








-
Rice ratooning is a cultivation technique that combines both innovative and traditional approaches. As early as 1,900 years ago, during the Xi Jin Dynasty, Guo Yigong recorded the practice of ratooning rice in Guangzhi(an ancient book). After harvesting the main crop, new plants with roots would grow from the stem base of the remaining stubble and mature by September. Similar explanations can also be found in YangshengYaoshu (An ancient book on skills necessary for health preservation) by Zhang Chen in the 4th century AD and Nongzheng Quanshu (An ancient book on the encyclopedia of Agricultural Affairs) by Xu Guangqi during the Ming Dynasty, respectively[1,2]. Therefore, rice ratooning can be defined as a farming approach where axillary buds sprout from leftover stubbles and develop into new panicles using appropriate cultivation techniques for a second ratoon rice crop following the harvest of the main crop. There has been a long history of rice ratooning in China's Yangtze River basin. However, ratoon rice yields were lower and unstable at that time primarily due to unsuitable varieties and cultivation techniques. Consequently, this technology was only applied to self-rescue after disasters[3−5]. In the 1930s, Professor Yang Kaiqu from the former Sichuan Agricultural College pioneered systematic research on ratoon rice's biological characteristics and cultivation techniques, laying the foundation for in-depth research and application of rice ratooning in China. Subsequently, southern provinces in China also initiated research efforts; however, they failed to scale up effectively, thus struggling to attract attention and support from agricultural management departments. In the late 1970s, with the development and release of dwarf rice varieties as well as advancements in hybrid rice technology, particularly high-regenerating ability hybrid rice like Shanyou 63 bred by Academician Xie Hua'an in Fujian Province, rice ratooning gained significant momentum and widespread adoption in production. However, during its early stages of development, ratoon rice was primarily limited to food-deficit provinces and regions in southern China where the population density was high, but arable land resources were insufficient for double-cropping due to sunlight and temperature constraints[6]. After the 1980s, with the continuous development of China's reform and opening-up policy, various factors such as rural labor migration and agricultural structural adjustment has driven significant changes in China's rice cropping system. Consequently, there has been a consistent decline in the cultivation area of double-cropping rice in southern provinces over the past years. This decline is primarily attributed to the conversion from double-cropping to single-cropping rice facilitated by market regulations. Additionally, efforts have been made to reduce the planting area of early rice with relatively poor quality, leading to a gradual expansion of single-cropping middle rice.
To ensure food security, research and application of rice ratooning, particularly mechanically harvested ratooning rice, have been objectively promoted. This has significantly enhanced its economic benefits and facilitated the continuous deepening of research on rice ratooning, resulting in a substantial increase in the planting area of ratoon rice over successive years[7]. Moreover, countries such as Brazil, India, Colombia, Japan, Pakistan, the Philippines, Thailand, and the United States have also initiated research and application of ratoon rice along with its cultivation technology[8,9]. As a result of continuous advancements in rice breeding methods and innovative mechanized cultivation techniques, such as water-fertilizer management, the development of double-channel and full-feeding harvesters, and the optimization of field routes for mechanical harvesting, there has been a notable increase in the proportion of ratoon rice planting within the overall cropping system. Consequently, this has led to a significant annual expansion by millions of acres in recent years[10]. According to statistics from Fujian, Jiangxi, Zhejiang, Hunan, Hubei, Anhui, Jiangsu, Sichuan, Chongqing, Yunnan, and Guizhou, 1.1 Mha were dedicated to cultivating ratoon rice in 2022, and therefore, there is an additional 4.5 Mha for the ratoon rice expansion. If ratoon rice increases grain yield by 3.75 t·ha−1 compared with the middle-season rice, 4.5 Mha of the ratoon rice could increase rice production by 16.9 million tons, which is about 8% of annual rice production in China (Fig. 1)[5,11] . This system of farming rice has emerged as one of China's most efficacious cultivation patterns[5,12−14].
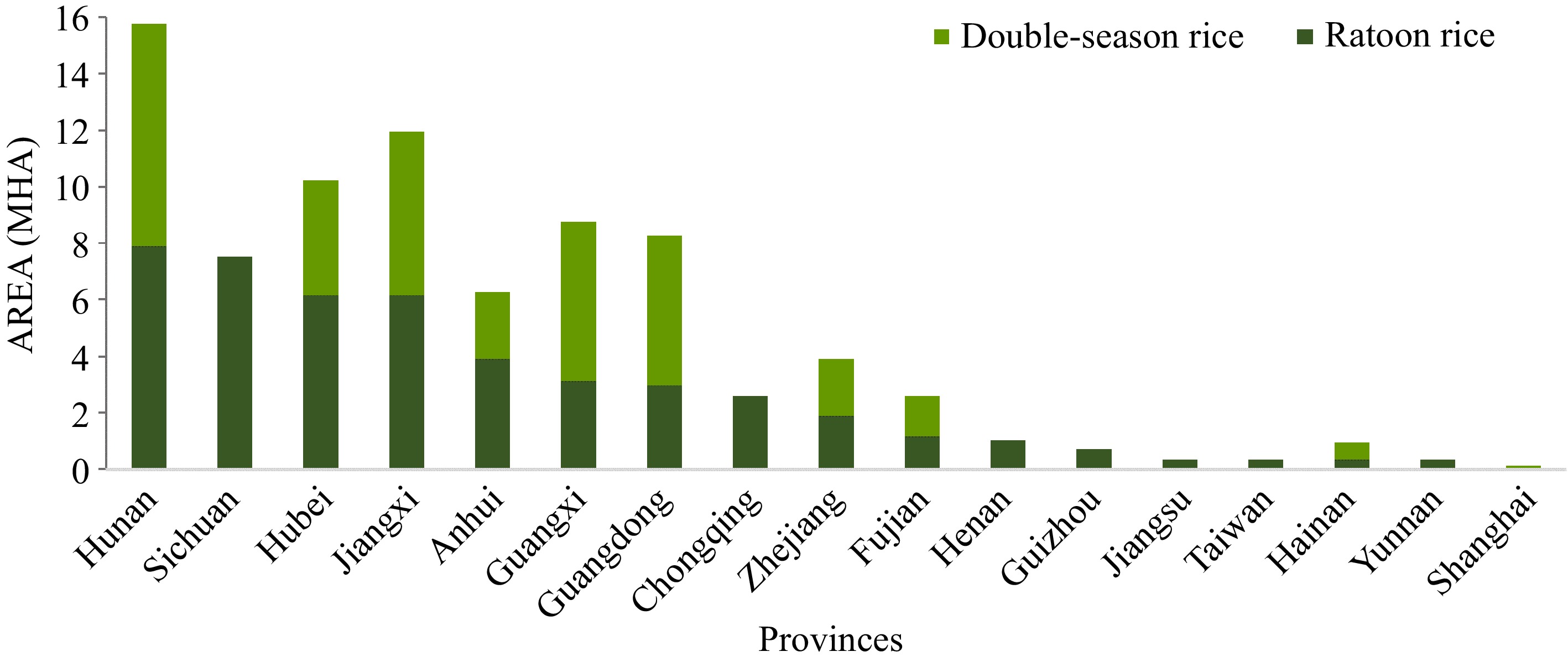
Figure 1.
Potential suitable regions and proportions for DR and RR: Area of DR and RR in each province[11].
However, numerous practical scientific challenges have also arisen, particularly regarding overcoming the substantial decline in yield caused by mechanical rolling damage to rice stubbles during main crop harvesting. This issue has become an urgent and prominent concern demanding resolution. Accordingly, during China's 14th Five-Year Ppan period and with support from the National Science and Technology special project, several provinces have engaged in collaborative research efforts and achieved significant progress[12−18]. Drawing upon years of research and practical experience from our team led by the corresponding author, this paper focuses on elucidating the current status and prospects of high-yielding formation in ratoon rice and its underlying physiological mechanism from a crop ecophysiology perspective.
-
Previous studies have revealed that the inheritance of regenerative and high-yield agronomic traits in rice presents a complex quantitative genetic challenge, where the interplay between genotype and environment influences its genetic expression. Recently, Yao et al.[19] successfully identified a promising candidate gene called RRA3 (Rice Ratooning Ability 3) through genome-wide association analysis (GWAS), which exhibits significant associations with various regeneration and yield traits specific to ratooning season rice. Phenotypic experiments conducted in potted conditions and field settings have convincingly shown that knockout variants of RRA3 substantially enhance rice regeneration capacity and ratoon crop yields when compared with the wild-type Huanghuazan (HHZ). Conversely, overexpressed variants noticeably diminish both aspects[18]. This complexity impedes genetic breeding research on ratooning rice[18,20]. The key to successful ratoon rice development lies in the meticulous screening of suitable cultivars; therefore, it is crucial to emphasize this aspect. Historically, due to limited research projects dedicated to ratoon rice breeding in China, the screening of ratooning rice cultivars was restricted to the current cultivated varieties for practical production purposes. From the 1970s to the 1980s, ratoon rice varieties in China was primarily selected from conventional and hybrid rice varieties renowned for their high yield and disease resistance. In the 1990s, there was a shift towards prioritizing comprehensive traits such as superior yield performance, stress resistance, and grain quality when selecting ratoon rice cultivars. Significant progress was made during this period in achieving a synergistic improvement of both yield and quality in ratoon rice.
Screening ratooning rice varieties with enhanced regeneration capacity from a wide range of options would present a formidable task. Moreover, it is not feasible for these same varieties to consistently achieve high yields across diverse regions due to varying ecological conditions. Hence, conducting in-depth investigations on specific agronomic traits closely related to regeneration ability, such as chlorophyll content, photosynthesis capacity, root vigor, and stem, strength is imperative. Additionally, establishing identification criteria for predicting elite lines with stronger regeneration capacity within both main crops and ratooning rice plants have become crucial. Addressing these issues will provide a solid foundation for expediting the screening process for desirable ratoon rice varieties. Accordingly, we further analyzed the relationship between the yield and its components of the main crop. The results indicated that the number of effective panicles and grain number per panicle were the primary factors influencing the yield of the main crop, contributing to 88.3% of the total yield. Moreover, we selected nine agronomic indicators from the agronomic traits of mature stage main crops that exhibited significant correlation with regenerating ability and ratooning rice yield. Based on this, we established a regeneration grading identification system with higher prediction accuracy for screening elite varieties of ratooning rice. Among these indicators, the SPAD attenuation index of the reverse-counted fourth leaf blade, root sap intensity, and stem diameter at the second internode base had the highest screening accuracy and reliability rate. The aforementioned factors thus play a pivotal role in assessing high-yield ratoon rice varieties.
During the 13th Five-Year plan period in China, supported by major national science and technology projects, Chinese rice breeders successfully developed and released high-quality ratoon rice varieties such as Jiayu 99, Luyou Mingzhan, Liangyou 688, and Nei6you 7075. Building upon this achievement, rice cultivation experts have further identified numerous varieties with axillary buds' stronger regenerative and tillering capabilities at medium and low node positions based on the environmental requirements and cultivation practices for rice ratooning. These include Yongyou 2640, Yongyou 1540, Zhenliangyou 60, Longliangyou Huazhan, Jingliangyou Huazhan, Yongyou 4949, Yuanyou 607, Weiliangyou 8216, and Yliangyou 900. These varieties have been extensively utilized in mechanized rice ratooning across various ecological regions in southeast and southwest China, including Fujian, Jiangxi, Zhejiang, Sichuan, and Chongqing provinces[4,5]. Based on the growth duration, the existing ratoon rice varieties can be classified into three types: early-maturing (shorter-duration) ratooning rice varieties, medium-maturing (medium-duration) ratooning rice, and late-maturing (longer-duration) ratooning rice varieties[2]. The diverse range of these types effectively caters to the varying ecological conditions and altitudes within different regions of China. It is crucial to fully consider their regeneration characteristics in conjunction with regional temperature and light conditions when selecting ratooning rice varieties from them. The sprouting of axillary buds from the lower nodes is particularly crucial for rice varieties dominated by this growth pattern. Despite an increase in cut stubble height from 5−35 cm, it does not affect the growth duration of ratooning seasonal rice. Ratooning rice varieties with low node axillary bud sprouting into effective panicles, such as Nanling No.1 and perennial rice PR25 and PR23, exhibit a distinct characteristic of forming effective panicles through sprouting at lower positions[21]. However, some ratooning rice varieties primarily rely on tiller regeneration characteristics for axillary bud sprouting, such as the Yongyou series indica × japonica hybrid rice varieties, including widely used excellent varieties like Yongyou 2640 and Yongyou 1540. Another type of variety exhibits a distinct apical advantage, where its axillary buds can sprout into panicles in multiple nodes distributed upwards and downwards from the stem base, including Quanyou 607. Therefore, selecting appropriate ratooning rice varieties based on regional ecological conditions is crucial for achieving high yields.
The optimization of stubble height and regulation of cultivation control
-
The precise determination of stubble height is critical in attaining high yields in rice ratooning, provided that appropriate cultivars are selected based on regional light, temperature, and rainfall conditions. Research has indicated that for main crop rice, prioritizing the culture of large panicles while ensuring an adequate number of effective panicles is essential for attaining high yield. However, to achieve a high yield in rice ratooning, the focus should be on maximizing the number of effectively regenerated panicles while maintaining a stable number of grains per panicle per unit area[18]. Therefore, the promotion of axillary bud sprouting and tillering and ensuring their successful development into productive panicles are crucial factors for consistently achieving optimal ratooning rice yields[22]. The results of further studies have demonstrated that the remaining stubble height and characteristics of axillary bud regeneration influences the number of effective panicles in ratooning season rice. The contribution of regenerated panicle yield to the overall output of ratooning season rice varies depending on the panicles produced from different nodal positions along the remaining stubble. In the cultivation mode with a high remaining stubble height (> 35 cm), significant contributions to the yield were observed from axillary bud sprouting into panicles at the 2nd and 4th node positions (D2 and D4) from the top for ratooning season rice due to apical dominance influence. Conversely, in low stubble height cultivation (< 25 cm), determining overall yield relied crucially on regenerated panicles yielded at the 3rd−5th node positions from the top for ratooning season rice[22,23] (Fig. 2). Therefore, appropriate fertilization measures should be determined based on regeneration characteristics of the ratooning season rice and cut stubble height.
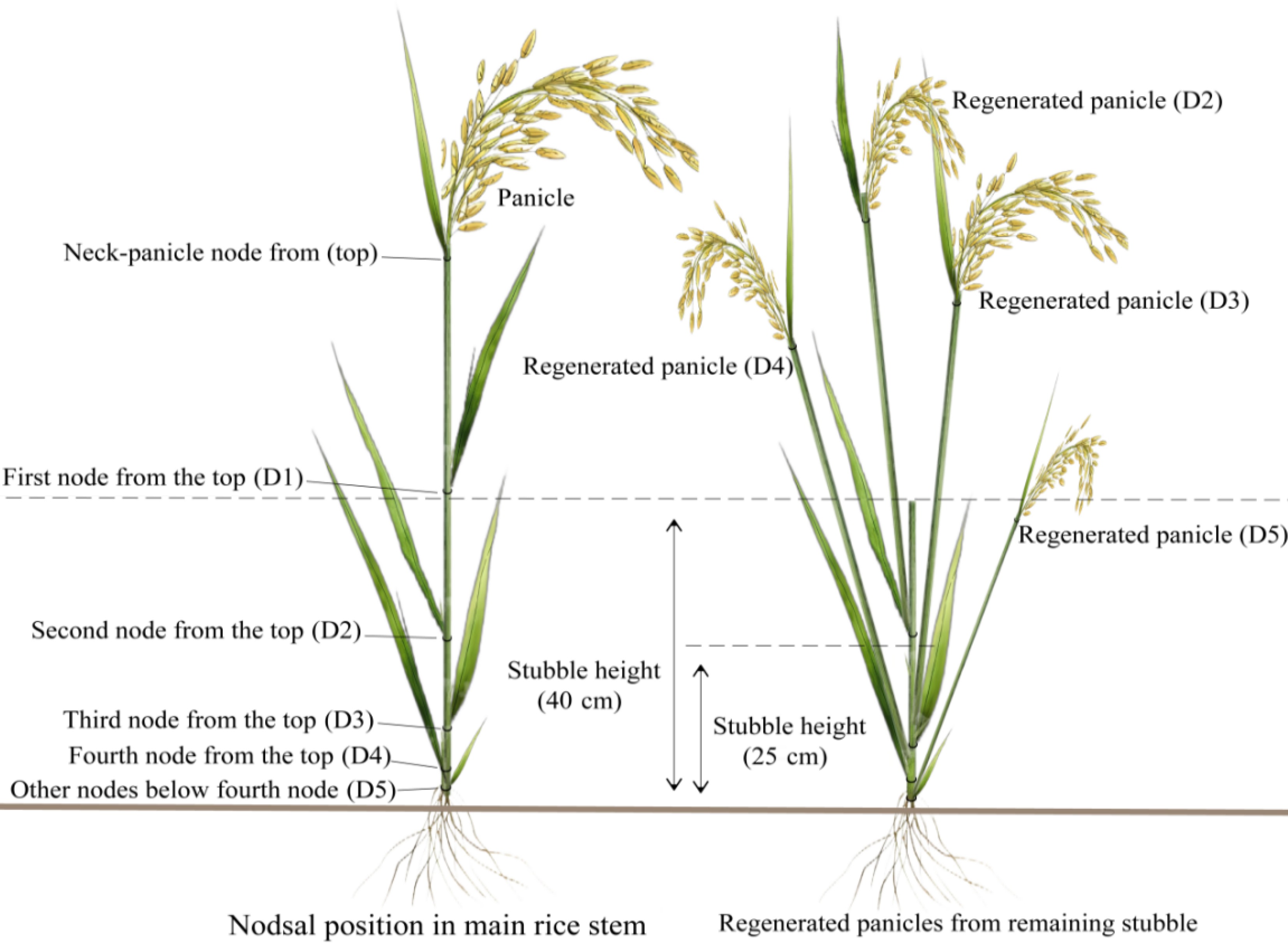
Figure 2.
Diagrams of the nodes and regenerated panicles in ratoon rice systems.
Further studies have consistently demonstrated a negative correlation between the growth duration of ratooning season rice and stubble height, with higher stubbles leading to shorter growth periods[17,22−24]. However, as previously mentioned, determining the appropriate stubble height is influenced by local light and temperature resources and the regeneration characteristics of different varieties. For instance, the growth duration of some varieties with more efficient panicles emerging from buds at lower node positions in the remaining stubbles is predetermined and fixed by their genetic traits, particularly their sensitivity to temperature and light. Changes in stubble height do not affect the growth duration of these ratooning season rice varieties. Conversely, varieties with more pronounced apex dominance that can sprout into panicles at both upper and lower node positions display a characteristic response of shortening when stubble height increases from 5−40 cm[21]. Therefore, determining the optimal stubble height should consider both variety regeneration characteristics and the availability of local light and temperature resources. The current high-stubble cultivation technology is generally suitable for rice ratooning in the lower-middle reaches of the Yangtze River basin, such as Wuhan, Anhui, and other rice-growing regions with relatively limited renewable seasonal light and temperature resources. On the other hand, low-stubble rice ratooning is more appropriate for areas in southern provinces of China that have relatively abundant light and temperature resources. However, these two ratooning techniques differ in water and fertilizer management. Specifically, it is crucial to fully consider the nutrient-dependent variation in axillary bud sprouting and tillering at different node positions of the remaining stubbles. In high-stubble cultivation, apart from selecting varieties with buds at higher or all node positions capable of sprouting into panicles (if the stubble height remains too low (< 25 cm) for such varieties, inevitably hampers apex advantage leading to reduced axillary bud sprouting into panicles which ultimately affects yield[21], fertilization strategy emphasizes protecting the sprout into panicles of high-node position axillary buds within remaining stubbles while ensuring that premature senescence does not occur in the main crop. Fertilization should be carried out before the main crop harvest to preserve root vigor and ensure bud survival.
Additionally, it is crucial to consider water and fertilizer management after the main crop harvest to facilitate rapid nutrient translocation from residual rice stubbles, stimulate sprout growth and elongation of high node position axillary buds, and prevent nutritional deficiencies that may hinder sprouting and tillering of lower node position buds. This ultimately affects the regeneration rate, subsequently impacting the number of regenerated panicles and overall yield. Effective water and fertilizer management in low remaining stubble cultivation involves not only selecting ratooning rice varieties with a propensity for sprouting into panicles at low-node positions but also prioritizing the prompt sprouting and tillering of axillary buds during the later growth stages of the main crop to enhance survival rates among low-node position axillary buds. This approach promotes orderly progression between vegetative and reproductive growth during the regeneration following the main crop harvest. In particular, special attention should be given to soil fertility. These varieties exhibit obvious advantages in low nodal position bud sprouting and tillering, allowing them to absorb nutrients from the soil through new roots. Moreover, the adoption of low-maintenance stubble cultivation extends the growth duration of rice during the ratooning season. Therefore, it is crucial to prioritize vegetative growth and dry matter accumulation from the main crop harvest until the full heading of the ratooning season rice. To achieve high yields with large panicles in ratooning season rice, it is essential to promote vegetative growth through nutrient regulation[23,25−27], followed by a timely transition into reproductive growth. Hence, meticulous attention must be given to achieving uniform and balanced fertilization and an optimal proportion of nutrient elements during the cultivation of ratooning season rice[23,28]. This is essential to prevent premature senescence and hinder the formation of abundant and filled grains caused by too early termination of irrigation and nutrient supplies.
In recent years, direct seeding of rice has been explored as a strategy to overcome the potential yield reduction in ratooning season rice caused by shortened growth duration due to the high cut height of main crop rice. This approach aims to shorten the growth duration of the main crop, enhance the flexibility of ratooning season rice's growth duration, promote sprouting of axillary buds and regenerated panicles from lower nodal positions on stubbles, and ultimately achieve larger panicle size and higher yield. Although some progress has been made in this research area[29], challenges such as ensuring stable yields still need to be addressed. Generally, poor and uneven establishment, inadequate weed control, lodging susceptibility and frequent cold spells can significantly hinder seed germination and seedling growth of direct-seeded rice under direct-seeding cultivation. Therefore, sustainable and effective technologies such as seed coating and priming treatments are required to improve seed germination and enhance seedling growth in direct-seeded rice under chilling stress[30].
Previous studies have also indicated two key fertilization periods for improving grain yield in main crop and ratooning season rice[17,18]. One aims to preserve root vigor and promote bud survival by applying fertilizer 15 d before harvesting the main crop, while the other focuses on promoting tillering 3−7 d after harvesting the main crop. The effects of the two fertilization timings on root vigor, bud survival, and tillering promotion in main crop and ratooning season rice was found to be influenced not only by the nitrogen nutrition status of the main crop at maturity but also by the height and quality of remaining stubbles after harvest[31]. However, it was observed that low remaining stubble ratooning rice mainly originated from the first and second nodes of the stem base. Applying a significant amount of budding fertilizer before harvesting the main crop accelerated sprouting and elongation of axillary buds, making them susceptible to being cut during the mature harvesting stage, particularly for ratooning rice varieties with prominent apical dominance. Furthermore, improper application of bud sprout-promoting and root-vigor preserving fertilizer during the late stages of the main crop can lead to a substantial increase in greenhouse gas emissions, especially nitrous oxide (N2O), as well as a decrease in nitrogen use efficiency, which ultimately affects rice quality[17,31−33]. Therefore, under low remaining stubble cultivation conditions, it is crucial to apply root-vigor preserving and budding fertilizer promptly and appropriately based on soil fertility[22,32]. This will ensure a high survival rate of axillary buds before the main crop harvest. The quality of rice stubbles and the health of axillary buds before and after the main crop harvest are closely related to soil moisture conditions and nutrient supply capacity at that time. Excessive soil moisture during main crop harvesting and insufficient or excessive fertility supply can hinder sprouting and regeneration of axillary buds. In some cases, rice stubbles may be damaged or buried in mud due to machine harvesting and rolling, leading to rotting and death, ultimately resulting in poor regeneration of remaining stubbles and uneven maturity between harvester rolling and non-rolling areas. This reduces the land utilization rate, leading to yield reduction[4,32]. Therefore, an in-depth exploration into water-fertilizer coupling regulation for low remaining stubble ratooning rice is essential for promoting timely sprouting and elongation of axillary buds at lower node positions while encouraging tillering into panicles. These factors are more important than the improvement of harvesters for achieving high-quality produce with ultra-high yields[31,32].
In light of this issue, we took the initiative to research and establish mechanized harvesting and cultivation technology for low-remaining stubble ratooning rice in different regions of southeast China, considering the specific characteristics of light-temperature resource endowment. This was done to support the development of high and stable yields in rice ratooning. The technical system consists of three key components[31,32]: (1) Emphasizing a moderate transfer of nitrogen fertilizer from the early stage to be applied during the later stage of the main crop. This means that while maintaining a constant total nitrogen application rate, an appropriate amount of nitrogen (accounting for 30%−40% of the total nitrogen application rate) should be allocated as young panicle differentiation and grain fertilizers. This helps maintain root vitality, prevent premature aging, and ensure axillary bud survival during the late growth stages of the main crop. Doing so can reduce excessive tillering peaks, which lead to an increase in ineffective tillers, less effective panicle formation, weakened stem strength, and ultimately decreased rice stubble quality. Additionally, it is recommended that root-vigor preserving and bud-promoting fertilizers be reduced or avoided 20 d after full heading stage completion to minimize N2O emissions and improve nitrogen utilization rates. (2) The ratooning season rice was subjected to a uniform fertilization method to facilitate the systematic progression of vegetative and reproductive growth. Given the synchronization between these two growth stages, proper nutrient regulation is crucial in promoting their orderly advancement. Therefore, applying effective nitrogen fertilizer on time after the main crop harvesting is essential to stimulate axillary bud sprouting and rapid tillering, enhance photosynthetic accumulation, and establish a solid foundation for achieving high yield. Emphasis should be placed on using complex fertilizers with high nitrogen and potassium elements during the full heading stage to promote young panicle differentiation, filling, and fruiting, as well as the development of large panicles, leading to higher yield. (3) It is advisable to conduct a secondary soil drying of the field before harvesting the main crop, followed by reducing the cut height of rice stubble (15−25 cm). This entails ensuring timely desiccation of the field soil (up to 30%−35% moisture content) after applying panicle and grain fertilizer to prevent paddy soil subsidence caused by mechanical rolling and damage to residual rice stubbles due to excessive soil moisture during machine harvest. After harvesting, it is crucial to maintain a mild alternating dry and wet condition in the field soil to promote rapid tiller regeneration. Additionally, natural drying should commence during the middle and late stages of heading and grain-filling (known as implementing the third time field soil drying in the regenerated season), facilitating the efficient transfer of dry matter and improving harvest index. This cultivation technology has been proven to result in even growth and neat maturity of ratooning season rice population subjected to low cut stubbles, significantly reduce rolling loss rate, ensure uniform rice heading, and remarkably enhanced yield and appearance quality (head rice rate). The maximum annual yield achieved on large demonstration areas for both main crop and ratooning rice has consistently exceeded 19 t·ha−1 for 8 years, with ratooning rice surpassing 9 t·ha−1. This signifies a significant milestone in global mechanized ratoon rice production[4,31,32].
Dry matter production and partitioning characteristics
-
Previous studies have indicated that carbohydrate accumulation in the main crop is primarily divided into two distinct processes. The initial process occurs before the differentiation of young rice panicles, during which carbohydrates are predominantly utilized for developing vegetative organs such as roots, stems, and leaves. The subsequent process involves the accumulation of carbohydrates in the young panicle of ratooning rice, mainly transited from the stems and sheaths as non-structural carbohydrates. This accumulation pattern demonstrates an initial increase followed by a subsequent decrease during grain filling[34]. The results of further investigations have revealed a strong correlation between carbohydrate accumulation in the stem-sheath and leaf area index (LAI)[35]. Zhou et al.[36] utilized a 14C isotope tracer method to examine the distribution patterns of the main crop photoassimilates at the full heading stage to the main crop and the ratooning season rice. The findings demonstrated that approximately 80%−90% of the main crop photosynthates during the full heading stage were allocated to the panicles of both the main and ratooning rice crops, while 10%−20% remained in flag leaves and only about 5% were distributed within the stem and sheaths. Furthermore, after harvesting the main crop, 52%−70% of residual photosynthates in rice stubbles were directed towards panicle development during regeneration season. It was observed that higher yields were achieved during regeneration season when more photosynthates remained below the second node position (counted reversely from the stem base) in the mature stage of the main crop[36]. We also observed that, in comparison with late rice synchronized in heading with the ratooning rice (late season), a significantly higher amount of 13C assimilates were transported to the panicles during the late growth stage of main crop rice. This transportation primarily occurred from the stem with sheath, and was consequently reduced for transfer to the underground part. These findings indicate that ratooning rice exhibits a stronger ability to export and transport materials from its stem and sheath compared to genotypically similar late rice that synchronously heads with the ratooning rice in late season. As a result, this leads to an increased harvest index and high daily yield of ratooning rice[37] (Table 1). Xie's studies confirm that the harvest index of high-yield main crops typically ranges from 0.50 to 0.53, while high-yield ratooning rice reaches as high as 0.67−0.70[38]. Yi et al. investigated the relationship between source and sink in regenerated rice. They observed that the leaf area index, number of grains per panicle, and yield of regenerated seasonal rice was significantly lower compared to late rice with the same genotype heading at the same time. However, its photosynthetic rate was 1.2−1.7 times greater than that of late rice with the same genotype during the early panicle development stage. The net assimilation rate of regenerated seasonal rice measured from the booting stage to the maturity stage exceeded that of single-season late rice with the same genotype by a factor ranging from 2.7 to 6.0 times, as well as its photosynthetic rate for early panicle development. Additionally, it was found that both photosynthetic product conversion efficiency and appearance quality were significantly higher in regenerated seasonal rice compared to single-season late rice with the same genotype heading at the same time[39]. These findings further confirm that regenerated rice belongs to a source-sink interaction type and highlight its close association between yield formation and its source-sink-flow characteristics. The results of our study indicated that the transport rates of dry matter and non-structural carbohydrates from various 'source' organs, namely roots, stems, and leaves and their contribution rates to the grain yield ('reservoir') of ratooning season rice of different varieties were significantly higher than those of the main crop, as well as late rice with the same genotype and heading time as ratooning rice (the ratooning rice counterpart)[37]. Quantitative analysis using Q-PCR demonstrated a general increase in sucrose transporter gene OsSUTs family expression in the root, stem, and leaf tissues of all 'source' organs during the full heading stage of regenerated season rice. In contrast, lower expression levels were observed for the INV gene family and RSUS gene family during the regeneration season compared to the main crop and the ratooning season counterpart. It is worth noting that INV gene family encodes enzymes related to sucrose transformation, while the RSUS gene family is associated with sucrose synthesis. Furthermore, we found that enzyme activity related to these processes was lower in roots, stems (sheaths), and leaves of ratooning season rice after full heading compared to the main crop and the ratooning season counterpart. This was primarily attributed to a significant improvement in material transport efficiency between 'source' organs and the 'reservoir' tissue in ratooning season rice. This facilitated the rapid transportation of multiple substances into grains, stimulating starch synthesis during the early grain-filling stage. Previous studies have suggested that increasing enzyme activity in sucrose conversion and starch synthesis during the early stages of grain filling while decreasing their activity later on is more favorable for dry matter production and achieving fuller grain filling[40]. These findings emphasize the significance of efficient source organs for achieving high yields in ratooning season rice. Moreover, our study also documented the elevation of the enzyme activity associated with sucrose and starch metabolism in grains of ratooning season rice compared to the main crop and the counterpart of ratooning season rice. Quantitative analysis using Q-PCR demonstrated the upregulation of genes encoding these enzyme activities in ratooning rice compared to that in the main crop and the ratooning rice counterpart. Furthermore, material transfer rates from stems and leaves in ratooning rice was significantly higher than in the main crop and ratooning rice counterpart. When comparing non-structural carbohydrates (NSC) between different varieties of ratooning season rice with the main crop and their ratooning rice counterparts at corresponding time courses, it was found that the stems (sheaths) and leaves of ratooning rice contributed a higher proportion towards NSC accumulation in grains compared to that of the main crop or the ratooning season rice counterparts. Further investigation utilizing 13CO2 pulse labeling technology demonstrated that ratooning rice reduced transport of 13C assimilates to underground parts during late grain development stages retentive to the main crop and the ratooning season rice counterparts. Therefore, it is suggested that this cultivation characteristic can be effectively applied to enhance the number of effective panicles of both the main crop and ratooning rice, promote source increase, sink expansion, and increase yield.
Table 1. Contents of 13C assimilates of rice plant organs and soil in different cropping patterns[37].
Cultivar Allocated part Contents at full heading stage
(mg pot–1)Contents at maturity stage
(mg pot–1)The content difference of the same accessions at maturity and full heading stages (mg pot–1) RR LR RR LR RR LR JFZ 13C-soil 4.87 Ecd* 0.40 d 3.02 Cbc 2.75 Cc −1.84 Db 2.35 Db* 13C-roots 1.54 Ed 2.66 Ed* 0.44 Cc 4.33 Cc* −1.10 Db 1.67 Db* 13C-stubbles 4.11 Ecd − 1.30 Cc − −2.82 Db − 13C-stem-sheath 38.57 EAb* 34.42 Ab 9.31 Bb 20.95 Ab* −29.25 Ac −13.46 Ad* 13C-leaves 6.30 Ec 7.73 Ec* 3.33 Cc 5.36 Cc* −2.97 Db −2.36 Dc 13C-panicles 70.25 Ba* 48.35 Ba 102.26 Aa* 60.97 Ba 32.01 Aa* 12.62 Aa YY2640 13C-soil 3.35 Ec* 0.41 Ed 2.36 Cc* 1.30 Cc −0.99 Db 0.89 Db* 13C-roots 0.97 Ec 1.41 Ed 0.30 Cc 2.74 Cc* −0.68 Db 1.34 Db* 13C-stubbles 2.04 Ec − 0.91 Cc − −1.13 Db − 13C-stem-sheath 19.12 Db 20.53 Db* 9.13 Bb 16.39 Bb* −9.99 Cc −4.13 Cc* 13C-leaves 3.00 Ec 9.95 Ec* 1.88 Cc 4.05 Cc* −1.12 Db* −5.89 Cd 13C-panicles 89.07 Ca* 62.77 Ca 104.93 Aa* 70.22 Ba 15.87 Ba* 7.46 Ba RR: ratooning rice. LR: late rice synchronously heading with RR. Different letters (lowercase or uppercase) in the same column of the same cultivar have significant difference at the 0.05 or 0.01 probability levels among the different allocated parts at ratooning season under the same nitrogen application treatment. * indicates significant difference at the 0.05 probability level between different nitrogen application treatments at the same allocated part in the ratooning season. It is worth mentioning that the high production of materials ('source') stored in the reservoir can provide a solid foundation for grain filling, and strong material mobility ('flow') is crucial for grain filling. Flow formation is closely related to endogenous hormones and vascular bundle density at the panicle base. Yang et al. discovered that abscisic acid (ABA) plays a role in regulating the transport of stored assimilates from the stems-sheaths to the grains[41,42], and its concentration (content) was positively correlated with the transport rate of non-structural carbohydrates (NSC) stored in the stems (sheaths) of the grains. Grain enrichment also depends on the ratio of ABA to ethylene (GAs)[43−45]. The present study also found that ratooning rice had higher ABA content than the ratooning season rice counterpart from the full heading stage to 15 d after the full heading. In addition, the GA content was significantly lower than that in the main crop and the ratooning season counterpart during this period (Fig. 3). These hormone levels promoted grain filling in ratooning rice. Previous studies have shown that increased zeatin riboside (ZR) and indole-3-acetic acid (IAA) content promote cell division and plant growth. At the same time, ABA may regulate substance movement and grain filling through calmodulin protein CIPKs[46]. IAA can also enhance cotton seed germination and seedling growth, improve biomass and photosynthetic capacity of leaves, increase sugar content in plants, activate key enzymes involved in sucrose metabolism, and balance key endogenous hormones antagonistically, leading to favorable changes in endogenous plant hormone levels as well as sucrose metabolism[47]. The results simultaneously demonstrated a significant elevation in ZR and IAA levels in the panicle of regenerated rice compared with the main crop and the ratooning season counterpart. This indicates a correlation between hormone regulation and the physiological foundation of material transport in regenerated rice. However, transporting NSC to the grains necessitates transportation through vascular bundle tissue as it is the primary conduit for material transport between the source and reservoir. A higher density of vascular bundles at the stem base of panicles were observed in different varieties of regenerated rice than in the main crop and the ratooning season rice counterpart. This suggests that efficient material transport in regenerated rice is associated with well-regulated endogenous hormones and an increased density of vascular bundles in the panicle necks. However, further investigation is required to elucidate its underlying mechanisms.
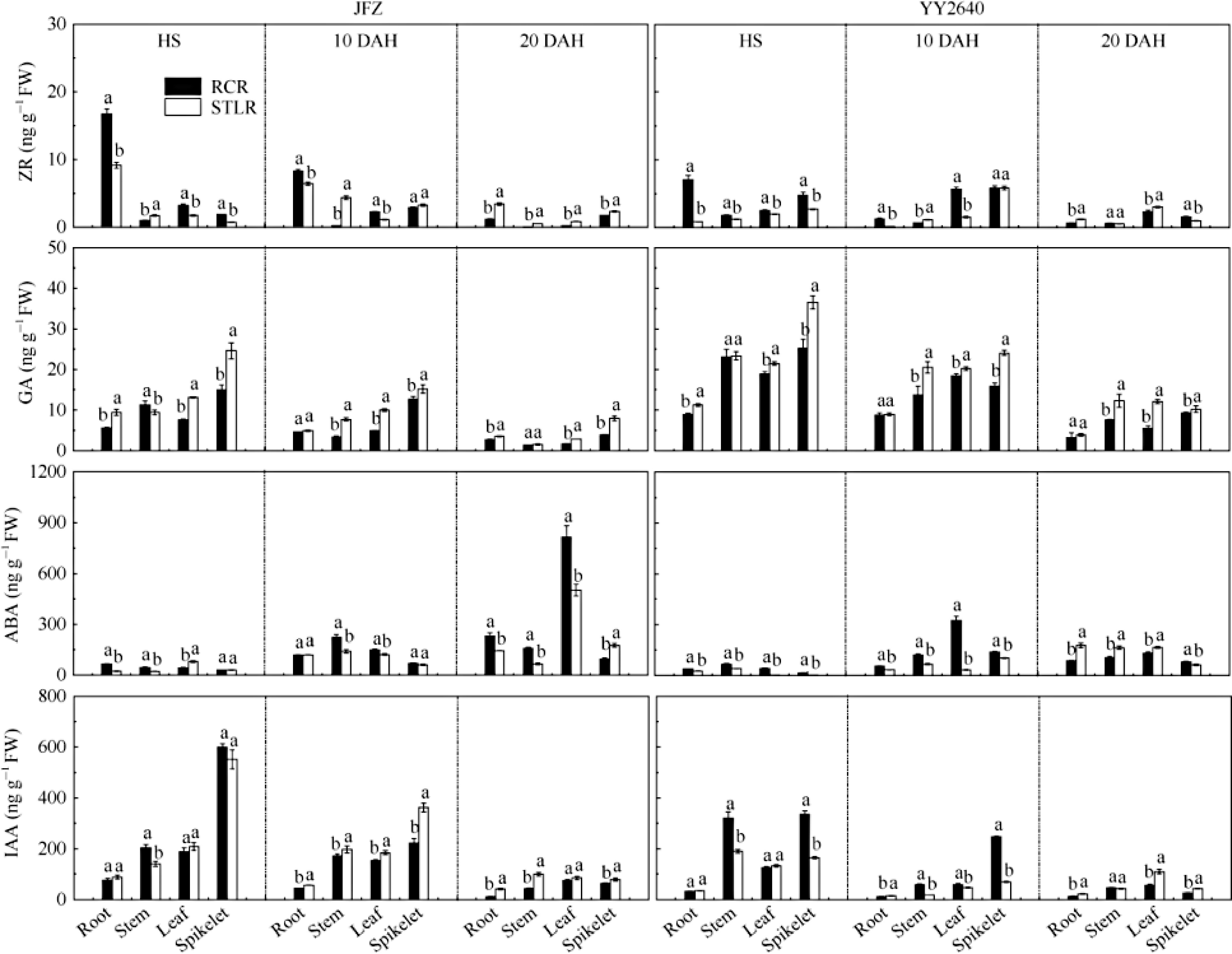
Figure 3.
Differences of hormone contents in roots, stems, leaves and spikelets of rice under different cropping patterns[37]. JFZ: conventional rice, Jiafuzhan; YY2640: hybrid rice, Yongyou 2640. RCR and STLR refer to ratooning rice and late rice with the same genotype of main crop synchronously heading with RR in late season. HS: heading stage. DAH: day after heading. Differences in lowercase or uppercase letters after each data in the same column in the same year indicate significant difference at the 0.05 or 0.01 probability levels, respectively.
Correlation between regeneration activity and rhizosphere microecological properties
-
The root system, comprising the main and ratooning rice crops consists of two components: one is the residual old root system on the rice stubble primordial base of the main crop, and the other is the regenerated seasonal root system that regenerates from the stem base nodes of the main crop stubbles or stem nodes of newly developed regenerated tillers. The growth of new roots during the regeneration season can be regulated by adjusting the stubbles left after harvesting the main crop. Lower stubble height significantly promotes new root growth in this season. Field fertilization management measures and above-ground biomass of the main crop also affect root activity in regenerated rice[48,49]. The vitality of roots in the main crop is closely associated with its regeneration ability, as a well-developed root system exhibits strong regeneration ability. Therefore, a significant positive correlation exists between root activity and regeneration ability during the late growth stages of the main crop[50,51]. The early growth and development of regenerated tillers rely on residual roots from the main crop, while the number of effective panicles and yield output in regenerated rice is linearly correlated with root function at the maturity stage for the main crop and the full heading stage for regenerated rice. Therefore, they are commonly employed as significant indicators for assessing the regenerative capacity of regenerated rice and screening rice varieties suitable for ratooning[51]. Consequently, we propose that by maintaining a constant total nitrogen application rate for the main crop (225.00 kg·ha−1), altering the distribution of nitrogen fertilizers during late growth stages can enhance dry matter production and allocation in regenerated rice. This alteration influences the function of rhizosphere soil microecosystems. It significantly contributes to sustaining high-root vitality while promoting the growth and development of ratooning rice, ultimately leading to increased overall yield from a single planting with two harvests[28].
In recent years, extensive research has been conducted on the correlation between soil communities and crop health. Crop root exudates modify the biogeochemical cycle and biocommunity composition of the surrounding soil, thereby creating a distinct and intricate ecological rhizosphere zone. Chaparro et al.[52] demonstrated significant differences in root microorganism composition between rice seedlings at different stages of growth. Edwards et al.[53], conducted a greenhouse experiment and observed comparable abundance and diversity of root microorganisms 13 d after transplanting compared to 43 d after transplanting. In field conditions, Zhang et al.[54] observed significant shifts in the root microorganism composition during the vegetative growth phase, while relative stability was noted during the initial stage of reproductive growth. Subsequently, minor variations occurred during the mature stage, with specific patterns depending on different rice varieties and geographical planting conditions. Studies have suggested that rhizosphere microbial community structure is selectively shaped by root exudates, leading to unique characteristics and genetic imprinting across different plant species[55,56]. Conversely, alterations in rhizosphere microbial diversity have significant impacts on plant-root secretion release as well as material circulation, energy flow, and information transfer within rhizosphere soil ecosystems, ultimately providing feedback mechanisms that regulate plant growth and development processes influencing productivity performance[57,58].
Provirus studies suggested that root secretions are pivotal in shaping rhizosphere microbiota selection. Altering patterns of rhizosphere secretions to recruit beneficial rhizosphere microorganisms present new opportunities for enhancing plant performance and positively impacting plant development and crop production as shown in Fig. 4[56,59]. Domestic and international research is focused on niche differentiation and life history countermeasures of rhizosphere microbial population evolution through various ecological enhancement techniques. Additionally, researchers are investigating the effect of feedback regulation on plant organ development in soil, which ultimately impacts the quality and quantity of their products. The primary focus of this research lies in understanding the priming effect formation, its influence on plant growth and development, and its regulatory mechanism. Previous studies have indicated that yield formation in ratooning rice is associated with greenhouse gas emission mitigation and dry matter production and partitioning characteristics within paddy fields[60,61]. However, it remains unclear how changes in dry matter (including root exudates) distribution within rhizosphere soil affects the microecology of ratooning rice's rhizosphere soil and subsequently impacts carbon emissions. It is warranted to specifically consider the relationship between plant source-sink-flow dynamics, further investigation into differences in photosynthetic product distribution to aboveground-underground parts in the main crop, and the response mechanisms of rhizosphere microecology to photosynthetic product distribution at later stages.
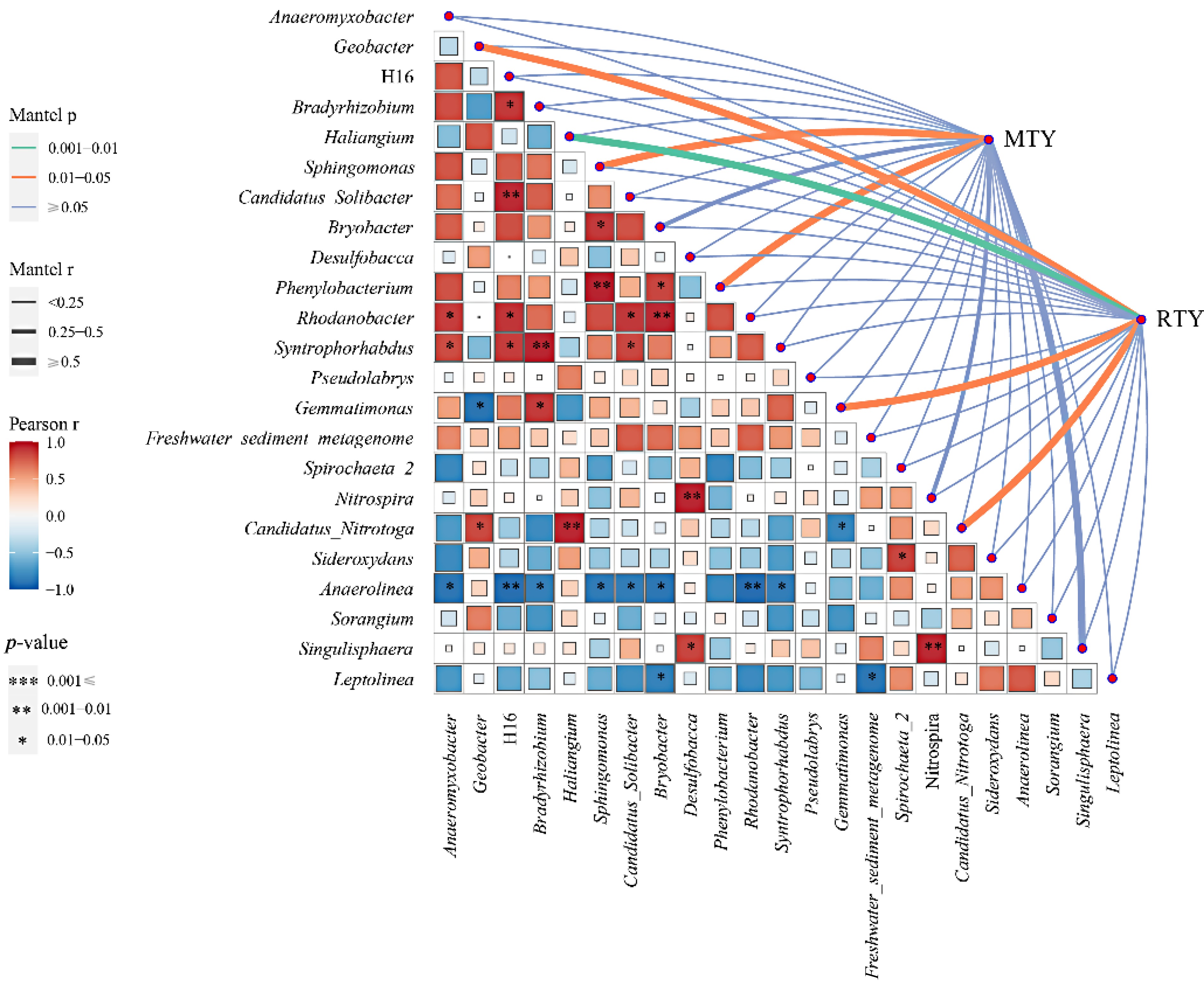
Figure 4.
The correlation analysis between the dominant bacteria at the mature stage of main crop rice and the yield of main crop rice and ratooning rice. MTY: main rice yield; RTY: ratooning rice yield[59].
-
In summary, China has made remarkable advancements in both theoretical research and yield levels of ratoon rice compared to the rest of the world[4,5]. However, numerous scientific challenges still require collaborative efforts from domestic and international counterparts. For instance, the regeneration coefficient (i.e., regeneration rate) of most ratoon rice in current production remains relatively low, fluctuating between 1.2 and 1.5. This means that under the current production level, an effective mother stem generally contains 5−6 potentially available axillary buds; however, less than two buds per stem can sprout and tiller into effective panicles. Therefore, enhancing the regeneration rate of rice is crucial for achieving high yields and super high yields of ratoon rice. Furthermore, while there have been significant advancements in researching medium maturing ratooning rice (middle seasonal rice) with notable application results regarding utilization types of ratoon rice, studies on early maturing rice (early rice) and late-maturing ratooning rice remain insufficiently explored and necessitate further investigation[2]. Overall, several issues need to be addressed to facilitate the further development of ratoon rice in China.
Firstly, the research on the enhancement mechanism of rice quality in the ratooning season and its regulatory technology needs further deepening. Recent studies have consistently demonstrated that low-stubble ratooning rice significantly improves the quality of ratooning season rice, particularly in terms of milled rice rate and head rice rate, due to a more synchronized ripening period during this season[32,62]. Moreover, additional investigations have suggested that the quality of ratooning season rice not only surpasses that of the main crop but it also exceeds that of its counterparts (late rice)[32,62,63]. However, further exploration is still required to elucidate its formation mechanism. Secondly, a limited selection of cultivated ratoon rice varieties suitable for mechanized production conditions fails to meet the actual production demand. Therefore, enhancing research efforts and establishing a theoretical and technical system for breeding ratoon rice varieties with robust regenerative capacity is imperative while directly selecting specialized ratoon rice varieties[4]. Thirdly, the implementation of key technologies needs improvement since a significant yield gap still exists between different management entities. This can be attributed to insufficient attention given by certain agricultural entities towards the regeneration season and inadequate grasp of mechanized production techniques for ratoon rice among farmer households. Consequently, implementing key technologies falls short, resulting in fluctuating ratoon rice yield levels. Henceforth, it is essential to strengthen training programs and demonstrations and scale-up initiatives related to regenerated rice cultivation technology while transforming the mindset of relevant management subjects and enhancing the success rate of key technological advancements[4].
Additionally, inadequate drainage and irrigation facilities lead to improper water discharge or delayed post-harvest irrigation for ratooning rice crops, affecting their yields. Therefore, it becomes essential to prioritize enhancing water conservancy infrastructure construction and other related facilities[4]. Moreover, there is a dearth of specialized small machinery for rice ratooning suitable for mountainous and hilly terrains. Mass production of specialized small harvesters for regenerated rice in such areas has not yet been realized. The commonly used harvesters in production are predominantly large and medium-sized ones. However, when utilized on small fields in mountainous and hilly regions, the challenge of water management leads to severe damage to the stubbles of first-season rice by the rolling harvester, consequently resulting in a decrease in the number of effective panicles and significantly reducing the yield of regenerated rice. Therefore, further development of specialized machinery specifically designed for ratooning rice on small fields in mountains and hills is imperative[4]. Furthermore, exploring the cultural significance and environmentally friendly qualities inherent in the ratoon rice industry will help construct a shared brand identity for this sector. Increasing publicity surrounding the green quality attributes of ratoon rice through effective marketing strategies can lead this brand initiative towards achieving high-quality standards along with premium pricing structures while simultaneously enhancing overall efficiency within the industry[4].
-
The authors confirm contribution to the paper as follows: study conception and design: Lin W, Chen H, Weng P, Chen T, Lin W; data collection: Lan C, Fallah N, Guo C, Zhang Z; analysis and interpretation of results: Lin W, Weng P, Chen T, Lin W. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study areavailable from the corresponding author on reasonable request.
The research was financially supported by the National Natural Science Foundation of China (32001109, 31871542, and 31871556), as well as the National Key Research and Development Projects of China (2017YFE0121800 and 2018YFD0301105).
-
The authors declare that they have no conflict of interest. Wenxiong Lin is the Editorial Board member of Technology in Agronomy who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen T, Weng P, Lan C, Nyumah F, Guo C, et al. 2024. Studies and prospectives of mechanically harvested ratooning rice in China. Technology in Agronomy 4: e015 doi: 10.48130/tia-0024-0012 

Studies and prospectives of mechanically harvested ratooning rice in China
- Received: 13 December 2023
- Revised: 25 April 2024
- Accepted: 30 April 2024
- Published online: 26 June 2024
Abstract: Continuous improvement of rice breeding methods and consecutive innovation of cultivation technology in China has resulted in a significant increase in the proportion of ratooning rice planting mode. Therefore, ratoon rice has become a prominent rice planting pattern in China. This study reviewed the current situation and prospects regarding ratoon rice's high-yielding formation and physiological mechanisms. We focused on four key aspects: screening and breeding of ratoon rice cultivars, classification, determining suitable stubble height, and water and fertilizer management considerations for rice ratooning. Additionally, we delved into characteristics related to dry matter production and allocation in ratoon rice. This study also explored the regenerative activity and root vigor within the rhizosphere zone while evaluating their relationship with micro-ecological characteristics specific to ratoon rice. We propose the implementation of proper regulation precise over the rhizosphere environment aimed at scaling up sustainable progress in the realm of research, aimed at expanding the utilization of mechanized low-cut stubbles (< 25 cm) for the advancement of the ratoon rice sector. This process is essential for stimulating the availability of vital soil nutrients to promote timely sprouting axillary buds while facilitating tiller growth coordination along with bud elongation, collectively referred to as achieving 'four highs' (high regeneration coefficient, high effective panicle percentage, high harvest index, and high daily yield) alongside 'one low' (low carbon emission) for safe and sustainable development purposes. Furthermore, we have also put forward corresponding policy recommendations in light of the challenges encountered during the development process of the ratoon rice industry.