









-
Camellia reticulata 'Tongzimian', also known as 'Yinhong Juban', with the English name 'Baby Face', originates from Dali, Yunnan province (China). It is one of the rare light-colored flowers among the cultivars of C. reticulata[1,2]. Its delicate flower, pale white with a red blush, resembles the tender face of a young child, making it famous as one of the traditional precious cultivars of C. reticulata (Supplementary Fig. S1). As a woody ornamental plant with distinct regional characteristics, C. reticulata is located at the top of the 'Eight Famous Flowers' of Yunnan and serves as the city flower of Kunming, Chuxiong, and Tengchong. C. reticulata has a long history of cultivation. After more than 1,500 years of natural hybridization and artificial cultivation and breeding, cultivars with diverse morphoglogical variations have been developed and spread to various parts of the world[3]. As of October 2024, the International Camellia Register (
https://camellia.iflora.cn ) has recorded 859 cultivars of C. reticulata, but the colors of these cultivars are mainly red, with few color combinations or rare light hues[4,5]. The lack of yellow, light, and other color schemes due to the single color scheme is always one of the key factors hindering the market promotion and application of C. reticulata. Therefore, enriching the color breeding of C. reticulata has always been the pursuit of camellia breeders.Flower color plays a significant role in plant survival. Additionally, flower color is one of the key traits of ornamental plants, therefore, research on flower colors has always been a hot topic in scientific research[6]. So far, research on flower colors mainly includes the measuring of flower color, the effects of cell morphology and ultrastructural features, pigment content and type, osmotic regulators and antioxidant enzymes, endogenous hormones, petal cell pH, light, and so on. The morphology and structure of epidermal cells significantly affect the absorption and reflection of light, thereby influencing flower color. In Paeonia suffruticosa, the stripes on epidermal cells are densest when chlorophyll content is at its highest, and most sparse when chlorophyll content is at its lowest[7]. In wild C. japonica, significant differences in cell shape were observed between single-petaled and double-petaled flowers[8]. In C. nitidissima, the size and shape of epidermal cells in the petals at different developmental stages were found to correlate with the formation of flower color[9].
The variation in flower color is primarily determined by the types and relative content of petal pigments, such as flavonoids, carotenoids, chlorophyll, and alkaloids. The synthesis, storage, and metabolism of these pigments are regulated by environmental factors and the plant's internal physiological state, resulting in a diversity of flower colors[10−12]. For example, the pale pink petals of Mimulus lewisii are due to low anthocyanin content, while the bright red petals of M. cardinalis are attributed to higher levels of anthocyanins and carotenoids[13]. In Chrysanthemum morifolium, flower color is primarily determined by anthocyanins[14]. In the post-flowering stage, the lightening of petal color in chrysanthemum is regulated by a decrease in anthocyanin accumulation[15]. Previous studies have shown that flavonoids and anthocyanins are the main determinants of flower color in C. reticulata[1,2].
Osmotic regulators and antioxidant enzyme activity also play a significant role in petal coloration to some extent. Among them, anthocyanins are composed of anthocyanins and sugars. The synthesis of anthocyanins in cells requires an adequate sugar content, which can influence anthocyanin synthesis to varying degrees[16]. In addition, changes in soluble protein content during anthocyanin synthesis can also impact petal coloration. The reduction in soluble protein content in the petals of Rosa chinensis significantly promotes the synthesis of anthocyanins[17]. Meanwhile, the primary protective enzymes that scavenge reactive oxygen species (ROS) in petals include superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD), and their activity plays an important role in color change[16]. In Vanda Orchids, petal fading is mediated by increased POD activity, which leads to the rapid degradation of plant anthocyanins[18]. During the transition from purple to white in Camptotheca acuminata flowers, the activities of SOD, POD, and CAT enzymes significantly increased, surpassing other indicators such as protein content[19]. Furthermore, endogenous hormones play a crucial role in the growth and development of plants. They can affect the synthesis and accumulation of anthocyanins, thereby producing different petal colors[20,21]. In particular, abscisic acid (ABA) can promote the synthesis of anthocyanins[22]. The color change in the petals of Lonicera japonica is closely related to changes in the concentration of endogenous hormones[23]. Therefore, in the process of flower color formation, osmotic regulators, antioxidant enzymes, and endogenous hormones are key physiological and biochemical indicators that significantly influence the production of flower pigments. In addition, Petal coloration is also influenced by pH, light, temperature, and metal ions. In plants rich in anthocyanins, the effect of pH is particularly significant. Yoshida et al. found that the vacuolar pH of colored epidermal cells in Ipomoea tricolor unusually increased from 6.6 to 7.7, resulting in a shift in flower color from red to blue[12]. Light and temperature are among the key factors influencing anthocyanin biosynthesis. Early studies have shown that light can induce anthocyanin accumulation in Arabidopsis thaliana[24]. High temperatures significantly reduce anthocyanin accumulation in chrysanthemums[14]. Additionally, in plant cells, metal ions can form complexes with anthocyanins, a concept known as metal chelation theory. However, the highly colored metal complexes resulting from chelation can also influence the manifestation of flower color[25].
As one of the lightest-colored cultivars of C. reticulata, the physicochemical mechanisms underlying the coloration of 'Tongzimian' remain unreported. This study utilized petals from six different flowering stages of 'Tongzimian' as research materials (Fig. 1a). By analyzing the color difference values, changes in cell structure and ultrastructure, and physiological changes at different coloration stages, we explored the physiological mechanisms underlying the coloration changes. The findings provide a theoretical foundation for exploring the molecular mechanisms of flower coloration in C. reticulata and support the optimization of camellia flower color breeding programs.
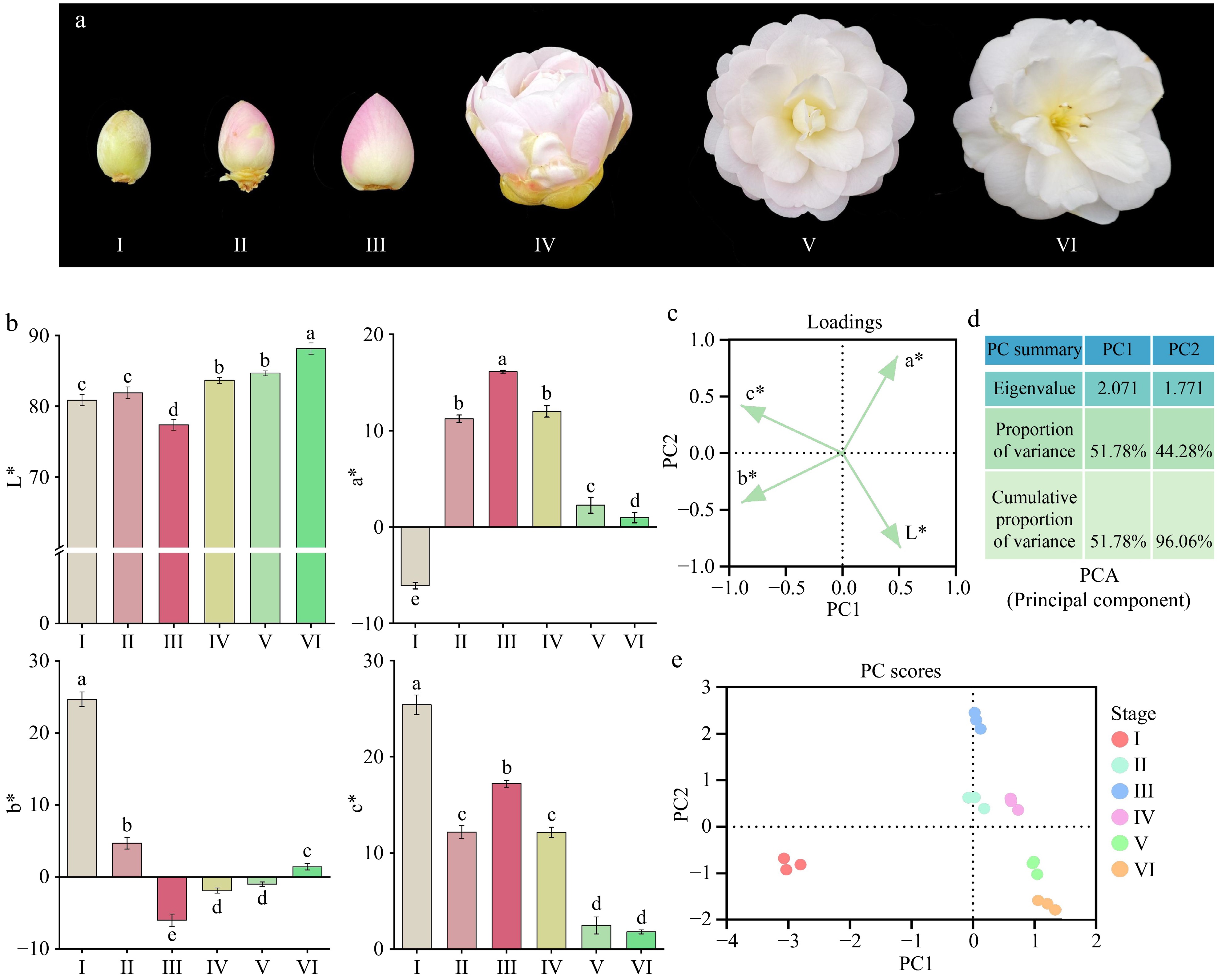
Figure 1.
(a) Color changes in 'Tongzimian' petals at six different flowering stages: I (yellow-green, bud stage), II (light pink, coloring stage), III (distinct pink, exposed color stage), IV (light pink, early bloom stage), V (whitish, full bloom stage), and VI (white, senescence stage). (b) CIELab* color parameters of petals at six coloration stages; different lowercase letters indicate significant differences (p < 0.05, ANOVA). Error bars represent the standard deviation of three biological replicates. (c) Loadings plot of Principal Component Analysis (PCA). (d) Principal Component Analysis (PCA) results. (e) Component analysis score plot.
-
This study uses petals from six different flowering stages of C. reticulata 'Tongzimian': bud stage (I), coloring stage (II), exposed color stage (III), early bloom stage (IV), full bloom stage (V), and senescence stage (VI) as materials (Fig. 1a). The petals were collected from January 16 to February 28, 2023, from healthy, disease-free 'Tongzimian' plants growing in good condition at the Yiliang Zhengyang Camellia Garden Landscaping Co., Ltd. (Coordinates: 24°88' N, 103°23' E, Altitude: 1,950 m). For each coloration stage, sampled petals were collected from three independent flowers per replicate and each flower selected from a single plant, three biological replicates were set up in total. The collected petals were immediately wrapped in aluminum foil, placed in liquid nitrogen, and stored at −80 °C until the completion of subsequent experiments.
Research contents
-
This study conducts measurements on the flower color, structural characteristics of petal epidermal cells, pigment composition and content, endogenous hormones, osmotic regulators, and antioxidant enzymes of petals from six different flowering stages of 'Tongzimian'. The specific details are as follows.
Flower color measurement
-
The flower color of the six different flowering stages of 'Tongzimian' was measured using a 3nh color difference meter (Model: NR110), obtaining the CIELab* parameters[26]. After calibration with a color chart, three points were randomly selected on the upper epidermis of the petals at 2/3 of their length, and the average value was recorded as the color difference for that stage. The average value from three petals was taken as the color value for a single plant at each stage, and the average of three plants was used as the color difference for each stage. The measurements provided values for brightness (L*) and the two chromatic components, a* and b*. The chroma (C*) was then calculated using the formula: C* = (a*2 + b*2)1/2.
Cell morphology and ultrastructural features
-
Petals from six different flowering stages of 'Tongzimian' were sampled and quickly placed in electron microscopy fixative and FAA fixative (70 % ethanol), then they were stored at 4 °C for analyzing the epidermal cell structure of the petal segments using scanning electron microscopy (SEM), transmission electron microscopy (TEM), and paraffin sectioning. The distribution pattern of pigment accumulation in the petals was also observed to analyze the structural basis for flower coloration (performed by Wuhan Maispu Biotechnology Co., Ltd., Wuhan, China). The anatomical structural parameters of the petals were measured using ImageJ 1.4.8 software (Supplementary Fig. S2), including petal thickness, size of single epidermal cells on both the upper and lower surfaces, thickness of the parenchyma tissue, and size of individual cells. For each parameter, six fields of view were selected, and measurements were repeated five times.
Pigment measurement
-
Flavonoid content was determined using the ethanol extraction method. Fresh petals (0.500 g) were precisely weighed, ground, and then extracted twice with 10 mL of 80 % ethanol at 75 °C in a water bath. The absorbance of the sample solution was measured at 510 nm. Rutin was used as the reference standard to construct a calibration curve, and the flavonoid content in the sample was calculated accordingly[26]. Anthocyanin content was determined using the hydrochloric acid-ethanol method[27]. Fresh petals (approximately 0.500 g) were precisely weighed and mixed with 25 mL of 0.1 mol/L hydrochloric acid-ethanol solution. The mixture was then extracted at 60 °C in a water bath for 60 min (extracted twice). After cooling, the solution was filtered. To prevent interference from chlorophyll in the extract, the absorbance of the solution was measured at 530, 620, and 650 nm using a spectrophotometer. The anthocyanin absorbance was then calculated using the Greey formula, and the anthocyanin content was determined accordingly. The total carotenoid content was determined using the extraction colorimetric method[28]. Fresh petals (0.200 g) were weighed and ground with 25 mL of 95 % ethanol in a mortar. The mixture was transferred to a centrifuge tube and left to soak in the dark for half a day. After filtration, the absorbance of the extract was measured at 665, 649, and 470 nm using a spectrophotometer. The total carotenoid content was calculated based on the Lambert-Beer law.
Measurement of osmotic regulators and antioxidant enzymes
-
The soluble sugar content was determined using the anthrone colorimetric method[29]. Fresh petals (0.200 g) were weighed and extracted in a boiling water bath. The absorbance of the extract was measured at 630 nm, and the soluble sugar content in the petals was calculated based on the standard linear equation. Soluble protein content was determined using the Coomassie Brilliant Blue method. Fresh petals (0.300 g) were weighed and extracted. After dilution, the absorbance of the extract was measured at 595 nm, and the soluble protein content in the petals was calculated based on the standard linear equation[29]. The levels of SOD, POD, CAT, and ROS were measured using assay kits provided by Nanjing Ruiyuan Biotechnology Co., Ltd (Nanjing, China)[30,31].
Measurement of endogenous hormones
-
Endogenous hormones in the flower petals were extracted and measured using a high-performance liquid chromatography (HPLC) system (Agilent 1290) coupled with an AB Sciex Qtrap 6500 mass spectrometer (HPLC-MS/MS)[32]. The levels of endogenous hormones, including indole-3-acetic acid (IAA), ABA, cytokinins (CKs), trans-zeatin riboside (tZR), zeatin (Zeatin), trans-zeatin (T-zeatin), isopentenyl adenine (IP), gibberellin (GA), jasmonic acid (JA), salicylic acid (SA), and indole-3-acetic acid-aspartic (IAA-ASP) were quantified at six different flower coloration stages of 'Tongzimian'. Internal standards were added to the extract to correct the results (assisted by Nanjing Ruiyuan Biotechnology Co., Ltd., Nanjing, China).
Statistical analysis
-
Data organization was performed using Excel 2019, and significance analysis was conducted with SPSS 22.0. Data visualization was completed using Origin 22.0, ImageJ 1.4.8, and GraphPad Prism 9.5.0. Image creation and assembly were carried out with Adobe Photoshop 2024.
-
As shown in Fig. 1a, the petals of 'Tongzimian' at stage I are yellow-green, and at stage III, the color is the deepest, a pink hue, which later transitions to white. The CIELab* analysis of the petal color (Fig. 1b) reveals that the petal brightness (L*) at stage VI is significantly higher than in other stages, followed by stage V and IV, and then stage II and I. Stage III has the lowest L* value, significantly lower than the others, likely due to the petals being the darkest at this stage and the lightest at stage VI, which is almost white. The a* (green-red) value significantly increases from stage I to stage III, then decreases significantly, with the highest a* value at stage III, when the petals are pink. The b* (yellow-blue) value decreases significantly from stage I to stage III, and then significantly increases from stage V to stage VI, with no significant changes observed between stage IV and V. The C* (chroma) value shows a trend of significant decrease from stage I to stage II, followed by a significant increase from stage II to stage III, then a significant decrease from stage III to stage V, and finally stabilizes from stage V to stage VI.
Principal component analysis (PCA) revealed that the trends of CIELab* and c* values at different flowering stages of 'Tongzimian' were captured by PC1 and PC2, which together explained 96.06 % of the total variance (Fig. 1c & d). PC1 was primarily associated with L*, a*, and C*, indicating that these variables showed significant variation along PC1. PC2, on the other hand, was mainly associated with b* and L*, reflecting substantial changes in these variables along PC2 (Fig. 1e). As shown in Supplementary Table S1, during the flowering process of 'Tongzimian', there were significant negative correlations between L* and C* (r = −0.763), and between a* and b* (r = −0.766), meaning that as the brightness (L*) of the petals increased, the color saturation and red-green components (a*) decreased. In contrast, there was a significant positive correlation between b* and C* (r = 0.610), indicating that as the yellow component (b*) of the petals increased, the overall color saturation also increased.
Cellular morphology and ultrastructural characteristics of 'Tongzimian' petals at six different flowering stages
Changes in petal cell tissue structure
-
During the different stages of flowering in 'Tongzimian', the morphology of the petal epidermal cells showed significant changes (Fig. 2; Supplementary Table S2).
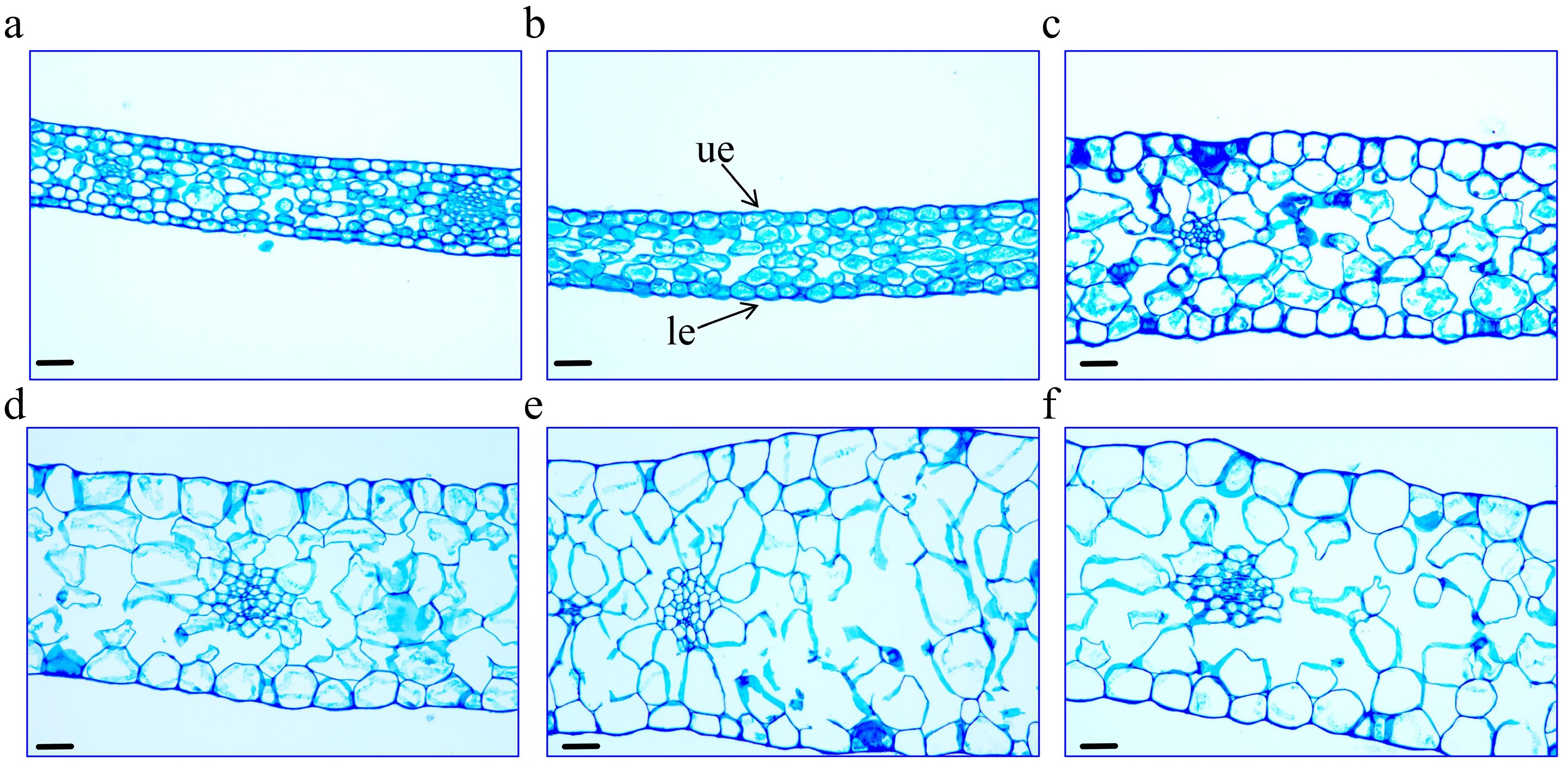
Figure 2.
Morphological changes in petal cells of 'Tongzimian' at flowering stages. (a) I (bud stage). (b) II (coloring stage). (c) III (exposed color stage). (d) IV (early bloom stage). (e) V (full bloom stage). (f) VI (senescence stage). ue: upper epidermal cells, le: lower epidermal cells. Scale bar = 40 μm.
Changes in petal thickness: From stage I to stage II, no significant change in petal thickness was observed. However, from stage III to stage V, the petal thickness increased significantly from 214.847 to 317.063 μm, after which it decreased significantly in stage VI.
Changes in petal epidermis: Throughout development, the size of the upper epidermal cells increased significantly, from 164.613 μm² at stage I to 2,221.718 μm2 at stage VI. This expansion correlates with the turgidity of the surface cells, particularly between stages IV and VI. As the cell area increased, the petal color gradually shifted towards white, becoming more uniform. Similarly, the lower epidermal cells showed a similar trend of expansion, especially from stage III to stage VI, where their size increased significantly from 740.490 to 1,345.528 μm2. This expansion corresponds to the maturation of the overall petal structure and the fading of the petal color.
Changes in parenchyma tissue thickness: The thickness of the parenchyma tissue followed a similar trend to that of the petal thickness. It significantly increased from 61.400 μm at stage I to 244.771 μm at stage V, then decreased significantly at stage VI. During this period, the arrangement of the parenchyma cells became more sparse, with increased intercellular spaces and significant cell expansion, from 192.884 μm2 at stage I to 2501.082 μm2 at stage V.
Overall, the development of 'Tongzimian' petals exhibited a significant expansion of the upper epidermal cells compared to the lower epidermal cells. This differential expansion may be closely linked to the functional structure of the petals, which could influence the changes in petal color during the flower development.
Changes in the ultrastructure of petal cells
-
During the development of 'Tongzimian' flowers, significant changes occurred in the microstructure of the petal epidermal cells. In the early stages, the cell walls were relatively thick, and the plastids and starch grains were abundant, indicating active energy storage and pigment synthesis. However, by stage V, plastids and cell walls began to degrade, which likely contributed to the fading of the petal color. As shown in Fig. 3a, the surface of the upper epidermal cells exhibited a transition from smooth to complex textures during the petal color changes. Notably, the cells at the top and base of the petals showed distinct differences, which could be one of the reasons for the color variation observed in 'Tongzimian' petals. At stage I, both the top and base cells had smooth surfaces and simple shapes, without any complex textural features. As the flower developed into stage III, the top cells began to show noticeable wrinkles and ripples, with the surface texture becoming more intricate. Although the base cells also exhibited some texture, it was less pronounced. By stage V, the textural features of the top cells became weaker, with some areas showing concavities, while the base cells retained relatively simple textures.
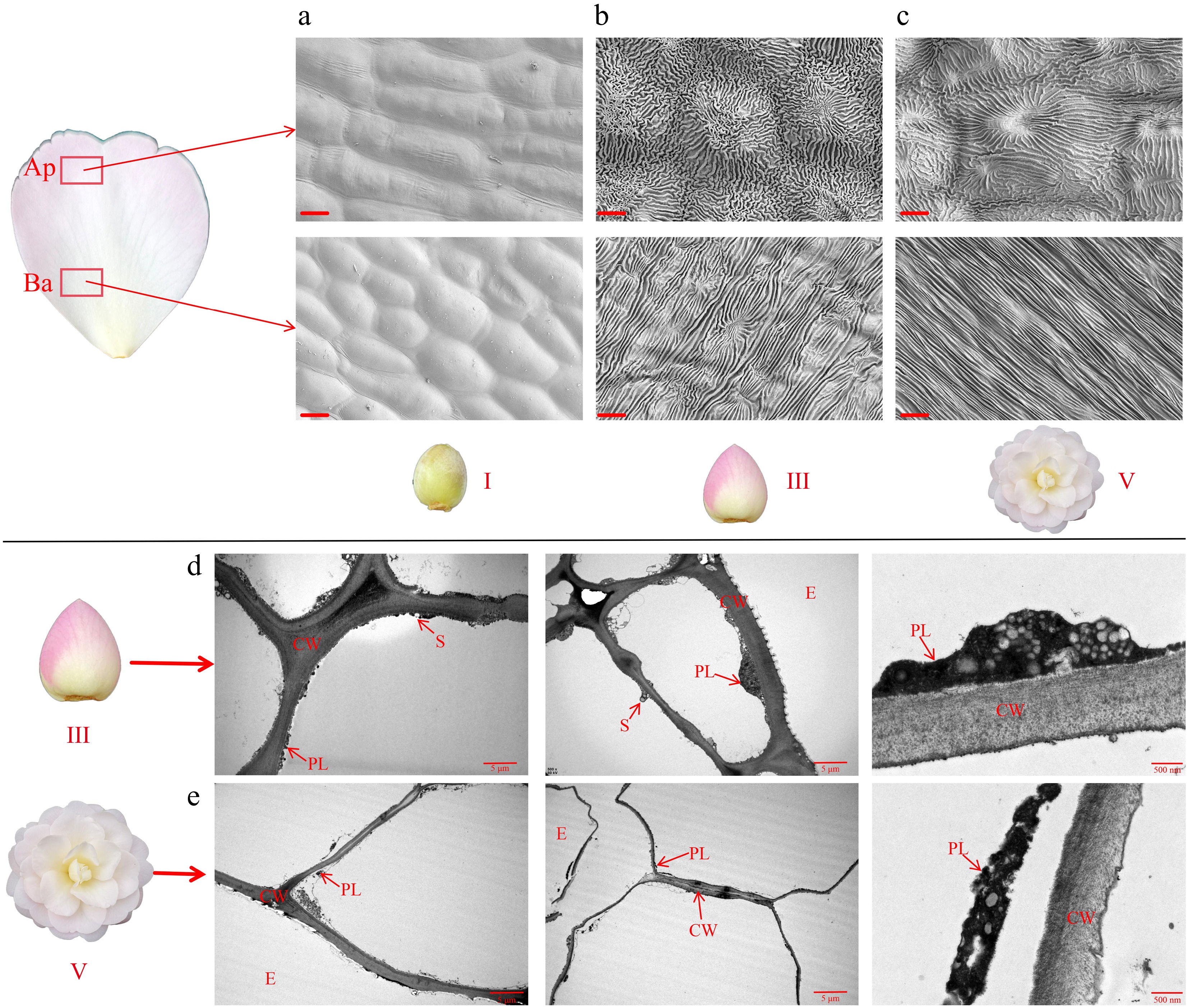
Figure 3.
Morphological changes in petal cells of 'Tongzimian'. (a)−(c) Ultrastructure of petal cells from SEM at stage I, stage III, and stage V; Ap: Apex position of the petal; Ba: Base position of the petal; scale bar = 10 μm. (d), (e) Ultrastructure of petal cells from TEM at stage III and stage V; PL: Plastid, CW: Cell wall, S: Starch granule, E: Epidermal cells; scale bar = 5 μm.
The ultrastructure of the epidermal cells of 'Tongzimian' petals showed significant differences between stage III and V. As shown in Fig. 3b, during stage III, the cell walls were thick, with a higher number of plastids and the presence of starch grains, indicating an active phase of energy storage and pigment synthesis. This likely contributed to the pink color of the petals. By stage V, the plastid morphology had changed, with starch grains largely degraded, and the cell walls showed signs of partial degradation. The pigment content had also significantly diminished, which likely led to the transition of the petal color to white.
Pigment composition analysis of 'Tongzimian' petals at six different flowering stages
-
During the development of 'Tongzimian' flowers, significant changes in pigment contents were obtained (Fig. 4). As shown in Fig. 4a, we know that the carotenoid content increased first and then decreased through the six coloring stages. From stage I to stage II, the carotenoid significantly increased from 9.108 to 10.065 μg/g, reaching the highest value of the entire blooming process. Subsequently, it significantly decreased at stages III, IV, and VI, with stage V showing a higher content than stages IV and VI, during which the petal base appeared pale yellow. Figure 4b shows that the flavonoid content decreased significantly from 40.684 to 16.804 mg/g. Stage I had obviously more flavonoids than stages II and III, with no significant difference between stage II and III, but both stages significantly had more content than stages IV, V, and VI. The latter three stages showed no significant difference in flavonoid content. According to Fig. 4c, stage I had the most anthocyanins, up to 16.358 μg/g, followed by stage III with a content of 14.952 μg/g, which was also significantly higher than stages II, IV, V, and VI. The content at stage I was also significantly higher than at stages II, V, and VI, with no significant difference among these three stages. This trend aligns with the gradual lightening of petal color from stages IV to VI.
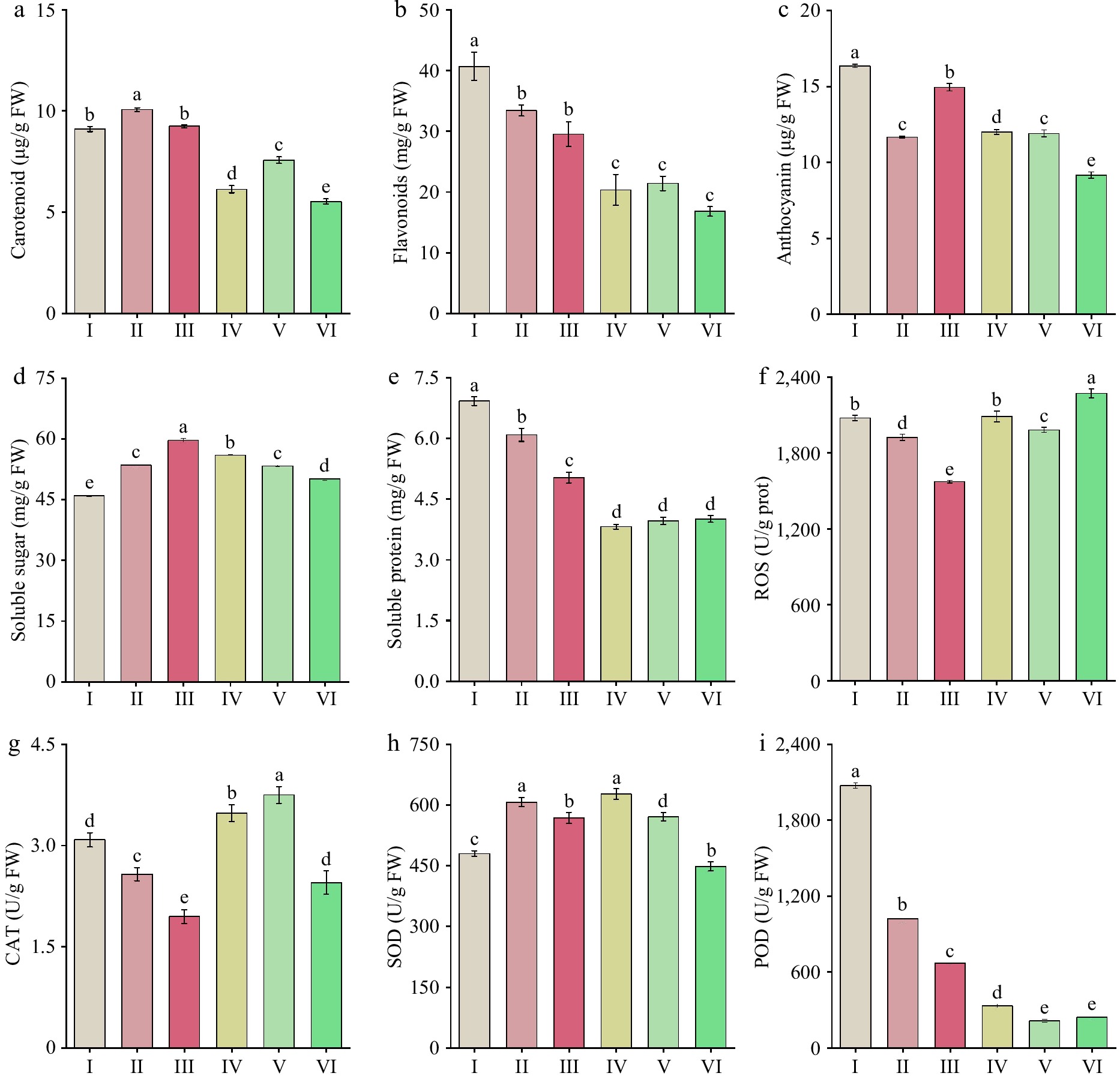
Figure 4.
Changes in pigment content, osmotic regulatory substances, and antioxidant enzyme activity in 'Tongzimian' petals at flowering stages. (a) Carotenoids. (b) Flavonoids. (c) Anthocyanin. (d) Soluble sugars. (e) Soluble protein. (f) Reactive oxygen species (ROS). (g) Catalase (CAT). (h) Superoxide dismutase (SOD). (i) Peroxidase (POD). Different lowercase letters indicate significant differences (p < 0.05, ANOVA).
Changes in osmoregulation substances in 'Tongzimian' petals at six different flowering stages
-
During the color development of 'Tongzimian' flowers, the content of soluble sugars in the petals (Fig. 4d) initially increased and then decreased. The content was relatively low at stage I (45.951 mg/g), but it significantly increased to the highest value (59.631 mg/g) from stage I to III, then it significantly decreased to 50.081 mg/g at stage VI. The highest content occurred at stage III, indicating that anthocyanin synthesis was the most active at this stage, which aligns with the changes in petal color. The content of soluble proteins (Fig. 4e) decreased significantly from stage I (6.922 mg/g) to stage IV (3.819 mg/g). From stage IV to stage VI, the contents remained relatively stable with no significant change. The variations in soluble protein content may reflect the dynamic changes in pigment synthesis and degradation, possibly influenced by the activity of anthocyanin-metabolizing enzymes, which may depend on the existence of soluble proteins.
Changes in antioxidant enzyme activities in 'Tongzimian' petals at six different flowering stages
-
ROS activity firstly decreased and then increased during the flowering process of 'Tongzimian' (Fig. 4f). ROS levels significantly decreased from stage I to III, followed by a significant increase at stage IV, and then a decrease at stage V. Finally, at stage VI, ROS activity significantly rose up to the peak value at 2,270.538 U/g. The changes in ROS activity were inversely correlated with the changes in anthocyanin content during the later stages. The activity of CAT (Fig. 4g) showed a trend of initially decreasing and then increasing. CAT activity significantly decreased from stage I to III, with no big difference between stages II and III. From stages III to V, CAT activity increased significantly, reaching its peak value (3.750 U/g). SOD (Fig. 4h) activity increased initially and then decreased, with the lowest activity at stages I and VI. The highest activity was suggested in the middle stages, especially at stages II and IV. POD (Fig. 4i) activity showed a significant decrease throughout the flowering period, which decreased from 2,072.722 to 214.616 U/g.
Changes in endogenous hormones in 'Tongzimian' petals at six different flowering stages
-
During the development of 'Tongzimian' flowers, significant changes were obtained in the levels of endogenous hormones at different flowering stages. The levels of IAA (Fig. 5a) showed no significant variation from stages I to III, but increased significantly from stages III to V, reaching the highest level (1.101 ng/g) at stage V. The content of IAA-ASP (Fig. 5b) remained stable from stages I to IV, but significantly increased from stages V to VI, peaking at 1.410 ng/g. ABA (Fig. 5c) showed a significant increase from stages I to VI, up to the highest level at stage VI (151.573 ng/g). The significant increasing in IAA from stages IV to V supported petal expansion, which likely led to the dilution or redistribution of pigments during cell expansion, contributing to the gradual fading of flower color.
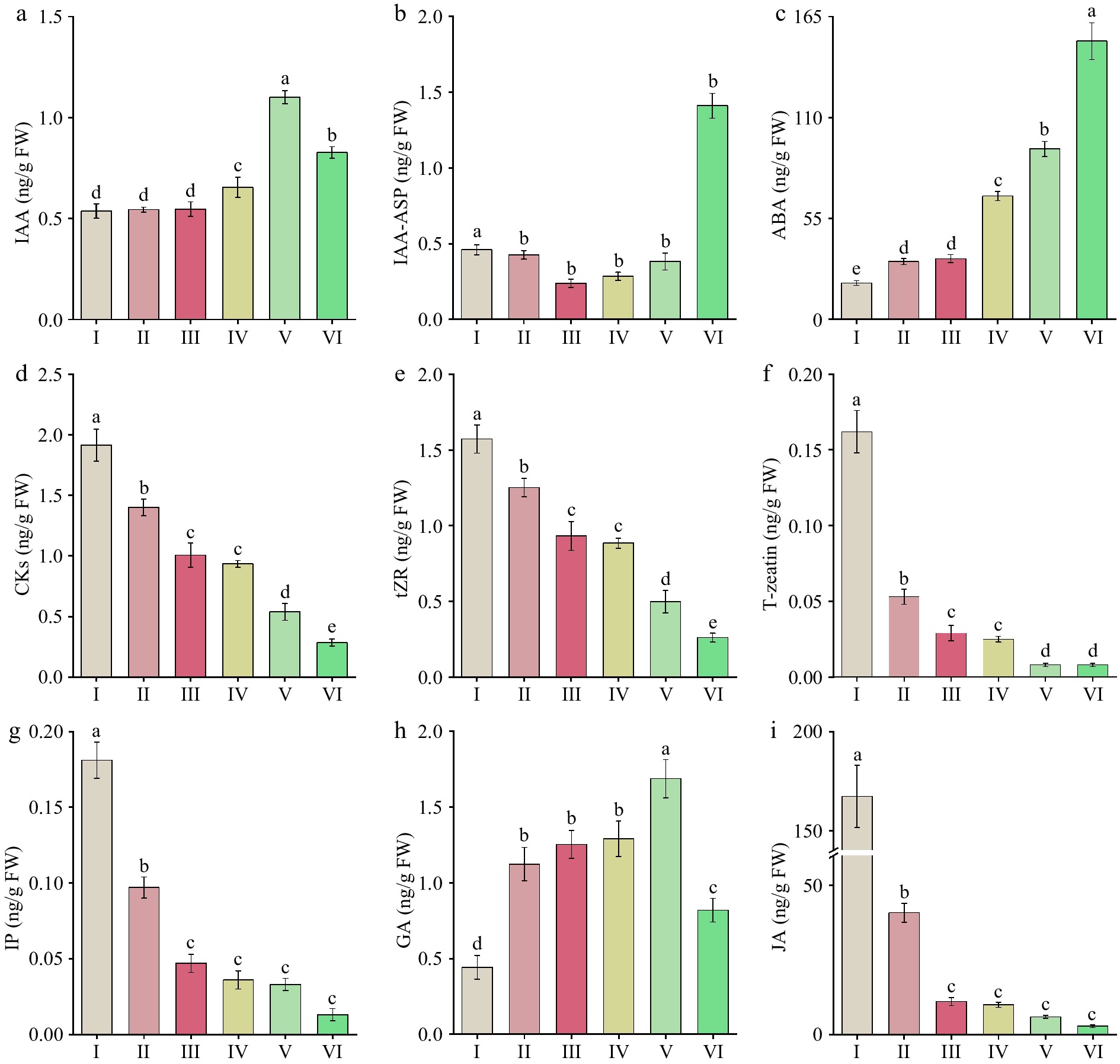
Figure 5.
Endogenous hormone levels in 'Tongzimian' petals at six different flowering stages. (a) Auxin (IAA). (b) Indole-3-acetic acid-aspartic (IAA-ASP). (c) Abscisic acid (ABA). (d) Cytokinins (CKs). (e) Trans-zeatin riboside (tZR). (f) Trans-zeatin (T-zeatin). (g) Isopentenyl adenine (IP). (h) Gibberellins (GA). (i) Jasmonic acid (JA). Different lowercase letters indicate significant differences (p < 0.05, ANOVA).
CKs (Fig. 5d) were at the highest level at stage I (1.945 ng/g), then significantly decreased to 0.350 ng/g, which was the lowest level through the whole flowering process. The changes in tZR (Fig. 5e) had the same trend as CKs, with a significant decline from stage I (1.572 ng/g) to stage VI (0.261 ng/g). T-zeatin (Fig. 5f) also showed a significant decline, peaking at stage I (0.162 ng/g) and remaining at low levels from stage V to VI (0.008 ng/g).
The content of IP (Fig. 5g) followed a similar downward trend, with the highest concentration at stage I (0.181 ng/g) and the lowest at stage V and VI (0.013 ng/g). GA (Fig. 5h) increased significantly from stage I to II, but remained relatively stable from II to VI. A significant increase occurred at stage V, reaching its peak (1.686 ng/g), followed by a notable decrease at stage VI. JA (Fig. 5i) showed an overall declining trend, with a significantly higher level at stage I (167.294 ng/g) compared to other stages. It then decreased significantly at stage II, and remained stable from stage III to VI. SA (Fig. 6a) showed an overall downward trend, with significantly higher levels at stage I (330.965 ng/g). There were no significant changes between stage III to V, and the lowest level was at stage VI (7.047 ng/g). The peak values of IP, JA, and SA during the early stages likely provided protective functions. The two peaks of GA at stages II and V supported petal expansion and the full opening of the flower.
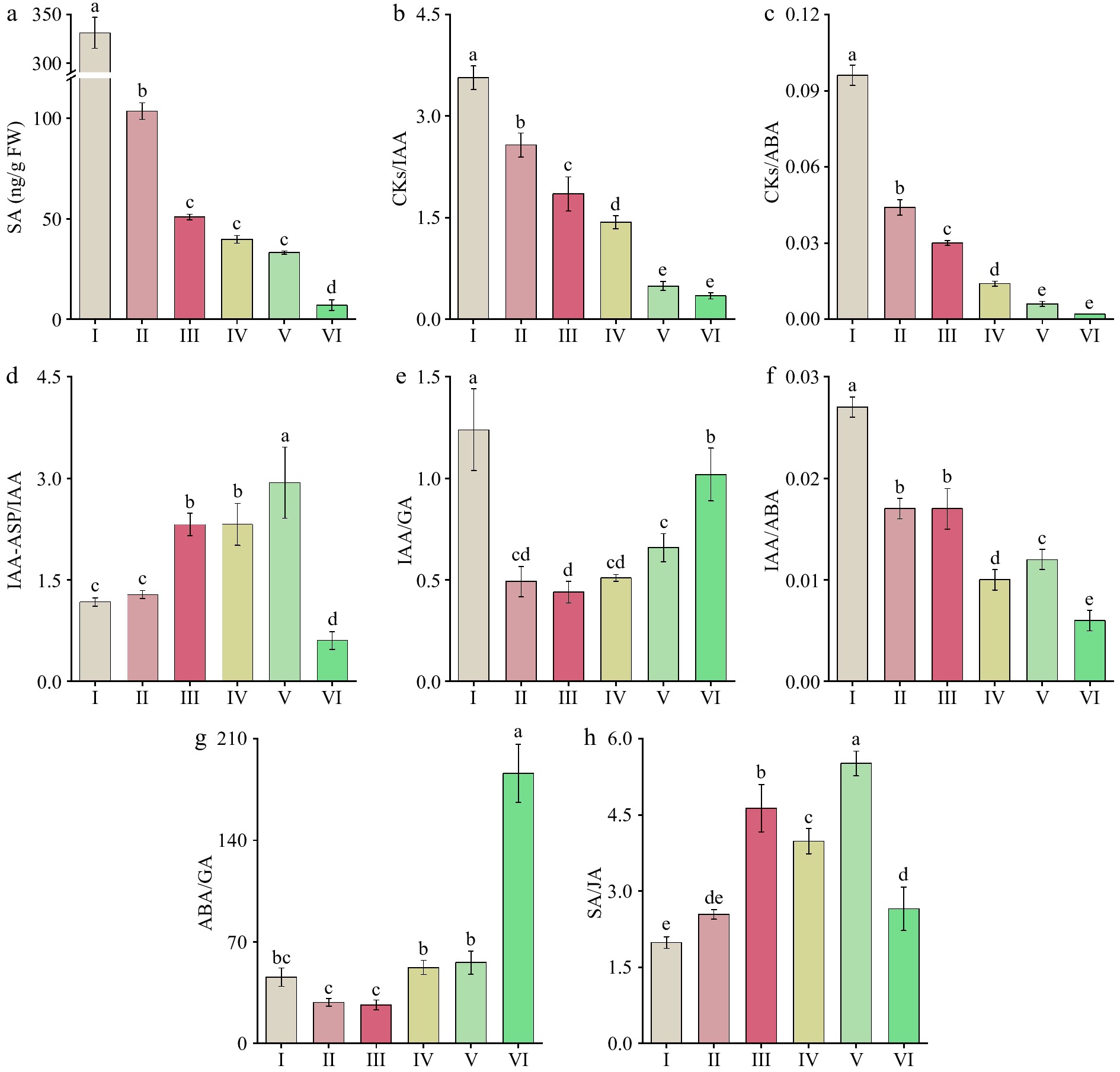
Figure 6.
Changes in endogenous hormone levels and their ratios at six different flowering stages of 'Tongzimian'. (a) Salicylic acid (SA). (b) Cytokinins/Auxin (CKs/IAA). (c) Cytokinins/Abscisic acid (CKs/ABA). (d) Auxin/Indole-3-acetyl-aspartic acid (IAA/IAA-ASP). (e) Auxin/Gibberellin (IAA/GA). (f) Auxin/Abscisic acid (IAA/ABA). (g) Abscisic acid/Gibberellin (ABA/GA). (h) Salicylic Acid/Jasmonic acid (SA/JA). Different lowercase letters indicate significant differences (p < 0.05, ANOVA).
Changes in the ratios of endogenous hormones in 'Tongzimian' petals at six different flowering stages
-
The content and ratios of endogenous hormones during the flowering stages of 'Tongzimian' as shown in Fig. 6. The CKs/IAA (Fig. 6b) ratio was the highest during stage I (3.564), then decreased significantly to the lowest value of 0.346 at stage VI, which decreases in the CKs/IAA ratio indicates a reduction in CKs relative to IAA, suggesting a decrease in cell division and growth-promoting activities as the flower developed. The CKs/ABA (Fig. 6c) ratio also peaked at stage I (0.096) and then decreased significantly to the lowest value of 0.002 at stage VI, which reflects a shift in the hormonal balance, with CKs decreasing relative to ABA during flower development. This suggests a transition from growth promotion (CKs) to stress and senescence (ABA) as the flower developed. The IAA/IAA-ASP (Fig. 6d) ratio increased significantly from stage I to IV, peaking at stage V (2.935) and then decreasing to its lowest point at stage VI (0.604), which suggests that the metabolism of IAA was slower during the early stages of flower development, but accelerated in the later stages (especially at stage V), possibly supporting the final stages of petal expansion and pigment distribution.
The IAA/GA (Fig. 6e) ratio was the highest during stage I (1.239) and significantly decreased over time, reaching its lowest value at stage III (0.439). The ratio then significantly increased again at stages V and VI, which aligns with the changes in flower color and suggests that IAA and GA contribute to the petal expansion and the redistribution of pigments. The reversed trend of this ratio in comparison to anthocyanin content indicates a connection between IAA and GA in regulating pigment distribution and flower color. The IAA/ABA (Fig. 6f) ratio was the highest at stage I (0.027), then significantly decreased to the lowest value at stage VI (0.006). This trend highlights the shift from IAA-dominated activities in the early flowering stages (promoting growth and expansion) to ABA-dominated processes in the later stages (involved in stress and aging).
The ABA/GA (Fig. 6g) ratio decreased significantly from stage I to stage III, reaching its lowest value at stage III (26.609), and then increased significantly through to stage VI, peaking at 186.094. This shift suggests that the balance between ABA and GA influences the transition from flower growth to senescence, with ABA playing a dominant role in the aging process. The SA/JA (Fig. 6h) ratio was the lowest during stage I (1.987) and then increased significantly, maintaining high levels through stages III to V. However, it declined significantly at stage VI, indicating a shift toward defense and senescence during the later stages of flower develops.
In summary, during the early bud stage (I to II), the higher ratios of CKs/IAA, CKs/ABA, and IAA/GA suggest that cell division and growth were more active, which corresponded with the lighter petal color. The hormonal balance supported cell proliferation and the formation of basic flower morphology. In the flower blooming stage (IV to V), the higher IAA/IAA-ASP and IAA/GA ratios supported petal expansion, with GA playing a significant role in facilitating the redistribution of pigments, leading to more uniform petal color. During stage VI, the significantly higher ABA/GA and CKs/ABA ratios indicated that ABA dominated the aging process, leading to color fading, while the decline in SA/JA ratios also pointed to the onset of defensive and senescence mechanisms.
Correlation analysis of physiological and biochemical indicators at six different flowering stages of 'Tongzimian' flowers
-
The correlations among the pigment composition and content, endogenous hormones, osmotic regulators, and antioxidant enzyme activities in the petals of 'Tongzimian' flowers at six different flowering stages were analyzed (Fig. 7). There are significant correlations among the endogenous hormones. IAA is significantly negatively correlated with CKs (r = −0.724), and ABA is also significantly negatively correlated with CKs (r = −0.875). On the other hand, SA shows a significant positive correlation with CKs (r = 0.886), and SA is highly positively correlated with JA (r = 0.995). CKs are significantly positively correlated with carotenoids (r = 0.724), flavonoids (r = 0.912), and anthocyanins (r = 0.836), suggesting that CKs may play a promotive role in regulating pigment synthesis in the petals. ABA shows a significant negative correlation with carotenoids (r = −0.843), flavonoids (r = −0.839), and anthocyanins (r = −0.898). Therefore, ABA is primarily involved in petal senescence and color fading, exhibiting a significant negative correlation with pigment components. This suggests that an increase in ABA leads to a reduction in pigment content, resulting in the gradual fading of the petal color.
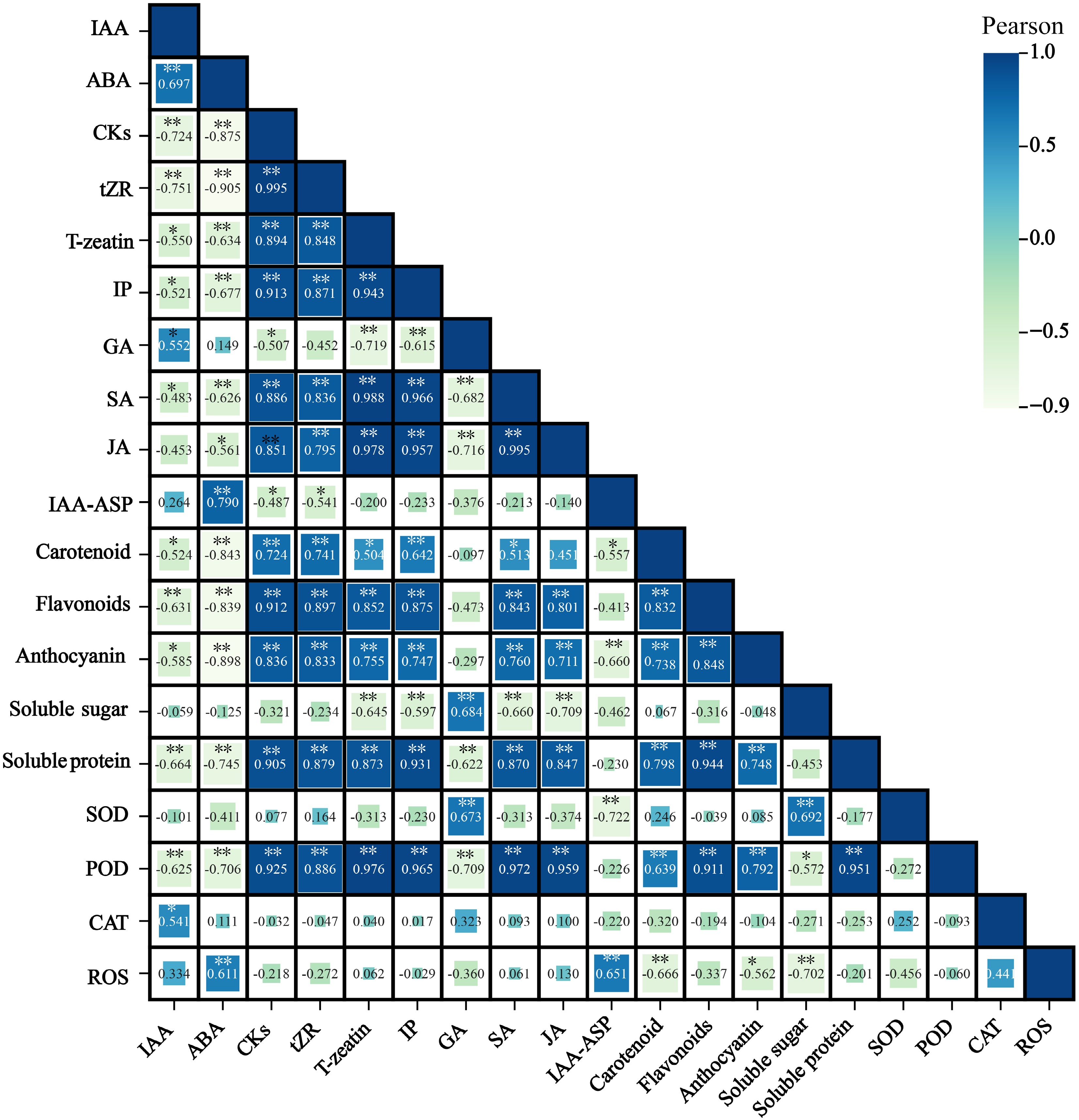
Figure 7.
Correlation between petal pigment composition and content, endogenous hormones, osmotic regulatory substances, and antioxidant enzymes at flowering stages of 'Tongzimian'. Auxin (IAA), Abscisic acid (ABA), Cytokinins (CKs), Trans-zeatin riboside (tZR), Trans-zeatin (T-zeatin), Isopentenyl adenine (IP), Gibberellins (GA), Salicylic acid (SA), Jasmonic acid (JA), Indole-3-acetic acid-aspartic (IAA-ASP), Superoxide dismutase (SOD), Peroxidase (POD), Catalase (CAT), Reactive oxygen species (ROS). Darker colors indicate positive correlations, while lighter colors indicate negative correlations. * Indicates significance at the 0.05 level, and ** indicates significance at the 0.01 level.
SA and JA work synergistically in the flower's defense mechanisms, potentially providing protection during the flowering period and aiding in pigment accumulation. Soluble sugars and proteins play an active role in osmotic pressure regulation, cell turgor, and pigment synthesis, showing a significant positive correlation with GA, POD, and pigment components. Both POD and SOD are closely related to pigments and antioxidant defense, with high levels of POD contributing to pigment stability and antioxidant capacity. Soluble sugars show a significant positive correlation with GA (r = 0.684) and SOD (r = 0.692), and a significant negative correlation with SA (r = −0.660), JA (r = −0.709), and ROS (r = −0.702). Soluble proteins are significantly positively correlated with CKs (r = 0.905), SA (r = 0.870), JA (r = 0.847), POD (r = 0.951), carotenoids (r = 0.798), flavonoids (r = 0.944), and anthocyanins (r = 0.748). These positive correlations indicate that proteins may play an auxiliary role in pigment synthesis and accumulation, enhancing antioxidant capacity and stress tolerance. Soluble proteins also show a significant negative correlation with ABA (r = −0.745).
POD is significantly negatively correlated with IAA (r = −0.625) and ABA (r = −0.706), but is significantly positively correlated with CKs (r = 0.925), SA (r = 0.972), and JA (r = 0.959). SOD is significantly positively correlated with GA (r = 0.673) and soluble sugars (r = 0.692), while it is significantly negatively correlated with IAA-ASP (r = −0.722). ROS is significantly negatively correlated with carotenoids (r = −0.666) and anthocyanins (r = −0.562). These correlation data highlight the complex interactions between plant hormones (such as IAA, ABA, and CKs), pigments (such as flavonoids and anthocyanins), nutrients (such as soluble sugars and proteins), and antioxidant enzymes (such as POD) during the flower development stages of 'Tongzimian'.
-
C. reticulata is famous for its evergreen shrub/tree shape, and large and diverse flower patterns, also known for its gorgeous flower color. However, its flower colors are generally limited and lack yellow, light, and other multicolored flowers[1−3]. Due to the absence of suitable models, research on the light-colored flowers of C. reticulata has been constrained. To address this gap, we have chosen the cultivar 'Tongzimian', which has the lightest color among C. reticulata cultivars, as the research model to reveal the physiological mechanisms underlying its nearly white petals. During the flower development and blooming process, the petal color of 'Tongzimian' transitions from yellow-green (I) to pink (III) and gradually lightens to almost white with a slight red tinge at full bloom. This study analyzes the phenotypic color differences, cell morphology, ultrastructure, and physiological-biochemical factors at flowering stages to preliminarily elucidate the physiological mechanisms behind the formation of nearly white flowers in 'Tongzimian'. The findings provide a theoretical basis for further molecular mechanism studies and lay a solid foundation for optimizing flower color breeding in C. reticulata.
During the flowering process, the conical structure of the epidermal cells in petals enhances the absorption of light by anthocyanins, thereby intensifying the petal color[33]. In this study, we observed changes in the cell morphology at the tops and bases of the epidermal cells on the petals of 'Tongzimian' at different stages: I, III, and V. From stage I to stage III, the cell surface gradually transforms from smooth to a complex, undulating structure with numerous wrinkles, during which the petal color is the deepest. As the flower transitions from stage III to stage V, the texture of the top epidermal cells weakens significantly, with some areas showing depressions. At this stage, the pink color gradually fades to white. These structural changes are believed to affect light absorption and reflection, indirectly influencing pigment accumulation and, consequently, the petal coloration. Similar phenomena have been observed in other plants. For example, in Paeonia suffruticosa, the epidermal cell striations are the densest when chlorophyll content is at its highest, and the striations become sparse when chlorophyll content is lowest[7].
The formation of flower color in plants is closely related to structural changes in the plastids within petal cells. Studies have shown that during petal development, chloroplasts, and amyloplasts gradually transform into chromoplasts, a process that is one of the key mechanisms responsible for the gradual color change of petals and fruits from yellow to orange[34,35]. In this study, we observed the accumulation of chromoplasts in the petals of 'Tongzimian' during stage III, resulting in a pink color. However, as the flower reached stage V, chromoplasts gradually degraded, and the petals turned white. This observation is consistent with the findings of Haratym et al., who reported similar results in Centaurea cyanus during its full flowering stage[36]. Additionally, during the transition of tobacco floral nectaries from green to orange, chloroplasts are converted into chromoplasts[37]. The combined effect of these cellular morphological changes promotes the color transition in 'Tongzimian' petals during flowering, providing a foundation for further research into its color presentation mechanism.
Flower color is primarily determined by the types of pigments present in the petals and their relative concentrations, including flavonoids, carotenoids, chlorophylls, and alkaloids. In most ornamental plants, changes in petal color during flower development are driven by variations in anthocyanin accumulation. For instance, in chrysanthemums such as 'Nannong Lvdong', 'Ibis Sunny', and 'Arctic Queen', the red color observed in the later stages of flowering is attributed to the accumulation of anthocyanins[38]. In peonies, the color change from coral to pink and then to light yellow in cultivars like 'Coral Sunset' and 'Pink Hawaiian Coral' is caused by fluctuations in anthocyanin content[39]. Similarly, in Malus hupehensis, the decrease in anthocyanin content during flowering results in a color change from red to white[40]. Generally, an increase in anthocyanin content deepens the red color of the petals, while a decrease leads to a lighter color. This trend is consistent with the color changes observed in 'Tongzimian', where anthocyanin content increases as the a* value rises, directly influencing the depth of red in the petals.
It is well known that anthocyanins not only scavenge free radicals but that a certain level of ROS can induce anthocyanin production[41]. Therefore, the reduction in anthocyanin content observed in the early stages of flowering in 'Tongzimian' may play a role in scavenging ROS in the petals. Later, a certain level of ROS could trigger the production of anthocyanins, causing the petals to turn pink. In the later stages, however, the significant increase in ROS levels leads to the involvement of anthocyanins in ROS scavenging, resulting in the fading of the petal color. At the same time, enzymes such as SOD, POD, and CAT all play crucial roles in ROS scavenging. These enzymes may work synergistically and have different action times depending on the type of ROS they target[42]. In this study, the changes in the activity of SOD, POD, and CAT complement the fluctuations in anthocyanin content. During periods of high anthocyanin content, the demand for antioxidant enzymes may be lower, while during periods of decreasing anthocyanin levels, the activity of antioxidant enzymes (especially SOD) increases, compensating for the reduced antioxidant capacity. Thus, anthocyanins and antioxidant enzymes exhibit a complementary mechanism during the different flowering stages of 'Tongzimian', ultimately leading to the observed color changes.
Anthocyanins typically present red, purple, or blue hues, while carotenoids generally appear in yellow, orange, or red colors[14]. The changes in carotenoid and anthocyanin content often work in tandem to regulate flower color and fruit development in many crops[43]. Their relative content and distribution directly influence the color expression of the petals. In the petals of 'Tongzimian', we also detected carotenoids, but their content was consistently lower than that of anthocyanins during each stage, which contributed to the dynamic pink coloration of the petals. Notably, the carotenoid content was relatively high in the early stages, likely giving the petals a yellow-green color. This trend is consistent with the changes in carotenoid levels observed in plants such as petunia, chrysanthemum, and rose during flower development[44]. Therefore, one future research direction is to explore how to regulate the balance of anthocyanins and carotenoids in the petals to develop cultivars of camellia with yellow hues. Additionally, we observed a significant decrease in flavonoid content, although it remained notably higher than that of anthocyanins and carotenoids. Further analysis of the specific components and their variation during the different flowering stages of 'Tongzimian' will be necessary for a more comprehensive understanding.
In the flavonoid metabolic pathway, soluble sugars and soluble proteins provide the energy and substrates necessary for anthocyanin synthesis. Variations in soluble proteins and sugars result in changes in carbohydrate content, thereby playing a significant but indirect role in the manifestation of petal color[45]. Soluble sugars and soluble proteins act as precursors or signaling molecules for anthocyanin synthesis and degradation, influencing the production of anthocyanins to some extent. In our study, we observed that soluble sugar content in 'Tongzimian' petals followed a trend of initial increase followed by a decrease across the six stages of flower development, which aligns with the changes in anthocyanin content from the coloring phase to the initial blooming stage. Additionally, the soluble protein content showed a significant decrease and then leveled off, which correlates with the degradation of petal pigments and the reduction in starch granules observed in the cellular ultrastructure. The exact cause of this decrease requires further verification. During 'Tongzimian' flower development, the content of soluble sugars and soluble proteins was found to be negatively correlated, a finding consistent with Bao et al.'s research on peony petals[45].
Endogenous hormones regulate flower color by affecting the accumulation of anthocyanins, leading to the display of different flowerings. It has been reported that IAA can suppress anthocyanin accumulation to some extent[20,21], and ABA positively regulates anthocyanin biosynthesis in the red-striped petals of C. japonica 'Joy Kendrick'[22]. In this study, ABA showed a significant negative correlation with anthocyanins (r = −0.898). The accumulation of ABA may inhibit the further synthesis of anthocyanins by triggering a negative feedback mechanism. Additionally, this negative correlation could be due to interactions between ABA and other hormones. In the correlation analysis, we also found that dynamic changes in endogenous hormones, such as CKs and IP, were significantly associated with changes in anthocyanin content (p < 0.01). Furthermore, anthocyanins are important antioxidants that scavenge ROS, and during the later stages of flower development, increased ROS levels could lead to a decrease in anthocyanin content.
-
This study investigates the cellular morphology and physiological mechanisms of 'Tongzimian' petals at different coloring stages of flower development. It was found that the dynamic changes in anthocyanin and carotenoid content, micro-adjustments in cellular morphology, and the temporal regulation of endogenous hormones collectively drive the transformation of petal color from yellow-green to pink and ultimately to white. Additionally, changes in antioxidant enzymes and ROS contribute to the alteration of petal coloration. The findings of this study provide a preliminary theoretical foundation for understanding the color development mechanism in 'Tongzimian', offering valuable insights for flower color breeding lighter-colored cultivars of C. reticulata in the future.
This research was funded by the National Natural Science Foundation of China (32260416), Yunnan agricultural basic research joint special general project (202101BD070001-095), and Yunnan Province Xingdian Talent Project 'Youth Talent Special Project' Support Project (YNQR-QNRC-2-19-104). Funders have no role in the design and publication of the manuscript.
-
The authors confirm contribution to the paper as follows: study conception and design: Xiao H, Geng F, Wang Z; data collection and analysis: Xiao H, Nie R, Zeng Y, Chen S, Yan F, Wu X, Wang Z, Geng F; draft manuscript preparation: Xiao H, Geng F. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Correlation analysis of petal color phenotypic data at flowering stages of 'Tongzimian'.
- Supplementary Table S2 Petal anatomical parameters of 'Tongzimian'.
- Supplementary Fig. S1 The pictures of C. reticulata 'Tongzimian'. (a) I (yellow-green, bud stage). (b)III (distinct pink, exposed color stage). (c) IV (light pink, early bloom stage). (d) (whitish, full bloom stage). (e) The morphological characteristics of the plant during the flowering period of 'Tongzimian'.
- Supplementary Fig. S2 Petal anatomical structure of 'Tongzimian'. Scale Bar=40 μm; Ue: Upper epidermis; Le: Lower epidermis; PT: Petal thickness; Pt: Parenchyma tissue; Vb: Vascular bundle.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xiao H, Nie R, Zeng Y, Chen S, Yan F, et al. 2025. The physiological mechanism of the flower color formation of Camellia reticulata 'Tongzimian'. Ornamental Plant Research 5: e024 doi: 10.48130/opr-0025-0022 

The physiological mechanism of the flower color formation of Camellia reticulata 'Tongzimian'
- Received: 09 December 2024
- Revised: 01 March 2025
- Accepted: 11 March 2025
- Published online: 09 June 2025
Abstract: 'Tongzimian', is the lightest colored cultivar of C. reticulata flowers, to explore the physiological mechanisms of its flower color formation is very important for camellia breeders. In this study, the color difference values of flower color, cell structure, and ultrastructure changes, as well as the physiological changes were analyzed by using the petals from six different flowering stages. The results showed that changes in color difference values were consistent with the phenotypic changes of the petals. The size and morphology of petal epidermal cells varied significantly across six different flowering stages, particularly with the apex petal cells transitioning from smooth to complex textures. Flavonoid levels exhibited an overall decreasing trend, while changes in anthocyanin content corresponded with petal color alterations, reaching 14.952 μg/g during the exposed color stage (III) when petal color was the deepest. Osmotic regulatory substances and antioxidant enzymes were also closely related to color changes, especially catalase (CAT) and reactive oxygen species (ROS), which decreased first and then increased. Furthermore, endogenous hormones, such as abscisic acid (ABA) and cytokinins (CKs) indicated significant changes across different stages and were significantly correlated with pigment composition and petal color changes (p < 0.01). Interestingly, ABA is significantly negatively correlated with anthocyanins (r = −0.898). Therefore, the study has preliminarily revealed the physiological mechanisms of color formation in 'Tongzimian', providing a theoretical basis for further analyzing the molecular mechanism of color formation in 'Tongzimian' and enriching camellia color breeding.
-
Key words:
- C. reticulata 'Tongzimian' /
- Flower color /
- Anthocyanidin /
- Physiological mechanisms