









-
The world is full of fragrances and scents, the fragrances usually derived from the presence of volatile compounds that plants produce and emit. In nature, plants synthesize numerous volatile substances with diverse biological properties and functions. These substances are generally small-molecular compounds characterized by low molecular weight, low boiling point and lipophilic properties[1]. Throughout the growth and development of plants, different tissues such as flowers, fruits, leaves, and rhizomes release a plethora of volatiles. Each plant has its unique aroma attributed to the distinct combinations and concentrations of these volatiles[2]. These aroma compounds play a pivotal role in various aspects, enabling plants to interact with other organisms and their surrounding environment. Moreover, they contribute to attracting pollinators and seed dispersers. For humans, these aroma compounds enrich our daily lives, permeating our food, health products, and cosmetics. Given their agricultural and economic significance, we focused on three parts: the major biosynthesis pathways of aroma compounds, the role of aroma compounds, and enhancing plant aroma utilizing CRISPR/Cas9 system.
-
Aroma compounds, generated through primary and secondary metabolism, are essential for the captivating scents in plants. According to biosynthetic pathways, aroma compounds are mainly derived from four major pathways: fatty acids, amino acids, terpenoids and carotenoid pathways. These pathways give rise to a diverse range of compounds, such as alcohols, aldehydes, esters, ketones, terpenes, sulfur compounds and so on (Fig. 1).
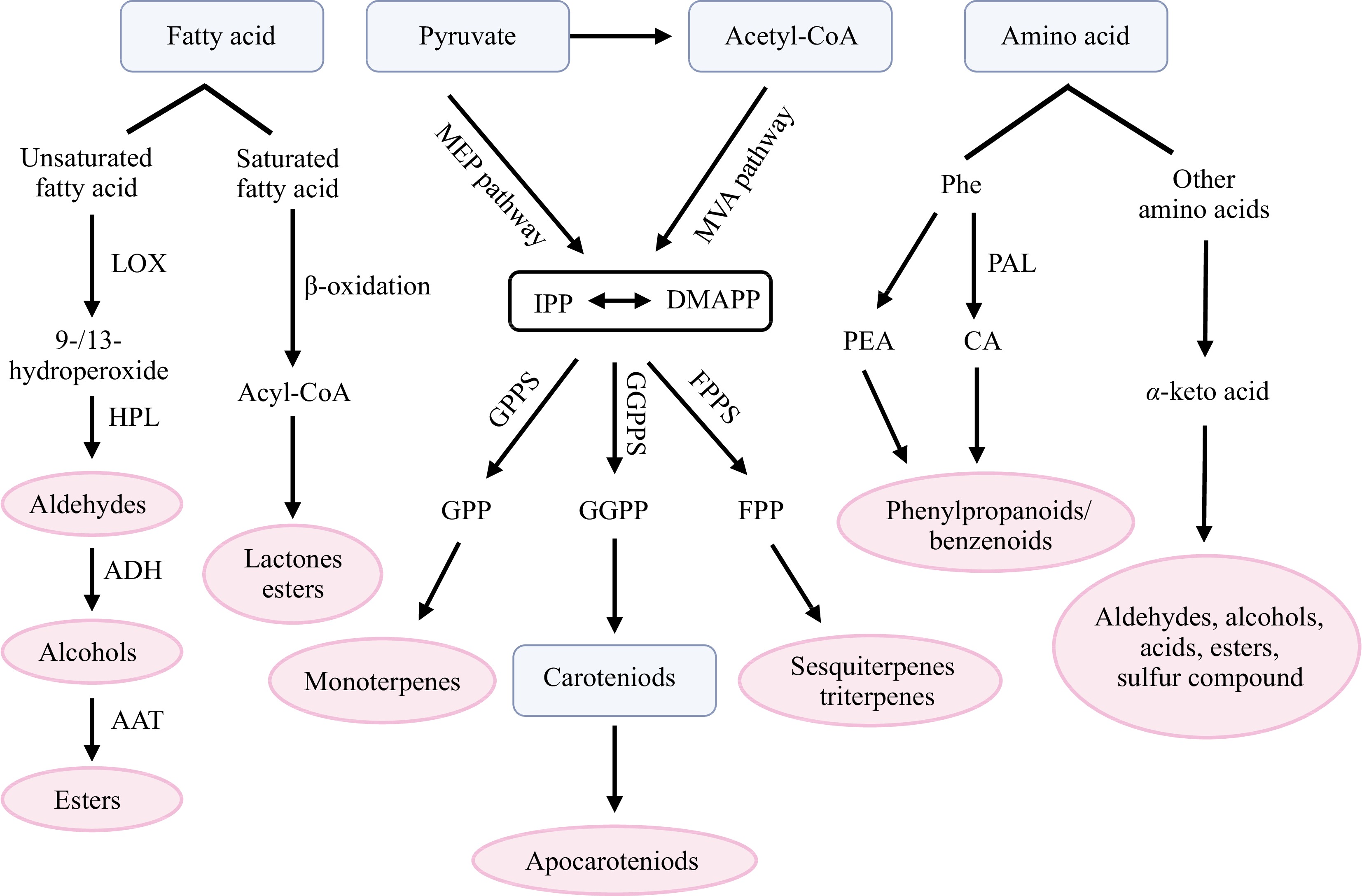
Figure 1.
Major biosynthetic pathways of aroma compounds in plants. AAT, alcohol acyltransferase; Acetyl-CoA, acetyl coenzyme-A; ADH, alcohol dehydrogenase; CA, cinnamic acid; DMAPP, dimethylallyl diphosphate; FPP, farnesyl diphosphate; FPPS, farnesyl pyrophosphate synthase; GGPP, geranylgeranyl diphosphate; GGPPS, Geranylgeranyl pyrophosphate synthase; GPP, geranyl diphosphate; GPPS, geranyl pyrophosphate synthase; HPL, hydroperoxide lyase; IPP, isopentenyl diphosphate; LOX, lipoxygenase; MEP, methylerythritol 4-phosphate; MVA, mevalonate; PAL, phenylalanine ammonia lyase; PEA, phenylethylamine; Phe, phenylalanine.
-
Fatty acids, such as linoleic acid and linolenic acid, are widely present in plants. These fatty acids serve as precursors of various aliphatic compounds, including alcohols, aldehydes, ketones, acids, esters and lactones, which are found ubiquitous in plants. In most fruits, fatty acid-derived compounds contribute to the flavor and affect human preferences. The formation of these flavor compounds basically are formed by β-oxidation and the lipoxygenase (LOX) pathway[3]. β-oxidation is a common pathway for fatty acid degradation in the peroxisomes, leading to the successive removal of C2 units from the parent fatty acid, that produce lactones and esters[4,5].
Lipoxygenase pathway is the metabolic pathway of plant polyunsaturated fatty acids, for example, linolenic acid (LeA) and linoleic acid (LA). Saturated and unsaturated volatile C6, C9 aldehydes and alcohols are produced via LOX pathway. These C6 aldehydes, alcohols, and corresponding esters are called "Green leaf volatiles" (GLVs), such as hexanal, hexanol and hexenyl acetate, which are important contributors to the flavor of fruits and vegetables[6]. In daily life, the smell of cut grass is also caused by GLVs[7] . For example, cis-3-Hexen-1-ol is an important volatile compound in plants, such as green tea, grapes, passion fruits, and toona sinensis[8−11]. This compound shows the smell of 'cut grass' or 'green grassy', contributes to the 'green, grass and freshness' of food flavor[12]. In the lipoxygenase pathway, LOX can catalyze unsaturated fatty acids into hydro peroxides. During this process, oxygen can be added to the 9th carbon atom of the hydrocarbon chain of substrates, or it can be added to the 13th carbon atom, so the plant LOX are divided into 9-LOX and 13-LOX. Then hydroperoxides serve as a reaction substrate, under the action of hydroperoxide lyase (HPL) enzymes, 9-hydroperoxide is cleaved to form a nine-carbon volatile and a nine-carbon oxoacid, and 13-hydroperoxide will form a six-carbon volatile and a 12-carbon oxoacid[13]. In this process, HPL is a member of the cytochrome P450 family, and catalyzes the lipid hydroperoxides without any co-factor[14,15]. The catalytic products of HPL are not only one of the main components of plant odor, but also related to disease and insect resistance[6,16−18]. According to their different substrates, plant HPL are divided into two enzymes: 9-HPL and 13-HPL. Research also showed that there is a class of 9/13 HPL that can catalyze both 9-HPOT and 13-HPOT[7]. Via the HPL pathway, short-chain aldehydes and oxyacids are synthesized, then aldehydes are catalyzed by alcohol dehydrogenase (ADH) to produce C6 alcohols through the removal of hydrogen. These C6 alcohols can be further converted into esters, through the esterification reaction between alcohols and acyl-CoAs catalyzed by alcohol acyltransferase (AAT)[19−21].
-
The metabolism of amino acids can produce aliphatic and aromatic compounds such as aldehydes, acids, alcohols, and esters. In fruits, the characteristic aroma components of 'fruity' scent are mostly produced from amino acid metabolism[6]. Amino acid derived aroma compounds can be divided into two branches. One way is to use phenylalanine (Phe) as substrate, and synthesize many aroma phenylpropanoids/benzenoids. Phe can be catalyzed by phenylalanine ammonia lyase (PAL) to cinnamic acid (CA), and then through different metabolic pathways to form methyl salicylate, guaiacol, catechol or eugenol[22,23]. Phe can also produce phenylpropanoid-related aroma compounds via other pathways. For example, in tomato, amino acid decarboxylases enzymes LeAADCs catalyze decarboxylation of Phe to phenylethylamine (PEA), which can be further catalyzed to synthesize 2-phenylacetaldehyde, 2-phenylacetonitrile, and 1-nitro-2-phenylethane[24]. Using 2-phenylacetaldehyde as substrate, 2-phenylethanol was generated from catalytic reduction of phenylacetaldehyde reductases LePARs, both of them are important contributors to flavor[25]. These results indicated that it is practicable to alter phenylalanine-based aroma volatiles in plants by regulating the expression of related enzymes.
Other amino acids, such as alanine, valine, leucine, isoleucine, and methionine, are also involved in some aroma compounds biosynthesis, producing aldehydes, alcohols, esters, acids, and nitrogen- and sulfur-containing volatiles. Firstly, amino acids can produce α-keto acid through a deamination or transamination. Subsequent reactions like decarboxylation, reductions, oxidations, and esterification lead to the formation of aldehydes, acids, alcohols and esters[6]. A kind of typical products are sulfur compounds, which are found in some fermented foods (cheese, beer, wine) and many tropical-subtropical fruits like durian, grapefruit, guava, lemon. Various volatile sulfur compounds have been identified and contribute to the juicy, fresh, and authentic aroma[26,27].
-
In plants, a large class of aroma compounds are derived from terpenoid pathways, which can be divided into two pathways, the mevalonate (MVA) pathway, active in the cytosol, and the methylerythritol 4-phosphate (MEP) pathway, active in the plastids[28,29]. Although the MEP and MVA pathways start from different substrates (acetyl CoA or pyruvate), they generate the same terpene precursors isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (DMAPP), the two five-carbon compounds are the basic unit for biosynthesis of terpenoids. IPP and DMAPP are combined with each other by the activity of prenyltransferases. With the catalysis of geranyl pyrophosphate synthase (GPPS), geranyl pyrophosphate (GPP) is synthesized by the condensation of one IPP molecule and one DMAPP molecule, a precursor of monoterpene substance. Farnesyl pyrophosphate (FPP) is a precursor of sesquiterpenes, which is synthesized by condensation of two IPP molecules with one DMAPP molecule results through farnesyl pyrophosphate synthase (FPPS). Geranylgeranyl pyrophosphate synthase (GGPPS) can condense three IPP molecules with one DMAPP molecule to produce GGPP, a precursor of diterpenoids. Various terpenes are synthesized from substrates such as DMAPP, GPP, FPP and GGPP under the catalysis of terpene synthase (TPS)[30]. Terpenoids (also known as isoprenoids), formed by oxidation of the terpenes, are the largest class of plant secondary metabolites[31].
In nature, terpenes and terpenoids are widely present in plants. Almost all the volatile components of floral aroma contain terpenes and terpenoids, and they are also the characteristic aroma components of some fruity aroma[32]. In addition, terpenes and terpenoids are one of the important components in the pharmacological effects of plant herbs. The aroma of citrus fruit mainly comes from terpenes and terpenoids. Limonene is the most abundant aroma substance in citrus fruit peel oils with antioxidant and anti-inflammatory properties[33]. β-Myrcene is also an abundant monoterpene in many plant species, such as cannabis and hops[34]. In apples, various terpenes have been identified, such as germacrene, β-caryophyllene, linalool and α-pinene[35]. Roses, an ornamental plant, also can be used to extract essential oils for perfumes and cosmetics, have high economic and health values. Research showed that the scent of roses is composed of hundreds of volatile molecules, one of the most important components are terpenes, in some cultivars, monoterpenes make up to 70% of the aroma content, including geraniol, nerol, and citronellol[36].
-
Carotenoid-derived aroma compounds are common in some fruits and vegetables, such as tomato and grape[37,38]. Usually, the biosynthesis of these compounds contains three steps: dioxygenase cleavage, then the formation of aroma precursors through enzymatic transformations of the cleavage products, and acid-catalyzed conversions of the precursors into the aroma compounds[39]. Carotenoid cleavage products (apocarotenoids), including 6-methyl-5-hepten-2-one (MHO), ionone, β-cyclocitral, and geranylacetone, are important compounds to the flavor of flowers and vegetables[40]. MHO is an important flavor component in plants, such as tomato, guava and papaya[41,42]. Ionone widely exists in fruits and vegetables that contain β-carotene, including α-ionone and β-ionone[43,44]. α-ionone has a woody floral scent and a honey smell, β-ionone has a woody floral scent, and is a major contributor to rose aroma, both are widely used as aroma compounds in various industries, including flavoring agents and cosmetics[45].
These apocarotenoids are derived from the cleavage of carotenoids by carotenoid cleavage dioxygenases (CCDs), forming an aldehyde or ketone at the site of cleavage[46]. CCDs often exhibit diversity of substrates and have different subcellular localization. For example, CCD1, the only CCD located in the cytosol, is involved in the formation of flavor and aroma volatiles. In plants, CCD1 usually cleaves the 9–10 and 9'–10' double bonds of their carotenoid substrates, and it can also act on 5–6 and 5'–6' double bonds of carotenoids[46]. Acting on different substrates, such as lutein, lycopene, and zeaxanthin, this cleavage way can produce various apocarotenoid products[47]. In tomato, these carotenoid-derived volatiles may greatly affect aroma and scent at low concentrations. For example, LeCCD1 can cleave zeaxanthin or lutein into 3-hydroxy-β-ionone, also generate geranylacetone via cleavage of phytoene, when lycopene serves as a precursor, pseudoionone is produced with LeCCD1[48].
-
Aroma compounds fill our lives with a diverse array of scents, and they play crucial roles in various fields. Here, we outlined the significance of aroma compounds for humans, their interactions with other species, and their role in attracting pollinators.
-
For humans, the aroma volatiles from trees and flowers emit distinctive smells that promote relaxation. These aroma compounds, with their good biological activity and pleasant flavors, find widespread applications in medicine, health care, cosmetics, and the food industry. Rice is one of the staple foods around the world. Aroma is the key criteria to evaluate the quality and value of rice. According to previous studies, rice aroma is composed of more than 500 different volatile compounds including aldehydes, festers, alcohols, ketones, organic acids, phenols, and other compounds[49]. These diverse compounds lead to 'popcorn', 'green', 'fruity', 'roasty', 'bitter' and 'nutty' scents in conventional rice cultivars. For instance, 2-acetyl-1-pyrroline (2-AP) is the major compound in fragrant rice that contributes to popcorn aroma; green smell is mainly due to the presence of aldehydes, some ketones and alcohol; fruity aroma is contributed by heptanone, ketones, and 6-methyl-5-hepten-2-one; (E,E)-2,4-nonadienal, 2-pentyfuran, 4-vinyl guaiacol, and benzaldehyde provides the nutty aroma while 2,3-octanedione caused the roasty smell[49−51]. Maize is an important food crop all over the world, and has different processing methods including fresh, freezing, boiling, pre-cooling, and others, each of them could produce a specific flavor. Using gas chromatography ion mobility spectrometry (GC-IMS) and gas chromatography-mass spectrometry (GC-MS), the key aroma compounds in maize with different processing technology identified include nonanal, n-hexanal, decanal, trans-2-octenal, trans-2-nonanal, 2-pentyl furan, 1-octene-3-ol, etc. Among them, nonanal, 2-pentylfuran, and 1-octene-3-ol is the main flavor compounds present in maize[52]. In addition, 2-acetyl-1-pyrroline and 2-acetyl-2-thiazoline were identified as the key aroma compounds of some frozen and fresh maize[53]. Fragrance plays a crucial role in enhancing the quality of crops, fruits, and vegetables, as the crop varieties are more easily appealing to consumers and command higher prices. A typical example is fragrant rice, known for its 'pop-corn like' smell caused by 2-AP, is favored by consumers. Previous studies have showed that the fragrance is controlled by a single recessive gene BADH2, encoding betaine aldehyde dehydrogenase, a key enzyme that can inhibits the biosynthesis of 2-AP[54,55]. The popcorn fragrance in various plants, such as rice, sorghum, millet, soybean, cucumber, mung bean and Pandan, is controlled by the recessive gene BADH2 responsible for inhibiting the biosynthesis of 2-AP[54,56−62]. The leaves of sorghum with BADH2 mutation also accumulates 2-AP, and can be used as forage grass, which is more attractive to ruminants and increase their feed take. The fragrant forage grass would improve the flavor of meat for humans[63].
Improving food aroma will further increase our demand and bring great economic benefits. In recent years, plant essential oils are widely used in the cosmetics industry, including toothpaste, soap, perfume, and all kinds of hair and skin care products. According to previous data, the aroma chemicals market is expected to exceed USD 9 billion dollars by 2032 (
www.gminsights.com/pressrelease/aroma-chemicals-market ), showcasing significant economic potential. Plant essential oils have been used for thousands of years. With so many aroma plants providing us with essential oils of different fragrances for multiple uses, the fragrance market offers around 300 essential oils, each of them is a kind of volatile secondary metabolite obtained by extracting aroma molecules from different plant tissues. The fragrant function of essential oils not only provide a sense of comfort for people but also improves product quality due to their unique biological activity. Nowadays, due to the increasing emphasis on health and wellness, 'back to nature' has become mainstream, and plant extracts have promising prospects in cosmetics. With essential oils offering various skin benefits, such as antiseptic and anti-inflammatory properties, promoting soft and smooth skin. Aromatherapy is gaining popularity worldwide, with essential oil having a positive impact on mood and body, such as peppermint for gut health and lavender for inducing relaxation[64]. It has become one of the best natural, organic, and anti-stress methods.In addition, aroma plants can be used as food flavoring agents, contributing to the creation of unique and enticing flavors in various culinary delights. Food additives derived from aroma plants, such as cloves, nutmeg, sage, are known for their aroma and flavorful properties. When added to food, these natural additives enhance the taste and overall sensory experience, making dishes more appealing to the palate.
-
As sessile organisms, plants are facing various challenges, including abiotic and biotic stress such as drought, heat, cold, pathogen infections, and herbivore attacks. Through long-term evolution, plants have evolved a highly sophisticated defense system to effectively cope with adverse environment and complete their life cycles. Previous studies have shown that plants emit volatile compounds as part of their defense mechanism when under stress[65,66].
Methylsalicylate (MeSA) is a volatile aroma compound that can be converted into salicylic acid (SA) by the SABP2 methyl esterase enzyme. MeSA is a signal molecule and plays an important role in systemic acquired resistance (SAR), plant-plant communication, and responses to various stress in plants. Experiments have demonstrated that MeSA can elicit plant tolerance and resistance against chilling injury when used as a seed soaking treatment in cucumber[67]. It has also been identified that MeSA is a pivotal mobile signal molecule for plant SAR, enabling it to travel through the phloem from infected to uninfected tissues[68]. Furthermore, MeSA is proposed as one of the herbivore-induced plant volatiles (HIPVs) emitted by various species, including apple, tomato and cucumber[69]. Interestingly, plants also emit MeSA to induce indirect defense by attracting the enemies of herbivores. For example, when a host plant is infested by soybean aphids, it released MeSA to attract Coccinella septempunctata, enhancing the biological control of these aphids[70].
Additionally, certain aroma compounds released by plants can attract their enemies. Bird damage is one of the main causes of agricultural production loss in recent years, affecting crops like wheat, millet, rice, sorghum, and barley[71]. It has been reported that fatty acid-derived volatiles can influence sparrows' feeding behaviors. Through GC-MS analysis, nine volatile organic compounds (VOCs) were detected at high level in bird-preferred accessions, and two VOCs, hexanal and 1-octen-3-ol, were used in bird feeding experiments. The results showed that sparrows prefer to eat seeds soaked in high hexanal or 1-octen-3-ol concentrations[72]. It is known that SbLOX3 was significantly upregulated in the bird-preferred accessions. Therefore, regulating specific fragrant volatiles derived from fatty acids can be an effective way against birds and mitigate their impact on crops. Understanding the role of these volatile compounds in plant defense mechanisms opens new avenues for developing environmentally friendly strategies to protect agricultural production from bird damage.
-
Interaction between plants and pollinators is crucial for natural ecosystems and human life. Approximately 75% of major food crops and 87.5% of flowering plants rely on pollinators, with insects being the primary carriers, to increase crop yields[73,74]. During the pollination process, volatiles are important chemical signals that attract pollinating insects. For most plants, flower volatiles enable plants to communicate with pollinators, while a few plants attract pollinators through leaf volatiles. The volatiles released by plants are specific and attract the corresponding pollinators. The composition and concentrations of volatile mixtures can influence pollinators' olfactory preference for plants and may even contribute to reproductive isolation between closely related plant species.
For example, Platanthera species have similar color and shape, only Platanthera obtusata orchid can attract Aedes aegypti as its effective pollinators, due to the specific volatiles emitted by Platanthera obtusata orchid, which Aedes aegypti can sense[75]. Similarly, Ficus species exclusively rely on their specific pollinating wasps for pollination. This mutualistic symbiosis is vital as the larvae of pollinating wasps can only develop in the syconia of the host Ficus species. The crucial factor for host identification by these pollinating wasps is their recognition of specific volatile substances released by the host figs. Using Ficus pumila var. pumila and its pollinating wasp Wiebesia pumilae as research materials, experiments confirmed that decanal was the attractive chemical compound in their relationship[76]. Bumble bees utilize pollen odor to assess pollen availability in flowers of Rosa rugosa. Adding components like tetradecyl acetate and eugenol to flowers significantly increased the landing frequency and pollen-collecting behavior, while the addition of 2-tridecanone and geranyl acetate reduced landing responses[77]. As a high-quality forage, the demand for alfalfa is increasing significantly. When introduced bees into alfalfa fields, the seed yield of alfalfa increased greatly[78]. Thus, studying the regulation of flower volatiles-insects interaction can provide a theoretical basis for improving seed yield.
Effective pollination networks can only form when flowering plants and insects appear simultaneously. In recent years, with the change of climate, there are rapid evolution and phenotypic changes in many plants, resulting in mismatches between flowering plants and their pollinators. This phenomenon has led to a loss of biodiversity. For example, due to global warming, the earliest flowering time of Erythronium grandiflorum is one month earlier than in previous years. Unfortunately, the lack of related insects leads to low pollination rate and reduced seed production[79]. Therefore, studying flower volatiles plays an important role in understanding and addressing these issues in pollination and production.
-
In the past few decades, with the development of molecular biology, genetic engineering has been used in plants to improve flavor. Compared with traditional breeding, the new technology is time-saving and convenient. In 2007, scientists from Israel have developed a genetically modified tomato with a more aroma smell. They achieved this by expressing the Ocimum basilicum geraniol synthase gene in tomato, leading to the accumulation of geraniol, which emits an intense rose scent. The volunteers responded well to tests using the genetically modified tomatoes[80]. Another important gene in the flavor enhancement of tomato is SlCCD1A, referred to as carotenoid cleavage dioxygenase (CCD), involved in the biosynthesis of carotenoid-derived volatiles. Experiments showed that overexpressing of SlCCD1A could improve the flavor quality of tomato. Through cleaving lycopene, α-carotene, and β-carotene, SlCCD1A could significantly increase the carotenoid-derived volatiles such as 6-methyl-5-hepten-2-one, geranylacetone, α-ionone, and β-ionone, thus enhanced the floral, fruity, and sweet-like aromas[81] (Table 1).
Table 1. Functional genes in the synthesis of aroma compounds mentioned.
Genes Aroma compounds Species References BADH2 2-AP Rice, sorghum, maize, millet, soybean, cucumber, mung bean, pandan [54,56−62] SlCCD1A Carotenoid-derived volatiles (6-methyl-5-hepten-2-one, geranylacetone, α-ionone, β-ionone) Tomato [81] TomLoxC Fatty acids-derived C5 and C6 volatiles and apocarotenoid
(1-penten-3-ol, 1-penten-3-one, geranial, etc.)Tomato [97,98] LiTPS2 Monoterpenoids (linalool, (E)-β-ocimene, trans-nerolidol) Lilium [87] CsTPS1 Valencene Sweet orange [89] RhNUDX1 Geraniol Rose [94] Indeed, in most cases of plant genetic engineering, foreign genes are introduced into plants and stably integrated into the plant genome. However, concerns about environmental and health impacts have led to restrictions on the cultivation and consumption of fresh genetically modified foods in many countries and regions.
Recently, gene editing technology has triggered a revolution. In this way, genomes can be precisely modified without introducing foreign DNA fragments. In fact, genome editing is not considered as a transgenic method, without the restrictions associated with genetically modified organisms in some countries. To date, CRISPR/Cas9 genome editing technology is widely used and has become a tool of choice for plant researchers. Plant bioengineers used the technology for innovative applications in biology, which have successfully created a lot of new varieties, including the creation of fragrant rice, maize, and sorghum. By designing specific target sites of BADH2 gene, there have created fragrant lines with high 2-AP concentration, which provides the basis for the breeding of aroma varieties and accelerates the improvement of flavor[63,82,83]. As the world's main cereal crops, the breeding of aroma varieties has greatly increased the economic value, especially in the field of food and livestock industry.
With the rapid development of advanced technology, the CRISPR/Cas9 system has been constantly improved. Through targeting eight genes or 12 genes using a construct, scientists obtained transgene-free octuple and duodecuple mutant plants in Nicotiana benthamiana and Arabidopsis thaliana within a single generation, which will contribute to the generation of complex genotypes[84]. Gene editing technology can not only knock out gene expression, but can also enhance gene expression. Nowadays with the improvement of technology, CRISPR/Cas9 systems can be used to increase the transcription level, thus upregulate gene expression, namely RNA-guided CRISPR activation (CRISPRa) systems. Researchers have developed CRISPR–Act3.0 which can enhance the function of multiple genes simultaneously through testing the effects of the system in rice, tomato, and Arabidopsis. Using this system, the expression of seven genes can be robustly increased in rice[85]. More interestingly, gene editing and gene activation can be achieved simultaneously in plants through the CRISPR-Combo system with a single Cas9 protein[86]. This system has been demonstrated in Arabidopsis, rice and poplar, presenting promising prospects for the development of plant breeding strategies. Using these techniques, we can obtain varieties with desired traits. For example, we can combine the fragrance characteristic with bird resistance trait in sorghum, through knock out SbBADH2 gene and relevant LOX or HPL genes in the Lipoxygenase pathway.
For fruit and flower aroma, we can also use CRISPR/Cas9 technology to regulate gene expression. The transcription factors (TF) have been reported to regulate aroma metabolite pathways and contribute to flavor of different species. LiTPS2 plays an important role in monoterpenoids production in Lilium, studies showed that LiMYB1, LiMYB305, and LiMYB330 directly bind to the promoter region of LiTPS2 and activate gene expression, thus improve the volatile terpenes and enhance the fragrance of flowers[87]. The fatty acid-derived volatiles are the important aroma components in pears, a bZIP TF PubZIP914 can regulate this pathway. Data analysis revealed the high accumulation of ethyl acetate, butanoic acid, ethyl ester, among others in the PubZIP914 overexpression plants[88]. CitAP2.10 can activate the expression of CsTPS1, which involved in valencene synthesis in sweet orange[89]. PbbHLH4 induces monoterpenoid production of orchid and led to floral scent[90]. Transcription complexes also exhibit important roles in the regulation of aroma compounds. For instance, PpERF5 and PpERF7 acts together as a protein complex to enhance the transcription of PpLOX4[91]. PhERF6 interacts with EOBI to negatively regulate floral volatile benzenoid/phenylpropanoid compounds biosynthesis in petunia flowers[92]. An ERF-MYB complex upregulates gene expression of quinone oxidoreductase (FaQR) and promotes the increased content of 4-hydroxy-2,5-dimethyl-3(2H)-furanone, which is a key volatile contributing to the aroma of strawberry[93]. These genes can all be used to increase the flavor of fruits and flowers. For the scent of roses, research shows that the Nudix hydrolase RhNUDX1, which has geranyl diphosphate diphosphohydrolase activity and supports geraniol biosynthesis, contributes to fragrance in roses[94]. Therefore, upregulating the expression of RhNUDX1 can improve rose flavor. Tomato is an important fruit and vegetable crop, which is favored by consumers because of its rich nutrition and unique flavor. Tomato had a global output of 177 million tons in 2016, with a total output value of USD 65.4 billion dollars (
www.fao.org/faostat ). However, compared with traditional varieties, the flavor of modern tomato varieties has decreased significantly. In recent years, breeders prefer to select high-yielding, disease-resistant and storage-resistant types, thus causing the flavor quality to largely decrease. With the development of the economy and the improvement of people's living standards, consumers pay more and more attention to the flavor of tomato, which is related to the type and content of volatile aroma compounds[95]. Also 33 major flavor substances were identified that could affect consumer preferences[96]. Furthermore, researchers found some loci associated with volatile compounds through GWAS. TomloxC is a key lipoxygenase involved in the generation of lipid-derived C5 and C6 volatiles, and involved in the production of apocarotenoid, thus influences fruit flavor[97,98]. Through CRISPR/Cas9 technology, we can change the expression of genes related to fragrance and create tomato with better taste.These methods are applicable to a wide variety of plants. Although there are increasing studies about fragrance in the past few years, the synthetic and metabolic pathways of many compounds are still unclear. To make better use of these aroma compounds, further research on aroma compounds should be carried out. These theoretical studies can provide a basis for cultivating high quality varieties. Therefore, we need to explore the gene regulatory network in detail, and experiments should be conducted to determine which genes jointly determine certain traits. These basic studies will lead to the creation of new varieties. Another challenge that needs to be solved is the establishment of a stable genetic transformation system in some species. Nowadays, there are still many plants without stable genetic transformation system. For instance, efficient transformation methods for sorghum are not available, only specific cultivars are amenable to transformation, thus limiting the creation and application of new varieties. With the maturity and improvement of genetic technology, it is feasible to create more palatable and nutritious products. γ-aminobutyric acid (GABA) is the predominant inhibitory neurotransmitter in the brain and helps lower blood pressure. In 2021, a company from Japan created GABA-enriched tomatoes using CRISPR/Cas9 technology, which became the world's first CRISPR-modified food to enter market[99]. This is an important milestone for CRISPR-edited foods. Encouraged by the market and demand, a lot of genome-edited crops, fruits and vegetables will be created, enhancing the flavor to suit more consumers, and improving product value. Because the CRISPR/Cas9 system is capable of multiple targeting and multiple regulating different genes, it will be a versatile tool for plant aroma improvement.
In addition to using aroma compound extracted from plants, scientists produce lot of metabolites through synthetic biology technology. This approach is mainly achieved by expressing critical enzymes in microorganisms, mostly including yeast and bacterium. Compared with the traditional plant extraction methods or chemical synthesis, metabolic engineering in microorganisms have many advantages, such as its directed synthesis, high efficiency, land conservation, easy operation and environmentally friendly[100,101]. Vanillin is one of the most popular aroma compounds in the world and is widely used in food, beverage cosmetics and pharmaceuticals. Natural vanillin is extracted from vanilla pods, which are time consuming, laborious, and costly. Most of the vanillin is chemically synthesized from petrochemicals or lignin, which also cause environmental pollution[102,103]. In recent years, with the development of synthetic biology, vanillin produced on an industrial scale has been successfully developed and modified in microbial hosts using substrates like ferulic acid, eugenol, and glucose, obtaining yields exceeding 10 g/L[102−104]. Raspberry ketone is the main aroma component of strawberries and raspberries. Expressing four heterologous genes encoding phenylalanine/tyrosine ammonia lyase, cinnamate-4-hydroxlase, coumarate-CoA ligase and benzalacetone synthase in yeast, L-phenylalanine and L-tyrosine can convert to raspberry ketone at high levels[104]. In addition, the production of three major rose oil monoterpenoids, including citronellol, geraniol and nerol, has been achieved with single yeast strains, which provides the rose aroma while using glucose as the substrate. The ratio of three citronellol (62.30%), geraniol (27.39%), and nerol (10.31%) via synthetic pathways was close to that of Rosa × damascena MILLER[105]. Further studies revealed that different enzymes and exogenous addition of p-coumaric acid produced high concentrations of raspberry ketone. To date, many important aroma compounds have been successfully synthesized in microorganisms, such as 2-phenylethanol, nootkatone and cinnamic alcohol[106−109]. Through various enzyme combinations and exploring more appropriate pathways, yields can reach higher levels to meet market preferences.
In summary, aroma compounds play a crucial role in various aspects. We have summarized four major biosynthetic pathways for these compounds and discussed the advances in genetic engineering to enhance plant aroma and flavor. Exciting prospects lie ahead as we anticipate a wide range of improved and delightful products. Continued research and innovation will undoubtedly unlock the full potential of aroma compounds, enriching our culinary experiences and benefiting the agricultural and food industries.
-
The authors confirm contribution to the paper as follows: draft manuscript preparation: Zhang D; conceptualization & manuscript editing: Yu F, Xie Q, Ma X. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
This research was funded by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA24010306), the National Natural Science Foundation of China (U1906204, 32222010), and the Agricultural Breeding Program in NingXia Province (2019NYYZ04).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang D, Ma X, Xie Q, Yu F. 2024. Understanding and engineering of aroma compounds in crops. Seed Biology 3: e001 doi: 10.48130/seedbio-0023-0025 

Understanding and engineering of aroma compounds in crops
- Received: 16 September 2023
- Revised: 30 November 2023
- Accepted: 13 December 2023
- Published online: 15 January 2024
Abstract: Plants produce and emit a wide array of aroma compounds. These aroma compounds are extensively utilized in cosmetics, healthcare, and the food industry. In the last few years, the research on aroma compounds has made great progress. For some valuable economic crops including gain crops, fruits, vegetables and flowers, the main fragrance compounds have been identified. In this review, we summarize the important role and great potential of aroma compounds for crops and humans. Aroma compounds mainly originate from four major biosynthetic pathways in plants, including fatty acids, amino acids, terpenoids and carotenoid pathways, producing all kinds of substances including esters, alcohols, aldehydes, ketones, terpene, and sulfur compounds, etc. Importantly, we discussed the development of genetic engineering and its application potential in enhancing plant fragrance, especially the CRISPR/Cas9 system. We hope this review will provide insights into the fragrance improvement of the economic crops.
-
Key words:
- Aroma compounds /
- Biosynthetic pathways /
- Genetic engineering