








-
Crop production in controlled environment agriculture (CEA) plays an important role in feeding an increasing world population as urbanization is expanding, and arable land is decreasing. Crops can be grown year-round with reduced water and pesticide usage. Spinach (Spinacia oleracea) is a leafy green vegetable and one of the most consumed salad crops. It is a nutrient-rich vegetable with high amounts of carotenoids, vitamin C, vitamin K, folic acid, iron, and calcium. It has numerous health benefits, including antioxidant activity and antidiabetic effects[1], reduction of macular degeneration, and blood pressure (
www.healthline.com/nutrition/foods/spinach ). Spinach production was valued at over US${\$} $ For CEA hydroponic production, crop yield is an important factor in selecting a crop since CEA has a large upfront investment, and higher yields expedite the return on investment. An alternative way to increase crop yield is to use plant growth-promoting bacteria (PGPB). In general, PGPB, including rhizosphere and endophytic bacteria, promote plant growth, increase nutrient uptake, enhance stress tolerance, and inhibit pathogen growth[5−7]. Mechanisms of plant growth promotion are diverse, and complex, and often depend on specific bacterial strains and their interactions with the host plant. Direct growth promotive functions include nitrogen fixation, phosphate solubilization, hormone production, and enhanced nutrient uptake[6,7]. Indirect mechanisms include induction of systemic resistance and inhibition of plant pathogens. Many studies across various systems and crops have reported plant growth promotion due to PGPB application. Commercial products are also available to producers that contain PGPB for both greenhouse and field crop production. However, consistent results are not always achieved due to environmental factors, interactions with existing microorganisms, application methods, and a lack of understanding of the PGPB mechanisms. Many PGPB have been shown to promote plant growth under in vitro conditions, likely because competitive microbes are eliminated, and the environmental factors are highly regulated. Bacterial endophytes from the Institute for Advanced Learning and Research (IALR) Plant Endophyte Research Center's collection were found to increase the growth of tall fescue KY31 up to 300%[8]. It has been observed that when PGPB identified in vitro are applied in the field, the percentage yield increase is greatly reduced or completely disappears[9−11]. When compared to field production, CEA crop production in hydroponic systems is more similar to in vitro conditions, and consistently higher yields may be expected.
PGPB applications in hydroponic production systems under CEA conditions have been found to improve plant growth and crop quality and reduce disease severity in various crops grown in hydroponic systems through a range of mechanisms[12−15]. Aini et al. evaluated the effects of nutrient concentration, PGPB, and arbuscular mycorrhizal fungi on the growth and nutrient uptake of hydroponically produced romaine lettuce (Lactuca sativa L. var. longifolia) and found the application of PGPB and arbuscular mycorrhizal fungi as a consortium reduced the nutrient concentration level needed by the plant[16]. A commercial biostimulant product TNC BactorrS13 containing several Bacillus spp. added to nutrient solutions enhanced salt tolerance of hydroponically grown lettuce[17]. Single PGPB strains applied in hydroponic production systems have also resulted in improved plant growth. Pseudomonas psychrotolerans IALR632 significantly increased the fresh weight of five lettuce cultivars in different CEA conditions and hydroponic systems including greenhouse deep water cultivation, greenhouse nutrient film technique (NFT) and indoor vertical NFT[18]. PGPB have been utilized to improve spinach growth in field production, with Pseudomonas species in particular found to increase the productivity of spinach (Spinacia oleracea)[19]. Inoculation of spinach cv. 'Tyee' with Pseudomonas strains S2 and S4 resulted in shoot dry weight increases of 69% and 63% compared to non-inoculated controls, respectively[20]. For hydroponically grown spinach, there are a few reports on PGPB applications. One example is a study conducted by Urashima & Hori where spinach grown in small-scale sterilized hydroponic culture was inoculated with Pseudomonas spp. The spinach plants that received the bacterial inoculation had increased root and shoot growth by 50%[21]. In another study, spinach grown in a floating hydroponic system at 50% normal nutrient concentration and inoculated with PGPB did not have improved yield measurements but did have improved quality measures such as nutrient leaf content and reduced nitrate concentrations[22]. However, other studies have shown that PGPB not only increased total harvest yields but also improved the food quality of basil (Ocimum basilicum) grown in a floating culture. Biofertilizers, including a mix of beneficial bacteria, improved leaf quality, and the leaves had lower nitrate content, increased vitamin C, phenols, flavonoids, and increased mineral elements (Ca, Mg, P, N, K, Fe, and Zn)[23].
While past studies have identified optimal nutrient requirements for spinach in hydroponic production, spinach production in CEA faces several challenges, including nutrient imbalances, insect pests and plant pathogens, water quality, and environmental factors (e.g. high temperatures can cause bolting while low temperatures can stunt growth). Further research is needed to improve the profitability of spinach production. The current hypothesis is that the application of PGPB to spinach grown hydroponically in an indoor vertical farm will result in higher yields. To evaluate this, bacterial endophytes Pseudomonas psychrotolerans IALR632 and Enterobacter asburiae IALR1379 were applied individually to two spinach cultivars ('Red Snapper' and 'Seaside') grown under indoor vertical farm conditions in NFT hydroponic systems. Growth parameters were evaluated to determine yield improvements, and presence of the target bacteria were confirmed with GFP-tagged bacterial endophytes.
-
Two cultivars of spinach (Spinacia oleracea L.) were used in this study: 'Seaside' F1, a slow bolting smooth-leaf variety with dark green leaves and 'Red Snapper' F1, a red-veined, baby-leaf spinach with a novel leaf shape purchased from Johnny's Selected Seeds (Fairfield, ME, USA).
Seed germination
-
Spinach seeds were sowed in wetted Oasis Grower Solutions Horticubes XL Foam Media (276 cells) (Kent, OH, USA). Three seeds were sowed per cell and covered with vermiculite to keep the seeds moist and germinate in the dark and then placed in propagation systems from AmHydro (Arcata, CA, USA). One seedling was kept in each cell for bacterial inoculation. The seed flats were irrigated with tap water until germination under natural lights in a greenhouse. After germination, the seedling flats were irrigated for 1 min, four times daily with Virginia Tech fertilizer solution[18]. The EC of the fertilizer was maintained at 1.0 ± 0.1 mS/cm and pH 5.9 ± 0.1. The EC and pH were monitored with Economy pH/EC Meter (Spectrum Technologies, Inc., Aurora, IL, USA) and adjusted as needed.
Bacterial culture preparation and inoculation
-
Two bacterial endophytes IALR632 and IALR1379 from the IALR Plant Endophyte Research Center's collection were chosen for spinach experiments in CEA indoor vertical conditions based on previous growth promotion results in other crops[8,18]. IALR632 was isolated from the leaves of a wild Sorghum halepense plant in the foothills of the Appalachian Mountains in Central Virginia, USA (geographic location: 37.125372, −79.298415). It was identified as Pseudomonas psychrotolerans (GenBank accession MZ519967). Endophyte IALR1379 was isolated from the roots of a wild Tragopogon dubius plant in Yanceyville, NC, USA (geographic location: 36.407037, −79.341545). It was identified as Enterobacter asburiae (GenBank accession OQ414238). Plant parts were surface-sterilized, and bacteria were isolated according to the method in a previous publication[8]. All endophytes were preserved in glycerol stocks at −80 °C for later use.
Treatments were applied to seedlings at the first true leaf stage. Bacterial endophyte inoculants were prepared by taking a loop of bacterial glycerol stock and culturing it in 25 mL of half-strength Luria-Bertani (LB) broth in a shaking incubator at 200 rpm at 30 °C overnight. The overnight culture was transferred to a flask with 500 mL of fresh half-strength LB medium and grown at 30 °C at 200 rpm for about 5 h until the OD600 was approximately 1.0 (the number of colony-forming units per mL for IALR632 and IALR1379 were 1.85 × 109 and 4.12 × 108, respectively). One mL of the bacterial culture was pipetted at the base of each seedling. The control treatment was 1 mL sterile half-strength LB broth.
Indoor vertical NFT systems
-
Seedlings with 3−4 true leaves were transplanted into vertical racks with 3 levels outfitted with NFT hydroponic systems (HydroCycle Vertical NFT Lettuce and Herb Systems, FarmTek, Dyersville, IA, USA). Each level contained 48 plants and the slope of all NFT channels were set to 2.5%–3%. Light was supplied with high output reproductive pink batten LED grow lights with a 6:1 red-to-blue ratio (GE First Generation ArizeTM Lynk and Life 1.5 HO Horticulture Batten LED Luminaire, Hort Americas, Bedford, TX, USA). The lights were fixed 30 cm above the NFT channel surface. The reservoir tank was 265 L (Fig. 1).

Figure 1.
Spinach plants were grown in the indoor vertical NFT units with three levels and a reservoir tank. The plants at both ends of the channels were not counted due to low light intensity.
Plant growth nutrient solution and environmental conditions
-
The vegetative hydroponic fertilizer solution developed at Virginia Tech (Blacksburg, VA, USA) was used for spinach growth[18]. The fertilizer solution does not account for mineral elements present in the city water source. Concentrated (100×) stock solutions were maintained in two parts (Stocks A and B). Equal amounts of Stocks A and B were used to create a final dilute solution (1:90) with EC 1.7 ± 0.1 mS/cm and adjusted to pH 5.9 ± 0.1 in reservoirs under each vertical tower. The EC was adjusted by adding nutrient solution or water. The pH was adjusted by adding 1 N H2SO4 or 1 N KOH as needed. The indoor vertical NFT system was set up as follows: temperatures at 21/19 °C (day/night), light length at 16 or 14 h, relative humidity at 58%, and the CO2 level at 350–500 ppm. The two bacterial endophytes were evaluated in three separate experiments. The real-time growth conditions (humidity and temperature) of each experiment were recorded with a WatchDog 2,475 Plant Growth Stations (Spectrum Technologies, Inc, Aurora, IL, USA), and data analysis was performed with SpecWare software version 9 (Table 1).
Table 1. Vertical NFT environmental conditions.
Dates
(transplant to harvest)No. of
plants/treatmentTemperature
(°C)Light
(h)PPFD
(μmol/m2/s)DLI
(mol/m2/d)Relative humidity
(%)29 Apr. – 19 May 2022 18 20.5 16 250 14.4 58.5 4 Oct. – 27 Oct. 2022 36 21.0 16 250 14.4 55.5 13 Sept. – 5 Oct. 2023 36 20.9 14 250 12.6 53.9 Plant growth and leaf greenness (SPAD) measurements
-
Approximately 3 weeks after transplant, plants were harvested, and growth measurements were conducted. Shoot and root fresh weights (FW) were measured separately. Dry weight (DW) was determined after shoots and roots were dried at 60 °C for 2 d. Leaf greenness was also measured at harvest by averaging three representative leaves per plant using SPAD 502 Plus Chlorophyll Meter (Spectrum Technologies, Inc., Aurora, IL, USA).
Colonization of spinach seedlings with GFP-tagged bacterial endophytes
-
To monitor bacterial colonization of spinach seedlings, IALR1379 was transformed with p519ngfp following a previous report[18]. Both IALR1379- and IALR632-tagged p519ngfp[18] were used to inoculate spinach seedlings of 'Red Snapper' and 'Seaside' one week after seeds were sowed using the same method described above. Samples were taken at 4, 11, and 18 d after bacterial inoculation with three biological replicates. Seedlings were separated into roots and shoots, and ground with sterile water. Ground samples were centrifuged at 2,000 rpm for 3 min. Supernatant was diluted into 10×, 100×, and 1,000×, and 0.1 mL was plated on LB + kanamycin (30 mg/L). The colonies with GFP were counted under a fluorescent microscope Olympus SZX12 (Tokyo, Japan).
Experimental design and statistical analysis
-
Three different experiments were carried out for spinach grown in indoor vertical NFT systems with bacterial inoculations. Due to limited vertical NFT units, one unit was used to grow spinach seedlings treated with one bacterial inoculation to avoid cross-contamination. Data collected from different experiments were placed together for statistical analysis with different experiments as blocks. Data for growth measurements and leaf greenness by bacterial treatments were analyzed with one-way ANOVA, and the least significant difference (LSD) was used for multiple treatment comparisons using SAS® Studio from SAS OnDemand for Academics. All significant levels were set at p < 0.05.
-
Two bacterial endophytes (IALR632 and IALR1379) were selected from the IALR's collection based on their plant growth-promoting traits and in vitro growth promotion. Both bacterial endophytes have multiple plant growth promoting traits (Table 2). Both have a relatively high ability to solubilize insoluble phosphate compounds as well as N-fixing ability. IALR1379 produces high levels of the plant growth promotive hormone auxin, and IALR632 has high siderophores and ACC deaminase activity.
Table 2. Plant growth promoting traits of bacterial endophytes.
Plant growth
promoting traitsaPseudomonas
psychrotolerans
IALR632Enterobacter
asburiae
IALR1379Auxins (μg/mL) 10.0 ± 2.9b 56.8 ± 1.2 N fixation + + P solubilization (μg/mL) 263.6 ± 12.4 238.1 ± 18.2 ACC deaminase ++ - Siderophore (%) 97 ± 0.5 26 ± 5.3 a Plant growth promoting traits were measured following previous publication[8]. b Data showed mean ± standard error. Spinach growth promotion by bacterial endophytes
-
For 'Red Snapper', the shoot FW and DW, total FW and DW were increased by 19.3%, 19.4%, 28.5%, and 17.3% with IALR632 inoculation, and 13.3%, 12.1%, 23.5%, and 11.1% with IALR1379 inoculation at harvest three weeks after transplant, compared with the control treatment, respectively (Fig. 2). Although they both increased root FW and DW, the two bacteria showed different effects on root growth. IALR1379 significantly increased root FW but not root DW. IALR632 significantly increased root DW but not root FW.
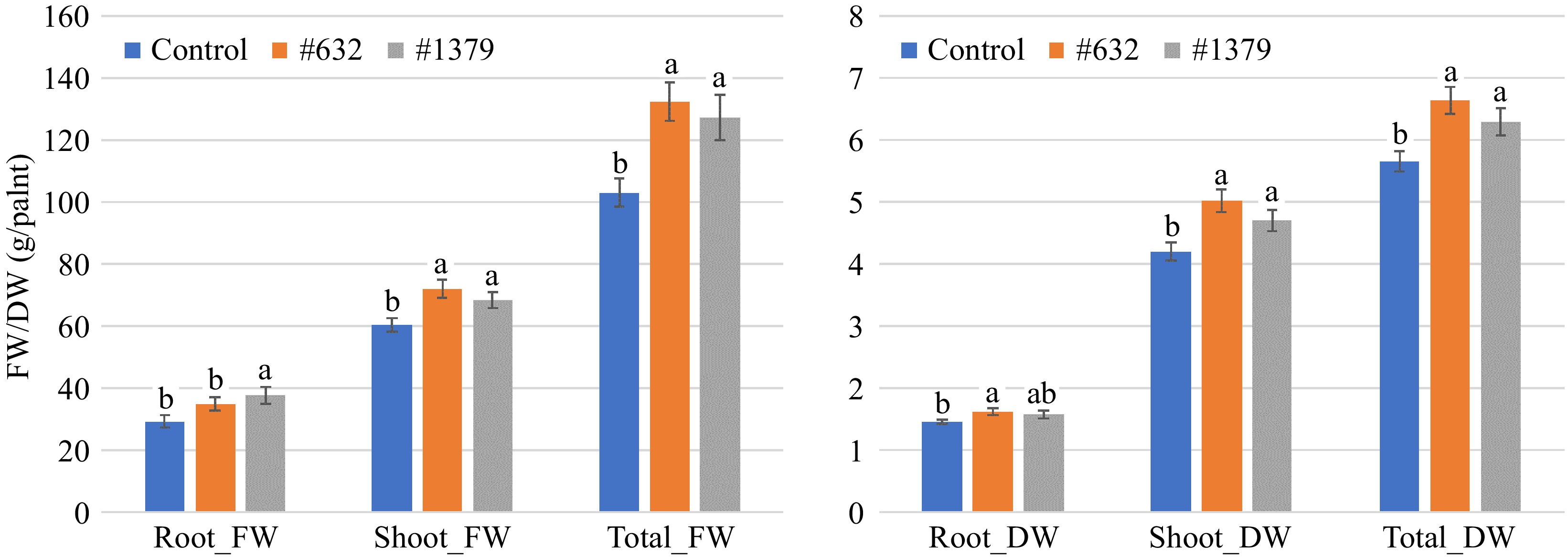
Figure 2.
Effects of bacterial endophytes on the growth of spinach cultivar 'Red Snapper' under indoor vertical farm conditions in NFT hydroponic systems. Bars with the same letters are not significantly different (p > 0.05). FW: fresh weight and DW: dry weight.
For 'Seaside', both IALR632 and IALR1379 bacterial inoculations significantly enhanced shoot FW by 17.8% and 14.1%, compared with the control treatment, respectively (Fig. 3). Only IALR632 significantly increased shoot DW and total DW by 12.7% and 10.6%, compared with the control treatment, respectively. However, bacterial inoculations did not affect the root growth of 'Seaside'.
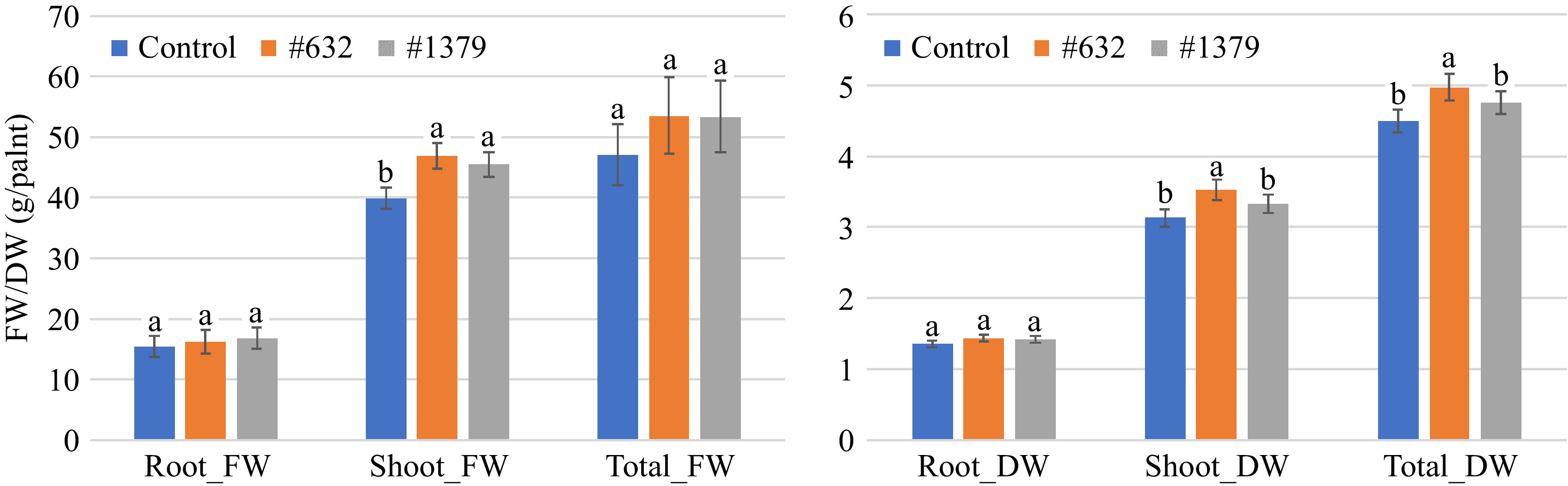
Figure 3.
Effects of bacterial endophytes on the growth of spinach cultivar 'Seaside' under indoor vertical farm conditions in NFT hydroponic systems. Bars with the same letters are not significantly different (p > 0.05). FW: fresh weight and DW: dry weight.
Effects of bacterial inoculation on spinach leaf greenness
-
Leaf greenness was measured with a SPAD meter at harvest. The results showed that IALR632 inoculation significantly increased leaf greenness in both 'Red Snapper' and 'Seaside' cultivars, compared to the controls (Fig. 4). However, there were no differences between the control plants and IALR1379 treated plants in leaf greenness of both cultivars.
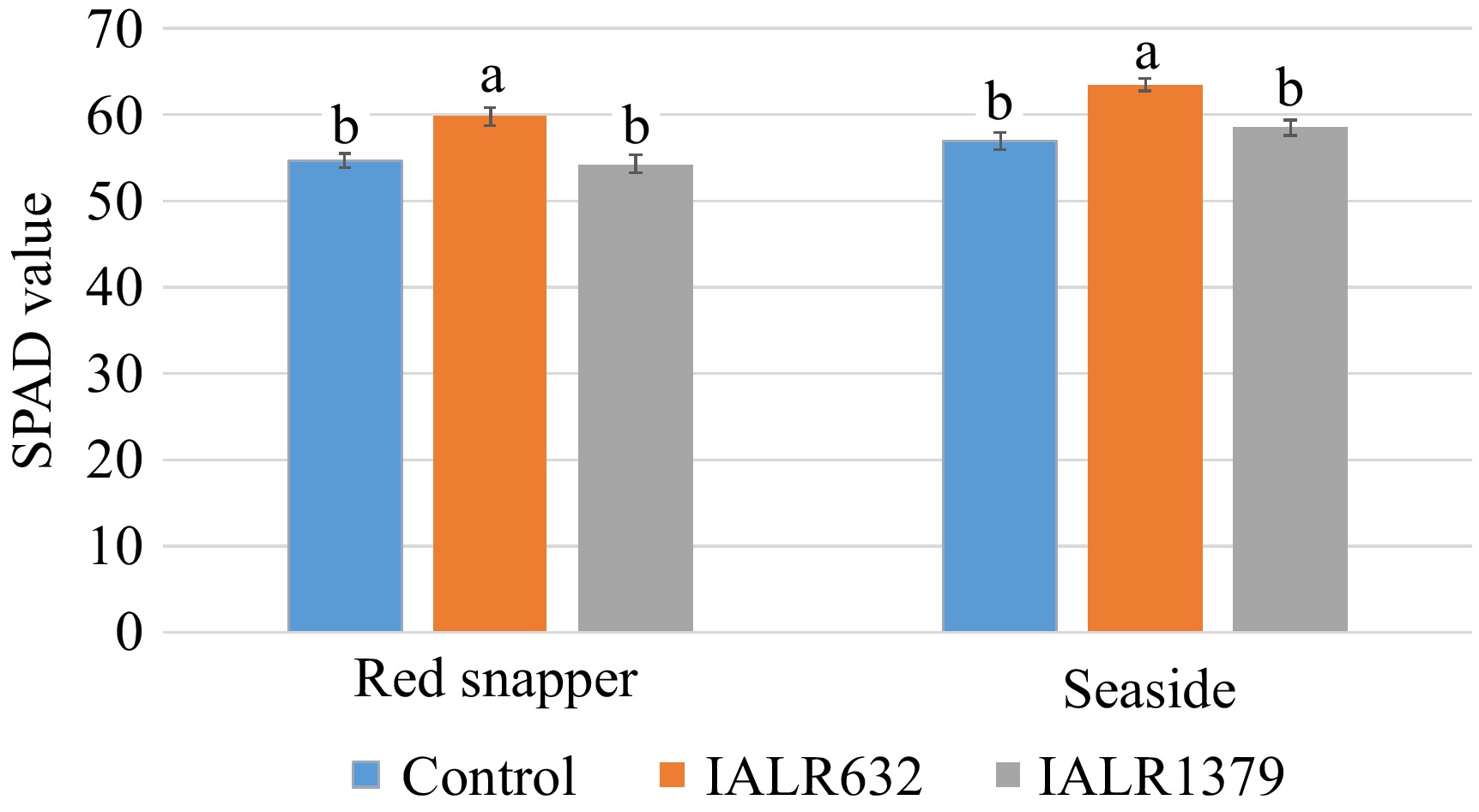
Figure 4.
Effects of bacterial inoculation on leaf greenness of spinach plants. Bars with the same letters are not significantly different (p > 0.05).
Bacterial colonization inside spinach seedlings with GFP-tagged endophytes
-
Bacteria IALR632 and IALR1379 were confirmed to be endophytic with GFP-tagged IALR632 and IALR1379. Figure 5 showed that bacterial endophytes were re-isolated from leaves of 'Seaside' 18 d after bacterial inoculation. The colonies of IALR632-GFP were smaller and less bright while those of IALR1379-GFP were bigger and much brighter. Table 3 shows colony-forming units per mg FW of roots and shoots of both 'Red Snapper' and 'Seaside' at 4, 11, and 18 d after bacterial inoculation. Overall, IALR632 had more colonies in shoots than those in the roots of both spinach cultivars, particularly at 18 d. IALR1379 was predominantly colonized in the roots.
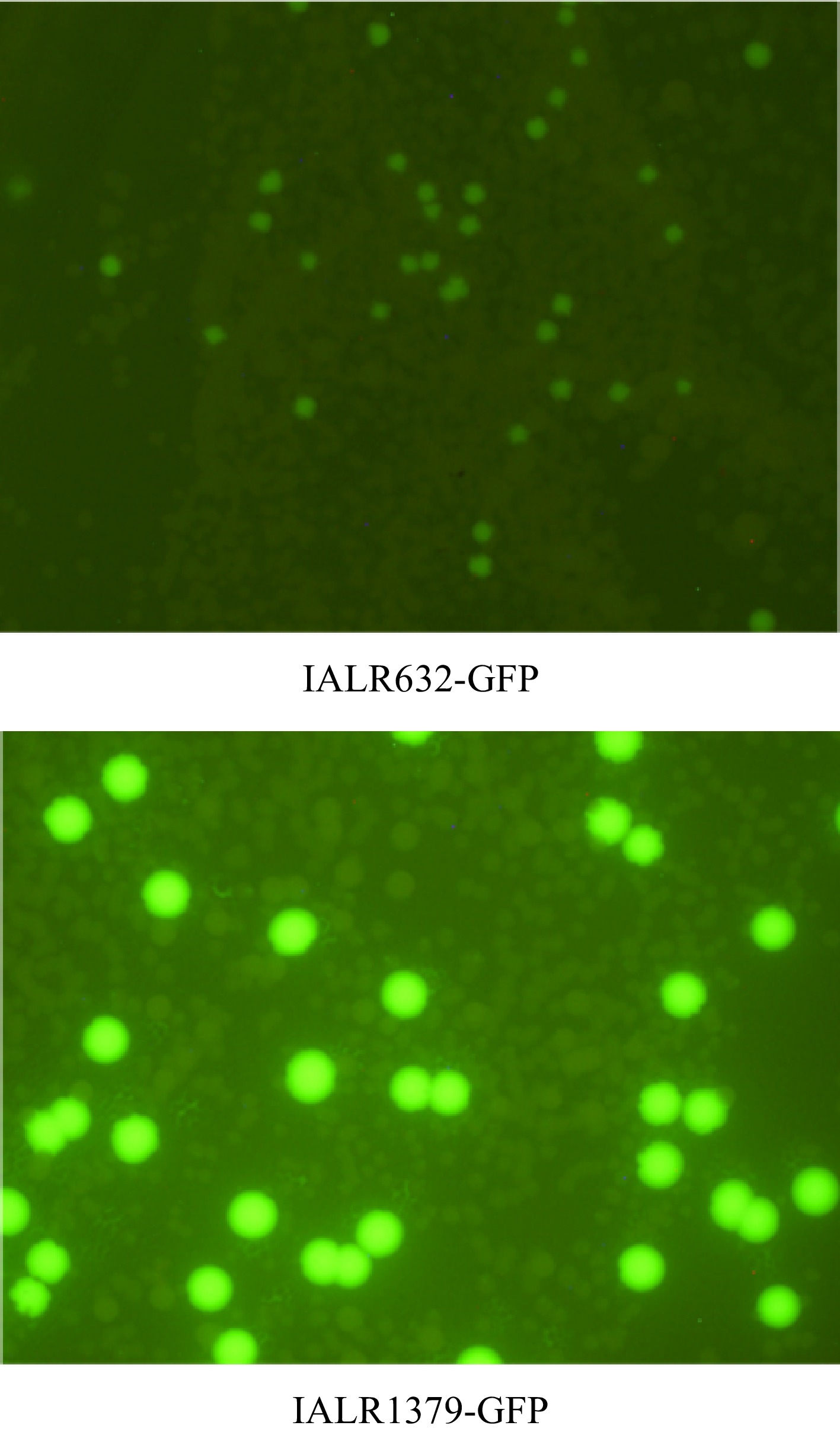
Figure 5.
Colonies of bacterial endophytes re-isolated from leaves of 'Seaside' at 18 d after inoculation of GFP-tagged bacteria. The images were taken under fluorescence microscope Olympus SZX12 with 5 s exposure and 11.2× magnification.
Table 3. Bacterial colonization of spinach seedlings using GFP-tagged bacterial endophytes.
Bacteria Cultivars Tissues Days after inoculation 4 11 18 IALR632 Red snapper Shoot 24.7 ± 8.5 209.4 ± 191.7 10.6 ± 9.1 Root 2.7 ± 0.4 200.2 ± 197.7 0.6 ± 0.4 Seaside Shoot 4.7 ± 3.4 2.3 ± 1.3 15.1 ± 8.1 Root 17.1 ± 16.7 4.3 ± 1.5 1.7 ± 0.6 IALR1379 Red snapper Shoot 4.9 ± 4.7 12.0 ± 6.0 5.0 ± 4.2 Root 26.6 ± 26.4 841.7 ± 836.6 27.4 ± 8.7 Seaside Shoot 7.8 ± 0.8 0.8 ± 0.5 6.9 ± 4.0 Root 38.3 ± 3.1 6.8 ± 1.9 19.6 ± 5.2 The values present averages of colonization form units (CFU) per mg fresh tissues from three biological replicates ± standard errors. -
There has been an increasing trend to use PGPB in agriculture production because PGPB not only can promote plant growth, increase nutrient uptake and enhance stress tolerance[5,6], but also is more environmentally friendly. Under in vitro conditions, PGPB greatly increases tall fescue KY31 growth, up to 300%[8]. Often the efficacy of PGPB in field applications is dramatically reduced and inconsistent, probably due to changing environmental factors and competition with other microorganisms. In CEA facilities, the growing conditions are controlled, and microorganism growth is restricted. Indoor vertical facilities control temperature, humidity, light duration and intensity, as well as CO2 concentrations. These controlled environmental conditions could provide stable conditions needed to increase PGPB efficacy in promoting plant growth. We tested two bacteria (Pseudomonas psychrotolerans IALR632 and Enterobacter asburiae IALR1379) with two spinach cultivars ('Red Snapper' and 'Seaside') under indoor vertical conditions in NFT hydroponic systems. IALR632 and IALR1379 inoculations achieved significantly higher yields, increasing shoot FW by 19.3% and 13.3% for 'Red Snapper' (Fig. 2), by 17.8% and 14.1% for 'Seaside' (Fig. 3), compared with their control treatments, respectively. Overall, IALR632 is more effective at increasing yield in spinach compared to IALR1379 under indoor vertical hydroponic systems. Moreover, both bacterial endophytes exhibited colonization in both spinach cultivars. Colonization within the plant corresponded with the original plant tissue they were isolated from (Table 3).
Mechanisms for bacterial endophytes for plant growth promotion have been elucidated in general. Both IALR632 and IALR1379 have a high ability to solubilize phosphate compounds, which could enhance phosphate uptake. IALR632 belongs to the Pseudomonas genus, which is widely used to promote plant growth[22] and has robust functional growth-promoting traits, which can result in increased plant biomass yields from photosynthesis of higher leaf greenness (Fig. 4). It also has ACC deaminase activity and higher siderophore production. ACC deaminase activity lowers ethylene levels in plants and may increase plant stress tolerance to changes in environmental conditions. Higher levels of siderophore compounds increase iron absorbance. Recently, Putra et al. found that two Pseudomonas strains promoted lettuce growth in hydroponic NFTs by enhancing IAA production and affecting 12 metabolic pathways, particularly in myo-inositol and acetic acid metabolic pathways[24]. IALR1379 is identified as Enterobacter asburiae and produces higher levels of plant growth promotive hormone auxin, which could promote plant growth.
PGPB application in hydroponically produced spinach has the potential to improve yield in vertical farming facilities. There are still many challenges for consistent efficacy for PGPB application in hydroponically grown spinach in CEA, including different fertilizer use, the dosage of bacterial cultures, effective colonization, variations of colonization in plants (Table 3), and application methods. In addition, more cultivars need to be tested for possible plant-microbe interactions.
-
In summary, plant growth-promoting bacterial endophytes Pseudomonas psychrotolerans IALR632 and Enterobacter asburiae IALR1379 have potential applications for improving yields of spinach grown in indoor vertical farming systems.
-
The authors confirm contribution to the paper as follows: study conception and design: Mei C, Lowman S, Evens M; data collection: Chretien RL, Zhou D, Mei C; analysis and interpretation of results: Zhou D, Chretien RL, South K, Mei C; draft manuscript preparation and edit: Chretien RL, Zhou D, South K, Mei C, Lowman S, Evens M. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This research was funded by USDA Specialty Crop Block Grant, Agreement Number: 2021B-570 for Utilizing Endophytes to Promote Hydroponic Vegetable Growth and Increase Profitability and Tobacco Commission for Promoting Controlled Environment Agriculture in Southern Virginia.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Dongfang Zhou, Robert L. Chretien
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou D, Chretien RL, South K, Evans M, Lowman S, et al. 2024. Beneficial bacterial endophytes promote spinach plant growth under indoor vertical hydroponics. Technology in Horticulture 4: e015 doi: 10.48130/tihort-0024-0012 

Beneficial bacterial endophytes promote spinach plant growth under indoor vertical hydroponics
- Received: 20 January 2024
- Revised: 22 May 2024
- Accepted: 12 June 2024
- Published online: 02 July 2024
Abstract: Hydroponic and vertical farming technologies have grown rapidly in recent years as there are many advantages including year-round production, short growth cycles, greater efficiency of water and nutrient use, prevention of soil-borne diseases and insects, longer shelf life, etc. Spinach (Spinacia oleracea) is a popular leafy green and a good source of vitamins and minerals. Although methods of spinach production in hydroponic systems have been improved including lighting, fertilizer solutions, and others, plant growth could be further promoted to increase profitability and compensate for initial startup costs. One way to increase spinach yields is to use plant growth-promoting bacteria, which can promote plant growth, enhance nutrient uptake, increase stress tolerance as well as inhibit pathogen growth. Two bacteria (Pseudomonas psychrotolerans IALR632 and Enterobacter asburiae IALR1379) were tested with two spinach cultivars ('Red Snapper' and 'Seaside') using indoor vertical nutrient film technique systems. Inoculations of IALR632 and IALR1379 significantly increased the shoot fresh weight of 'Red Snapper' by 19.3% and 13.3%, and that of 'Seaside' by 17.8% and 14.1% at harvest, compared with their control treatments, respectively. Both bacterial inoculations enhanced root growth of 'Red Snapper', but not 'Seaside'. Additionally, inoculation of IALR632 significantly increased leaf greenness of both 'Red Snapper' and 'Seaside', but IALR1379 did not. Both bacterial endophytes colonized the inside of the roots and translocated to shoots of 'Red Snapper' and 'Seaside'. Overall, IALR632 is more effective at increasing spinach yields, root growth, and leaf greenness compared to IALR1379 under indoor vertical hydroponic systems.