) infiltration experiments.
The GC-MS based profiling of VOCs in R. chinensis
-
The collection of floral scent was performed according to a previous study[32]. Briefly, the fresh rose flowers were excised from plants, and approximately 0.7 g of flower samples were transformed into a collection bottle. Four independent replicates were performed at the S3 stage of R. chinensis 'OB' and 'CH' petals. The solid-phase microextraction was conducted to collect floral metabolites and the extracts were injected into GC-MS (QP-2010, Shimadzu, Japan) for identifying VOCs with a setup as described previously[31]. After obtaining the chromatograms and mass spectrograms of floral compounds, their qualitative analysis was performed by searching the mass spectral and retention index (RI) against the National Institute of Standards and Technology (NIST, USA). The final relative contents of detected metabolites were based on the percentage of the peak area of selected mass to the total peak area of detected metabolites. The VOCs with an area of > 1% in 'OB' and 'CH' were considered to be the predominant VOCs of that sample. The odor of detected floral compounds were annotated according to the description from public website Flavornet (
www.flavornet.org ).Transcriptome analysis
-
The raw RNA sequencing data of different flower stages of 'CH' and 'OB' were obtained from a previous study[31] and public database NCBI (PRJNA351281 and PRJNA546486), respectively. Those datasets include nine (three biological replications for each stage) and six samples (two biological replications for each stage) for 'CH' and 'OB', respectively. Quality control was performed using Fastqc v0.11.9, adapter and low-quality reads (Q < 20) were removed using Trimmomatic[33]. The clean reads were mapped to the R. chinensis 'CH' reference genome using HISAT2 v2.2.1[34], and the mapped reads were assembled using StringTie v2.1.6[35] and expression levels were predicted. The read counts of these samples were calculated using HTSeq[36] and used for differential gene expression analysis.
The gene expression level was evaluated by fragments per kilobase of transcript per million mapped fragments (FPKM). Transcripts with FPKM values of > 1 in at least one sample were defined as the threshold expression of that gene. Differential expression genes (DEGs) between two cultivars at different developmental stages (OB-S1 vs CH-S1, OB-S2 vs CH-S2, and OB-S3 vs CH-S3) was obtained using DESeq2 of TBtools-II v2.086[37]. The |log2(Fold Change)| ≥ 1 and adjusted p-value < 0.05 were set as the criteria for screening DEGs. Principal component analysis (PCA) and correlation analysis matrix were conducted using online tools of Metware Cloud (
https://cloud.metware.cn , accessed on 1 January 2024). Identification of DEGs, gene ontology (GO) analysis, and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis were performed (p value < 0.05). Combined analyses of DEG functions and expression levels were conducted with MapMan 3.6.0RC1, which is a user-driven tool that visualizes large datasets as diagrams of metabolic pathways or other processes. The function annotations of DEGs derived from MapMan were visualized as a network using Cytoscape 3.10.1.Gene cloning and sequence analysis
-
Total RNA was extracted from 'CH' flowers using a FastPure® Universal Plant Total RNA Isolation Kit (Vazyme, Nanjing, China) according to the manufacturer's instructions. First-strand cDNA was then synthesized using a HiScript® III 1st Strand cDNA Synthesis Kit (+gDNA wiper) (Vazyme, Nanjing, China) according to the manufacturer's instructions.
The motif identification of amino acid sequences for RcCH_AADC1, RcCH_PAR1, and RcCH_PAR1_Like was conducted using MEME (
https://meme-suite.org/meme/index.html ). The NCBI Conserved Domain Database (www.ncbi.nlm.nih.gov/cdd ) was used to identify domains from the amino acid sequences of RcOB_OOMT1, RcOB_OOMT2, RcCH_OOMT1 and RcCH_OOMT2. The amino acid sequences of above-mentioned candidate genes from R. chinensis, R. wichuraiana Basye's Thornless, R. rugosa, R. damascene, R. hybrida, Fragaria vesca (F. vesca), P. hybrida, S. lycopersicum, and Eucalyptus gunnii were aligned using ClustalW and the neighbor-joining (NJ) trees were generated using Molecular Evolutionary Genetics Analysis, version 7 (MEGA, version 7). The bootstrap values were obtained with 1,000 replications. The motifs and trees were drawn with Chiplot[38].Transient transformation in tobacco leaves
-
The full-length CDS of RcCH_AADC1, RcCH_PAR1, and RcCH_PAR1_Like were amplificated from the cDNA of R. chinensis 'CH' using designed primers (Supplementary Table S1). The candidate genes were inserted into vector pEAQ-HT. The pEAQ-HT vector with the target gene was introduced into A. tumefaciens strain GV3101[39]. The positive colonies were collected and re-dissolved in the buffer containing 10 mM 2-(N-morpholino) ethanesulfonic acid (MES), 10 mM MgCl2, and 200 μM acetosyringone (AS) and adjusted to OD600 of 0.8. After 2 h of incubation, the suspensions were pressure infiltrated into 4-week-old N. benthamiana leaves. After infiltration for 4 d, 1 g infected leaves were collected and placed in enclosed bottles for volatile compound analysis. The pEAQ-HT with empty vector was used as a negative control. The compound of 2-PE served as positive control.
-
The two rose cultivars used in this study showed similar phenotype of double-petal flowers, and both cultivars bloom in all seasons (Supplementary Table S2). Some differences were observed in the flower color and size that 'OB' displays larger flowers than 'CH' (Fig. 1a & b; Supplementary Table S2).
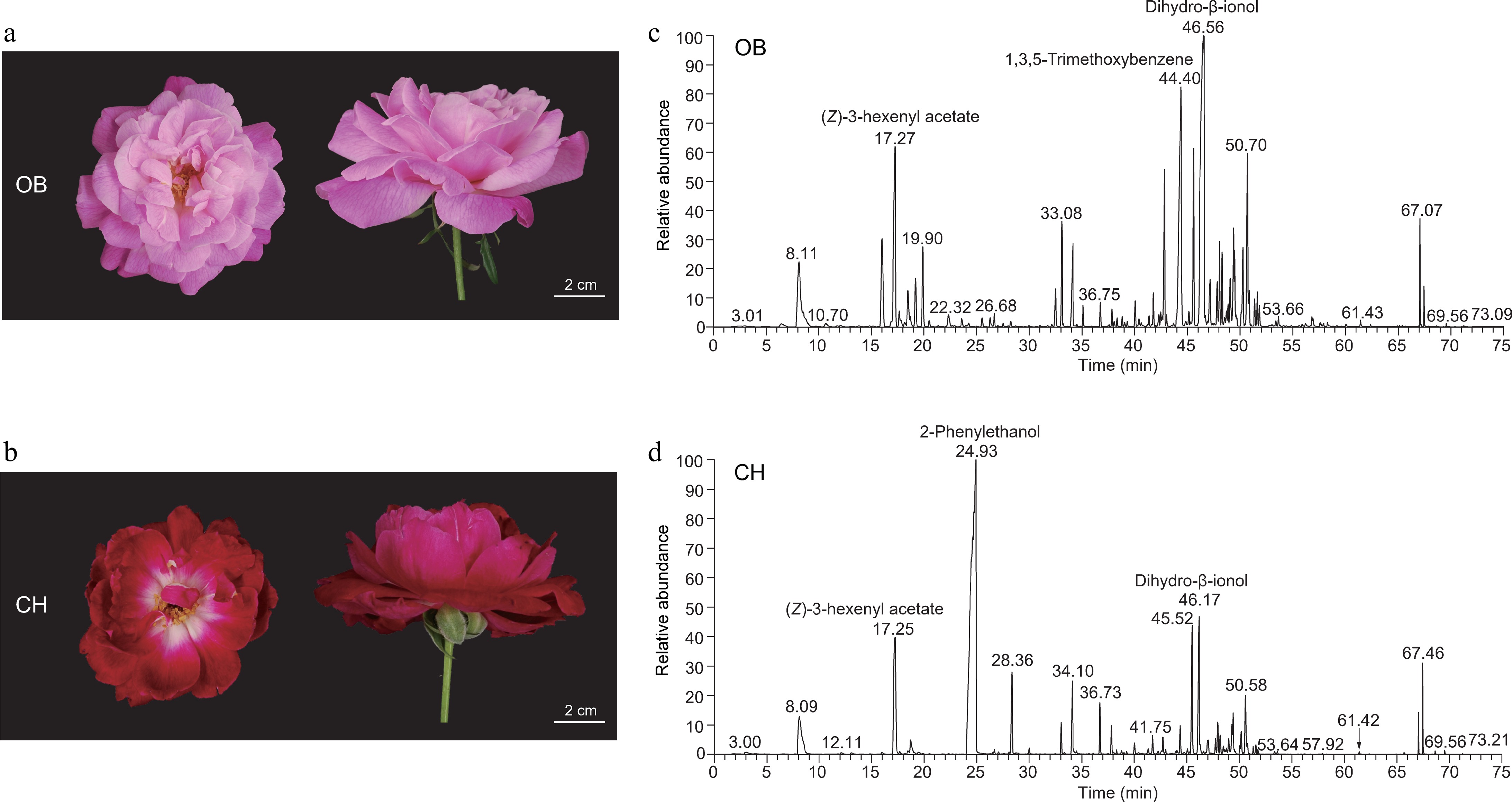
Figure 1.
Flower phenotypes and volatile compounds of 'OB' and 'CH'. (a), (b) Floral phenotypes of (a) 'OB', and (b) 'CH' at S3 (full opening stage). (c), (d) Chromatograms for the S3 petals of (c) 'OB', and (d) 'CH'. The three highest peaks were labeled with annotations in chromatograms of two cultivars.
To clarify the floral characteristics of 'OB' and 'CH', we performed GC-MS-based VOC profiling on the flower at the S3 stage of 'OB' and 'CH' (Fig. 1c, d). A total of 22 primary floral components (area > 1%) were identified either in 'OB' or 'CH', including 14 terpenoids, five fatty acid derivatives, three phenylpropanoids and their derivatives. The primary floral components of 'OB' and 'CH' accounted for 74.90% and 88.52% of total VOCs, respectively (Supplementary Table S3). Considering 22 primary floral components, terpenoids were found to be the most abundant VOC group (57%) emitted from the 'OB' flower, whereas in 'CH' flower the phenylpropanoids were the major VOC group (60%) compared with other VOC groups (Fig. 2a). Among the primary VOCs, 12 VOCs showed significant (p < 0.05) differences in their contents between two cultivars (Fig. 2b). Specifically, 2-PE and TMB were observed with the greatest variations between 'OB' and 'CH' (Fig. 2c). The content of 2-PE in 'CH' was accumulated significantly higher (p < 0.05) than in 'OB', while the trend in the content of TMB was reversed between the two cultivars (Fig. 2b).
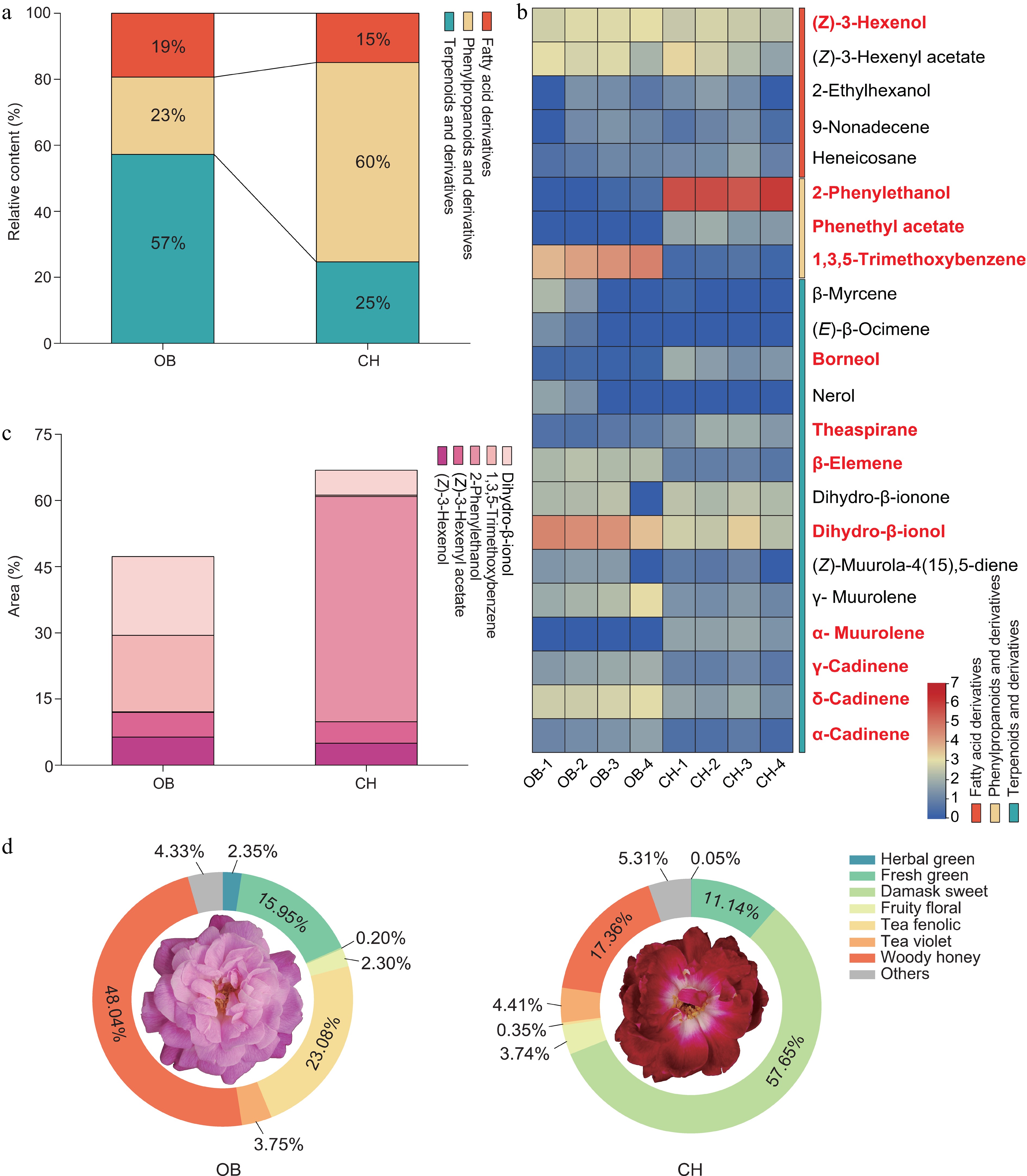
Figure 2.
The main VOCs of 'OB' and 'CH'. (a) Relative content of VOCs in different scent components of two cultivars. (b) Heatmap showed the relative content (log2-transformed peak area) of 22 major floral fragrance components. The text in bold and red indicates the significantly changed VOCs. (c) Area of the top five VOCs. (d) Classification of the aroma types of 'OB' and 'CH'.
With respect to the aroma odor, the two cultivars also exhibited a diversity of aroma types. Twenty two major VOCs have been classified into eight groups based on their fragrance descriptions following a previous report[40]. These groups consist of herbal green, fresh green, damask sweet, fruity floral, tea fenolic, tea violet, woody honey, and others (Fig. 2d). The dominant flavor profile of 'OB' flowers was woody honey, characterized by a high presence of terpenes and dihydro-β-ionol. The 'OB' flower contained another type of aroma, a tea scent, associated with TMB. In contrast, 'CH' flowers emitted a pronounced damask sweet scent predominantly composed of 2-PE, accounting for approximately 51.03% of total VOCs.
Analysis of the DEGs in R. chinensis flowers at different stages
-
To investigate the molecular basis for the metabolic differences between 'OB' and 'CH', we conducted a transcriptomic analysis of both cultivars at different developmental stages of flowers (S1−S3). PCA score plot based on gene expressions revealed a great variation between cultivars as two cultivars widely separated from each other. In addition, a notable separation was observed between S3 and the other two stages (S1−S2) in both 'OB'and 'CH' (Fig. 3a). Both PCA and correlation analysis demonstrated excellent repeatability among biological replicates (Fig. 3a, b). By comparing the gene expression profiles between two cultivars at different stages, a total of 1,899 (879 up-regulated and 1,020 down-regulated), 1,535 (702 up-regulated and 833 down-regulated), and 3,663 DEGs (1,767 up-regulated and 1,896 down-regulated) were identified in the comparisons of OB-S1 vs CH-S1, OB-S2 vs CH-S2, and OB-S3 vs CH-S3, respectively (Fig. 3c). The results of GO and KEGG enrichment analyses based on DEGs revealed the gene functions primarily in metabolism, chaperones, and folding catalysts, signal transduction, and biosynthesis of other secondary metabolites (Supplementary Fig. S1). As the flower at the S3 stage displayed the greatest variation in gene expression and VOC contents, we used MapMan to cluster the genes involved in the same metabolism using the DEGs that were associated with secondary metabolism at the S3 stage. A total of 155 genes were obtained and fed into MapMan for further analysis. Those DEGs were clustered into 18 different metabolism pathways, and they were mainly associated with flavonoids, isoprenoids, and phenylpropanoids biosynthesis, linking 52, 32, and 25 DEGs (Fig. 3d).
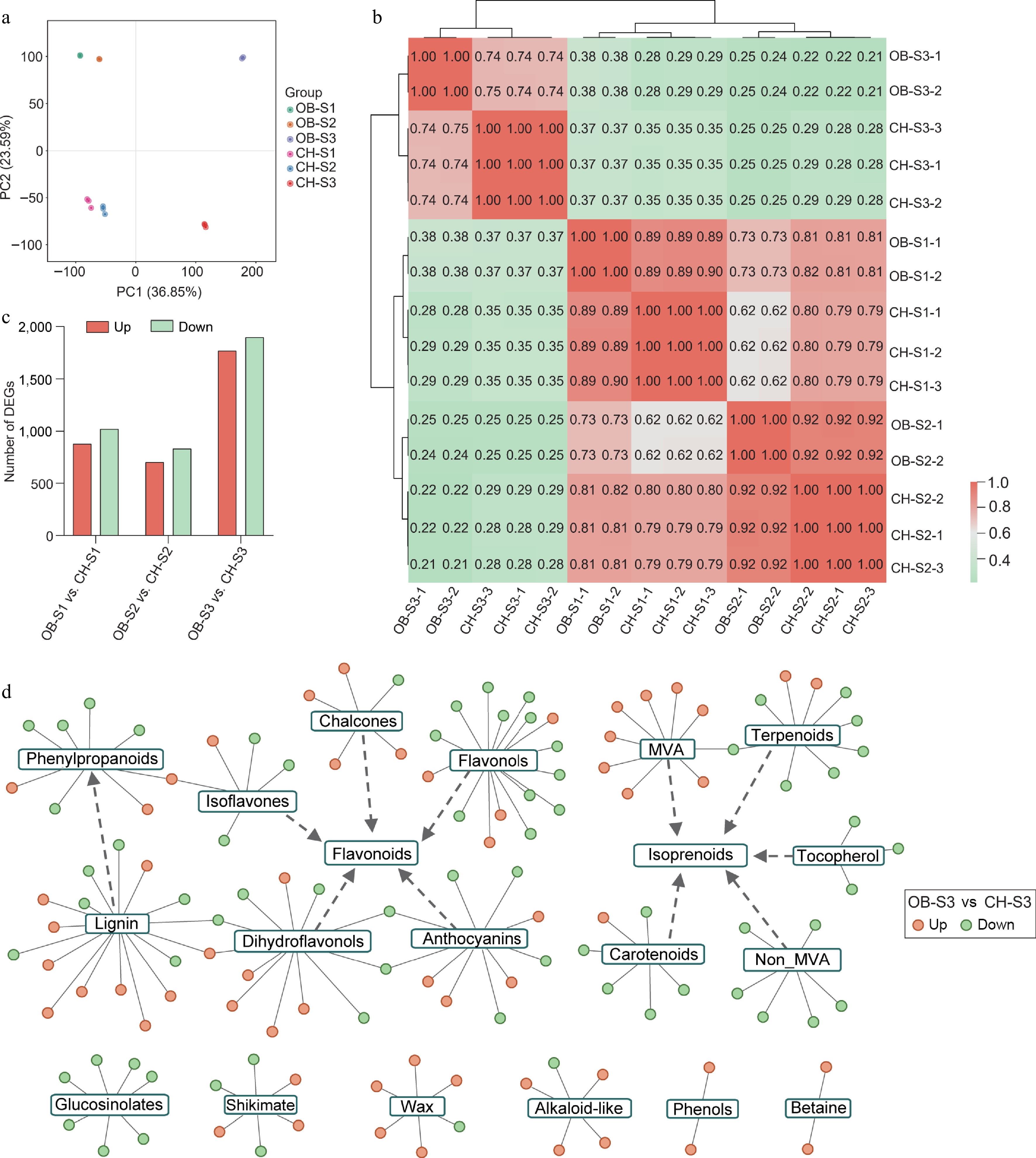
Figure 3.
Analysis of transcriptomes of 'OB' and 'CH' at S1−S3. (a) PCA showing the clustering of transcriptome of floral development stages (S1−S3) in R. chinensis petals. (b) Correlation analysis matrix among 'OB' and 'CH' at different development stages based on transcriptomes. (c) Statistics of DEGs in the comparisons of 'OB' and 'CH'. (d) Network of secondary metabolism of identified DEGs between two cultivars at the S3 stage. The orange and green cycles represent up- and down-regulated DEGs in 'OB' compared to 'CH'. The edges connect the DEGs with corresponding pathways.
Pathways analysis for floral fragrance between 'OB' and 'CH'
-
The blooming flowers (S3) of 'OB' and 'CH' were characterized by some typical floral VOCs, with TMB identified as the predominant aroma compound in 'OB' flowers and 2-PE as the key VOC in 'CH' flowers. The MapMan results indicate that the DEGs associated with phenylpropanoid pathway significantly contribute to the floral fragrance of the two cultivars (Fig. 3d). Based on these results and previous research, we identified 15 key structural genes related to phenylpropanoids biosynthesis in rose (Supplementary Table S4 & Fig. 4). There were two phloroglucinol O-methyltransferase (POMT) and four orcinol O-methyltransferases (OOMT) genes within the biosynthetic pathway of TMB, and one AAAT, four AADC, five PPDC, and two PAR genes related to the biosynthesis of 2-PE. Among them, the RcPOMT gene was highly expressed at the S1 and S2 stages, followed by a decrease at the S3 stage of two cultivars. In contrast to POMTs, the OOMTs genes showed variations in expression patterns. The OOMTs of 'OB', RcOB_OOMT1 (RcCH_v1.0_chr2.3178), and RcOB_OOMT2 (RcCH_v1.0_chr2.2828), exhibited their highest expression levels exclusively at the S3 stage, whereas the RcCH_OOMTs genes of 'CH' remained unexpressed in flowers across all three stages. The expression of RcCH_OOMT1 was observed primarily in the root and stem, whereas that of RcCH_OOMT2 was not detected in any of the six tissues. Aligning the promoter sequences of OOMTs genes between 'OB' and 'CH', we found more similarity in the promoter sequences between RcOB_OOMT1 and RcCH_OOMT1 (Supplementary Fig. S2) than in that between RcOB_OOMT2 and RcCH_OOMT2 (Supplementary Fig. S3). In addition, RcCH_OOMT2 amino acid sequence was shorter than that of three RcOOMTs and lacked the dimerization domain (Supplementary Fig. S4a).
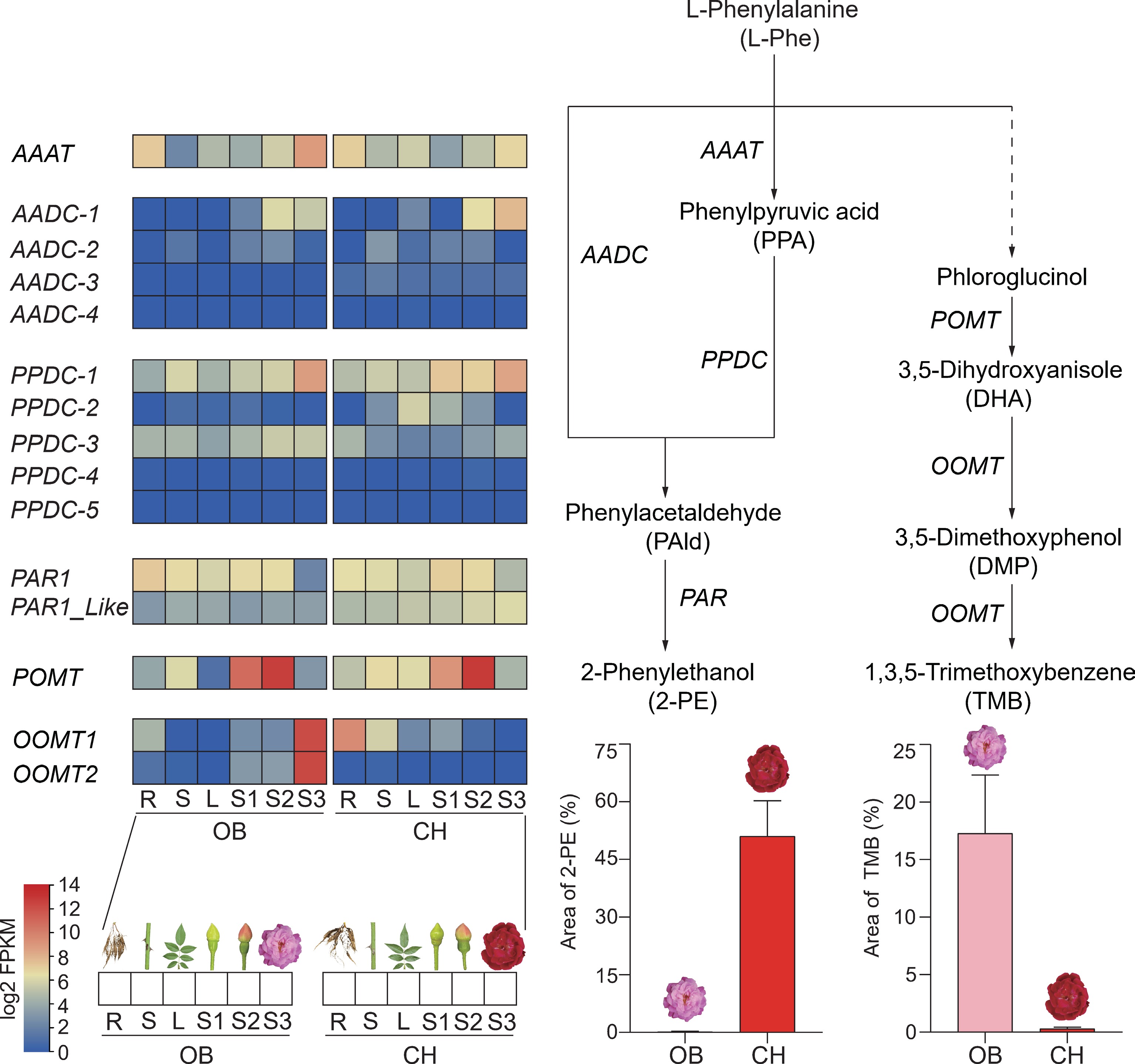
Figure 4.
Metabolomic and transcriptomic analysis of phenylpropanoid synthesis pathways in six tissues of 'OB' and 'CH'. POMT, phloroglucinol O-methyltransferase; OOMT, orcinol O-methyltransferase; AAAT, aromatic amino acid aminotransferase; AADC, aromatic amino acid decarboxylase; PPDC, phenylpyruvic acid decarboxylase; PAR, phenylacetaldehyde reductase; R, root; S, stem; L, leaf.
In the biosynthesis pathway of 2-PE, the expression level of RcOB_AAAT (RcCH_v1.0_chr5.2131) increased as the flower developed (Fig. 4). Although the expression trend of RcCH_AAAT followed a similar pattern to that of RcOB_AAAT, the expression level of RcCH_AAAT at the S3 stage was significantly lower than that of RcOB_AAAT. Among the five PPDC genes identified in the two cultivars, only one gene, PPDC-1 (RcCH_v1.0_chr3.1054), displayed notable expression levels in flowers, indicating a distinct functional role. In an alternative route of 2-PE biosynthesis, the AADC-1 gene (RcCH_v1.0_chr6.304) showed notable increases along with flower development in both cultivars, distinguishing it from the other three AADC genes identified in the two cultivars (Fig. 4).
PAR gene plays a critical role in catalyzing the reduction of PAld to 2-PE. The tissue-specific expression pattern of the RcPAR1 (RcCH_v1.0_chr5.4505) gene exhibited a striking similarity between the two cultivars, displaying a gradual decline in root, stem, and leaf, as well as during the stages of flower development. At stage S3, the expression of RcOB_PAR1 (RcCH_v1.0_chr5.4505) was decreased compared to that at the stages S1 and S2, while the expression of RcOB_PAR1 was notably inferior to that of RcCH_PAR1. The expression level of RcOB_PAR1_Like (RcCH_v1.0_chr7.2995) was found to be similar in all six tissues. RcCH_PAR1_Like consistently exhibited higher expression levels compared to RcOB_PAR1_Like in all three stages.
Identification of RcCH_AADC-1 and RcCH_PARs genes involved in the synthesis of 2-PE
-
2-PE plays a crucial role in perfumes, essential oils, and cosmetics, contributing to the reputation of rose as 'liquid gold'[41]. In this study, we conducted phylogenetic analysis and transient transformation on selected genes, RcCH_AADC-1 and RcCH_PARs, to explore the connection between these genes and 2-PE synthesis (Fig. 5 & Supplementary Fig. S4b). The AADC amino acid sequences from different species showed similar motif structure (Supplementary Fig. S4b). AADC-3 and AADC-4 were found to be clustered together in both cultivars, whereas AADC-1 and AADC-2 were not. The sequences of AADC-1 from two cultivars were found to cluster in a group with the reported RdAADC, RhAADC, and RhPAAS.
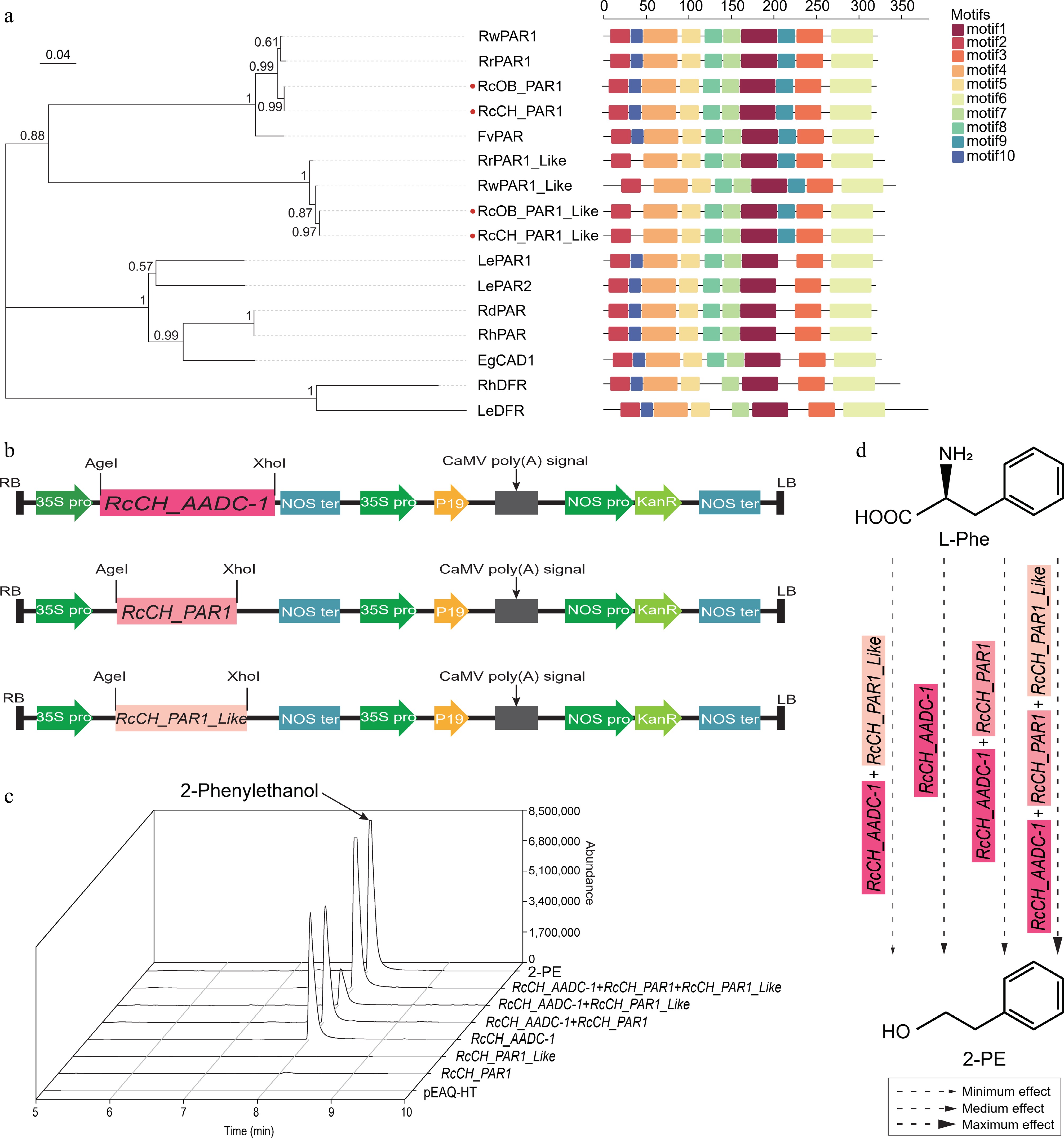
Figure 5.
The characterization of the RcCH_AADC-1 and RcCH_PARs genes. (a) Phylogenetic analysis of PARs from R. chinensis and other plants by MEGA. Protein IDs are shown in Supplementary Table S6. (b) Recombinant pEAQ-HT vectors with RcCH_AADC-1, RcCH_PAR1, and RcCH_PAR1_Like. (c) The relative contents of 2-PE in empty pEAQ-HT vector, RcCH_AADC-1, and RcCH_PARs transgenic lines. (d) The potential 2-PE synthesis pathway. Arrows of different sizes indicate the effect of RcCH_AADC-1 and RcCH_PARs on the formation of 2-PE. Thin, medium, and coarse arrows represent minimum, medium, and maximum effect.
The genes of PARs and DFRs were identified as PF01370 on the SDR website (
www.sdr-enzymes.org ) (Supplementary Table S5). The PARs amino acid sequences exhibited conserved motifs (motifs 1 and 2) that are characteristic of the SDRs superfamily, containing the cofactor binding site (TGxxxGxG) and the active site (YxxxK) in both cultivars (Supplementary Fig. S4c). By constructing phylogenetic trees, four PARs of 'OB' and 'CH' were divided into two clades (PAR1 and 1_Like clades), and orthologous genes from different cultivars clustered into one clade (Fig. 5a). Among them, we found that the amino acid sequences of RcOB_PAR1 and RcCH_PAR1 are identical, as well as RcOB_PAR1_Like and RcCH_PAR1_Like. While the similarity of amino acid sequences between the two PARs from the same cultivar was found to be 73.65%.To clarify the functional characteristics of the highly expressed RcCH_AADC-1 and RcCH_PARs, these genes were overexpressed by the transient transformation of the vectors into tobacco via A. tumefaciens (Fig. 5b). The levels of 2-PE in transgenic tobacco leaves were detected by GC-MS. We found that overexpression of RcCH_AADC-1 can promote the accumulation of 2-PE (Fig. 5c). Furthermore, the co-expression of RcCH_AADC-1 and RcCH_PAR1_Like exhibited the lowest level of 2-PE, while the co-expression of RcCH_AADC-1 and two RcCH_PARs resulted in higher content of 2-PE than that of RcCH_AADC-1 overexpression. In addition, either RcCH_PAR1 or RcCH_PAR1_Like was individually transformed into tobacco plants, we did not detect any significant increase in the level of 2-PE compared with that of the empty vector. Consequently, we proposed a potential model of the pathways governing the biosynthesis of 2-PE that co-expressing RcCH_PARs with RcCH_AADC-1 presented different regulations in the biosynthesis of 2-PE (Fig. 5d).
-
This study investigated two famous R. chinensis cultivars 'OB' and 'CH' for an assessment of the reason for differences in floral scent. Because the phenotypic divergence between these two cultivars seemed likely to provide good material for research on floral fragrances. The total content of volatile compounds in 'OB' was significantly lower than that in 'CH', 'OB' possessed more types of terpenoids, and occupied 57% of the total volatiles. Our results indicated that TMB and 2-PE are the most representative volatile constituents in 'OB' and 'CH' flowers, respectively, which is consistent with previous reports[4,25]. Combining the transcriptomic and transformation analysis, our findings revealed that OOMTs play a crucial role in TMB synthesis and AADC-1 serves as a pivotal regulator in 2-PE synthesis.
Sequence differences of RcOOMTs gene
-
TMB is a crucial component of the 'Chinese rose odor' as a phenylpropanoid compound[23,42]. This volatile compound is a potent sedative and has been utilized as an ingredient in cosmetics[42]. Interestingly, high levels of TMB were specifically detected in the Chinenses section of the Rosa subgenus, while trace amounts of TMB were found in other sections like Gymnocarpae, Gallicanae, Synstylae, and Banksianae[43]. In this study, there were higher levels of TMB in 'OB' than in 'CH' flowers (Fig. 2b), in line with previous research findings[6,20]. This indicates that variations in TMB contents are present not only between different sections in rosa, but also among Chinenses section.
A phylogenetic analysis of the OOMT gene family in five Chinese roses and 13 Eurasian rose species revealed that only Chinese roses possess both the OOMT1 and OOMT2 genes[4]. In the present study, both OOMT1 and OOMT2 were identified in 'OB' and 'CH' flowers (Fig. 4). The genes RcOB_OOMT1, RcOB_OOMT2, and RcCH_OOMT1 contain the dimerization and AdoMet_MTases superfamily domain, and their expression levels changed along with the flower development in both cultivars. RcCH_OOMT2 only possesses a partial segment of the AdoMet_MTases superfamily domain, and its expression level remained (Fig. 4 & Supplementary Fig. S4a). This suggests that two domains are essential for the function of OOMT genes. The OOMT2 gene may be critical for the biosynthesis of TMB in R. chinensis.
Previous research has demonstrated European roses expressed OOMT genes in their stamens rather than in flower petals. In contrast, levels of OOMT1 and OOMT2 were higher in the petals of Chinese roses instead of in other tissues[44]. As the key genes in the biosynthesis of TMB, the upregulation of OOMTs genes can promote the synthesis of TMB in roses. In our study, RcOB_OOMTs were specifically expressed in full blooming 'OB' petals (S3). While in 'CH', minimal levels of RcCH_OOMT1 were observed in S1 and S2 petals, RcCH_OOMT2 showed no detectable mRNA transcripts in petals (Fig. 4). The finding suggests that the development of TMB biosynthesis in Chinese roses relies on the concurrent expression of OOMT1 and OOMT2, accompanied by an upregulation of OOMT expression in petals.
Functional differentiation of RcPARs genes
-
The 2-PE is a prevalent scent compound abundantly emitted by rose flowers, contributing to its characteristic rose-like scent[25]. As the predominant component commonly present in perfumes and essential oils, the 2-PE is usually extracted from the Damask rose[20]. Universally, L-Phe is gradually converted into 2-PE through the catalytic action of AADC and PAR[24]. After overexpressing RcCHAADC-1 in tobacco leaves, a high level of 2-PE was noticed (Fig. 5), which reinforces the previous findings that modulating AADC expression can influence the synthesis of phenylalanine-based aroma volatiles in R. rugosa and R. hybrida[29,45]. A previous study showed that the reduction of PAld to 2-PE can be catalyzed by PAR or alcohol dehydrogenase (ADH)[26]. Therefore, it cannot be ruled out that endogenous dehydrogenase (e.g., ADH) in tobacco may replace PAR in the reduction reaction. The upregulation of RcCH_AADC-1 gene expression might serve as a key factor in 2-PE synthesis in roses.
PAR functions as the key enzyme in the last step of 2-PE biosynthesis[26]. PAR gene has one copy in R. rugosa 'Tanghong'[41], R. damascene[17], and R. hybrida 'Yves Piaget'[46]. The expression of RdPAR, RrPAR, and LePARs increased consistently from the budding stage to the half-opening stage, and decreased at the full-opening stage[17,28,41]. In our study, two PAR genes were identified in both cultivars, one was PAR1, whose expression pattern was consistent with previous reports[17,28,41]. The other one is PAR1_Like, whose expression pattern was not. In 'OB', RcOB_PAR1_Like was lowly expressed at the three stages of petal, while RcCH_PAR1_Like expression increased from S1 to S3 in 'CH' (Fig. 4). In the presence of nicotinamide adenine dinucleotide phosphate (NADPH) or nicotinamide adenine dinucleotide (NADH), PAR efficiently converted PAld to 2-PE[17,26]. This may explain to why neither overexpression of RcPAR1 nor RcPAR1_Like independently regulated 2-PE synthesis in this study. It is also possible that the PAld contents in tobacco may not be sufficient to synthesize more 2-PE. In addition, the analysis of sequences, evolutionary trees, and transient transformation of RcPAR1 and RcPAR1_Like revealed the functional diversity in these genes. When one RcPAR copy was co-expressed with RcAADC-1, it exhibited significantly lower levels of 2-PE compared to the situation where two RcPARs were co-expressed with RcAADC-1. Hence, the functional differentiation of RcPAR1 and RcPAR1_Like, along with their involvement in the regulatory mechanisms of 2-PE formation, requires further investigation.
-
This study investigated the floral fragrances in the blooming flower of two R. chinensis cultivars. The main specific focus on essential volatile floral components were identified: TMB in 'OB' and 2-PE in 'CH'. In addition, 15 key genes involved in the biosynthesis of phenylpropanoid were identified in R. chinensis flowers by analysis of transcriptome data. For example, RcOOMTs are involved in the synthesis of TMB, RcPARs and RcAADC-1 are involved in the synthesis of 2-PE. Transgenic experiments suggested that RcAADC-1 gene acts as a pivotal regulator in the biosynthesis of 2-PE. These findings provide valuable insight for practical production of rose essential oils.
This work was supported in part by the Chinese Academy of Agricultural Sciences Elite Youth Program to Zhiqiang Wu; and this work was also supported by funding from the Scientific Research Foundation for Principle Investigator, Kunpeng Institute of Modern Agriculture at Foshan (KIMA-QD2022004); the Funding of Major Scientific Research Tasks, Kunpeng Institute of Modern Agriculture at Foshan (KIMA-ZDKY2022004); and Foshan self-funded scientific and technological innovation projects (2320001007191). We thank members of the Wu laboratory. We thank Prof. Li Wei (Agricultural Genomics Institute at Shenzhen, Chinese Academy of Agricultural Sciences) for providing the pEAQ-HT vector.
-
The authors confirm contribution to the paper as follows: study conception: Zhang X, Wu Z; methodology: Luo K, Song C, Huang R, Leng L; investigation: Luo K; formal analysis: Luo K, Song C, Liao X; data curation: Song C, Yang Y, Leng L, Gu C; validation: Bao T; writing - original draft: Luo K, Song C; writing - review & editing: Luo K, Song C, Bao T, Lin S, Zhang X, Wu Z; supervision: Zhang X, Wu Z; funding acquisition, project administration: Wu Z. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Kaiqing Luo, Chao Song
- Supplementary Table S1 Primer sequences used for constructing vectors.
- Supplementary Table S2 Phenotypic traits and collection locations of two tested roses.
- Supplementary Table S3 Major Volatile compounds of the two R. chinensis cultivars.
- Supplementary Table S4 The genes expression of metabolic pathways for floral scent in 'OB' and 'CH'.
- Supplementary Table S5 SDR family prediction of PARs proteins from R. chinensis and other plants.
- Supplementary Table S6 Protein IDs of PARs and AADCs from different species in the evolutionary tree.
- Supplementary Fig. S1 GO terms and KEGG pathways for differentially expressed genes (DEGs). (a)−(c) GO terms for comparisons of samples (OB-S1 vs. CH-S1, OB-S2 vs. CH-S2 and OB-S3 vs. CH-S3). (d)−(f) KEGG pathways for comparisons of samples (OB-S1 vs. CH-S1, OB-S2 vs. CH-S2 and OB-S3 vs. CH-S3).
- Supplementary Fig. S2 Alignment of promoter sequences of RcOOMT1 in R. chinensis 'OB' and 'CH'.
- Supplementary Fig. S3 Alignment of promoter sequences of RcOOMT2 in R. chinensis 'OB' and 'CH'.
- Supplementary Fig. S4 OOMTs, AADCs, and PARs amino acid sequences analysis. (a) Domain analysis of OOMTs amino acid sequences from R. chinensis. The green box indicates the dimerization domain. The orange boxes represent the AdoMet_MTases superfamily domain. (b) Phylogenetic analysis of AADCs amino acid sequences from R. chinensis and other plants. Protein IDs were shown in Table S6. (c) Motif analysis of PARs amino acid sequences from R. chinensis and other plants.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Luo K, Song C, Bao T, Huang R, Lin S, et al. 2025. GC-MS metabolite and transcriptome analyses reveal the differences of volatile synthesis and gene expression profiling between two Chinese rose cultivars. Ornamental Plant Research 5: e009 doi: 10.48130/opr-0025-0003 

GC-MS metabolite and transcriptome analyses reveal the differences of volatile synthesis and gene expression profiling between two Chinese rose cultivars
- Received: 03 September 2024
- Revised: 21 November 2024
- Accepted: 25 December 2024
- Published online: 04 March 2025
Abstract: Floral fragrance is an important ornamental trait of horticultural crops. However, research on the physiological and molecular biology of the floral volatile compounds in Chinese rose is limited. This study conducted metabolomic and transcriptomic analyses on the floral volatile compounds of Rosa chinensis (R. chinensis) 'Old Blush' ('OB') and R. chinensis 'Chilong Hanzhu' ('CH'). A total of 22 major volatile organic compounds (VOCs) were detected in the blooming flowers of 'OB' and 'CH'. The main VOCs emitted from 'OB' petals was 1,3,5-trimethoxybenzene (TMB), while that from 'CH' petals was 2-phenylethanol (2-PE). The GO and KEGG enrichment on differentially expressed genes (DEGs) revealed the essential role of phenylpropanoid pathway in the difference of floral fragrance between two cultivars, and a total of 15 key genes involved in the biosynthesis of phenylpropanoid were identified in R. chinensis flowers. The overexpression of RcCH_AADC-1 in transgenic tobacco plants led to the accumulation of 2-PE. In contrast, no significant change in the level of 2-PE was observed after the transient transformation of RcCH_PAR1 or RcCH_PAR1_Like into tobacco plants. These findings provide valuable insight for enhancing the accumulation of 2-PE in R. chinensis.
-
Key words:
- Rosa chinensis /
- Volatile organic compounds (VOCs) /
- Phenylpropanoid pathway /
- Genes /
- 2-PE