









-
Seed dormancy is an evolutionary adaptation that allows plants to withstand unfavorable conditions[1]. Some plants in nature may not germinate under suitable conditions due to their complex dormancy mechanism, thus threatening their survival[2]. Hence, comprehending the underlying mechanisms governing seed dormancy release holds paramount importance. To date, the precise regulatory mechanisms involved in seed dormancy release remain elusive. Early investigations have demonstrated that the dormancy of plant seed is a complex characteristic shaped by various internal and external elements[3]. Genes associated with seed dormancy serves as a pivotal components in the entire process spanning throughout the seed-to-seedling transition, serving as crucial internal factors contributing to the alleviation of seed dormancy[4]. Certain genes modulate seed dormancy through their differential expression in diverse environmental conditions. In contrast, others regulate seed dormancy by actively participating in the anabolic processes or signal transduction pathways of plant hormones associated with dormancy[5]. Recently, DOG1 has emerged as a pivotal gene in modulating dormancy in the model plant Arabidopsis, garnering increasing attention from scholars in this field. However, the function of this gene in P. lactiflora seeds remains poorly elucidated.
The heme-binding protein DOG1, initially discovered in A. thaliana, constitutes essential elements in the process of the dormancy of plant seed, particularly temperature-dependent dormancy induction[6−8]. Extensive investigations have been conducted to elucidate the structural characteristics of DOG1. In A. thaliana, DOG1 comprises three exons and two introns, with the 2nd intron undergoing alternative splicing to generate five transcript variants[9]. Among these variants, the translation of β, γ, and ε transcripts yields identical proteins, thus resulting in only three distinct protein isomers being produced[10]. All DOG1 isoforms demonstrate intrinsic self-association capabilities crucial for functional enhancement, as impaired self-binding maintains protein abundance but results in dormancy deficiency. Consequently, dimer formation within different DOG1 isomers becomes indispensable for the precise regulation of DOG1 functionality[11].
There are five DOG1 homologs identified in the A. thaliana genome, namely AtDOGL1, AtDOGL2, AtDOGL3, AtDOGL4, and AtDOGL5[12,13]. AtDOGL1 to AtDOGL4 are located adjacent to each other on chromosome 4, while AtDOGL5 is located on chromosome 3. AtDOGL1, AtDOGL2, and AtDOGL3 are relatively similar to AtDOG1, while the amino acid sequences of AtDOGL4 and AtDOGL5 are not very similar to those of AtDOG1[12,14]. Similar to A. thaliana, the DOG1 gene has been found in some dicotyledonous and many monocotyledonous plants[15]. Among the dicotyledonous plants studied, the amino acid sequences of DOG1 are highly similar[16]. DOG1 genes are also found in many monocotyledonous plants, such as barley, wheat, rice, maize, sorghum, and Brachypodium distachyon[17−19].
DOG1 is involved in seed dormancy-related processes. The expression of DOG1 determines the depth of the dormancy of plant seed[11,20,21]. The expression of DOG1 is specifically enriched in seeds, exhibiting a gradual increase in its level during seed embryo maturation[11]. Freshly harvested seeds exhibited increased dormancy, linked to higher DOG1 gene expression, whereas germinated seeds exhibited reduced DOG1 transcriptional activity[22]. In recent years, it has been discovered that DOG1 exerts regulatory control over seed dormancy and germination through diverse pathways, with multiple mechanisms on the extensively investigated involvement of plant hormones[23,24]. Scholars have discovered that the simultaneous presence of ABA and DOG1 is essential for inducing dormancy, as the absence of either regulatory factor results in a complete loss of dormancy[8,25]. Additionally, DOG1 also regulates germination by inhibiting GA-responsive genes involved in cell wall modification[26,27]. Moreover, DOG1 interacts with ethylene signaling to control seed dormancy and associated physiological responses[15].
P. lactiflora is a perennial herbaceous plant with important landscape, pharmaceutical, and economic values and is distributed throughout the world[28]. P. lactiflora seeds exhibit unique dormancy characteristics, typically requiring 3−5 years from germination to flowering under natural conditions. However, traditional sowing methods for reproduction are time-consuming and inefficient, posing challenges in meeting the demands of commercial production[29,30]. Based on this situation, understanding the mechanism of seed dormancy relief can improve the artificial breeding efficiency of the peony, accelerate its commercial production, and provide theoretical support for genotype development[31]. However, research on dormancy-breaking methods in P. lactiflora has mainly concentrated on physical scarification, cold stratification, and hormone treatments, with limited investigation into the molecular basis of seed dormancy in peonies. Here, we obtained significantly differentially expressed PlDOG1 from our previous RNA-sequencing data[32]. We cloned PlDOG1, analyzed it bioinformatically, and studied its role in regulating seed germination with A. thaliana and P. lactiflora. In addition, our results suggest that P. lactiflora auxin response factor PlARF2, as a transcription factor, can bind to the promoter of PlDOG1, thereby affecting seed dormancy release. These findings provide fundamental insights into the regulatory mechanisms of DOG1 and auxin in seed dormancy control and elucidate the related regulatory mechanism of peony seed germination from the perspective of molecular biology.
-
In August 2022, seeds of P. lactiflora cultivar 'Fen Yu Nu' × 'Fen Yu Lou' were collected from the germplasm repository at Shenyang Agricultural University, located in Shenyang, Liaoning Province, China. The SALK_000867 mutant in Arabidopsis Col-0 ecotype was acquired from the Arabidopsis Biological Resource Center. Seeds of wild type (WT) and mutant A. thaliana were sown following method reported previously[30].
Acquisition of RNA and cDNA, and procedural methodology for qRT-PCR
-
RNA was isolated from the hybrid seeds of P. lactiflora with the Easy spin Plus Complex Plant RNA Kit (Aidlab, Beijing, China). For cloning, cDNA was generated using the PrimeScript™ II 1st Strand cDNA Synthesis Kit (Takara, Dalian, China). Meanwhile, for qRT-PCR, cDNA was synthesized by the PrimeScript™ RT Master Mix kit (Takara, Dalian, China). Primers for qRT-PCR were devised with Primer Premier 5.0 software, taking the coding sequence (CDS) of PlDOG1 as the basis. In qRT-PCR, PlACTIN (GenBank number JN105299.1) served as the reference gene[33]. The qRT-PCR was carried out using the TB Green® Premix Ex Taq™ II (Tli RNaseH Plus) kit from Takara (Dalian, China). All the tests were repeated three times, and the gene expression levels were quantified using the comparative 2−ΔΔCᴛ method. The obtained data were analyzed by IBM SPSS Statistics 26 software for standard deviation and significance, and the images were created using GraphPad Prism 8.0.
Sequence acquisition and bioinformatics analysis
-
We compared our transcriptomic data[32] with the CDS sequence of AtDOG1 in A. thaliana, and obtained the homologous gene of AtDOG1 in P. lactiflora and named it PlDOG1. Then, we used the NCBI ORF Finder website (
www.ncbi.nlm.nih.gov/orffinder ) to produce its amino acid sequence. Using MEGA 7.0 software and evolview (www.evolgenius.info//evolview-v2 ) online software to generate a PlDOG1 protein phylogenetic tree. The conserved domains of proteins were analyzed using the NCBI structure tool (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ). The physical and chemical properties of the protein were analyzed using Expasy Prot-Param (https://web.expasy.org/protparam ). PlDOG1-specific primers were designed with Primer Premier 5.0 according to its CDS, followed by PCR amplification using 2×Taq Plus MasterMix (Aidlab, Beijing, China) for cDNA cloning.Ectopic expression and genetic complementation of PlDOG1 in A. thaliana
-
To acquire overexpressing transformants and gain-of-function mutants, the coding sequence of PlDOG1 was cloned into pCAMBIA1301 and subsequently introduced into Agrobacterium GV3101. The recombinant vector was introduced into both WT and mutant plants via Agrobacterium-mediated transformation. Briefly, the construct was transferred into Agrobacterium GV3101 and cultured at 28 °C with 200 rpm shaking until reaching OD600 was 1.0. Healthy Arabidopsis plants at the flowering stage were selected for floral dip transformation, where unopened flower buds were immersed in the bacterial suspension. After 48-h dark incubation, plants were grown under normal conditions for seed production. T0 seeds were collected and germinated on 1/2 MS medium supplemented with 50 mg/L kanamycin for transgenic selection. Homozygous lines were identified through successive screening until T3 generation. All seed types (wt, mutant, overexpression, and complementation lines) were subsequently germinated on 1/2 MS medium for phenotypic analysis. Germination timing and rates were recorded, with data analysis performed using Microsoft Excel 2019 and graphical representations generated through GraphPad Prism 8.0.
Transient expression of P. lactiflora seeds
-
The sources and conditions of P. lactiflora seeds used in this part of the experiment are the same as described above. The overexpressed vector obtained in the previous description was transferred to Agrobacterium EHA105 to prepare the infected solution. To inhibit the expression of PlDOG1, 248 bp sequence was cloned according to the CDS of PlDOG1, and cloned into pTRV2 vector to develop a transcriptionally silent fusion vector. The fusion constructs were subsequently introduced into Agrobacterium EHA105. After that, the infected solution was prepared by combining Agrobacterium harboring pTRV1 with the fusion vector and the empty vector in a 1:1 ratio. The seeds also needed to be treated while the infected solution was prepared. First, the seeds were washed followed by a 48-h water immersion, then seeds were surface-sterilized sequentially with ethanol (75% concentration) for 30 s and 0.1% (w/v) mercuric chloride solution for 5.5 min. Subsequently, the seeds were rinsed four times with distilled water (1 min per wash). Post-sterilization, they were trimmed to 3 mm3 pieces and punctured using a needle in a 1 mL syringe. These punctured seeds underwent Agrobacterium-mediated transformation under negative pressure for 3 min, followed by a 15-min immersion in the bacterial suspension. Excess solution was blotted with filter paper, and the seeds were then subjected to a 36-h dark incubation on MS medium. After dark treatment, the transiently transformed seeds were rinsed for 25 min in sterile water containing 250 mg/L cefotaxime. Subsequently, seed embryos were excised and transferred to MS medium supplemented with 0.5 mg/L GA3 for further culture. The germination phenotype was observed, and the germination time, and germination rate were calculated. The instantaneous conversion effect was detected by qRT-PCR. For each type, 30 embryos were inoculated, with three independent biological replicates prepared.
Cloning of PlDOG1 promoter and analysis of promoter activity
-
Since P. lactiflora lacks genomic reference, we obtained the promoter of PlDOG1 by the chromosome walking method. Firstly, the plant genome DNA Rapid extraction kit (Aidlab, Beijing, China) was used to extract the genomic DNA of P. lactiflora. Then, the amplification primer was designed with reference to the CDS sequence of PlDOG1 gene, and the DNA sequence obtained in the above step was used as the template for amplification. Next, the amplified sequence was compared with the PlDOG1 sequence in the existing transcriptome data, and the primer sequence of the chromosome step method was designed with reference to the sequence without introns. In accordance with the user manual for the Genome Walking Kit (Takara, China), the PlDOG1 promoter sequence was reverse cloned via triple nested PCR using the specifically designed primer sequences. The promoter regions were examined for cis-acting elements using the PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html ) and NewPLACE (www.dna.affrc.go.jp/PLACE ) online databases.To investigate the promoter activity of the obtained promoter, we selectively deleted the 5' end of the acquired promoter sequence according to the location of cis-acting elements within it, generating two distinct 5' end deletion fragments. Subsequently, these two fragments and the promoter sequence were cloned into the empty vector pCAMBIA1301, substituting the native promoter, followed by transferring into the Agrobacterium EHA105 strain. As a control, the empty vector was also employed. Then, the Agrobacterium suspension was infiltrated into healthy tobacco leaves (4-week-old), followed by a 36-h incubation in darkness. After culture, different leaves were treated with GUS Stain Kit (RealJumes, Beijing, China), followed by decolorization with 95% ethanol. The activity of the promoter was determined by observing the staining of each leaf after decolorization.
Y1H, EMSA, and Dual-LUC
-
For the Y1H, a 300 bp sequence containing RY motif 'CATGCAT' in the obtained promoter sequence was cloned into the pHIS2 Y1H vector and transformed into the Y187 yeast strain with the fusion vector pGADT7-PlARF2 preserved in our laboratory. Positive clones were further identified by continuous dilution (1/1, 1/10, 1/100, 1/1000) of yeast cells on triple dropout growth medium without Histidine, Leucine, and Tryptophan (TDO) containing 50 mM 3-aminotriazole (3-AT). Transformants grown on double dropout medium lacking Leucine and Tryptophan (DDO) served as growth controls.
For the EMSA, the 300 bp promoter sequence used for the Y1H assay was selected as the competitive probe, and the competitive probe was labeled with biotin as the labeled probe. Labeling of the probe with biotin was completed by Shenggong Biotechnology Co., LTD (Shanghai, China). Before initiating the probe-protein binding reaction, ddH2O was added to both probes and diluted to 1, 1/10, and 1/100 times the original concentration for reserve. Meanwhile, the PlARF2 protein was induced in Escherichia coli by prokaryotic expression method. The CDS of PlARF2 was first inserted into the pGEX6p vector and subsequently introduced into E. coli Rosetta (DE3) cells. Fusion protein expression was induced with 0.4 mM IPTG for 16 h. The protein was then purified using a GST-tag protein purification kit (Beyotime Biotechnology, Shanghai, China). Interactions between the PlARF2 protein and the probe were analyzed through EMSA performed with an EMSA kit (Beyotime Biotechnology, Shanghai, China).
For the Dual-LUC, the PlDOG1 promoter was cloned into the pGREENII 0800-LUC vector, and the CDS of PlARF2 was inserted into the pGREENII 62-SK vector. Both fusion constructs were introduced into Agrobacterium GV3101 (pSoup) and cultured following the protocol provided with the Dual-LUC kit (Shenggong Biotechnology, Shanghai, China). The bacterial suspensions were then infiltrated into healthy tobacco leaves. Relative LUC/REN activity was measured using a Thermo Scientific Varioskan®Flash system (Thermo Fisher Scientific, Massachusetts, USA) and the Dual-LUC kit (Shenggong Biotechnology, Shanghai, China). LUC signals were visualized and saved using a plant cell imaging system (NightSHADE LB985, Berthold, Germany).
Primer sequences
-
The primers involved in the present study are shown in Supplementary Table S1.
Statistical analyses
-
All experiments in this study were conducted with three biological replicates for statistical analysis. Data were processed using IBM SPSS Statistics 26. The significance of differences was evaluated through one-way analysis of variance (ANOVA) followed by Duncan's post hoc test. Graphical representations, including bar and line charts, were generated using GraphPad Prism 8.0, with error bars indicating standard deviation (SD).
-
The CDS of PlDOG1 was cloned and submitted to GenBank with the accession number PP158216. The PlDOG1 gene contains a 786 bp open reading frame (ORF), encoding a 261-amino acid protein with a predicted molecular weight of 29.72 kDa. Conserved domain analysis indicated that PlDOG1 contains a highly conserved DOG1 superfamily domain, commonly found in plants, indicating its high specificity and involvement in seed dormancy regulation (Supplementary Fig. S1). A phylogenetic tree was generated to analyze the evolutionary relationships of PlDOG1 with DOG1 proteins from diverse plant species, demonstrating a close evolutionary affinity between PlDOG1 and the DOG1 protein of Papaver somniferum (Fig. 1).
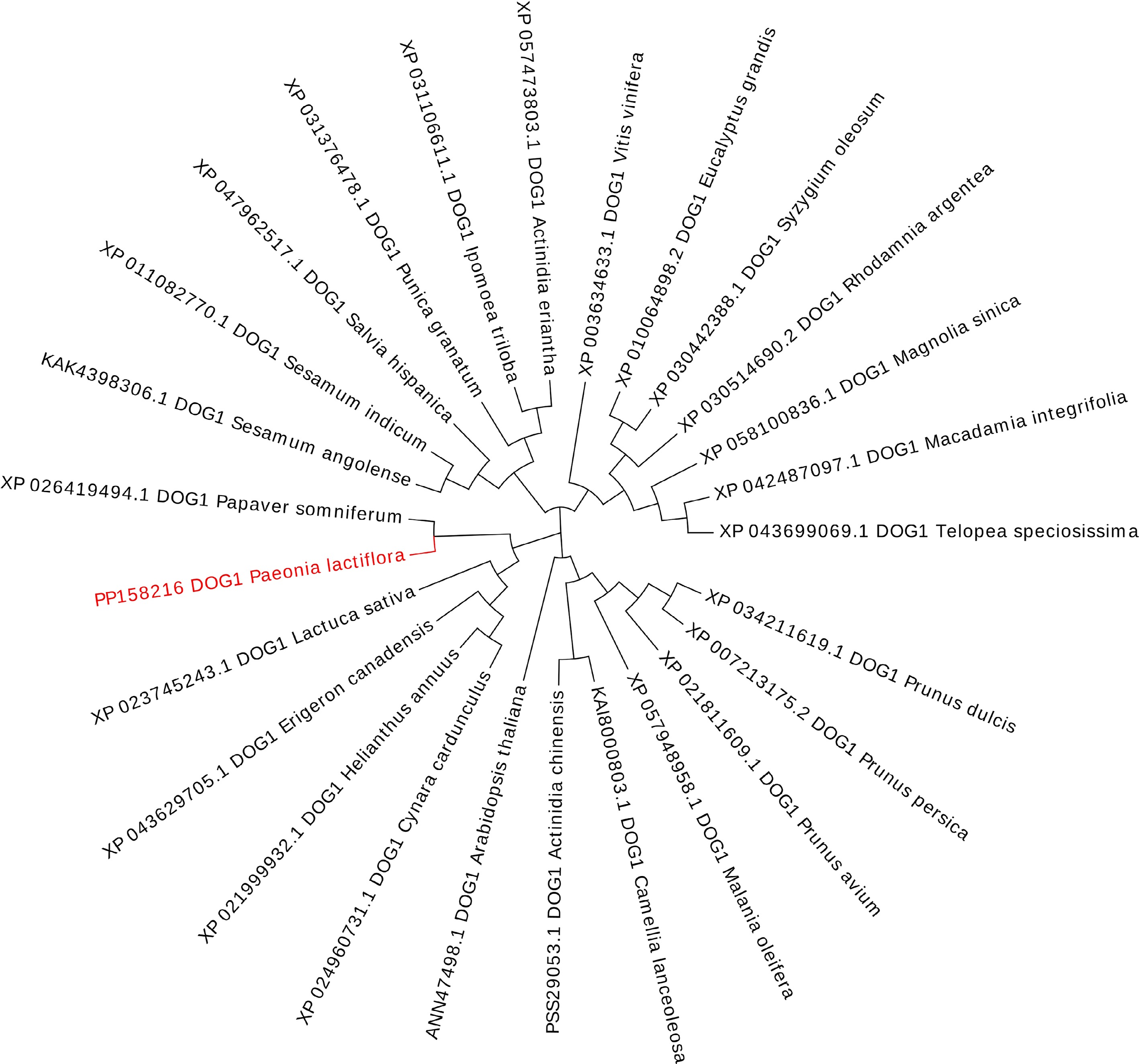
Figure 1.
A phylogenetic tree of PlDOG1 and DOG1 homologs from various plant species. The tree was generated by MEGA 7.0 and evolview software. The robustness of the tree branches was evaluated through 1,000 bootstrap replicates, and the corresponding bootstrap values were determined.
Dormancy of A. thaliana seeds was enhanced by the overexpression of PlDOG1
-
The results of seed germination observation revealed that all four types of seeds initiated germination 24 h after sowing, albeit with varying speed. The majority of WT seeds exhibited germination between 24−36 h, achieving 80% and 90% germination rates at 48 and 72 h, respectively. The results revealed accelerated seed germination in mutants relative to WT, showing significant germination between 24−36 h and surpassing an 80% germination rate at the end of the first two days. However, PlDOG1-OE and PlDOG1-FC seeds demonstrated delayed germination as most did not initiate until after 36 h, achieving approximately an 80% germination rate by the end of three days (Fig. 2a). These findings suggest that there are variations in seed germination rates among different types of seeds, which can be readily observed after 60 h of culture. At this time point, almost all WT and mutant seeds had successfully undergone germination, however, a small proportion of PlDOG1-OE and PlDOG1-FC seeds either remained ungerminated or had just begun to sprout (Fig. 2b). Consequently, these outcomes suggest that PlDOG1 may extend A. thaliana seed's dormancy period while further implying its potential negative regulatory role in plant seed germination.
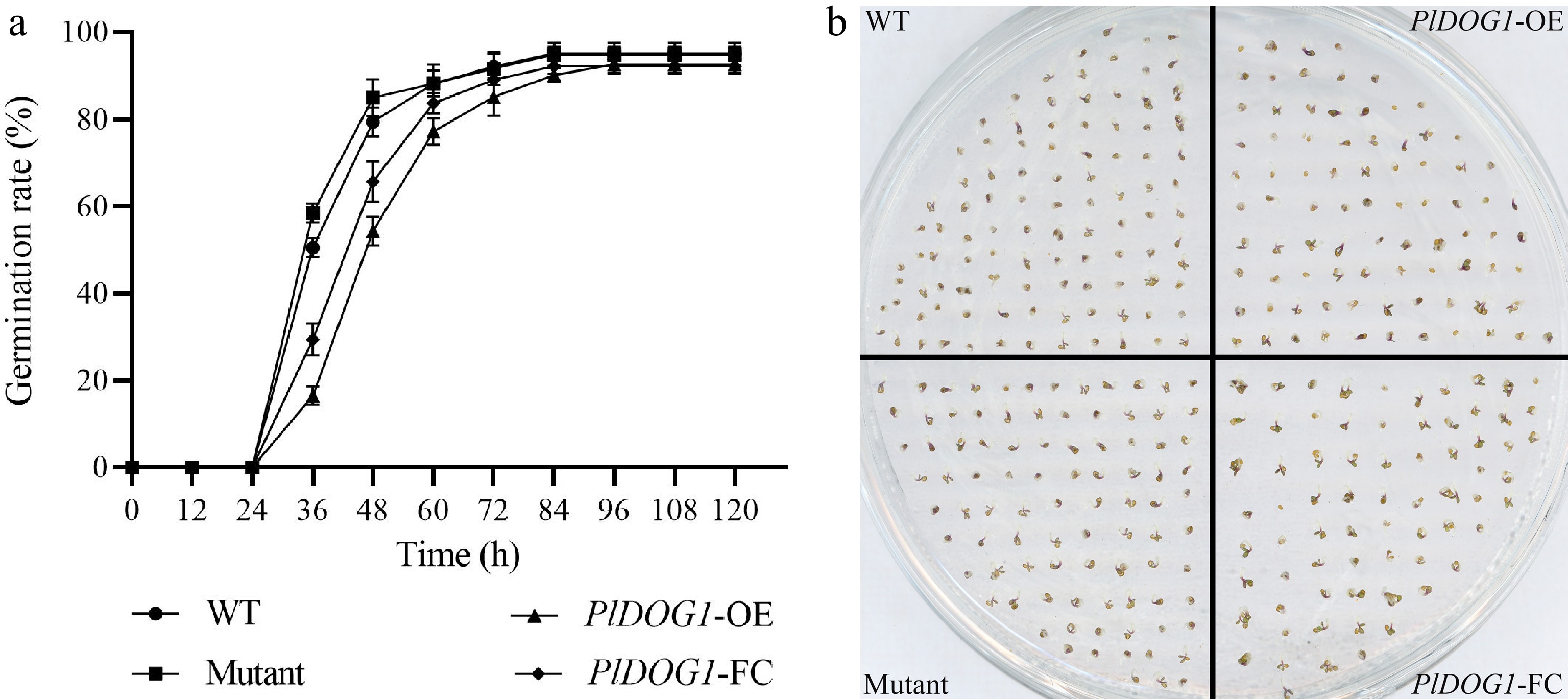
Figure 2.
Germination analysis of wild type, PlDOG1 overexpression, PlDOG1 complement expression and PlDOG1 function deficient Arabidopsis seeds. (a) Germination rates statistical analysis of different A. thaliana seeds. (b) Germination phenotype at 60 h. WT, wild type; PlDOG1-OE, transgenic overexpression seeds; PlDOG1-FC, mutant plants with functional complementation expression; Mutant, DOG1 deficient mutant seeds. Three independent biological replicates were prepared. Data differences were analyzed for significance using one-way analysis of variance (ANOVA) followed by Duncan's test.
Overexpression of PlDOG1 extends the duration needed for dormancy release in P. lactiflora seeds
-
To delve deeper into how PlDOG1 regulates seed dormancy in P. lactiflora, we utilized the Agrobacterium-based transformation and Virus-induced gene silencing (VIGS). These were applied to transiently overexpress and silence PlDOG1 within the seed embryos of P. lactiflora. The successful acquisition of required transient overexpression and silencing embryos was confirmed by qRT-PCR results (Supplementary Fig. S2). The sprouting behavior of these embryos revealed that the silenced embryos started to germinate on the 1st day after dark culture. By the 5th day, the germination rate approached 80%, and it remained above 80% on the 6th day. Control (CK) embryos transformed with ddH2O started germinating 2 d after cessation of dark culture, reaching a germination rate of 60% after 6 d. In contrast, embryos with transient overexpression exhibited the minimum germination rate, remaining under 40% even following 6 d of cultivation (Fig. 3). Based on these findings, we propose that PlDOG1 exerts negative regulation on the release of seed dormancy in P. lactiflora.
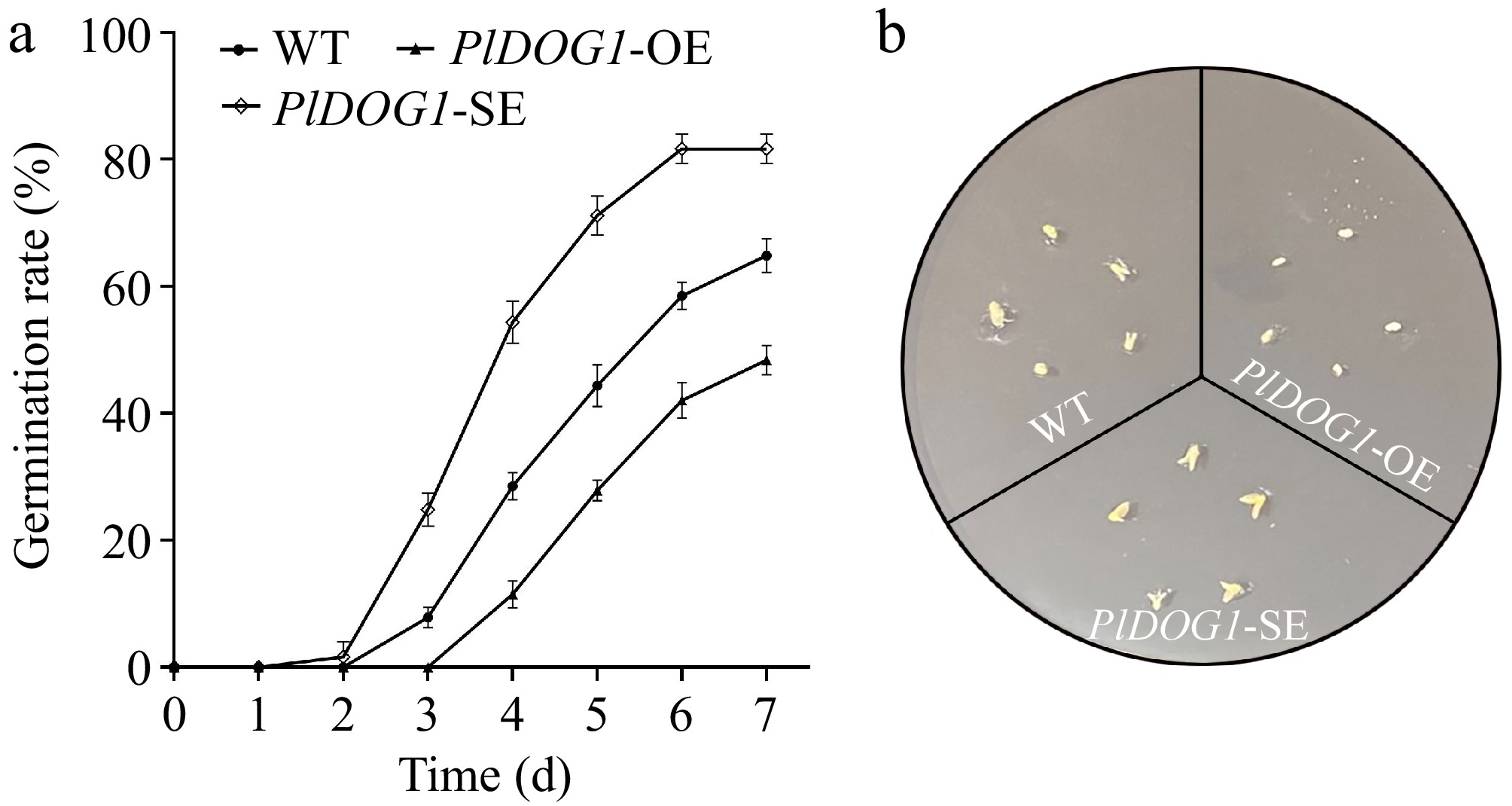
Figure 3.
Phenotypic and statistics analysis of PlDOG1 transgenic P. lactiflora embryos. (a) Germination rates statistical analysis of different P. lactiflora embryos. (b) Embryos germination phenotype observed on MS medium containing 0.5 mg/L GA3 after 4-d cultivation. WT, ddH2O water-transformed embryos; PlDOG1-OE, PlDOG1 overexpressed embryos; PlDOG1-SE, PlDOG1 silenced embryos. Three independent biological replicates were prepared. Data differences were analyzed for significance using one-way analysis of variance (ANOVA) followed by Duncan's test.
Cloning and bioinformatics analysis of the promoter of PlDOG1
-
It was previously reported that the RY-motif is very conserved in the promoters of DOG1 genes in different plants, so we hypothesized that the RY-motif may also exist in the promoter of PlDOG1 in P. lactiflora. To test this hypothesis, the promoter sequence of PlDOG1 was cloned, and the possible cis-acting elements on it were analyzed. Given the absence of genomic data for reference, we employed the chromosome walking method to amplify the 5' end sequence of PlDOG1. Following multiple amplifications, sequencing alignments, and sequence splicing, a 1,602 bp-long 5' end sequence was obtained. Subsequently, we analyzed the cis-acting elements within this sequence. The results showed that the sequence contained TATA-box and CAAT-box elements that are normally found in general gene promoter sequences, as well as some key elements involved in plant growth regulation and morphogenesis. Notably, the RY-motif was found to be linked to seed formation, development, and dormancy. Additionally, components such as CGTCA-motif were identified as being associated with MeJA-responsiveness, while G-box was linked to light sensitivity (Table 1). These findings underscore the significant role of PlDOG1 throughout multiple phases of plant survival, particularly in biological processes associated with seeds.
Table 1. Important motifs contained in the PlDOG1 promoter sequence.
Motif name Sequence Site Function RY-motif CATGCAT +517 Seed formation, development,
and dormancyCGTCA-motif CGTCA +1,453 MeJA-responsiveness G-box CACGAC +1,262 Light responsive element GA-motif ATAGATAA −990 Light responsive element MYB TAACCA +1,548 Light responsive element TC-rich repeats GTTTTCTTAC −1,241 Protective and stress response mechanisms TGACG-motif TGACG −1,453 MeJA-responsiveness W-box TTGACC +381 Abiotic stress response To further investigate whether the sequence has promoter activity, we inserted the whole and the various deletion sequences of the 5' end of the sequence into the pCAMBIA1301 vector that has a GUS gene, replaced the original promoter in it, injected them into healthy tobacco leaves by Agrobacterium-mediated transformation, and observed the GUS gene expression level. The findings revealed that GUS gene expression led to the presence of blue coloration uniformly across all tobacco leaves. This observation indicates that these fragments possess promoter activity and effectively initiate gene expression (Fig. 4).
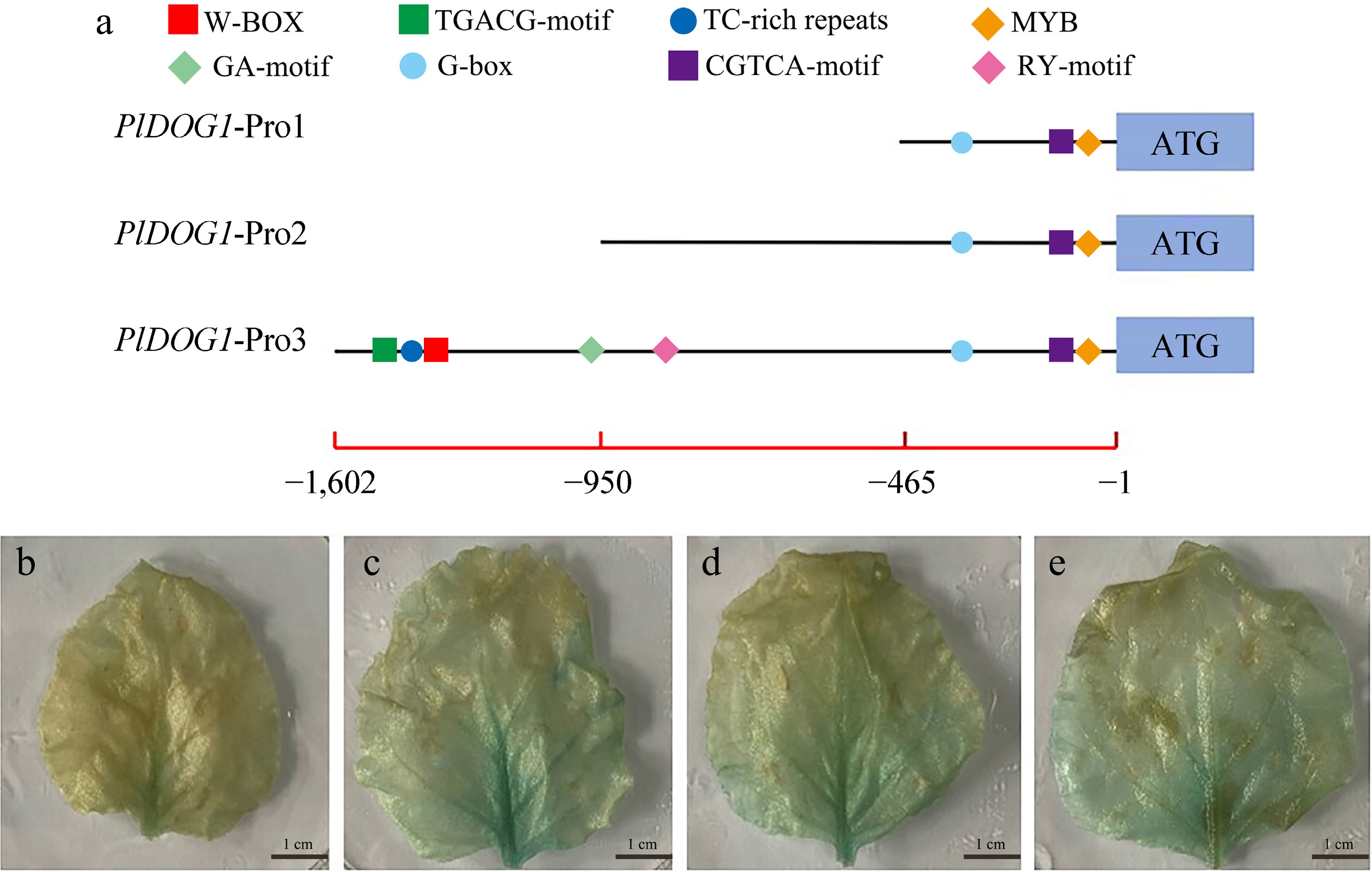
Figure 4.
Analysis of promoter activity of PlDOG1 promoter. (a) Schematic diagram of the absence of the 5' end of the PlDOG1 promoter sequence. GUS gene expression driven by different promoter sequences in healthy tobacco leaves: (b) 35S promoter, (c) PlDOG1-Pro1, (d) PlDOG1-Pro2, (e) PlDOG1-Pro3. Bars: 1 cm.
PlARF2 can bind to the promoter of PlDOG1
-
Our previous studies found that PlARF2 could regulate downstream gene expression by binding RY-motif[34]. Considering the existence of RY-motif in the promoter region of PlDOG1, we hypothesized that PlARF2 (GenBank Accession No. OQ715342) may bind to the promoter of PlDOG1 to participate in regulating seed dormancy release. To validate this hypothesis, we performed a Y1H experiment based on PlARF2 and the promoters of PlDOG1. The results demonstrated that the experimental group exhibited normal growth in the medium following appropriate incubation time (Fig. 5a), mirroring the phenotype observed in the positive control group. Consequently, this finding suggested that PlARF2 is capable of binding to the promoter region of PlDOG1 in yeast.
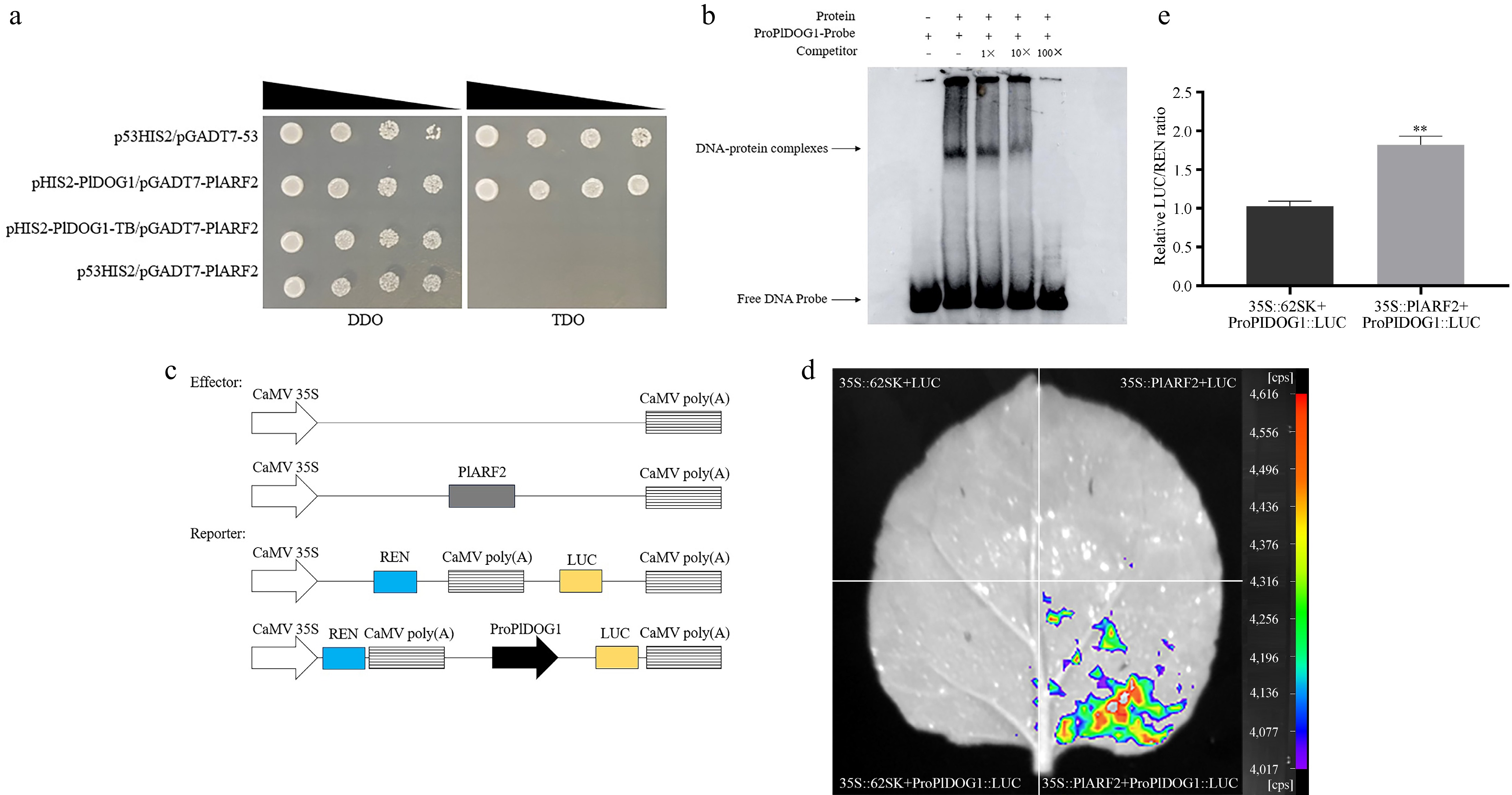
Figure 5.
Identification of the binding between PlARF2 and the PlDOG1 promoter. (a) The association of PlARF2 with the PlDOG1 promoter determination by using the Y1H system. p53HIS2/pGADT7-53 was the positive control group, pHIS2-PlDOG1/pGADT7-PlARF2 was the experimental group, pHIS2-PlDOG1-TB /pGADT7-PlARF2 was the mutant group, and p53HIS2/pGADT7-PlARF2 was the negative control group. Yeast transformants were serially diluted (1/1, 1/10, 1/100, 1/1000) and plated on TDO selective medium lacking Leucine, Tryptophan, and Histidine, supplemented with 50 mM 3-AT for identification. Parallel cultures grown on DDO medium without Tryptophan and Leucine served as growth controls. (b) EMSA analysis of PlARF2 combined with the promoter of PlDOG1. (c) Diagrammatic representation of Effector and Reporter assembly. (d) LUC fluorescence was detected using a live plant imaging system. (e) Analysis of the relative LUC/REN value. Asterisks in each column denote significant differences (p < 0.01).
To prevent false positive results and ensure the reliability of our findings, we employed EMSA testing to further validate the binding capacity between PlARF2 and the PlDOG1 promoter. We obtained purified PlARF2 protein through prokaryotic expression and conducted an EMSA assay using a biotin-labeled probe derived from the promoter sequence of PlDOG1. The results demonstrated that the free probes labeled by biotin remained unbound in reactions lacking PlARF2. Upon protein addition, distinct shifted bands corresponding to protein-DNA complexes were detected, with migration retardation relative to free probes. These complexes exhibited a concentration-dependent reduction in intensity upon the addition of unlabeled competitor probes (Fig. 5b), confirming PlARF2's capacity for sequence-specific interaction with the PlDOG1 promoter in vitro.
The binding ability of PlARF2 to the PlDOG1 promoter was further confirmed through a Dual-luciferase reporter gene detection assay. In vivo images revealed fluorescence solely when the PlARF2 and PlDOG1 promoters co-existed, indicating their interaction in plants and PlARF2 can positively regulate the expression of PlDOG1 (Fig. 5c−e). In light of these findings, we propose that PlARF2 can bind to the PlDOG1 promoter sequence.
-
P. lactiflora, a perennial plant belonging to the genus Paeonia, exhibits abundant and vibrant flowers with diverse colors. The captivating flower color of this plant, coupled with its remarkable adaptability to diverse environmental conditions, renders it an exceptional candidate for landscape design. Moreover, certain P. lactiflora varieties also yield valuable medicinal roots used in traditional Chinese medicine[30]. Consequently, P. lactiflora holds immense value in terms of landscape aesthetics, economic benefits, and medicinal applications. Nevertheless, the cultivation and propagation of P. lactiflora still encounter significant challenges due to their unique and intricate dormancy mechanism that hampers seed germination under natural conditions resulting in delayed emergence rates and low germination percentages. These factors pose substantial obstacles to the industry's productivity as well as hinder the selection and breeding of novel P. lactiflora cultivars[35]. The regulation of seed dormancy involves several crucial genes, among which DOG1 plays a significant role. DOG1 is present in various plant species and exhibits a relatively conserved mechanism in controlling seed dormancy[36]. Therefore, comprehending the molecular regulatory framework behind DOG1-mediated regulation of seed dormancy in P. lactiflora is of great scientific value.
The DOG1 gene was initially discovered in A. thaliana, and it represents a plant-specific gene family[37]. However, recent scientific advancements have unveiled the presence of the DOG1 gene in an expanding array of plant species, encompassing barley, rice, wheat, sorghum, maize, and soybean[19,38]. Although multiple members of the DOG1 family exist within the same species, their roles in seed dormancy exhibit limited redundancy. Within a single species, distinct members of the DOG1 protein family might exert disparate regulatory influences on seed dormancy. For instance, in A. thaliana, the single mutants atdogl1, atdogl2, and atdogl3 did not exhibit any alteration in dormancy levels. Conversely, overexpression of AtDOGL3 resulted in heightened sensitivity of seed germination to ABA. However, overexpression of AtDOGL5 had no impact on the ABA sensitivity of the seed germination process. Additionally, the mutant atdogl4 displayed an augmented level of seed dormancy indicating that AtDOGL4 acts as a negative modulator of seed dormancy[11,39]. Consequently, it is crucial to select a key member from the DOG1 gene family for further investigation. In this study, in order to identify the homologous gene of AtDOG1, a critical member of the DOG1 family in A. thaliana, in P. lactiflora, we compared the coding sequence (CDS) sequences of differentially expressed genes (DEGs) from our preliminary RNA-sequencing data at various stages of P. lactiflora seed germination with AtDOG1. As a result, we identified a homologous gene named PlDOG1. In parallel, we executed a conserved domain assessment of the polypeptide chain specified by this gene and identified a highly conserved DOG1 superfamily domain. This discovery enabled us to recognize this gene as a homologous counterpart of AtDOG1 in P. lactiflora, thereby prompting further research.
DOG1 serves as the principal regulator governing seed dormancy. Although DOG1 has been found in different species, research on it has focused on A. thaliana. Previous studies have demonstrated that DOG1 predominantly exerts a positive regulatory role in seed dormancy[40]. Ethylene receptor 1 (ETR1) and Ethylene response factor12 (ERF12) exert inhibitory effects on AtDOG1 expression, thereby promoting early seed germination[15]. AtDOG1 can also function as an upstream regulator of ABA-HYPERSENSITIVE GERMINATION1 (AHG1), thereby modulating the ABA signaling pathway and consequently enhancing seed dormancy levels[41]. Low temperature could increase the expression of AtDOG1, leading to the deepening of seed dormancy[42]. Furthermore, DOG1 and its family members in other species have also been involved in the modulation of seed germination. For instance, in rice, it was found that there existed a positive correlation of the expression of OsDOG1L3 with seed dormancy[11]. Similarly, in wheat and barley, seeds from plants that overexpressed TaDOG1L4 and HvDOG1L1 exhibited enhanced levels of dormancy compared to those from the wild type[36]. In this study, we utilized A. thaliana and P. lactiflora as plant materials to conduct heterologous and homologous transformation of PlDOG1 to elucidate its role in seed dormancy. The results demonstrated that PlDOG1 negatively regulated seed germination, which is consistent with previous findings in other species[7,37,43]. However, further investigation is needed to explore the specific molecular mechanism underlying PlDOG1's regulation of seed dormancy and potential additional functions.
The promoter, a specialized sequence located upstream of the gene coding region, encompasses diverse cis-acting elements. By analyzing the cis-acting elements within the sequence, we can make preliminary predictions regarding the potential binding of important transcription factors to the promoter sequence, thereby gaining insights into the possible functions and underlying mechanisms of genes associated with this promoter[44]. As early as 2006, when DOG1 was first discovered, it was suggested that the divergence between enhanced and diminished degrees of dormancy in seeds may be caused by the difference in DOG1 gene expression, which may be related to the cis-acting elements in its promoter region[37]. In this study, using the chromosome walking technique, we acquired the 1,602-bp promoter sequence of PlDOG1. In the obtained promoter sequence, we recognized numerous cis-acting elements related to plant survival, including RY elements linked to seed development and dormancy, G-box elements related to light response, as well as other crucial components such as W-box and MYB. This hinted that PlDOG1 could play a role in controlling associated processes by binding to transcription factors that recognize these elements. Notably, some of these elements appear relatively conserved in DOG1 gene promoters across different plant species. For instance, we detected the presence of G-box and RY motifs within the promoter region of AtDOG1[45]. Furthermore, we assessed the promoter activity of the obtained sequence and found that it could initiate gene transcription normally while successfully expressing the target gene. These results provide valuable insights into which transcription factors can bind to the promoter of PlDOG1 and how PlDOG1 is regulated. Relevant studies have previously examined the transcription factors likely to be engaged in the governance of DOG1. In A. thaliana, several key transcription factors associated with seed maturation or dormancy were previously thought to be upstream transcription factors of AtDOG1, such as AtABI3, AtFUS3, and AtLEC2[43,46]. These transcription factors share a common feature of possessing a B3 binding domain that specifically targets RY (CATGCA (TG)) motifs located within the promoter regions of multiple genes related to seed development and dormancy[47−49]. In our previous study, we discovered that P. lactiflora auxin response factor PlARF2 possesses a B3-binding domain and can regulate downstream gene expression by binding to the RY motif. In this study, we recognized the occurrence of the RY motif of PlDOG1 promoter and subsequently confirmed the interaction between PlARF2 and the PlDOG1 promoter through Y1H, EMSA, and Dual-LUC. This finding significantly contributes to understanding the molecular framework governing DOG1-related regulation of seed dormancy while offering precious perspectives on the regulation of seed dormancy by auxin signals. Previous studies have demonstrated the presence of numerous transcription factors in plants harboring the B3 binding domain, but not all transcription factors possessing this domain exhibit successful interaction with the RY motif for downstream gene expression regulation[50]. Therefore, whether other transcription factors regulate its expression by binding the RY motif or binding the promoter of DOG1 in other ways remains to be investigated. Nevertheless, in light of the results from this study, combined with our prior research, we proposed a concise regulatory model whereby PlARF2 is regulated by auxin signaling and subsequently interacts with PlDOG1 to modulate seed dormancy (Fig. 6).
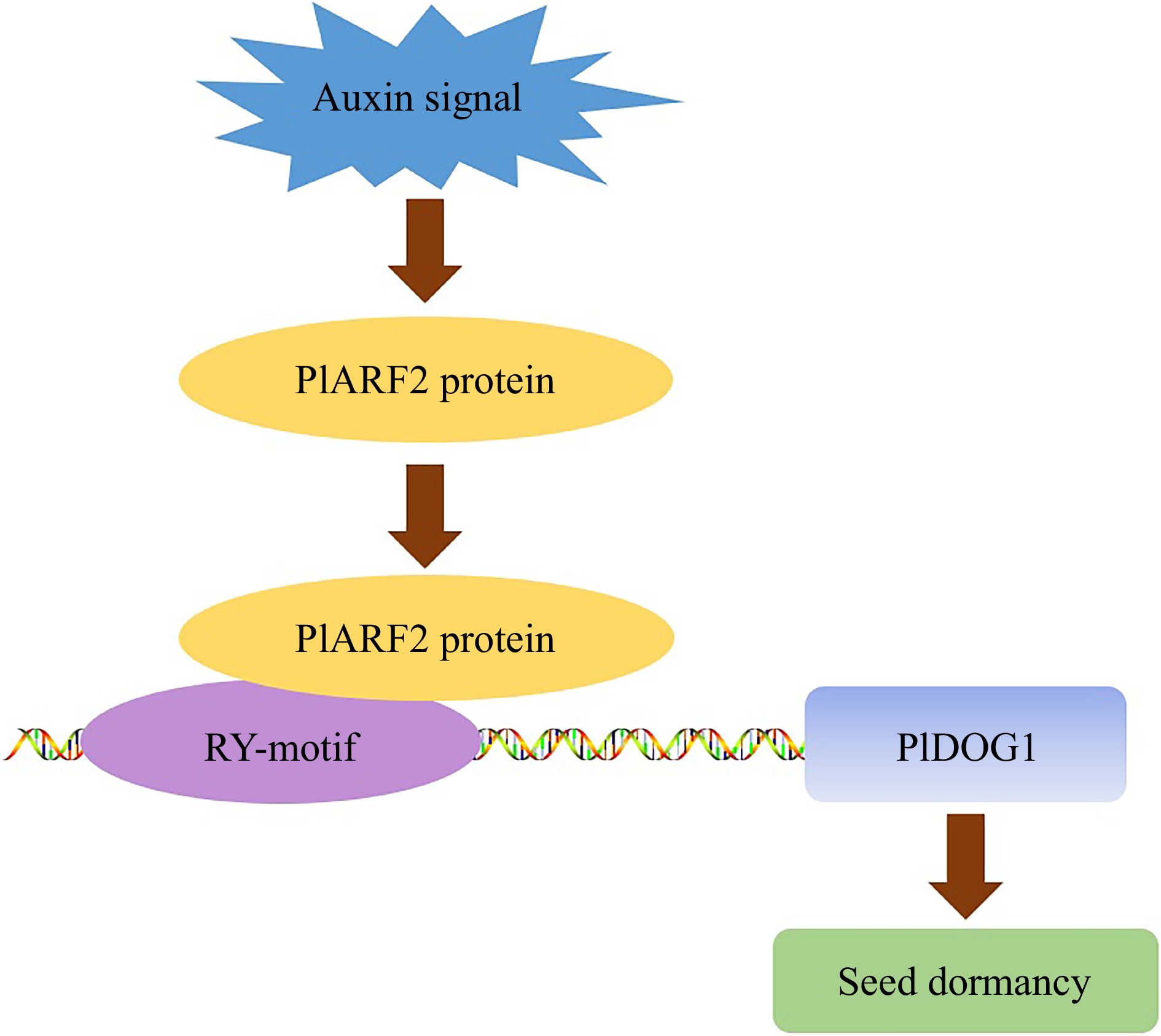
Figure 6.
A proposed model illustrates that PlDOG1 regulates seed dormancy through binding to PlARF2. Auxin signals trigger the expression of PlARF2, and then the PlARF2 protein binds to the PlDOG1 promoter to regulate seed dormancy.
-
In summary, this investigation cloned the PlDOG1 gene and its promoter sequence through a series of experimental procedures, verified the activity of its promoter sequence and the binding of upstream transcription factor PlARF2 with the PlDOG1 promoter sequence, and characterized the function of the PlDOG1 gene. The results show that the PlDOG1 amino acid sequence is highly homologous to other plants and contains a typical DOG1 family conserved superfamily domain. The outcome of the phylogenetic analysis indicated a close phylogenetic affinity between PlDOG1 and PsDOG1. Y1H, EMSA, and Dual-LUC showed that PlARF2 is capable of binding to the PlDOG1 promoter and exerting a positive regulatory effect on PlDOG1 expression. The germination test showed that the activation of PlDOG1 led to an increase in the time taken for the dormancy release of P. lactiflora seeds. In summary, our results revealed a new regulation mode of P. lactiflora seed dormancy, that is, PlARF2 can be involved in the modulation of P. lactiflora seed dormancy via the binding to the PlDOG1 promoter sequence. The findings of this study provide a theoretical framework for subsequent investigations into the mechanistic roles of DOG1 genes and auxin regulating plant seed dormancy and elucidate the related regulatory mechanism of peony seed germination from the perspective of molecular biology.
This work was supported by the Scientific Research Fund of Liaoning Provincial Education Department (LJKMZ20221027) and the National Natural Science Foundation of China (32071814 and 31470696).
-
The authors confirm contribution to the paper as follows: study conception and design: Duan S; data collection: Duan S, Song W, Kang X, Sun T, Xin R, Kan M; analysis and interpretation of results: Duan S, Fei R; draft manuscript preparation: Duan S, Guan S, Sun X. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primer sequence.
- Supplementary Fig. S1 Analysis of protein conserved domain of PlDOG1.
- Supplementary Fig. S2 Relative expression of PlDOG1 in transiently transformed P. lactiflora seeds. WT, seeds treated with ddH2O; PlDOG1-OE, PlDOG1 transient overexpression seeds; PlDOG1-SE, PlDOG1 transient silenced seeds. Three biological replicates were set up. One-way analysis of variance (ANOVA) and Duncan's method was used to analyze the significance of data differences. The different letters within a column marked significant differences (p < 0.05).
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Duan S, Guan S, Fei R, Sun T, Kang X, et al. 2025. PlDOG1 can be directly regulated by PlARF2 and participate in the regulation of Paeonia lactiflora seed dormancy. Ornamental Plant Research 5: e020 doi: 10.48130/opr-0025-0018 

PlDOG1 can be directly regulated by PlARF2 and participate in the regulation of Paeonia lactiflora seed dormancy
- Received: 16 October 2024
- Revised: 20 March 2025
- Accepted: 25 March 2025
- Published online: 16 May 2025
Abstract: Paeonia lactiflora Pall. is a significant ornamental, medicinal, and economic flower with a global distribution. Its prolonged dormancy period or incomplete dormancy release results in low germination rates and extended reproduction cycles, which severely hinders the development of related industries. The DELAY OF GERMINATION1 (DOG1) gene functions as a pivotal factor in seed dormancy regulation. However, it is still unknown how the DOG1 gene mediates seed dormancy in P. lactiflora. In this study, the function and regulatory mechanism of the PlDOG1 gene were comprehensively analyzed. Through cloning the PlDOG1 gene and its promoter sequence, it was revealed that the encoded protein possesses a characteristic DOG1 superfamily conserved domain and exhibits high similarity with the DOG1 gene in other plants, as demonstrated by phylogenetic analysis. Functional studies indicated that PlDOG1 delays the release of seed dormancy. Moreover, employing Y1H, EMSA, and Dual-LUC assays, we demonstrated that PlARF2 specifically binds to the PlDOG1 promoter and enhances its transcriptional activity. This creates a new framework for the interaction between the auxin signaling pathway and DOG1. The 'PlARF2-PlDOG1' regulatory model identified in this study provides a key target for molecular breeding in P. lactiflora, offering significant practical value for overcoming technical bottlenecks in P. lactiflora seedling cultivation. Additionally, it provides a theoretical basis for understanding the molecular interaction network of auxin and plant hormones in the dormancy regulation of plant seed, significantly advancing seed biology theory and guiding genetic enhancements in horticultural crops.