








-
In recent years, there has been a significant increase in research focusing on the taxonomy and phylogeny of saprobic fungi worldwide. These include an extensive examination of the diversity of lignicolous freshwater fungi in China[1−8]. The Tibetan Plateau, as the largest and most unique geographical region on Earth encompasses a remarkable range of endemic diversity[9−11]. Recent advancements in the study of lignicolous freshwater fungi on the Tibetan Plateau have led to the discovery of an increasing number of species, underscoring its critical importance for global biodiversity conservation and scientific research[12−14].
Savoryella was introduced by Jones & Eaton[15] with S. lignicola as the type species. The sexual morphs of Savoryella are characterized by immersed, partly immersed, or superficial, globose, subglobose, or ellipsoidal ascostromata, typically 8-spored, occasionally 2-spored, cylindrical or clavate, unitunicate asci, and ellipsoidal, three-septate ascospores[16−18]. By contrast, the asexual morphs are characterized by glistening, punctiform colonies; micronematous, mononematous conidiophores; holoblastic, determinate, integrated, terminal, and intercalary conidiogenous cells, and solitary or aggregated, pyriform to obovoid, septate conidia[19]. Zhang et al.[19] synonymized Trichocladium nypae with Savoryella nypae and introduced an asexual species, S. sarushimana, into the genus Savoryella, based on morphological and phylogenetic analyses. Subsequently, Tian et al.[20] reported two additional asexual species, S. cocois, and S. chiangraiensis, collected from decaying leaves of the Arecaceae, based on phylogenetic analysis and morphological characters. Savoryella, recognized as a holomorphic genus predominantly inhabits submerged, decaying woody debris within both aquatic and terrestrial ecosystems. It has been systematically described and illustrated by mycologists worldwide[15,18,21−25].
During the investigation of the diversity of lignicolous freshwater fungi on the Tibetan Plateau, two collections were made from freshwater habitats of taxa in their hyphomycetous forms. Multigene phylogenetic analysis showed that these two isolates belong to Savoryella. In this study, one new species, Savoryella claviformis, is introduced with morphological description and phylogenetic placement. These discoveries further add to the diversity of freshwater fungi on the Tibetan Plateau.
-
Submerged decaying wood samples were collected from freshwater habitats in the Tibetan Plateau, China. Samples were obtained from freshwater lakes and rivers, encompassing various substrates such as parts of tree trunks, branches, twigs, and litter. The specimens were studied following the methods of Senanayake et al.[26]. Microscopic structures were examined by using a stereomicroscope (SteREO Discovery.V12, Carl Zeiss Microscopy GmBH, Germany), photographed by using a Nikon ECLIPSE 80i compound microscope fitted with a NikonDS-Ri2 digital camera, macro morphological characters were examined by using a dissection microscope (Nikon SMZ745T, Nikon Instruments Inc., Japan), photographed by using a Canon 6D Mark II camera, measured by using the Tarosoft (R) Image Frame Work program. The illustrated figures were processed by using Adobe Photoshop CS6 v. 10.0 software (Adobe Systems, San Jose, CA, USA).
Single spores were isolated on potato dextrose agar (PDA) plates using the techniques outlined in Senanayake et al.[26]. Both the holotype and pure cultures were deposited at the Herbarium of Cryptogams, Kunming Institute of Botany, Chinese Academy of Sciences (KUN-HKAS), and the Kunming Institute of Botany Culture Collection (KUNCC), Kunming, China. Taxonomic novelties were submitted to the Faces of Fungi database[27] and Index Fungorum 2024[28].
DNA extraction, polymerase chain reaction (PCR) amplification, and sequencing
-
Fresh mycelia were scraped from colonies grown on PDA plates and transferred to a 1.5 mL microcentrifuge tube using a sterilized lancet for genomic DNA extraction. Fungal genomic DNA was extracted using the TOLOBIO Plant Genomic DNA Extraction Kit (Shanghai Co. Ltd, China), following the protocols in the manufacturer's instructions.
PCR amplifications were undertaken using the following primer pairs: ITS5/ITS4 for the internal transcribed spacer ribosomal DNA (rDNA) region, encompassing the 5.8S rDNA coding region (ITS); LR0R/LR5 for the 28S rDNA of the nuclear ribosomal large subunit (LSU); NS1/NS4 for the 18S rDNA of the nuclear ribosomal small subunit (SSU)[29,30]. DNA preparation was conducted in a 25 μL mixture, which included 21 μL of 1× Power Taq PCR Master Mix, 1 μL of each primer from a 10 μL stock, and 2 μL of genomic DNA, and amplification was performed in the BioTeke GT9612 thermocycler (Beijing, China). The PCR conditions for ITS, LSU, and SSU involved an initial denaturation at 98 °C for 3 min, followed by 35 cycles of 98 °C for 20 s for denaturation, 53 °C for 10 s for annealing, and 72 °C for 20 s for extension, and then the final extension at 72 °C for 5 min.
The PCR products were examined using 1% agarose gel electrophoresis with ethidium bromide staining. The presence of distinct bands was confirmed using the Compact Desktop UV Transilluminator Analyzer GL-3120 gel documentation system. The PCR products were sequenced by Tsingke Company (Beijing, China).
Phylogenetic analyses
-
Newly sequences were blasted to search for closely related taxa in GenBank database (
www.ncbi.nlm.nih.gov/blast ). Sequences generated from the ITS, LSU, and SSU gene regions were verified before further analyses, using BioEdit v. 7.0.9[31]. Sequences with high similarity percentages were determined to find the closest matches with taxa and from recently published data in Table 1[19−21]. Multiple sequence alignments were aligned with MAFFT v. 7 (http://mafft.cbrc.jp/alignment/server/index.html )[32] and automatically trimmed by using TrimA1 (http://phylemon2.bioinfo.cipf.es/index.html )[33]. A combined sequence dataset was performed with the SquenceMatrix v. 1.7.8[34].Table 1. Taxa used in the phylogenetic analyses and their corresponding GenBank accession numbers.
Taxa Vouchers/strains/isolates GenBank accession numbers ITS LSU SSU Aquabispora sp. MFLU 18-1002 MK421951 MK421953 MK421952 Aquabispora setosa GZCC 20-0492 OP377819 OP377918 OP378003 Ascotaiwania latericolla ICMP 22739 MN699390 MN699407 – Ascotaiwania lignicola NIL 00005 HQ446341 HQ446364 HQ446284 Ascotaiwania lignicola NIL 00006 HQ446342 HQ446365 HQ446285 Bactrodesmium abruptum CBS 145967 MN699393 MN699410 MN699367 Bactrodesmium diversum CBS 142448 MN699352 MN699412 MN699369 Bactrodesmium diversum CBS 144080 MN699355 MN699415 MN699371 Bactrodesmium leptopus CBS 144542 MN699388 MN699423 MN699374 Bactrodesmium obovatum CBS 144407 MN699397 MN699426 MN699377 Bactrodesmium pallidum CBS 142449 MN699363 MN699428 MN699379 Bactrodesmium pallidum CBS 145349 MN699364 MN699429 MN699380 Canalisporium jinghongense SS 03491 GQ390287 GQ390272 GQ390257 Canalisporium kenyense MFLU17-1086 MH701998 MH701999 – Canalisporium krabiense MFLU 16-1888 MH275051 MH260283 – Canalisporium pallidum SS 00498 GQ390295 GQ390280 GQ390265 Canalisporium paulopallidum NCYU-106A2-3-1 MT946658 – – Canalisporium paulopallidum NCYU-106A2-3-2 MT946659 – – Canalisporium pulchrum SS 03773 GQ390293 GQ390278 GQ390263 Canalisporium sichuanense CGMCC 3.23926 OQ428270 OQ428262 OQ428254 Canalisporium sichuanense UESTCC 22.0060 OQ428271 OQ428263 OQ428255 Canalisporium taiwanense NCYU-108ZQ-D1-1-1 MT946663 – – Canalisporium taiwanense NCYU-108ZQ-D1-1-2 MT946664 – – Canalisporium thailandense MFLU 16-1900 MH275052 MH260284 – Dematiosporium aquaticum CBS 144793 MN699402 MN699433 MN699385 Dematiosporium bambusicola CGMCC 3.23774 OQ428268 OQ428260 OQ428252 Dematiosporium bambusicola UESTCC 22.0059 OQ428273 OQ428265 OQ428256 Neoascotaiwania fusiformis MFLUCC 15-0621 MG388215 KX550893 – Neoascotaiwania limnetica CBS 126576 KY853452 KY853513 KT278689 Neoascotaiwania terrestris CBS 142291 KY853454 KY853515 KY853547 Pleurotheciella aquatica MFLUCC 17-0464 MF399236 MF399253 MF399220 Pleurotheciella erumpens CBS 142447 MN699406 MN699435 MN699387 Savoryella appendiculata NF 00206 HQ446350 – HQ446293 Savoryella aquatica SS 03801 HQ446349 HQ446372 HQ446292 Savoryella bambusicola CGMCC 3.23775 OQ428269 OQ428261 OQ428253 Savoryella bambusicola UESTCC 22.0057 OQ428267 OQ428259 OQ428251 Savoryella claviformis KUNCC 10408 OP626331 PP577958 PP577960 Savoryella claviformis KUNCC 10495 PP580830 PP577959 PP577961 Savoryella fusiformis SS 00783 HQ446351 – HQ446294 Savoryella lignicola NF 00204 HQ446357 HQ446378 HQ446300 Savoryella longispora SAT 00320 HQ446358 HQ446379 HQ446301 Savoryella nypae MFLUCC 18-1570 MK543219 MK543210 MK543237 Savoryella paucispora SAT 00867 HQ446361 HQ446382 HQ446304 Savoryella sarushimana NBRC 105262 – MK411004 MK411005 Savoryella verrucosa SS 03331 HQ446355 HQ446376 HQ446298 Savoryella yunnanensis MFLUCC 18-1395 – MK411422 MK411423 Savoryella sp. NF 00205 HQ446362 – HQ446305 Savoryella cocois MFLU 23-0227 OR581911 OR438867 OR458366 Savoryella cocois GZAAS 23-0589 OR581912 OR438868 OR458367 Savoryella chiangraiensis GZAAS 23-0590 OR581914 OR438870 OR458369 Savoryella chiangraiensis MFLU 23-0228 OR581913 OR438869 OR458368 The newly generated sequences are indicated in blue. The ex-type strains are in bold and '–' indicates unavailable sequences. A maximum likelihood (ML) analysis was conducted using RAxML-HPC2 v. 8.2.12[35] on the CIPRES Science Gateway web server[36] (
www.phylo.org/portal2 ), employing 1,000 rapid bootstrap replicates and the GTRGAMMA + I model.The model of evolution for the Bayesian inference (BI) analysis was performed by using MrModeltest v. 2.3[37,38]. GTR + I + G was selected as the best-fitting model for the ITS, LSU, and SSU dataset. For the nucleotide substitution model BI analysis was conducted by Markov chain Monte Carlo sampling (BMCMC) to assess posterior probabilities (PP) by using MrBayes v. 3.2.7[38]. Six simultaneous Markov chains were run for random trees for 1,000,000 generations and trees were sampled every 200th generation. Bootstrap support values for ML equal to or greater than 75% and Bayesian posterior probabilities (PP) equal to or greater than 0.95 were given above the nodes in the phylogenetic tree (Fig. 1). Phylograms were created using FigTree v. 1.4.0[39] and subsequently modified in Adobe Photoshop CS6 (Adobe Systems, USA). The completed alignments and phylogenetic trees were then submitted to TreeBASE, with the submission ID 31293 (
www.treebase.org ).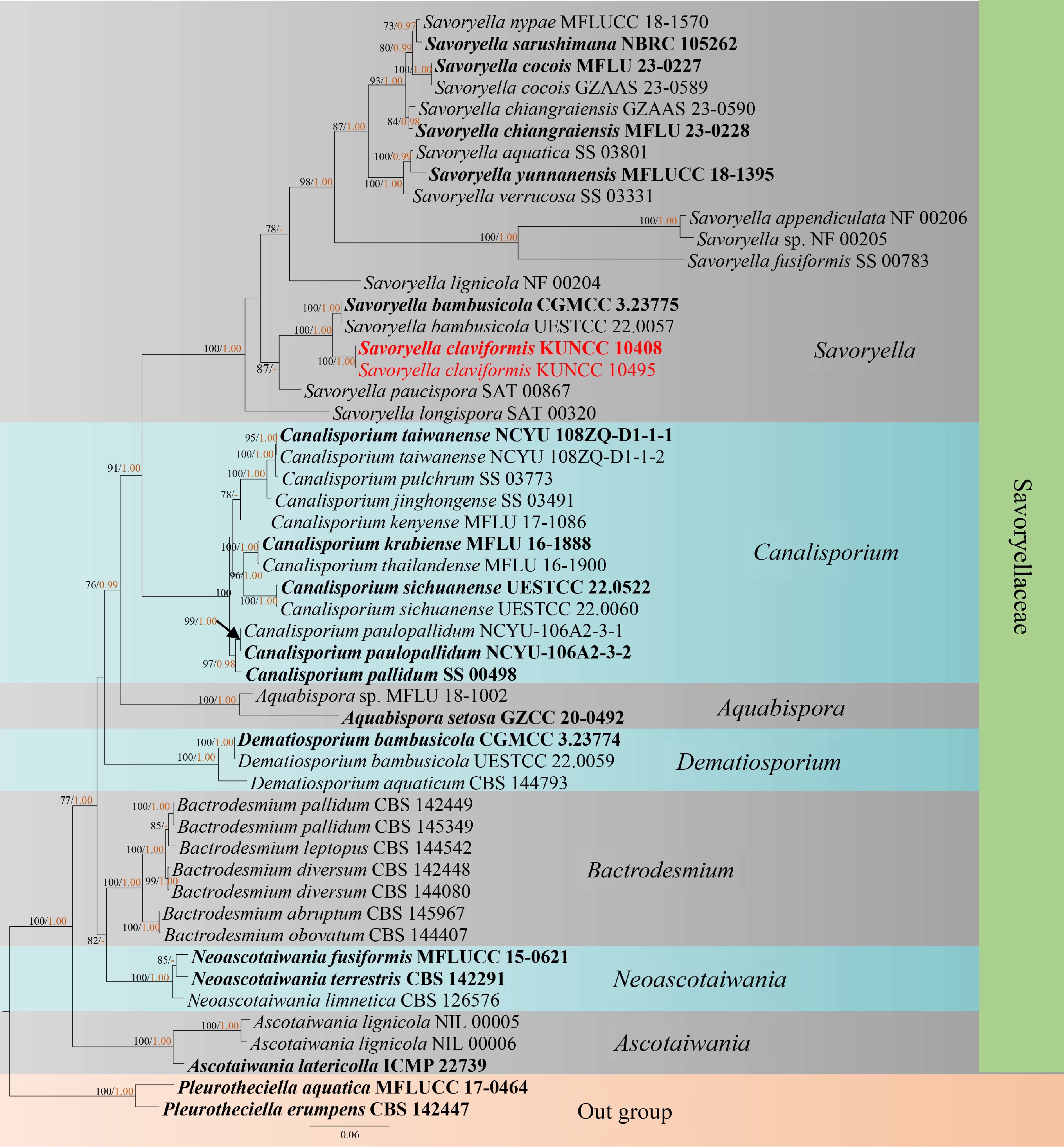
Figure 1.
RAxML tree based on analysis of a combined LSU, SSU, and ITS sequences for Savoryellaceae. Bootstrap support values for maximum likelihood (ML) equal to or greater than 75% were given above the nodes (left). Bayesian posterior probability (BIPP) equal to or greater than 0.95 were given above the nodes (right) and hyphen (−) were marked as values below 0.95. The tree was rooted to Pleurotheciella aquatica (MFLUCC 17-0464) and P. erumpens (CBS 142447)[18]. Two new isolates were shown in red, and ex-type strains are bold.
-
Best-scoring RA x ML tree for Savoryellaceae based on analysis of the combined LSU, SSU, and ITS datasets. The combined dataset comprised 51 strains with 3,268 characters including gaps (LSU: 1–1,845 bp; SSU: 1,846–2,759 bp; ITS: 2,760–3,268 bp). The tree is rooted with Pleurotheciella aquatica (MFLUCC 17-0464) and P. erumpens (CBS 142447)[18] and has a final ML likelihood value of –22,514.850213. The matrix had 1,326 distinct alignment patterns, with 36.62% undetermined characters or gaps. The estimated base frequencies were A = 0.226977, C = 0.268195, G = 0.306489, T = 0.198339; the substitution rates were AC = 1.307677, AG = 2.608024, AT = 1.966100, CG = 0.982783, CT = 6.087648, and GT = 1.000000; and the gamma distribution shape parameter α = 0.0010000000. The tree topologies of combined sequence data obtained from ML. BI analyses were not significantly different (Fig. 1).
Phylogenetic analysis showed that Savoryella claviformis (KUNCC 10408 and KUNCC 10495) formed a distinct lineage within the genus, and formed a distinct and sister group (100% ML, 1.00 BIPP) with S. bambusicola (UESTCC 22-0057 and CGMCC 3.23775) (Fig. 1).
Taxonomy
-
Savoryella claviformis R.J. Xu, S. Boonmee, K.D. Hyde & Q. Zhao, sp. nov. (Fig. 2)
MycoBank: MB853230; Facesoffungi number: FoF 15685
Etymology: The specific epithet 'claviformis' refers to the claviform conidia.
Holotype: HKAS 133191

Figure 2.
Savoryella claviformis (HKAS 133191, holotype). (a) Colony on nature substrates. (b), (c) Conidiophores and apical conidia. (d) Conidiophore. (e)–(g) Conidia. (h) Culture colony on PDA medium, from surface. (i) Culture colony on PDA medium, from reverse. Scale bars: (b)–(g) = 50 μm.
Saprobic on decaying stems of wood submerged in a freshwater stream habitat. Asexual morph: Colonies effuse, scattered, brown to dark brown. Mycelium immersed, subhyaline to pale brown, composed of branched, septate hyphae. Conidiophores 66–151 × 3–6 μm (
$ \overline{x} $ $ \overline{x} $ Culture characteristics: Conidium germinated on PDA within 48 h. Mycelia superficial, velvet, irregular, circular, raised near the center, surface villiform, dense, grey mycelium in the center, brown to grey from above, dark brown from below, becoming sparse and paler at the entire margin.
Material examined: CHINA, Xinjiang Province, Bayingoleng Mongolian Autonomous Prefecture, Kaidu River, 41°52'4.8" N, 86°43'39.8" E, 1049 msl, saprobic on submerged decaying wood in freshwater habitats, 22 July 2021, R.J. Xu, MD-325 (HKAS 133191, holotype), ex-type culture KUNCC 10408. Bayingoleng Mongolian Autonomous Prefecture, Bosten Lake, 42°3'4.69" N, 87°8'47.71" E, 1051 msl, saprobic on submerged decaying wood in freshwater habitats, 22 July 2021, R.J. Xu, MD-376 (HKAS 133192), living culture KUNCC 10495.
Notes: Savoryella claviformis can be distinguished from all asexual species in Savoryella by its semi-micronematous conidiophores, terminal, determinate conidiogenous cells, and acrogenous, solitary, fusiform, claviform, rostrate conidia[19,20]. Additionally, phylogenetic analysis showed that S. claviformis clustered into a distinct subclade and is sister to S. bambusicola, with (100% ML/1.00 BIPP) bootstrap support (Fig. 1). Since only the sexual morph of S. bambusicola has been discovered, it is not possible to compare their morphological differences[18]. Further comparisons of ITS sequences demonstrate a 7.9% (33/417 bp, excluding gaps) difference between S. claviformis and S. bambusicola. Therefore, following the guidelines of Chethana et al.[40,41], S. claviformis has been identified as a new species, supported by both morphological and phylogenetic evidence.
-
Savoryellaceae currently comprises seven genera: Aquabispora, Ascotaiwania, Bactrodesmium, Canalisporium, Dematiosporium, Neoascotaiwania, and Savoryella[7,17,18,21,42−44]. The asexual morphs of these genera are typically characterized by micronematous, often reduced to undifferentiated hyphal conidiophores and monodictys-like conidia. Specifically, Ascotaiwania, which has terminal, blastic, globose, dictyosporous conidia; Bactrodesmium known for its sporodochium-like conidiomata; Canalisporium is characterized by dark brown and muriform conidia; Dematiosporium is known for monodictys-like conidia; and Neoascotaiwania, recognized for its fusiform, dark brown, transversely septate conidia[17,21,42−45].
The specimens described in this study were collected from freshwater habitats on the Tibetan Plateau in China, increasing our understanding of fungal diversity in this region and enabling comparisons across a north-south gradient in Asia[1]. In addition, among the 17 accepted species in Savoryella, only four species, including S. cocois, S. chiangraiense, S. nypae, and S. sarushimana, are known as asexual morphs[19,20]. Savoryella limnetica was observed producing an asexual morph in culture, as reported by Réblová et al.[46]. However, based on molecular data and culture characteristics, it was synonymized with Neoascotaiwania limnetica by Hernández-Restrepo et al.[47].
This study exposes a new ascomycetous taxon from the freshwater ecosystems of the Kaidu River and Bosten Lake in Xinjiang, China, marking a new discovery in the region's fungal diversity. Originating from elevations above 4,000 m on the Tibetan Plateau, the unique geographical and climatic conditions of the Kaidu River and Bosten Lake create distinctive niches that support a rich biodiversity, including previously undocumented fungal species. This discovery enriches our understanding of ascomycetes in freshwater ecosystems.
-
The authors confirm contribution to the paper as follows: study conception and design: Xu RJ, Hyde KD, Zhao Q; data collection: Xu RJ, Guo YY, Yang QY; analysis and interpretation of results: Xu RJ, Dong W, Boonmee S; draft manuscript preparation: Xu RJ. All authors reviewed the results and approved the final version of the manuscript.
-
The authors of the manuscript confirm that data supporting our study findings are available in the article. Data regarding species/specimen DNA sequences are publically available on the accession provided in Table 1, in the GenBank data base of NCBI.
The authors appreciate the support given by the Thesis Writing Grant of Mae Fah Luang University, Thailand, to Rong-Ju Xu. This study is supported by the Second Tibetan Plateau Scientific Expedition and Research (STEP) Program (Grant No. 2019QZKK0503), the Survey of Wildlife Resources in Key Areas of Tibet (ZL202303601); Major science and technology projects and key R&D plans/programs, Yunnan Province (202202AE090001). Wei Dong thanks the National Natural Science Foundation of China (grant No. 32200015), the foundation of guangzhou bureau of science and technology (grant No. 2023A04J1425).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu RJ, Boonmee S, Dong W, Guo YY, Yang QY, et al. 2024. Savoryella claviformis (Savoryellaceae), a new freshwater hyphomycetous species from the Tibetan Plateau, China. Studies in Fungi 9: e009 doi: 10.48130/sif-0024-0009 

Savoryella claviformis (Savoryellaceae), a new freshwater hyphomycetous species from the Tibetan Plateau, China
- Received: 11 April 2024
- Revised: 29 May 2024
- Accepted: 14 June 2024
- Published online: 16 July 2024
Abstract: During an investigation of lignicolous freshwater fungi on the Tibetan Plateau, China, two collections were obtained from submerged wood in freshwater habitats. The morphological examinations and phylogenetic analysis using LSU, SSU, and ITS sequence data have identified that the two collections represent a novel species within the genus Savoryella, namely S. claviformis. Savoryella claviformis forms a distinct clade within Savoryella, and possesses unique characteristics compared with existing asexual species in having semi-micronematous, solitary, cylindrical conidiophores, terminal, determinate conidiogenous cells, and acrogenous, claviform, rostrate conidia. Detailed descriptions and illustrations of this species are provided.
-
Key words:
- 1 new taxon /
- Hyphomycetes /
- Phylogeny /
- Taxonomy /
- Sordariomycetes