









-
Southern highbush blueberries (Vaccinium corymbosum L., interspecific hybrids) thrive in tropical and subtropical climates, such as those found in the Northeastern region of Argentina (NEA). The growing global demand for healthy and nutritious foods has led to increased consumption of berries worldwide. Agronomists and farmers are particularly focused on ensuring high-quality attributes, given that the majority of the product is transported and marketed as fresh fruit.
Unfortunately, in recent decades, the frequency of hailstorms and windstorms has risen due to climate change. Consequently, the use of anti-hail covers (AHC) is recommended to mitigate potential damage to both the fruit and the trees. These covers, serving as a safeguard against both biotic and chemical threats, have emerged as an environmentally friendly alternative to pesticides, particularly in organic farming[1]. However, this physical defense method often results in a reduction of photosynthetically active radiation (PAR) and the induction of temperature variations, which can significantly impact crop development, yield, and quality standards[2]. Colored netting, used in light-regulated physiological processes, has been found to influence growth and delay the partitioning of photoassimilates by up to three weeks[3]. Additionally, shade covers, as suggested by certain studies, protect against excessive radiation levels throughout the day, thereby reducing heat stress and inhibition of photosynthesis[4]. Milivojević et al.[5] reported that the utilization of AHC diminishes the incidence of light and the maximum daily temperature in 'Duke' blueberry plants. These researchers also reported increased fruit yields compared to fruit grown in open fields, despite unchanged total phenolic and anthocyanin levels. A study on the 'Eliot' variety, using various colored shade covers, found higher fruit weight than controls but lower fruit soluble solids and a delay in the peak of the harvest season[6]. Meanwhile, compared to control fruit, utilizing covers with varied colors and shading resulted in more fruit per plant, lower weight per fruit, and increased firmness for the 'Brigitta' variety[7]. These authors also reported higher fruit yields compared to fruit grown in open fields, although total phenolic and anthocyanin levels remained unchanged.
The contrasting results described above, along with the extended practice of using AHC, prompted the further investigation of the consequences that the use of covers might have on relevant quality indicators for two varieties of blueberries most adapted to temperate climates: 'Emerald' and 'Snowchaser'. These two southern highbush varieties are widely produced in the NEA and are highly valued for their superior qualities, such as appealing taste and texture. 'Snowchaser', more recently introduced than 'Emerald' in Argentina, is the earliest available variety, attracting the interest of farmers and consumers, and it has been scarcely studied. The standard variables determining fruit compositional quality, such as soluble solids, titratable acidity, and their ratio have been studied, at harvest and after a postharvest period. In addition, the unique metabolite profile, provided by sugars, sugar alcohols, amino acids, and organic acids, as well as the total phenolic content, was included in the current analysis. Given the importance of cell wall integrity and fruit firmness, variations in the activities of cell wall-modifying enzymes were also explored. This type of study, to our knowledge, is infrequently found in the literature and may contribute to a better understanding of the potential implications for blueberry quality when grown under cover. The findings of this research will enhance our comprehension of the impact of this practice.
-
The studies were conducted in commercial blueberry orchards in Concordia, Entre Ríos, Argentina (Agroberries S. A., latitude 31°32'42'' S, longitude 58°08'07'' W). 'Emerald' and 'Snowchaser' southern highbush varieties were assessed in contiguous orchards separated by a forest curtain. One had a three-meter-high anti-hail cover, while the other was fully open to the sky. According to the manufacturer, the anti-hail cover (TextilKopruck,
https://textilkopruch.com/ ) utilized was white with a mesh size of 330 μm and decreased the incidence of photosynthetically active radiation by roughly 17%. Incident photosynthetically active radiation (PAR) was measured in μmol·m−2·s−1 (data logger Bar-Rad Dual USB,https://cavadevices.com ). Soil and leaf temperatures were measured with an infrared thermometer (Testo 830 T1,https://testo.com ). From August through October (2018 and 2019 seasons), ten readings were taken daily per treatment at the same hours at regular intervals. Supplementary Figure S1 shows the average precipitation amounts and temperature fluctuations during the growth and harvest periods of each variety. Standard techniques for this crop were applied during fruit development. All fruits were harvested at commercial maturity, as determined by complete blue skin color (100% blue coloration). Internal quality parameters and firmness were determined on immediately harvested fresh fruits. Each experiment's samples were frozen and stored at −80 °C for further analyses.Phenological stages examination
-
To determine bloom progression in each cultivar, 12 flower clusters were flagged and fortnightly observed throughout the entire period of bloom using the methodology developed at INTA EEA Concordia[8]. The alphanumeric Spiers scale[9] was adapted and used to determine the reproductive stages (the phenological scale from 1 to 9, see Supplementary Fig. S2 for further information).
Conservation period
-
Homogeneously selected fruits at the fully mature stage were used to fill eight clamshells (125 g each) per replicate. Clamshells were weighed and then stored at 0.5 ± 0.5 °C and 90%–95% RH, being sampled at 7 and 30 d for postharvest evaluations. Twenty fruits from four clamshells were used to determine fruit firmness as described below. The remaining four clamshells were weighed individually on the first day of storage (day 0) and then again during postharvest to measure weight loss, which was calculated as follows, correcting the values by clamshell weight:
[(Weight at day 0 − Weight at x time of storage)]/(Weight at day 0) × 100
Determination of agronomic parameters
-
The fruit was squeezed, and the juices obtained were used in the experiments. Five fruits were used for each determination, with three replicates per treatment. Using a digital refractometer, the total soluble solid (SS) content of the juice was calculated and expressed as a percentage of weight in volume (% w/v). The titratable acidity (TA) was measured by titrating with 0.1 N NaOH to pH 8.2. The results were expressed as the percentage of anhydrous citric acid per volume in the berry juice. Four to six fresh berries were weighed, sliced, and vacuum-dried for 24 h in Petri dishes to determine dry matter content. The fruit was then placed in a desiccator and weighed. This was performed in triplicate for each treatment. The results were given as a percentage (g of dry matter for every 100 g of fresh weight).
Firmness analysis
-
Firmness analysis was carried out by compression measure with a TA.XT Plus Texture Analyzer (Stable Micro Systems Ltd, UK), according to the following conditions: load, 5 kg; cylindrical plunger diameter, 75 mm; compression force at 10% of axial deformation; speed, 1 mm·s−1. Thirty fruits for each treatment and variety samples were used for individual determination, and maximal force was registered in Newtons (N).
Sampling
-
For the following determinations, tissue from a minimum of three different fruits were processed as a pool, and three replicates were conducted in all instances.
Total protein extraction and determination of protein concentration
-
One-half gram of tissue was ground with liquid nitrogen and mixed with 2 mL of extraction buffer (1 M NaCl, 12.5 mM citric acid, 50 mM Na2HPO4, and PMSF 1:100 v/v, pH 6.5). Homogenates were shaken for 1.5 h at 4 °C and centrifuged at 10,000 × g for 20 min. For protein and enzyme activity measurements, the recovered supernatants were kept at 4 °C. The protein content was measured using the Bradford protein assay method[10], with bovine serum albumin as the reference, in an EPOCH microplate reader.
Determination of pectin methyl esterase (PME), pectinase, and β -Galactosidase (β-Gal) activity
-
Total protein extracts obtained as described above were utilized to determine the activities of cell wall-related enzymes, as detailed by Montecchiarini et al.[11]. PME and pectinase activity were measured following the pectoplate technique[12], in agar plates containing citrus pectin. Pectoplate wells were loaded with 2 μg of total protein (Supplementary Fig. S3). β-gal activity was assayed by measuring the hydrolysis rate of the synthetic substrate p-nitro phenyl-D-galacto-pyranoside (pNPG) at 415 nm using an εpNp = 18,000 M−1·cm−1. The reaction mixture contained 35 μL of crude extract, 175 μL of 0.1 M sodium acetate-acetic acid pH 4.5, 150 μL of 0.1% BSA, and 150 μL of 13 mM pNPG. The mixture was incubated at 37 °C for 25 min, the reaction was stopped with 0.2 M sodium carbonate. One unit of β-gal activity is considered as the amount of extract used to release 1 nmol p-nitro phenol (pNp) per min. Boiled protein extracts were used as negative controls, and the linearity of activity over the time of analysis was verified in all cases. Results were expressed in units per microgram of total protein in the crude extracts.
Total phenolic content (TPC) determination
-
Total phenolic content was determined spectrophotometrically using the Folin-Ciocalteu reagent[13]. Absorbance was read at 730 nm in a microplate reader (EPOCH), and results were expressed as gallic acid equivalents/gram of fresh weight.
Metabolite extraction, derivatization, and analysis
-
Samples were essentially processed in the same way as in earlier work[14] with ribitol as the internal standard and N-methyl-N-trimethylsilyl-trifluoroacetamide (MSTFA) as the derivatizing reactive. The OpenChrom program (
www.openchrom.net ) was used to determine peak area and the Automated Mass Spectral Deconvolution and Identification System software (AMDIS) was used to identify compounds. For a minimum of three independent replicates, chromatograms were assessed by comparing individual peak areas for each metabolite relative to the internal standard. The representative ions for each component were selected for relative quantification[11]. Chemical identity was confirmed by updating data using the online software Mass Spectra & Retention Time Index (MSRI) from the Golm Metabolomic Institute (Germany) available athttp://gmd.mpimp-golm.mpg.de/analysisinput.aspx .Statistical procedures
-
Data were analyzed using the statistical program InfoStat[15]. Measured variables were expressed as mean ± standard error (SE), and the statistical significance of the parameters under study was compared using the t-student test between pairs of samples. A p-value of 0.05 was chosen as the level of statistical significance. After z-score normalization, data were utilized to perform Principal Component Analysis, Heatmaps, and Correlation analyses in MetaboAnalyst 5.0[16].
-
Several parameters were evaluated in fully mature fruit to gain a comprehensive understanding of the influence of growth circumstances on blueberry quality. Following this investigation, it was discovered that the 'Snowchaser' variety grown in the field (F) had considerably higher values of soluble solids, SS/TA ratio, and equatorial diameter in the 2018 season, while the titratable acidity and firmness were lower than under anti-hail cover (AHC) (Table 1). Equatorial diameter and firmness were lower in 'Emerald' fruit cultivated in the field than in fruit grown under AHC.
Table 1. Blueberry agronomic parameters. Data for the cvs 'Emerald' and 'Snowchaser' were obtained throughout the 2018 and 2019 seasons.
Agronomic parameter E-F E-AHC S-F S-AHC Season 2018 Soluble solids (SS, °Brix) 12.0 ± 0.2 (a, A) 11.9 ± 0.2 (a, A) 12.8 ± 0.3 (a, B) 11.3 ± 0.5 (b, B) Titratable acidity (TA,% acid) 1.44 ± 0.07 (a, A) 1.51 ± 0.22 (a, A) 1.01± 0.04 (a, B) 1.38 ± 0.11 (b, A) SS/TA ratio 8.3 ± 0.46 (a, A) 7.9 ± 1.2 (a, A) 12.7 ± 0.8 (a, B) 8.2 ± 1.1 (b, A) pH 2.65 ± 0.10 (a, A) 2.61 ± 0.01 (a, A) 2.87 ± 0.05 (a, A) 2.77 ± 0.14 (a, A) Equatorial diameter (mm) 16.44 ± 0.73 (a, A) 17.59 ± 1.21 (b, A) 16.44 ± 0.80 (a, A) 16.08 ± 0.60 (b, B) Weight (g) 1.63 ± 0.10 (a, A) 1.43 ± 0.20 (a, A) 1.53 ± 0.02 (a, A) 1.52 ± 0.06 (a, A) Dry matter content (%) 12.97 ± 0.14 (a, A) 12.97 ± 0.12 (a, A) 12.63 ± 0.17 (a, A) 12.03 ± 0.52 (a, B) Firmness (N) 1.83 ± 0.23 (a, A) 2.16 ± 0.30 (b, A) 1.60 ± 0.20 (a, B) 1.83 ± 0.23 (b, B) Season 2019 Soluble solids (SS, °Brix) 13.4 ± 1.4 (a, A) 12.5 ± 0.9 (a, A) 9.9 ± 1.1 (a, B) 10.1 ± 0.8 (a, A) Titratable acidity (TA,% acid) 0.88 ± 0.11 (a, A) 1.13 ± 0.03 (b, A) 0.90±0.08 (a, A) 1.05 ± 0.08 (a, A) SS/TA ratio 15.22 ± 1.9 (a, A) 11.06 ± 1.0 (b, A) 11 ± 0.6 (a, B) 9.6 ± 0.3 (b, B) pH 2.87 ± 0.04 (a, A) 2.67 ± 0.01 (b, A) 2.79 ± 0.06 (a, A) 2.82 ± 0.05 (a, A) Equatorial diameter (mm) 14.82 ± 0.18 (a, A) 15.15 ± 0.58 (a, A) 15.43 ± 0.73 (a, A) 15.04 ± 1.06 (a, A) Weight (g) 1.72 ± 0.04 (a, A) 1.76 ± 0.15 (a, A) 1.76 ± 0.09 (a, A) 1.73 ± 0.06 (a, A) Dry matter content (%) 13.70 ± 0.04 (a, A) 11.99 ± 0.62 (b, A) 11.92 ± 0.04 (a, B) 12.42 ± 0.24 (a, A) Firmness (N) 2.19 ± 0.24 (a, A) 2.24 ± 0.21 (a, A) 1.86 ± 0.22 (a, B) 2.16 ± 0.30 (b, A) F: Field growth; AHC: Anti-hail cover growth. Different letters indicate significant differences (p < 0.5) between treatments for the same variety (lowercase letters) or between varieties with the same treatment condition (capital letters). The quantities in bold indicate major seasonal changes. The values are the mean and standard deviation of at least three replicates. Considering data from the 2019 season, the main difference for 'Snowchaser' was observed in firmness, which was significantly higher in AHC-grown fruit than in open field-produced fruit. The SS/TA ratio was lower for both varieties under AHC compared to field-produced fruit. 'Emerald' had a firmness value equivalent to the greatest seen in 'Snowchaser', without treatment effect. Significant variations were seen in the dry matter content and pH, which were lower under AHC while the titratable acidity was higher.
Phenological stage changes under different growth conditions
-
The temperature decreased by an average of 0.8 °C and 0.3 °C from August to November in 2018 and 2019, respectively, under cover compared to the open field. Meanwhile, for fruit cultivated under AHC, photosynthetically active radiation (PAR) was reduced by an average of 17% in both seasons. Consequently, a delay of several days was observed at each phenological stage for both varieties under these conditions (Supplementary Fig. S2). However, this delay was more pronounced for 'Emerald' than for 'Snowchaser' in both seasons, being almost negligible for the latter cultivar in 2019. The average delay for 'Emerald' was 10 d in 2018 and 17 d in 2019. The average delay for 'Snowchaser' was 5 d in 2018. With the approach of the final phases of fruit ripening (8−9), stages were closer under each growth condition and organoleptic maturity was reached at approximately the same time.
Total phenolic content (TPC) and cell wall-related enzyme activities in blueberries under different growth conditions
-
TPC was investigated as it represents an essential trait that not only fluctuates among different varieties but also within tissues, influenced by seasonal variations, climate, and growth conditions. No significant differences were observed in the fruit grown under diverse conditions (Fig. 1a). TPC was significantly higher in 'Emerald' than in 'Snowchaser' for both seasons, with the difference being more noticeable in 2019.
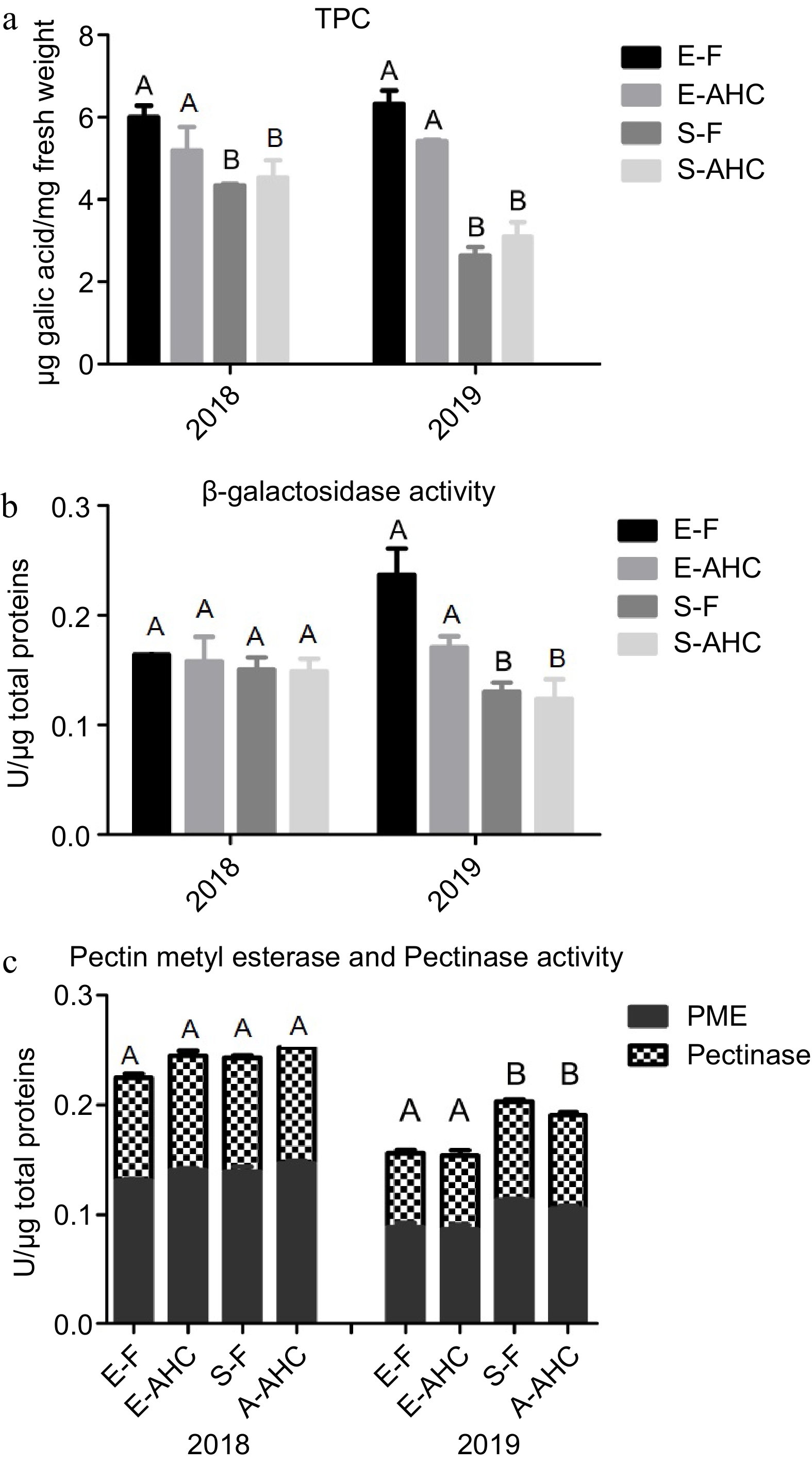
Figure 1.
Total phenolic content and enzymatic activities of cell wall related enzymes in mature blueberries from 'Emerald' and 'Snowchaser' throughout the 2018 and 2019 growing seasons. (a) Total phenolic content. (b) β-gal activity. (c) PME and Pectinase activities (for further information, see Material and methods). E-AHC: 'Emerald' grown under anti-hail cover; E-F: 'Emerald' grown in the field; S-AHC: 'Snowchaser' grown under anti-hail cover; S-F: 'Snowchaser' grown in the field. The values are the mean and standard deviation of at least three replicates per cultivar in the 2018 and 2019 seasons. Significant differences (p < 0.5) between varieties are denoted by capital letters. In either season, no significant changes were recorded between treated and untreated fruit.
Additionally, given that changes in the cell wall can impact fruit firmness and postharvest life, specific enzyme activities related to wall metabolism were measured (Fig. 1b & c). The treatment had no impact in either season or variety, as no significant variations were observed for β-galactosidase, pectin methylesterase (PME), or pectinase in field-grown fruit. However, in 2019, 'Emerald' exhibited increased β-galactosidase activity but reduced PME and pectinase activity compared to 'Snowchaser'.
Metabolites: sugar, amino acids, organic acids, sugar alcohols, and other compounds in blueberries vary according to growth conditions
-
To investigate the impact of anti-hail covers on fruit biochemistry, the levels of various metabolites in samples from the 2019 season were analyzed using GC-MS (Table 2). The representative ions for each component were selected for relative quantification with the internal standard (ribitol), and normalized by grams of fresh weight. It was observed that the relative levels of specific sugars changed with treatment within the same variety. Six sugars increased in 'Emerald' under AHC compared to field-grown fruit (cellobiose, fructose, maltose, sucrose, trehalose, and turanose), while one decreased (raffinose). Rhamnose was not detected in this variety. In contrast, five sugars decreased under AHC in 'Snowchaser' (cellobiose, maltose, ribose, sucrose, and xylose), and two increased (raffinose and trehalose). Galactose was higher in 'Snowchaser' under cover than in the field and rhamnose, only detected in 'Snowchaser', was higher in the field than under cover. Fructose, galactose, and glucose were higher in 'Snowchaser' than in 'Emerald' grown in the field. The most abundant sugar alcohol found in all samples was myo-inositol. The relative content of xylitol, mannitol, and myo-inositol in 'Emerald' fruit produced under AHC was higher than in the field. Simultaneously, mannitol was not detected in 'Snowchaser' under AHC, while xylitol and glycerol were lower than in the field. Citric, gulonic, and quinic acids increased considerably in 'Emerald' when AHC was applied. Meanwhile, gulonic acid levels were higher in 'Snowchaser' field-grown fruit, with no significant differences by treatment in the remaining acids identified. Regarding amino acids detectable by this approach, the relative amount of alanine, aspartic acid, gamma amino butyric acid (GABA), and serine increased in 'Emerald' when exposed to AHC. Serine levels were higher in 'Snowchaser' cultivated in the field, but aspartic acid was not detected, and other amino acids did not differ between treatments. Considering other compounds, it is noteworthy that glucuronic acid was only detected in 'Snowchaser' cultivated in the field, and hexadecanoic acid was higher under AHC in this variety.
Table 2. Metabolites level variation in 'Emerald' and 'Snowchaser' blueberries grown in the field or under anti-hail covers in the 2019 season.
Metabolites E-F E-AHC S-F S-AHC Sugars Cellobiose 12.90 ± 0.60 (a, A) 20.20 ± 0.80 (b, A) 14.2 ± 0.20 (a, B) 12.90 ± 0.20 (b, B) Fructose 328.24 ± 5.83 (a, A) 433.81 ± 79.74 (b, A) 338.43 ± 3.31 (a, B) 385.59 ± 27.60 (a, A) Galactose 234.01 ± 17.42 (a, A) 322.86 ± 66.09 (a, A) 272.90 ± 0.91 (a, B) 298.73 ± 9.97 (b, A) Glucose 588.96 ± 11.86 (a, A) 821.06 ± 163.92 (a, A) 619 ± 2.79 (a, B) 708.09 ± 62.58 (a, A) Maltose 77.10 ± 2.90 (a, A) 87.9 ± 3.00 (b, A) 73.8 ± 5.80 (a, A) 57.4 ± 6.70 (b, B) Raffinose 1.60 ± 0.06 (a, A) 0.90 ± 0.05 (b, A) 1.1 ± 0.20 (a, B) 2.60 ± 0.40 (b, B) Rhamnose ND ND 0.33 ± 0.08 (a, B) 0.22 ± 0.05 (b, B) Ribose 0.51 ± 0.09 (a, A) 0.61 ± 0.01 (a, A) 1.73 ± 0.13 (a, B) 0.95 ± 0.18 (b, B) Sucrose 49.98 ± 1.29 (a, A) 70.00 ± 7.86 (b, A) 54.94 ± 3.12 (a, B) 48.03 ± 0.27 (b, B) Trehalose 12.36 ± 0.09 (a, A) 16.73 ± 3.35 (b, A) 3.79 ± 0.30 (a, B) 17.24 ± 2.93 (b, A) Turanose 18.79 ± 0.30 (a, A) 22.76 ± 2.32 (b, A) 19.51 ± 2.13 (a, A) 21.69 ± 1.59 (a, A) Xylose 2.61 ± 0.09 (a, A) 2.58 ± 0.02 (a, A) 9.55 ± 1.11 (a, B) 3.32 ± 0.88 (b, A) Sugar alcohols Xylitol 1.30 ±0.13 (a, A) 1.67 ± 0.13 (b, A) 1.54 ±0.16 (a, A) 1.20 ± 0.05 (b, B) Glycerol 0.52 ± 0.05 (a, A) 0.44 ± 0.05 (a, A) 0.55 ± 0.04 (a, A) 0.42 ± 0.03 (b, A) Mannitol 5.22 ± 0.17 (a, A) 10.92 ± 1.45 (b, A) 4.75 ± 0.09 (a, B) ND Myo-Inositol 26.11 ± 0.37 (a, A) 36.08 ± 1.67 (b, A) 30.36 ± 3.28 (a, A) 28.41 ± 0.14 (a, B) Organic acids Citric acid 93.72 ± 4.73 (a, A) 110.23 ± 1.66 (b, A) 102.76 ± 1.65 (a, A) 168.91 ± 9.69 (a, B) Gulonic acid 0.10 ± 0.02 (a, A) 0.17 ± 0.02 (b, A) 0.27 ± 0.05 (a, B) 0.14 ± 0.02 (b, A) Malic acid 0.56 ± 0.04 (a, A) 0.64 ± 0.06 (b, A) 0.72 ± 0.12 (a, A) 0.56 ± 0.04 (a, A) Phosphoric acid 2.73 ± 0.15 (a, A) 2.64 ± 0.02 (a, A) 3.66 ± 0.60 (a, A) 3.69 ± 0.47 (a, B) Quinic acid 34.02 ± 0.06 (a, A) 40.22 ± 1.98 (b, A) 35.87 ± 4.19 (a, A) 33.41 ± 0.71 (a, B) Shikimic acid 1.24 ± 0.12 (a, A) 1.28 ± 0.03 (a, A) 1.38 ± 0.17 (a, A) 1.52 ± 0.03 (a, B) Amino acids Alanine 0.18 ± 0.03 (a, A) 0.56 ± 0.06 (b, A) 0.21 ± 0.03 (a, A) 0.20 ± 0.028 (a, B) Aspartic acid 0.20 ± 0.03 (a, A) 0.28 ± 0.00 (b, A) ND ND GABA 0.44 ± 0.04 (a, A) 0.61 ± 0.04 (b, A) 0.29 ± 0.03 (a, B) 0.32 ± 0.02 (a, B) Glycine ND ND 0.14 ± 0.04 (a, B) 0.11 ±0.03 (a, B) Serine 0.17 ± 0.02 (a, A) 0.20 ± 0.00 (b, A) 0.19 ±0.02 (a, A) 0.15 ±0.02 (b, B) Others Ethanolamine 0.32 ± 0.02 (a, A) 0.46 ± 0.01 (a, A) 0.28 ± 0.06 (a, A) 0.33 ± 0.06 (a, B) Glucuronic acid ND ND 0.16 ± 0.03 (a, B) ND Hexadecanoic acid 3.66 ± 0.23 (a, A) 3.67 ± 0.24 (a, A) 0.52 ± 0.08 (a, B) 0.910 ± 0.02 (b, B) The values show each metabolite's area relative to the standard (ribitol) area, determined per gram of fresh weight and expressed as the mean and standard deviation of at least three replicates. E-AHC: 'Emerald' under anti-hail cover; E-F: 'Emerald' grown in the field; S-AHC: 'Snowchaser' under anti-hail cover; S-F: 'Snowchaser' grown in the field. Different letters indicate significant differences (p < 0.5) between treatments for the same variety (lowercase letters) or between varieties with the same treatment condition (capital letters). ND: Not detected. It is worth noting that when only the factor of variety was studied under the same cultivation conditions, certain metabolites fluctuated. Raffinose, trehalose, mannitol, GABA, and hexadecanoic acid were significantly more abundant in 'Emerald' than in 'Snowchaser' in field-grown fruit. Cellobiose, ribose, sucrose, xylose, and gulonic acids, on the other hand, were lower in 'Emerald' than in 'Snowchaser'. Most metabolites were much more prevalent in 'Emerald' than in 'Snowchaser' in AHC-grown fruit; and only raffinose, ribose, citric, and phosphoric acids were more abundant in 'Snowchaser'.
To better visualize the biochemical properties of each sample and understand how growth conditions and/or variety influence them, a heatmap with all measured attributes and metabolite content was created (Fig. 2). At first glance, it is evident that the fruit of 'Emerald' grown under AHC undergoes a substantial increase in the level of several compounds and agronomic traits compared to the rest of the samples (Cluster 3: xylitol, mannitol, maltose, aspartic acid, galactose, turanose, titratable acidity, fructose, glucose, myo-inositol, alanine, quinic acid, sucrose, and cellobiose). A few chemicals appear prevalent in the field-grown fruit of 'Snowchaser' in this cluster (glucuronic acid, xylose, gulonic acid, ribose, glycerol, and malic acid). Meanwhile, Clusters 1 and 2 collect the compounds and parameters that distinguish the 'Emerald' and 'Snowchaser' varieties, respectively, regardless of growth conditions. Thus, in 'Snowchaser,' rhamnose, serine, glycine, phosphoric, and shikimic acids, PME, and pectinase activities predominated, but in 'Emerald,' hexadecanoic acid, β-gal activity, soluble solids, TPC, and GABA were more prominent. Surprisingly, five chemicals were not discovered in specific samples, including aspartic acid, mannitol, rhamnose, glycine, and glucuronic acid.
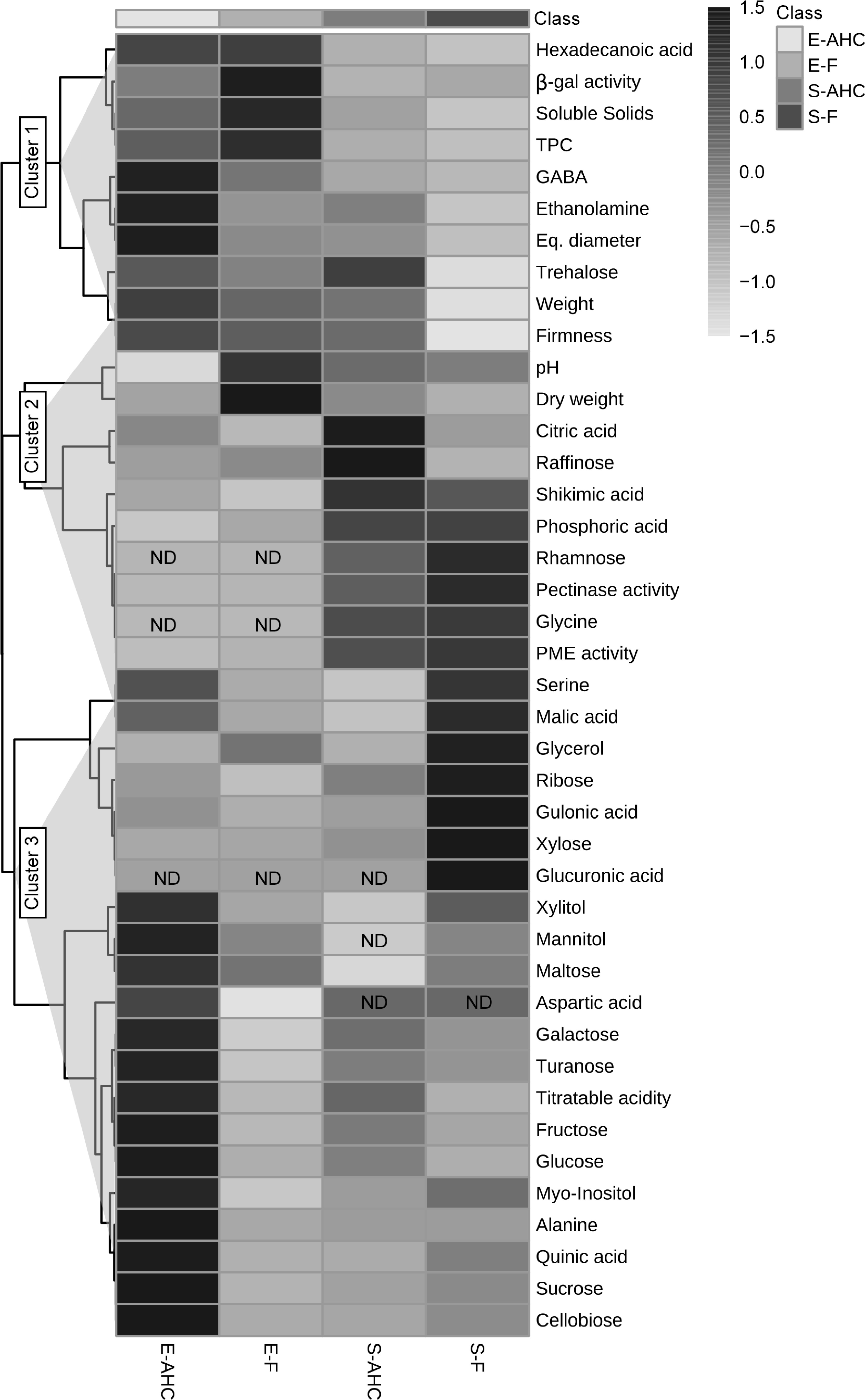
Figure 2.
Heatmap depicting metabolite levels, variations in cell wall-associated enzyme activity, and quality parameters in blueberries cultivated under different conditions. E-AHC: 'Emerald' grown under anti-hail cover; E-F: 'Emerald' grown in the field; S-AHC: 'Snowchaser' grown under anti-hail cover; S-F: 'Snowchaser' grown in the field. Clusters 1, 2, and 3 are further described in the text. Normalized data values are displayed on a grey scale (shown to the right of the figure) and are proportional to the content of each metabolite. Values from three separate determinations were normalized and examined as specified in the Materials and methods. ND indicates chemicals that were not detected in the sample.
A principal component analysis (PCA) was performed to identify the chemicals, enzymes, and/or qualitative features that best define each sample (Fig. 3). From PC1 and PC2, it was possible to describe 40.8% and 26.2% of the variation, respectively. The cumulative explained variance was 67%, indicating that this resource has excellent descriptive ability. PC1 and PC2 assisted in depicting each variety and treatment. As a result, 'Snowchaser' fruits cultivated under AHC had higher levels of citric acid and raffinose metabolites, whereas those grown in the field had higher levels of glucuronic acid, xylose, rhamnose, ribose, and pectinase activity. The metabolites that contributed the most to characterize 'Emerald' fruit cultivated under AHC were glucose, fructose, alanine, mannitol, and cellobiose, whereas dry weight, pH, soluble solids, and β-gal activity did so for fruit grown in the field (Fig. 3b).
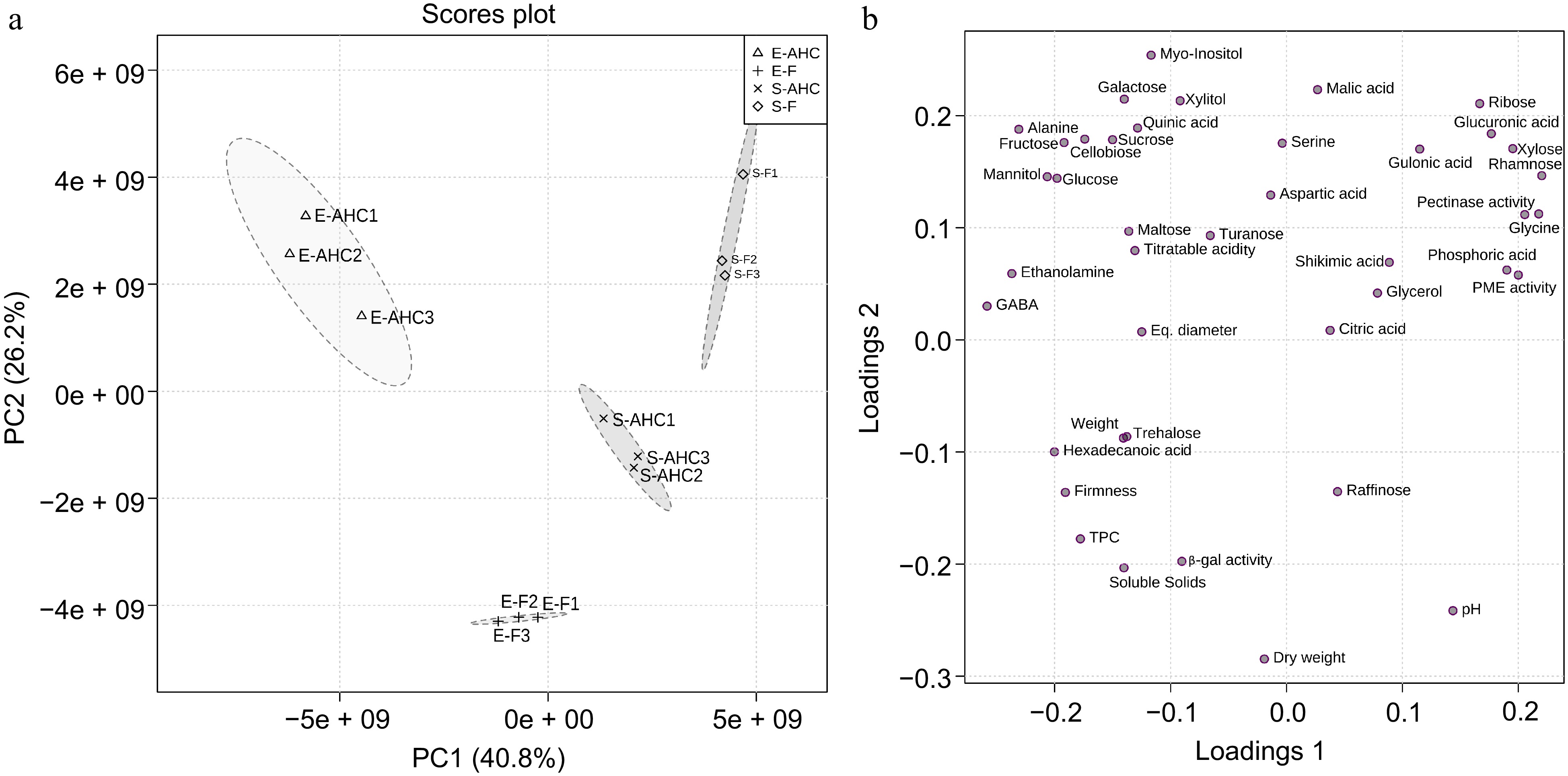
Figure 3.
Principal components analyses (PCA). PCA was modeled on the standardized data matrix for z-score to observe its grouping trend. The data matrix of each group was analyzed by unsupervised PCA. (a) PC1 and PC2 with the (b) related loading plot. Each component's contribution is indicated as a percentage (%). E-AHC: 'Emerald' under anti-hail cover; E-F: 'Emerald' grown in the field; S-AHC: 'Snowchaser' under anti-hail cover; S-F: 'Snowchaser' grown in the field.
A pairwise correlation analysis was performed to learn more about the metabolites or other factors associated with firmness, one of the most essential quality features (Supplementary Fig. S4). Trehalose, ethanolamine, hexadecanoic acid, GABA, weight, TPC, and soluble solids were all substantially and positively linked with firmness. On the other hand, this trait was inversely linked with xylose, glucuronic, gulonic, and phosphoric acids, ribose, rhamnose, glycine, pectinase, and PME activity.
Postharvest conservation
-
To assess the potential impacts of treatments on fruit preservation, various quality parameters were measured after 7 and 30 d of storage at 0.5 °C (Table 3). Only the results from the 2019 season are presented. The decrease in soluble solids (SS) and increase in titratable acidity (TA) for 'Emerald' grown under cover resulted in a significant decline in the SS/TA ratio after 7 d of storage, compared to field-grown fruit. However, after 30 d of conservation, this ratio exceeded ten. Meanwhile, weight loss and firmness were lower in open-field berries than in AHC-treated fruit. When considering the conservation period, the TA, SS/TA ratio, pH, and firmness of the latter samples changed. The storage length, on the other hand, had less impact on 'Snowchaser' than on 'Emerald,' as only firmness rose significantly after 7 and 30 d of conservation in fruit from all treatments. Weight loss was much greater after 7 d of storage for field-produced fruit than after 30 d of storage. Finally, for fruit grown under AHC, 'Emerald' had the greatest weight loss, whereas 'Snowchaser' had the lowest weight loss of all samples examined during storage.
Table 3. Variation in agronomic parameters during postharvest conservation of 'Emerald' and 'Snowchaser' blueberries grown in the field (F) or under anti-hail cover (AHC) throughout the 2019 growing season.
Agronomic parameter E-F E-AHC S-F S-AHC 7 d 30 d 7 d 30 d 7 d 30 d 7 d 30 d SS (°Brix) 12.8 a, A 12.7 a, A 10.8 b, A 10.8 b, A 10.8 a, A 10.5 a, A 10.9 a, A 11, 0 a, A TA (% acid) 0.96 a, A 0.93 a, A 1.37 b, A 1.03 b, B 0.80 a, A 0.75 a, A 0.91 a, A 0.83 a, A SS/TA ratio 13.3 a, A 13.7 a, A 7.9 b, A 10.5 b, B 13.5 a, A 14.0 a, A 12.0 a, A 13.3 a, A pH 2.69 a, A 2.96 a, B 2.64 a, A 2.88 a, B 3.02 a 3.05 2.90 b 3.00 a, A Weight loss (%) 0.48 a, A 0.25 a, A 0.70 b, A 0.73 b, A 0.60 a, A 0.10 b, A 0.10 a, A 0.10 a, A Firmness (N) 2.16 a, A 2.64 a, B 2.62 b, A 2.89 b, B 1.92 a, A 2.08 a, A 2.36 b, A 2.55 b, B AHC: Anti-hail cover growth; F: Field growth; 7 d: 7 days of conservation at 0 °C; 30 d: 30 days of conservation at 0 °C. Different letters indicate significant differences (p < 0.5) between treatments for the same variety (lowercase letters) or between conservation periods with the same treatment condition (capital letters). The quantities in bold indicate significant differences between varieties for the same treatment and conservation time. The values are the average of at least three replicates. For all measures, standard errors were less than 15% and were omitted for simplicity. -
The present study aimed to investigate whether the use of anti-hail covers (AHC) in blueberry cultivation impacts key quality indices, including fruit firmness, sugar content, organic acid content, and phenolic compounds. Measuring the relative quantity of primary metabolites and the activity of two cell wall-related enzymes may provide a more comprehensive understanding of the varieties' behavior. Despite the convenience of AHC for protecting trees and fruit from hailstorm damage, its potentially detrimental impact on fruit quality should not be overlooked.
The use of anti-hail covers resulted in modifications to various agronomic parameters, depending on the season and variety under investigation. The average photosynthetically active radiation (PAR) and diurnal temperature reductions recorded for fruit produced under AHC were lower than those observed for the 'Duke' variety[5]. These measurements were significantly influenced by the color of the cover; white (as utilized in this study) and red covers create less shade and more scattered light than black covers, as demonstrated for kiwifruit[17]. Consequently, white covers were more effective in terms of dry matter buildup.
In the present casse, the effect of growing conditions on quality attributes were more evident in 2018 than in 2019 for 'Snowchaser,' while the opposite occurred for 'Emerald' (Table 1). This difference may be related, in part, to changes in precipitation levels, which were much lower in 2018 than in 2019 during the primary harvest period of 'Snowchaser' (Supplementary Fig. S1). With the exception of soluble solids and ratio SS/TA, which were lower in field-grown fruit (2018), the attributes that changed were always higher for 'Emerald'. Firmness was either higher under AHC or did not alter with treatment (Table 1), consistent with observations in peaches and nectarines[18,19]. As previously stated, the protective cover may reduce photosynthesis, decreasing fruit size. In comparison to field-produced fruit, the equatorial diameter was larger in 'Emerald' and lower in 'Snowchaser' under AHC (only in the 2018 season, Table 1). In no case did the fresh weight of the fruit vary.
Blueberry quality requirements have been established as soluble solids (SS) greater than 10 °Brix, acidity between 0.3% and 1.3%, and an SS/TA ratio between 10 and 33[20]. One notable response is the change in the SS/TA ratio, which was lower under AHC. This ratio was extremely low in 2018 and can be attributed to a generally high amount of titratable acidity, as observed by other scientists[21]. Peaches and nectarines exhibited a similar drop in the SS/TA ratio as open-air-produced fruit[18,19]. Regarding the berry maturation delay (Supplementary Fig. S2), a lag in fruit growth was also observed for the 'Elliot' blueberry variety when tested with white and colored shade covers[6].
The findings indicate that the only change blueberries undergo when grown under protective covers is a decrease in SS/TA ratio, while other agronomic and qualitative aspects remain unaffected. Additional research was conducted to better understand the behavior of the varieties.
The content of total phenolic compounds and the activity of cell wall-associated enzymes changed with season and variety but remained unaffected by growing conditions. Phenolic compounds are highly valued for their health-promoting characteristics, nutritional value, and potential involvement in avoiding significant postharvest deterioration[22]. Hence, the discovery that growth conditions did not affect total phenolic content was a favorable result. Similar outcomes were observed for the 'Elliot' blueberry variety under AHC[6], while phenolic content in kiwifruit was low at harvest time but increased during storage to reach control (field-grown) values[17].
Increased activity of enzymes involved in cell wall metabolism and disassembly is a common occurrence during ripening. While this is important to make the fruit edible and appealing, excessive softness might be undesirable. The primary cell wall is composed of several polysaccharides, including pectins high in α-1,4-linked galacturonic acid (GalA), hemicelluloses, and cellulose[23]. GalA residues can be methylated to varying degrees, controlled by pectin methyl esterase (PME), which removes pectin methyl groups. Unesterified pectins can be cross-linked with Ca2+ ions to form an egg-box structure that supports the wall[24], or they can serve as a substrate for various pectinases or hydrolases that promote wall loosening[25]. The measurement of these activities, which could affect fruit firmness in part, revealed no differences between fruit produced under AHC and in the field. However, significantly altered values were found regarding seasons and varieties, with fluctuations more evident in the 2019 season. 'Snowchaser' exhibited lower TPC and β-gal activity than 'Emerald' but higher PME and pectinase activity. Thus, while the aforementioned observation could explain the lesser firmness of the first variety cultivated in the field, the same explanation does not apply to fruit grown under AHC. To our knowledge, the behavior of cell wall-related enzymes in fruit under covers has never been studied before, even though varietal divergence of these features has already been documented[11,14,26], and based on the current findings, is more important than the influence of AHC.
Differential metabolite content further assisted in defining the effect of growth conditions and variety on the fruit's biochemistry. The assessment of changes in the levels of around thirty metabolites in the fruit from the 2019 season revealed variations influenced by both variety and cultivation conditions (Table 2). Several chemicals were more abundant in 'Emerald' under AHC. Despite the taste of the fruit being influenced by various factors, the higher MI in 'Emerald' under both growth conditions (Table 1, 2019 season), mainly due to changes in citric acid content (Table 2), is expected to result in an improved taste. The same can be anticipated for aspartic acid, which possesses flavor intensifier properties, as glutamic acid does[27,28]. Aspartic acid was not detected in 'Snowchaser,' probably due to its very low level.
The heatmap-defined groups demonstrated that the trend of several variables is intrinsic to a variety and almost independent of the growth situation (Fig. 2, Cluster 1 for 'Emerald' and Cluster 2 for 'Snowchaser'). Meanwhile, parameters connected with the 'Emerald' variety cultivated under AHC (Fig. 2, Cluster 3) are primarily greater than those associated with 'Snowchaser'. When provided in the symplastic compartment, solute buildup, which correlates with a higher soluble solids (SS) concentration in 'Emerald' compared to 'Snowchaser,' is connected with firmness[29,30]. The PCA enhanced this varietal distinctiveness. Some of the characteristics that define the 'Emerald' variety, such as the quantity of GABA, mannitol, firmness, and TPC, are important quality indicators. GABA serves a variety of activities, including cytosolic pH regulation, signaling of biotic and abiotic stressors, and maintenance of a proper C-N ratio[31−33]. It was also detected in greater abundance in the 'Emerald' variety than in the 'O'Neal' variety[14]. Mannitol is a suitable solute synthesized in source tissues and transferred to sink tissues[34]. It protects protein and cellular structures from a significant reduction in osmotic potential and functions as an antioxidant and a reactive oxygen species (ROS) quencher[35,36]. In contrast, 'Snowchaser' has a large contribution of rhamnose, xylose, pectinase, and PME activity. This variety demonstrates enrichment in cell wall lytic activities as well as certain free sugars that comprise pectins and hemicelluloses when cultivated in the field, indicating a more active solubilization of this tissue. Mannitol and glucuronic acid were not detected in 'Snowchaser' fruit under AHC. Given that glucuronic acid is a component of hemicelluloses, it is reasonable to speculate that this polysaccharide is avoiding significant hydrolysis, hence boosting firmness (Table 1). Furthermore, the prevalence of raffinose in 'Snowchaser' under AHC over the other samples is noteworthy because this sugar has been linked to various types of stressors in plants such as salinity, drought, and temperature change[37−39]. Raffinose may operate as an osmoprotectant via functioning as a phloem-mobile signal, scavenging ROS, and membrane stabilization[39,40]. Several substances and characteristics, such as trehalose, ethanolamine, hexadecanoic acid, GABA, or TPC, were shown to be favorably connected with firmness, whereas others, such as ribose, xylose, PME activity, and glucuronic acid, were found to be negatively correlated (Supplementary Fig. S4). Taken together, these changes may be indicating physiological varietal diversification that might be associated with enhanced firmness and possibly better nutritional quality for 'Emerald'. However, the use of AHC improved firmness in 'Snowchaser', and the rise in raffinose concentration could indicate a varietal divergence in stress perception, which requires more investigation.
Postharvest storage affects 'Emerald' blueberries more than 'Snowchaser' blueberries cultivated under AHC
-
To the best of our knowledge, no one has researched the postharvest behavior of blueberries cultivated under AHC versus open field-grown fruit. The conservation period had a greater impact on 'Emerald' fruit cultivated under cover than on 'Snowchaser' fruit. First, the SS/TA ratio (which was lower at harvest and after 7 d of storage than in field-produced fruit) recovered to acceptable levels (> 10) after 30 d of storage at low temperatures (Table 3). Second, after 30 d of storage, firmness rose and was higher in all cases for fruit cultivated under AHC than for berries grown in the open field. This was also observed for 'Snowchaser,' whose fruit had enhanced SS/TA ratio and firmness following storage compared to immediately after harvest (Table 1, 2019 season). Shade, surprisingly, increased calcium uptake by apples and improved their ability to incorporate this ion into the cell wall network[41]. This conclusion was attributed mostly to the fact that shade reduced leaf transpiration, hence promoting fruit transpiration and, as a result, calcium intake through xylem vessels. Although the calcium content of the berries was not evaluated in this investigation, the enhanced firmness found under AHC for both varieties could be explained in part by improved calcium transfer to the berries. Finally, while the measured weight loss varied between samples, it was always less than 1%. These values are similar to those reported for the blueberry variety 'O'Neal'[42].
-
The use of protective white covers in berry production is a useful option for dealing with hail storms. However, some points deserve consideration, as the delay of several days (1−2 weeks) observed in fruit under cover, which could potentially postpone the onset of the commercialization period. Furthermore, varietal and seasonal variances have been noticed in terms of SS/TA ratio, dry matter content, and metabolites. Even when both cultivars have excellent quality, it was found that 'Emerald' preserved some attributes more efficiently than 'Snowchaser,' such as GABA and mannitol levels, high firmness, and cell wall integrity. However, the use of AHC for 'Snowchaser' increased firmness and likely reduced the solubilization of some cell wall components, which was a favorable result. In this last variety, the rise in the amounts of raffinose, a stress-related sugar, could be signaling a divergence with 'Emerald' in terms of stress sensing. Finally, both varieties showed good postharvest behavior, as some relevant quality indices, such as firmness and maturity index, improved after storage without significant weight loss.
This work was supported by a grant from the Agencia Nacional de Promoción Científica y Tecnológica to KEJT (PICT 2016-0091). KEJT is member of the investigator Career from the Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). LM is a Doctoral Fellow of CONICET. Agustín Gollán was a Doctoral Fellow of CONICET/INTA. Fernando Bello, María F. Rivadeneira and Daniel Vázquez are members of the Instituto Nacional de Tecnología Agropecuaria (INTA). The authors want to acknowledge the collaboration of Bioq. Mónica Hourcade for GC-MS runnings.
-
The authors confirm contribution to the paper as follows: study conceptualization, manuscritp writing, review and editing, visualization, supervision, project administration, funding acquisition: Tripodi KEJ; methodology, investigation, statistical software analyses: Morales L, Gollán A; biochemistry assays: Morales L; quality parameter determinations, field fruit treatments and determinations: Gollán A; plant treatments and conditioning, supervision of plant phenology and growth, collection of samples: Rivadeneira MF, Bello F; validation, supervision of plant treatments and field determinations: Vázquez D. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Fig. S1 Precipitation levels and temperature fluctuation during the period of growth and harvest of each variety in the 2018 and 2019 years. Arrows signal the month of harvest for mature fruit of 'Snowchaser' (S) and 'Emerald' (E).
- Supplementary Fig. S2 Bloom curve in 'Emerald' and 'Snowchaser' southern highbush blueberry at different growth conditions and for 2018/2019 seasons. E-AHC: 'Emerald' under anti hail cover; E-F: 'Emerald' grown in the field; S-AHC: 'Snowchaser' under anti-hail cover; S-F: 'Snowchaser' grown in the field. In both graphics, the abscissa shows the date of phenological stage determination. Number code for phenological stages was adapted from the alphanumeric scale (Spiers, 1978): 1-visible swelling of bud; 2: bud scales separating; 3-individual pink corollas visible; 4: white corolla expanded; 5: corolla completely opened; 6: corolla drop; 7: initial small green fruit; 8: growth of fruit, 9: fruit change colour for green to pink (Rivadeneira, 2008).
- Supplementary Fig. S3 Example of one simultaneous assay of PME and pectinase activity. The inner clear halo, delimited by a black line, corresponds to the pectinase activity, while the external darkest halo is due to the PME activity (Lionetti, 2015).
- Supplementary Fig. S4 Correlation graphs. (a) Correlation between firmness and measured parameters/compounds. Correlation coefficients for the top 25 parameters/compounds most correlated with the firmness are shown as bars, indicating positive (right) and negative (left) correlation. Parameters demonstrating a statistically significant correlation are denoted with an asterisk (p-value<0.05). (b) Correlation Heat map. The correlation coefficients (Pearson) among metabolites, internal quality parameters, total phenolic content (TPC), and enzymatic activities across all samples are displayed. A correlation cut-off of 0.65 was applied; therefore, pair-wise correlations failing to meet this threshold were set to 0.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Morales L, Gollán A, Bello F, Rivadeneira MF, Vázquez D, et al. 2025. Anti-hail covers causes changes in the quality and biochemistry in two blueberry (Vaccinium corymbosum L.) cultivars. Technology in Horticulture 5: e003 doi: 10.48130/tihort-0024-0031 

Anti-hail covers causes changes in the quality and biochemistry in two blueberry (Vaccinium corymbosum L.) cultivars
- Received: 20 June 2024
- Revised: 07 November 2024
- Accepted: 18 November 2024
- Published online: 23 January 2025
Abstract: Given the increased frequency of hailstorms that pose a threat to blueberry fruit and bushes, the utilization of anti-hail covers has become prevalent in Argentina's North-Eastern region. However, this practice may alter the orchard microclimate, potentially affecting fruit quality. The impact of these covers on the biochemical composition and agronomic attributes of two extensively cultivated blueberry cultivars, 'Emerald' and 'Snowchaser' was investigated. The early phenological stages exhibited a delay of several days in fruit under cover, which could potentially postpone the onset of the commercialization period. Seasonal and varietal variations in phenolic compound levels and cell wall-associated enzyme activity were observed, although these differences were not attributable to the growing conditions. The 'Emerald' cultivar demonstrated an overall positive response to the net application, with measured traits either remaining unaltered or showing improvement in comparison to field growth. The 'Snowchaser' cultivar displayed enhanced firmness when grown under cover, which is a favorable outcome. In this variety, the increase in the levels of raffinose, a stress-related sugar, may indicate a divergence from 'Emerald' with regard to stress sensing. Both varieties demonstrated resistance to decay in the postharvest period, with an increased SS/TA ratio and firmness resulting from the use of covers. Despite some varietal differences, this study supports the efficacy of anti-hail covers in safeguarding blueberries without compromising overall crop health and fruit quality. This outcome is relevant since it validates this practice for two blueberry varieties of commercial importance, thereby reducing the potential loss of fruit and bush damage.
-
Key words:
- Blueberries /
- Biochemistry /
- Anti-hail cover /
- Fruit quality /
- Metabolomic