






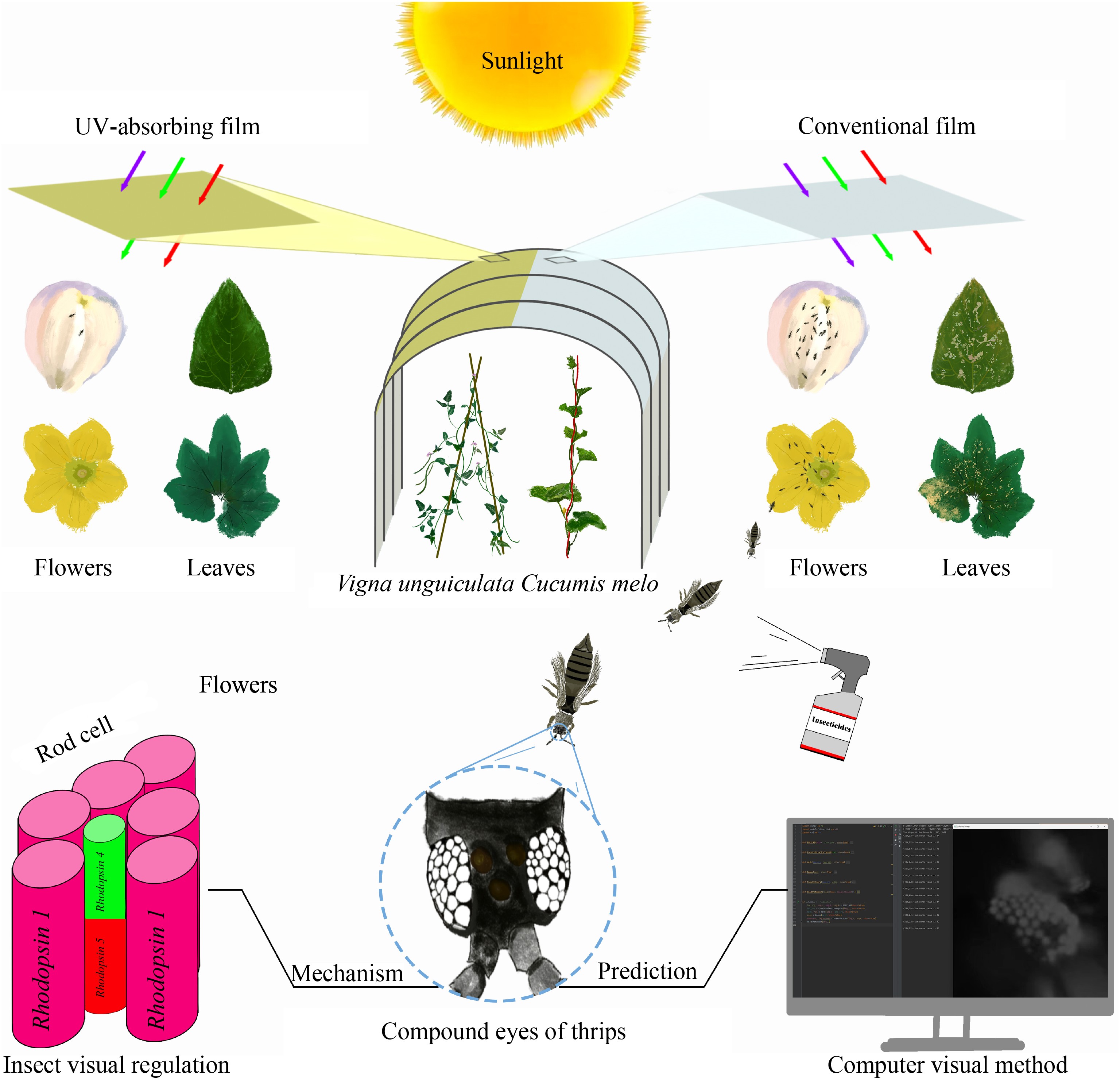



-
In China, cowpea cultivation has hundreds of years of history, cultivation area is large, mainly planted in temperate and tropical areas. Thrips are widely distributed in the world as 'small bugs and great hazards'[1,2]. Megalurothrips usitatus and Thrips palmi are two important pest thrips in agriculture. M. usitatus are mainly distributed in tropical areas and harms legumes, while T. palmi has a wide distribution and can harm many fruits and vegetables. They damage plants by filing and sucking young parts of plants (including shoots, tender leaves, and fresh fruits)[3,4] and cause a significant deterioration in fruit quality, leading to a substantial decrease in marketability[5].
There exist numerous management measures aimed at regulating the population of thrips[6,7]. Unfortunately, the use of chemical agents is still the most used measure, due to the thrips' traits of short generations, parthenogenesis, and high reproduction rate[8]. There are more than 30 types of chemical agents on the market, of which neonicotinoids and pyrethroids account for 24% and 15% sales, respectively[9]. The use of neonicotinoid such as imidacloprid (Gaucho) and thiacloprid (Cruiser) to treate seeds is the most widely used measure at-planting for thrips control[10]. At the same time, foliar applications of insecticides such as pyrethroids may be needed for optimal plant protection[10].
The irrational use of chemical agents has resulted in great resistance to chemical agents in the field thrips population[11,12]. Currently, M. usitatus and T. palmi have developed high levels of resistance to lambda-cyhalothrin, deltamethrin, and spinetoram in recent years[1,13−15]. These insecticides have almost no effect on the thrips. Nerve insensitivity associated with mutations in sodium channels is known as a major resistance mechanism to pyrethroid[16], DDT and indoxacarb[17], while neonicotinoids and spinosyns act on nicotinic acetylcholine receptors (nAChRs)[18,19]. In addition, transcript truncation of neuroreceptors is also one of the mechanisms by which thrips resistance to insecticides[20,21].
Consequently, an effective technique is urgently needed to control thrips. The UV-absorbing films, which influence insect visual responses and behaviors by controlling the UV luminous flux, is recognized as an effective strategy in Integrated Pest Management (IPM) systems[22]. So far, the UV-absorbing films have been proven to be effective in controlling aphids[23,24] and thrips (Frankliniella occidentalis Pergande, T. palmi)[25,26]. In contrast to other pest management methods, the manipulation of insect visual response is the specialization toward the visual ecology of insect pests. This has greatly improved in recent years because of the availability of LEDs and color glue trap in controlling the light intensity and specific wavelengths on thrips behavior, and the adaptation of simulating relevant lighting environments in field[27].
Thrips' light specificity and sensitivity as well as intriguing regionalization of structural adaptations in their eyes mediates the visually guided behavior to feeding habitats, mating, egg laying and orientation towards host plants[28]. Previous experiments have shown that thrips have wavelength-specific behavior and true color vision. The light preference varies between populations, species, and experimental conditions[3]. It has been demonstrated that thrips’ sensitivity curve is produced by three visual pigments peaking in the green, blue, and UV[29,30]. This corresponds to short-wavelength-sensitivity (SWS), blue clade (SWS-B) and UV clade (SWS-UV) and long-wavelength-sensitive clade (LWS) of opsins, and is conserved in many trichromatic insect vision systems[31]. The green peak varies slightly between species and studies: 535 nm[32] or 500 nm[30] for Frankliniella occidentalis and the L1 biotype of Thrips tabaci[29], and 520 nm for Scirtothrips dorsalis[33]. Interestingly, the L2 biotype of T. tabaci lacks the UV peak and displays a novel sensitivity curve[34].
In this study, to develop a better control measure, the resistance of two thrips species were monitored. An ultraviolet-absorbing film (UVa-F) was developed for the control and growth evaluation of highly resistant thrips in the greenhouse. The mechanism of UVa-F effect on the thrips was further studied. The compound eyes of M. usitatus were carefully observed and analyzed for the light-sensitive regions under UVa-F treatment by scanning electron microscope and fluorescence microscope. A computer vision image processing program was designed to assist in the analysis of the light-sensitive regions of thrips. Then five opsin genes belonging to G-protein-coupled receptor were identified in the transcriptome of M. usitatus, and their expression in M. usitatus compound eyes was accurately evaluated under different light conditions.
-
M. usitatus and T. palmiwas were collected in Haikou and Sanya, Hainan province (China) from 2021 to 2023 (Supplemental Table S1). These field populations were taken to the laboratory for bioassay and visual studies. They were raised in quartz glass tubes in incubators with 25 ± 5 °C, 65%−75% relative humidity and 12-h of light/dark photoperiod. The fresh cowpea segments were changed every 2 d, and the cowpea with laid eggs were put into a new quartz tube and marked with the algebra and date.
Bioassay
-
Seven insecticides (lambda-cyhalothrin, deltamethrin, imidacloprid, thiamethoxam, dinotefuran, spinetoram, and emamectin-benzoate) used in the bioassay were all purchased from Shanghai Yuanye Bio-Technology Co., Ltd (China). The bioassay was carried out using the TIBS (thrips insecticide bioassay system)[13,15] as follows: First, pre-tests were conducted to determine the appropriate concentration range of each insecticide. Second, seven concentrations of each insecticide were set up with three replicates per concentration and 0.1% Triton X-100 (Sigma, Shanghai) in distilled water as a control. Third, the insecticides were added in 1.5 mL centrifuge tubes for 4 h, and the 1.5 cm Cowpea segments were immersed for 10 s and placed at room temperature for drying. Fourthly, the tube caps were punched, and 10−12 thrips were aspirated and placed in the tubes. Cowpea and gauze were placed at the mouth of the tube and the lid was tightly closed. Finally, tubes were placed at room temperature and the total number of insects and the number of deaths were recorded after 48 h.
Detection of neuroreceptor mutations
-
RNA from 30 adults of M. usitatus and T. palmi was extracted separately with TRIzol (Invitrogen, USA), and the RNA was reverse-transcribed into cDNA using the PrimeScript™ II 1st Strand cDNA Synthesis Kit instructions (Takara, Dalian). PCR amplification of the sodium channels and nAChR subunits were carried out using the primers listed in Supplemental Table S2, and the PCR products were sent directly out for sequencing (Nanshan, Haikou).
Investigation of thrips in different greenhouses
-
The geographical location and setting of the greenhouse were described previously[26]. Treatment groups were covered with independent R&D UV-absorbing film (UVa-F), while control groups were covered with conventional films (C-F, XiTian Plastic Factory, Shandong, China). UVa-F is independently developed and produced by our laboratory, and the specific spectral parameters and physical properties have been referred to in a previously published article[26]. The transmittance of UVa-F in the ultraviolet spectrum (≤ 400 nm) is almost zero, and the transmittance of C-F in the ultraviolet spectrum is 48.86%. Cowpea (Vigna unguiculata) and Hami melon (Cucumis melo) were selected as test crops in 2023. Different greenhouses adopt the same cultivation, water, and fertilizer management regime.
The occurrence of thrips in the greenhouse was evaluated by investigating the number of thrips on the flowers and blue sticky traps (double-sided, size: 20 cm × 30 cm) with five-point sampling method. Three samples were collected at each point, repeated every 3 d in three greenhouses with each of two films, and a total of 15 and 10 continuous surveys of cowpea and Hami melon were conducted, respectively.
Prevention and control rate (P):
$ P=\dfrac{{N}_{C-FG}-{N}_{UVa-FG}}{{N}_{C-FG}}\times 100{\text{%}} $ (1) The number of thrips (N) is the average value of thrips on the flower plus on the blue sticky traps in each kind of greenhouse (C-FG and UVa-FG).
Investigation of quality and yield of Cowpea and Hami melon in different greenhouse
-
Cowpea and Hami melon fruits were randomly collected from greenhouses with UVa-F and C-F using the five-point sampling method. We collected three samples at each sampling point. At every sampling point in all greenhouses, the median of the three samples was selected as the value of the sample point in the greenhouse which was repeated three time during the harvest period. The collected fruits were used for quality measurements, including cowpea fruit weight and fruit length, and Hami melon fruit longitudinal diameter, transverse diameter, and perimeter.
Yield statistics from the beginning of the harvest period to the end of the harvesting period to obtain the total production of each greenhouse.
Yield increase rate (Y):
$ Y=\dfrac{{N}_{UVa-FG}-{N}_{C-FG}}{{N}_{C-FG}}\times 100{\text{%}}$ (2) Where, N is the number of total crop production from three repeated samples in each greenhouse.
Observation of ocelli and compound eyes structure by scanning electron microscope (SEM)
-
Ten M. usitatus adults were placed directly into a pre-cooled 2.5% glutaraldehyde solution and fixed for 24 h at 4 °C in the dark. The fixed sample was rinsed with 0.1 mol/L PBS (pH 7.2) six times. Each thrips samples were then cleaned using an ozone cleaner (ZoneSEM, Hitachi High-Technologies, Rexdale, Canada) and dehydrated with a gradient alcohol solution (10%, 30%, 50%, 70%, 80%, 90%, and 100%) followed by carbon dioxide critical point redrying. Next, the samples were stuck with double-sided conductive tape and sprayed with Au in a vacuum spraying machine. Finally, the samples were observed and photographed under a JEOL scanning electron microscope (Japan).
Observation of compound eyes in thrips under fluorescence microscope
-
The M. usitatus were gently placed on ice to freeze. Then frozen thrips were placed on a microscope slide for fluorescence observation with OLYMPUS BX53 biomicroscopy (Shinjuku, Tokyo, Japan) under fluorescence excitation wavelength UNA (360−370 nm), BVW (400−440 nm), BNA (470−495 nm), GW (530−550 nm). All pictures were analyzed using cellSens Dimension software.
Computer visual inspection and aided analysis
-
OpenCV (Open-Source Computer Vision Library) was used to convert the RGB three-channel image taken with OLYMPUS BX53 biomicroscopy into the LAB channel image, extract the luminance channel (L channel) in the LAB-channel image, and convert it into a single-channel grayscale image displaying luminance difference automatically. First, noise reduction was performed on the L-channel image, including tophat operation. Then binary operation was used on the processed image to form a mask. In the mask, the canny algorithm was used to obtain clear contour information, and the minimum enclosing rectangle was drawn on the original image according to the contour position. At the same time, the luminance value which was the fluorescent region in the original image was compared with that of the UV-sensitive ommatidium in the original image. The luminance region information was obtained by manually marking the UV-sensitive ommatidium position. Finally, the luminance value of the automatically obtained fluorescence region by OpenCV and that of the manually marked UV-sensitive region was analyzed. The above image processing methods are written in Python, the Python IDE is PyCharm 2022.2.4 (
www.jetbrains.com/community/opensource/#support ), and the OpenCV vision is 3.4.1 (https://opencv.org/license ). The original code and the program running video are shown in Supplemental File S1 and Supplemental Video S1.Phylogenetic analysis
-
Five opsin-related genes came from the transcriptome (Supplemental File S2). The opsin sequences of 10 insects except for M. usitatus were downloaded from NCBI and aligned (Supplemental File S2). DNAMAN 7.0.2.176 software (USA) was used to predict gene CDS. Subsequently, MEGA 7.0 (Mega Limited, Auckland, New Zealand) was used to construct the molecular phylogenetic tree by Neighbor-Joining Tree (NG). iTOL was used to evaluate secondary self-development (
https://itol.embl.de/itol_account.cgi# ).Real-time quantitative PCR (RT-qPCR)
-
The thrips raised in the laboratory were treated in darkness for 8 h, then exposed to LED light (10 W at 400−760 nm) and UV light (10 W at 315−400 nm) for 3, 6, and 9 h, respectively. The thrips exposed to dark for 3, 6, and 9 h served as controls. After rapid freezing in liquid nitrogen, all samples were stored at −80 °C. The LED light and UV light both were purchased from Xuzhou Aijia Electronic Technology Co., Ltd (China). The adult thrips' heads are then collected to extract the RNA. Total RNA extraction by Trizol reagent (Invitrogen, USA) following the manufacturer’s protocol. The concentration of RNA was determined by Micro Drop (BIO-DL Corporation, Shanghai, China), and its integrity were detected with 1% agarose gel electrophoresis. RNA with an A260/280 ratio range of 1.8–2.1 and a 260/230 ratio > 2.0 was used for the synthesis of cDNA. By using the PrimeScript RT Reagent Kit with gDNA Eraser (Takara, Dalian, China) Kit, cDNA was synthesized from total RNA using the kit’s instructions and stored at −20 °C until use.
Standard curves and RT-qPCR were carried out in a 20 μL reaction system using AriaMx Real-Time PCR System equipment (Agilent, USA). The program operation is carried out according to the instructions of ChamQ Universal SYBR qPCR Master Mix (Vazyme, China), which included 2 μL 81-fold diluted cDNA template and 10 μmol primers. The reliability of the qRT-PCR results was confirmed by standard curve and melting curve analysis. Standard curves were created by using a 3-fold dilution series of cDNA as a template for each treatment using the linear regression model[35]. The efficiencies (E) of corresponding primers used in qRT-PCR were calculated using Eqn (3).
$ E=\left({3}^{\frac{-1}{slope}}-1\right)\times 100{\text{%}} $ (3) The relative expression of the target gene was calculated according to the 2−ΔCᴛ (Ct: cyclic threshold) method[36], using the actin (ACT) as the internal reference gene[37]. The primers for RT-qPCR are listed in Supplemental Table S3.
Statistical analysis
-
Bioassay results for each field population of M. usitatus and T. palmi were calculated by POLO-Plus version 2.0. Sequence comparison was performed using DNASTAR Lasergene V7.1 software (Madison, USA) to count the mutation sites on the sodium channel and nAChR subunit genes of both thrips separately. Significant differences between different sets of data were analyzed using an unpaired t-test. Graphs were plotted by GraphPad Prism 9 (GraphPad, San Diego, USA), Biorender (Canada), RStudio 2023.06.1 (USA), R version 4.3.1 (New Zealand) and Adobe Photoshop 24.0.0 (USA).
-
We conducted insecticide surveys on the field populations of M. usitatus and T. palmi in Haikou and Sanya regions (China) between 2021 and 2023 to evaluate their resistance to pyrethroids (lambda-cyhalothrin, deltamethrin), neonicotinoids (imidacloprid, thiamethoxam, dinotefuran), spinetoram, and emamectin-benzoate (Fig. 1).
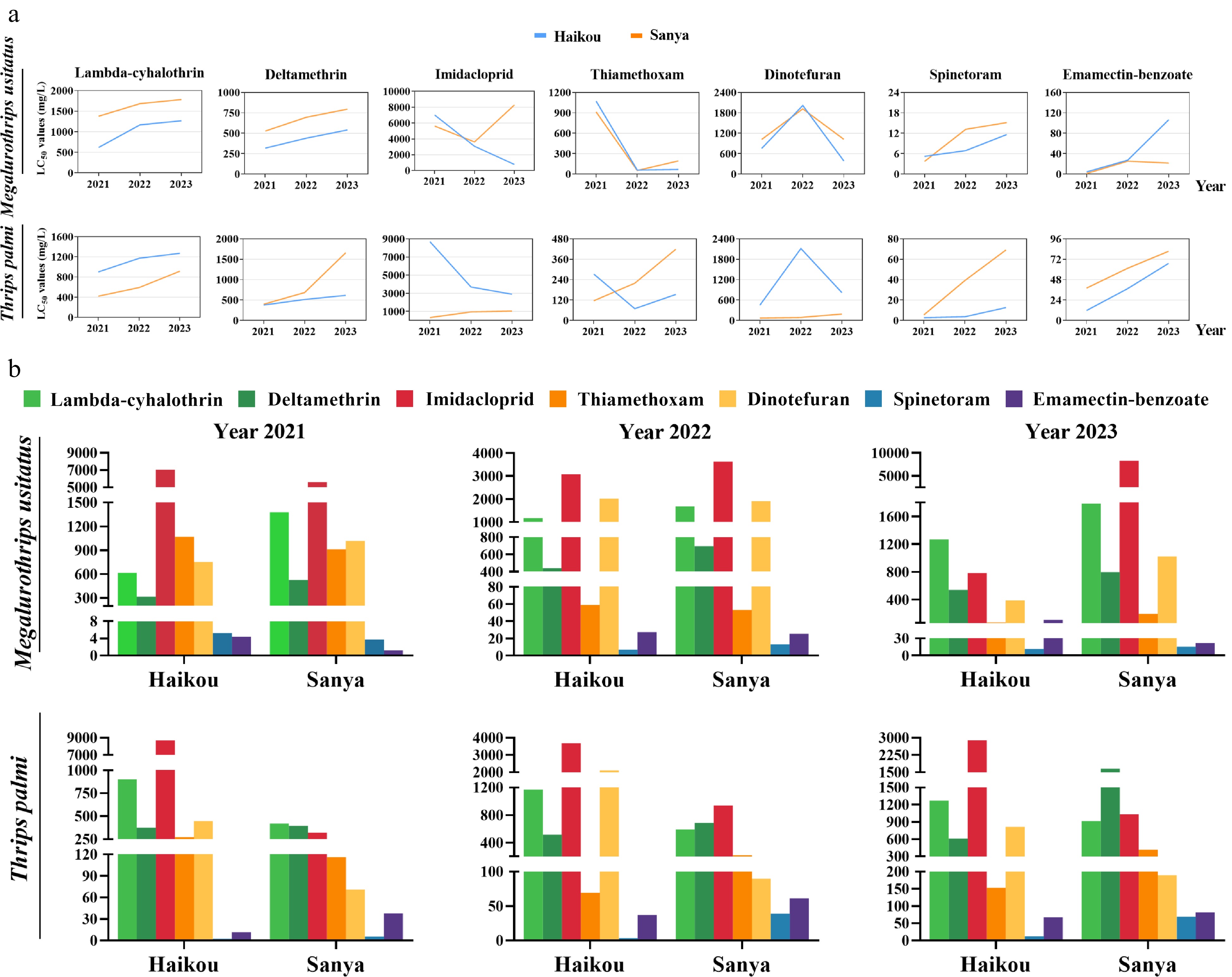
Figure 1.
Insecticide resistance of M. usitatus and T. palmi. (a) The resistance levels of two field populations to each insecticide from 2021 to 2023. (b) The annual resistance levels of the thrip populations to seven insecticides in the Haikou and Sanya regions (China).
Over the last three years, the resistance levels of two thrips to pyrethroids have increased. T. palmi resistance to deltamethrin in the Sanya region was significantly higher in 2023 than in 2022. The annual increase of LC50 has doubled from 72% between 2021 and 2022 to 147% between 2022 and 2023 (Supplemental Table S4).
The resistance of two thrips to neonicotinoids in the Haikou region declined back to relatively low levels. On the contrary, the resistance in the Sanya region kept a rapid upward trend after a decline (Fig. 1a). There was a very modest decline in the resistance of M. usitatus to emamectin-benzoate in the Sanya region from 2022 to 2023. The resistance to spinetoram and emamectin-benzoate in both regions maintained a rising trend (Fig. 1b).
Sodium channel and nAChRs mutation in M. usitatus and T. palmi
-
One of the resistance mechanisms to insecticides are mutations in insect neuroreceptors. Ten mutations (V218I, A221V, V225I, M292R, G969C, T995L, D996N, M1020I, V1391I, and K1638E) were detected in the sodium channel of the field M. usitatus population, with a 100% frequency of the V1391I (Supplemental Table S4). More mutations were detected in T. palmi sodium channel, about 15 mutations (S58T, C167G, E182D, A457V, P502H, R680Q, P726L, T772P, I843M, A850V, G939R, S1050Y, M1420T, R1730W, and M1943I), of which the I843M frequency was 93.33% (Supplemental Table S4). The length of the open reading frame (ORF) of the sodium channel was 6,300 bp in M. usitatus and 6,288 bp in T. palmi (Supplemental File S3).
Five mutations in the nicotinic acetylcholine receptor (nAChRs) were detected on each of α2 subunits (G213C, R241T, R406Q, S441N, and L535R) and α8 subunits (V19A, L26I, M87T, W227C, and D231H) of M. usitatus, while only three mutations were found on the β1 subunit (M3I, G269D, and F309I), and the lowest mutation frequencies were two out of 30 mutations, with a high rate of 33.3% for V19A. Five mutations were found on α1 subunit of T. palmi nAChRs, of which N118S was a homozygous mutation (Supplemental Table S4). The ORF lengths of M. usitatus α2, α8, and β1 subunits were 1,656, 1,671, and 1,638 bp, respectively, while T. palmi α1 subunit was 1,662 bp (Supplemental File S3 and Supplemental Fig. S1).
Effects of optical films on pest occurrence and plant yield and quality
-
The population occurrence of thrips in different greenhouses protected by using different films was then investigated (Fig. 2a, c, f & i). The number of thrips on the flowers and blue sticky traps in different greenhouses were significantly different (Fig. 2e, i). The average number of M. usitatus in the greenhouses covered by conventional film (C-FG) and by UV-absorbing film (UVa-FG) was 813.24 and 27.05 (t = 12.67, df = 14, p < 0.0001), while the average number of T. palmi in C-FG and UVa-FG was 514.30 and 11.89 (t = 2.59, df = 9, p = 0.0293). The prevention and control rates by UVa-FG were 96.67% to M. usitatus and 97.69% to T. palmi (Fig. 2e).
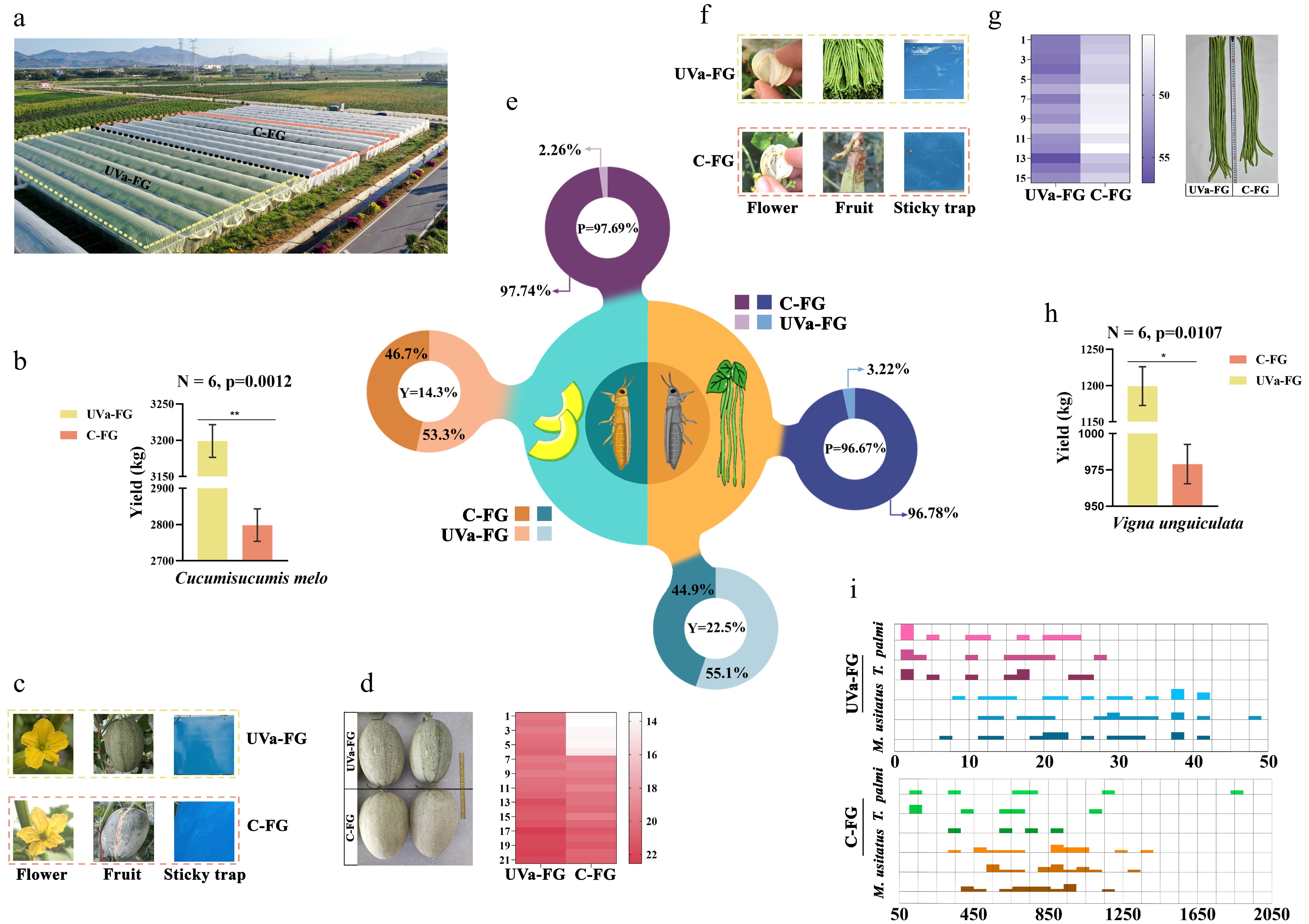
Figure 2.
Effect of UV-absorbing film greenhouse on thrips control, crop quality and yield of cowpea and Hami melon. (a) Greenhouses covered with two different light-transmitting films, conventional film greenhouses (C-FG) and UV-absorbing film greenhouses (UVa-FG), respectively. (b) Yield of Hami melon in two greenhouses (UVa-FG and C-FG). (c) Comparison of number of T. palmi on flowers and sticky trap and damage to Hami melon fruits of UVa-FG and C-FG. (d) Effect on the size of Hami melon. Left panel shows the size of the Hami melon in UVa-FG and C-FG. Right panel shows a heat map of the geometric mean of the longitudinal diameter, transverse diameter and perimeter of the Hami melon. (e) Prevention and control rate (P) and yield increase rate (Y) in UVa-FG and C-FG. (f) Comparison of the numbers of M. usitatus on flowers and sticky traps and damage to cowpea fruits of UVa-FG and C-FG. (g) Effect on the size of cowpeas. Left panel shows the size of the cowpea in UVa-FG and C-FG. Right panel shows a heat map of the geometric mean of the weight and length of the cowpea. (h) Yield of cowpea in UVa-FG and C-FG. (i) The histogram showed the occurrence of two species of thrips in UVa-FG and C-FG. The height of the column shows the density of the distribution under this number. The width of the column is shown in the number distribution range (triple repetition).
There were clear differences in the appearance of cowpeas and Hami melon grown in different greenhouses (Fig. 2d, g). The single fruit weight and length of cowpeas under UVa-FG were higher than those under C-FG (t1 = 7.064, df = 28, p < 0.0001; t2 = 5.221, df = 28, p < 0.0001) (Fig. 2g). The external quality (longitudinal diameter, transverse diameter, and perimeter) of UVa-FG Hami melons was higher than that of C-FG melons (t1 = 4.896, df = 28, p < 0.0001; t2 = 0.342, df = 28, p = 0.0256; t3 = 2.167, df = 28, p =0.0389) (Fig. 2d).
The yield of cowpeas and Hami melon in UVa-FG was significantly higher than that in C-FG (Fig. 2b, e & h). The yields of cowpea and Hami melon were 979.00 kg and 2798.33 kg (Fig. 2b, h), respectively in C-FG. The average yields of cowpea and Hami melon in UVa-FG were 1199.33 kg and 3199.00 kg, respectively, an increase of 22.5% and 14.3% by UVa-FG for cowpea and Hami melon respectively (Fig. 2e).
The response of compound eyes of M. usitatus to different wavelengths
-
The possible mechanisms of crop protection by UVa-FG were further investigated by the structural observation of compound eyes of M. usitatus and the measurement of their response to different wavelengths as well as computer vision simulation. The three dorsal ocelli of M. usitatus are triangular in distribution on the upper side of the compound eye (Fig. 3aII). The dorsal ocellar is flanked by a pair of ocellar bristles (Fig. 3aIII). The number of ommatidium in female and male adults is 65−75 in each eye. The ommatidia are oval, smooth surface, convex, and arranged closely in the edge region and loosely in the central region (Fig. 3aI, II & III). There are irregularly distributed trichoid sensilla around ommatidium, and each trichoid sensillum is between 11.07 and 13.24 μm in length (Fig. 3aIII). The dorsal rim area (DRA) of the compound eye is the area where polarized light is perceived to navigate (Fig. 3aII).
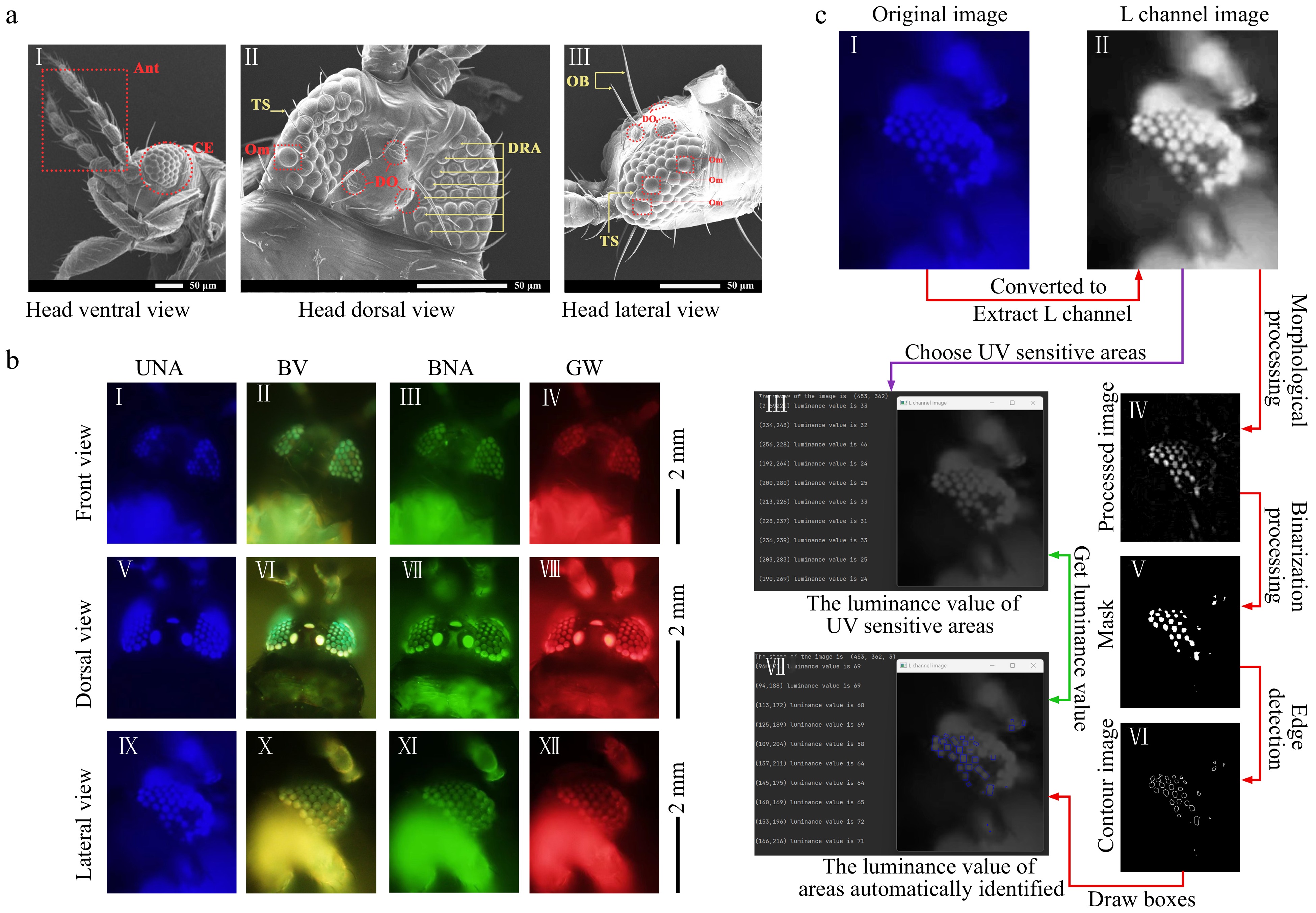
Figure 3.
External morphology, ultrastructure and the response of compound eyes of Megalurothrips usitatus under different wavelengths. (a) Microscopic observation of visual receptors of Megalurothrips usitatus. (aI) Ventral view of head; Ant: Antenna; CE: Compound eye. (aII) Dorsal view of head; TS: Trichoid sensillum; Om: Ommatidium; DO: Dorsal ocellus; DRA: Dorsal rim area. (aIII) Lateral view of head; OB: ocellar bristles; DO: Dorsal ocellus; Om: Ommatidium; TS: Trichoid sensillum. (b) Fluorescent area of Megalurothrips usitatus compound eyes irradiated by UNA (360−370 nm), BVW (400−440 nm), BNA (470−495 nm) and GW (530−550 nm) in the front, dorsal and lateral view. The triangles (▲) mark the position of the UV-sensitive ommatidium. (c) The images of computer vision aided analysis by OpenCV. (cI) The original image which shows the fluorescent area of Megalurothrips usitatus compound eyes irradiated by UNA in the lateral view. (cII) A grayscale image of the original luminance channel. (cIII) The image with clear fluorescent responsive areas morphologically processed of the L channel image. (cIV) The binary image binarized from Fig. 3c-III. (cV) The contours of the mask (cVI) Left panel shows the luminance values for ten points located in the UV-sensitive ommatidium areas and the position coordinates of the corresponding points, and right panel is the corresponding image. (cVII) Left panel shows the luminance values for ten points located in the clear fluorescent responsive areas chosen by OpenCV with box and the position coordinates of the corresponding points, and right panel is the corresponding image with blue boxes.
Six ommatidia of M. usitatus in the lateral view and seven ommatidia in the front view did not respond to the fluorescent light at between 360 nm and 370 nm wavelength, while other ommatidia around these ommatidia emitted bright blue-purple fluorescence (Fig. 3bI, V & IX). The absence of a bright fluorescence in responding to ultraviolet lights indicates that there is no UV-shielding pigment in these ommatidia. As the wavelength increased, these ommatidia became brighter and approached the luminance of the surrounding ommatidia`s color gradually (Fig. 3bI, II, III & IV). It should be noted that the fluorescence of the ommatidium in the marginal region on the back was stronger than that of others (Fig. 3bVI, VII, VIII). Under the irradiation of four bands of light, three dorsal ocellus showed bright fluorescence, reflecting the corresponding colors of four bands (Fig. 3bV, VI, VII & VIII). It was observed from both the UNA-lateral view images and the images of the luminance channel (L channel) that some ommatidia showed high brightness, while a few ommatidia showed low brightness in contrast (Fig. 3cI, II). OpenCV was used, a method of computer vision inspection, to extract from the fluorescence images the fluorescence-responsive regions in thrip compound eyes and to quantify the brightness of the responses (Supplemental File S1 & Supplemental Video S1). The highlighted areas selected by OpenCV in the blue box mostly overlapped with the areas where the ommatidia showed high brightness (Fig. 3cI & II). This means that the luminance value of these highlighted areas in the blue box can be considered as a luminance value of the ommatidium with fluorescence response (Fig. 3cIII, IV & V). The results presented in Fig. 3b demonstrated the consistencey between them. This allows us to use the luminance value of UV-sensitive ommatidium areas given by OpenCV to quantify the brightness of the manually observed UV-sensitive ommatidium areas (Fig. 3cVI, VII).
Analysis of expression level of vision-related genes in different light environments
-
At the molecular level, five vision-related genes were retrieved from the sequenced transcriptome (Genbank: SRP453564) of M. usitatus (Supplemental File S2), and 93 protein sequences of other insects were downloaded from NCBI (Supplemental File S2). Phylogenetic trees showed Rhodopsin 1 of M. usitatus had 100% similarity with Rhodopsin of F. occidentalis. Rhodopsin 2 and 3 of M. usitatus were highly homologous with Rhodopsin of other insects (Fig. 4a). Additionally, Ultraviolet sensitive opsin 2 of M. usitatus had 100% similarity with Ultraviolet sensitive opsin 1 of T. palmi (Fig. 4a). Homology analysis revealed that ultraviolet sensitive opsin 1 and 2 of M. usitatus were highly similar to ultraviolet sensitive opsin of A. mellifera and D. melanogaster which were sensitive to short wavelength (< 400 nm). While Rhodopsin 1 and 2 of M. usitatus were sensitive to long wavelength (Fig. 4a). According to Fan et al.[38], the peak diagram of the wavelength sensitivity curve was drawn (Fig. 4b). Thrips have three wavelength sensitivity peaks in the UV region of 375 nm, the blue region of 440 nm, and the green region of 525−550 nm (Fig. 4b).
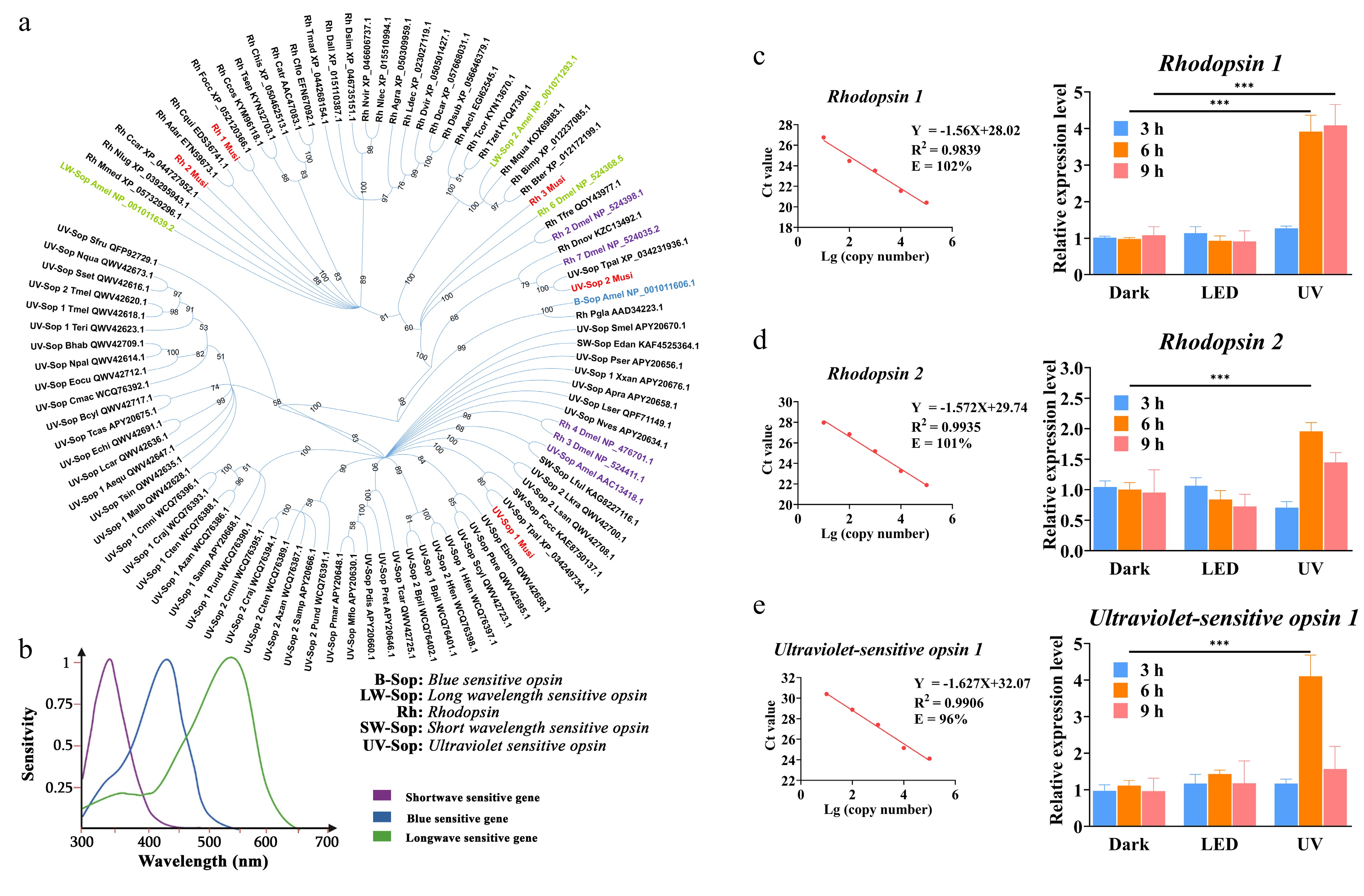
Figure 4.
Homology and expression analysis of opsin genes of Megalurothrips usitatus and their sensitivity to light range. (a) Homology analysis of five vision-related proteins of Megalurothrips usitatus to other vision-related proteins of photosensitive insects which have been reported. Purple font: short wavelength (< 400 nm), blue font: blue wavelength (400−500 nm), green font: long wavelength (500−550 nm). Note: Full names of all species are listed in Supplemental Table S5. (b) Wavelength sensitive peak of thrips, modified from Makabe et al.[29]. (c), (d) and (e) Expression levels of vision related genes in thrips. RT-qPCR analysis of three vision genes after 3, 6, and 9 h without UV and UV stress. *** p < 0.0001. The line chart shows the range of parameters of a standard curve.
Three genes related to thrips vision were further analyzed. Relative expression levels in the heads of M. usitatus showed that the Rhodopsin 1, Rhodopsin 2, and Ultraviolet sensitive opsin 1 changed greatly under UV light environment at 6 h compared with dark treatment (p < 0.0001) with the fold change of 3.97-, 1.98- and 3.71-folds, respectively (Fig. 4c, d & e). Furthermore, the expression level of Rhodopsin 1 at 9 h was significantly upregulated compared to the control group (p < 0.001). However, there was no change in the expression levels of the three genes under LED light treatment and UV at 3 h treatment (p > 0.05).
-
Hainan is an important tropical fruit production base, and the town of Foluo in Ledong County is famous for growing cantaloupe. Cowpea is one of the vegetables favored by people all over the country, is one of the main planting varieties of Hainan winter melon and vegetables, and is one of the main sources of income for Hainan farmers. Thrips are considered as one of the serious pests that harm commercial crops[39]. In Hainan, M. usitatus and T. palmi feed on the leaves, flowers, and fruits of their host plants and spread plant diseases, seriously affecting crop yield and quality, and causing huge commercial losses to the local vegetable market[13,40]. The control of thrips mainly relies on insecticides. In this study, the Hainan populations of M. usitatus and T. palmi showed a continuously decreased susceptibility to seven kinds of insecticides between 2021 and 2023.
Many factors contribute to the difference in resistance between the two regions. The Sanya region is the primary cowpea-producing region in Hainan. Insecticides were used more frequently than in the Haikou region to maintain product quality, which puts a great deal of selection pressure on thrips. Insecticides combinations may also cause cross or negative cross-resistance, which could cause varying degrees of insecticide resistance.
Analysis of the mechanism of action benefits greatly from a thorough understanding of sodium channels and nAChRs[16,18]. The mutations M918T, T929I, and L1014F of sodium channels resulted in pyrethroid resistance in T. palmi[41]. F. occidentalis was shown to have a high level of pyrethroid resistance due to the mutations L1014F and T929C[42]. The mutations of M918T/L, T929I, V1010A, L1014F, and T929I + K1774N were reported in several pyrethroid-resistant T. tabaci populations[43−45]. In this study, the mutations of sodium channels in M. usitatus and T. palmi field populations were detected. In addition to the reported mutations, 25 new mutations (located in domains and linkers) were found that may be involved in the pyrethroids resistance.
So far, neonicotinoid resistance of thrips populations due to mutations of nAChR subunits has not been reported. In this study, three β1 subunit mutations were found. This subunit was reported to mediate neonicotinoid resistance in fruit fly and aphids[46,47]. In addition, 15 new mutations were detected in the α1, α2, and α8 subunits of nAChR thrips. It was reported that the G275E/V mutation in the thrips nAChR α6 subunit was involved in the resistance to spinosyns, especially in T. palmi[48−50].
The ability to replace pesticides with novel chemistry and control technologies is nearly being outpaced by the rapid development of insecticide resistance in pests[9]. It is urgent to seek green comprehensive prevention and control technology. Insects use olfactory stimuli in their host location and other behavioral activities. The sense of smell plays a more stable role over long distances compared with others, such as locust migration[51] and ants looking for food[52]. However, more and more studies have demonstrated that insect vision is also indispensable in insect activity, in particular, during accurate location of close range. The specific activation of light-sensitive channels and the precise spectral sensitivity of individual photoreceptors help insects to respond to specific colors and intensities. Aphids and many other insects directly use external light signals and lay their eggs on the back of leaves that lack ultraviolet light[53]. Nymphalida and other insects indirectly use natural light and reflect light to the opposite sex through the vibration frequency of their wings to transmit mating information[54].
Recent studies have shown that new technologies such as environmental light field measurement, serial block-face electron, and microscopy micro-computed tomography[55,56] could be more efficient to face the challenge of working with insects as small as thrips. Many insects, such as whiteflies, thrips, and aphids, choose environments where UV radiation is present or disperse less under an environment without UV radiation[25,57]. The UV absorption or blocking films were found to have good protective effects on greenhouse pests[25,58,59] as a promising physical control method[60,61]. They could modify insect behavior by disrupting color perception[62]. The study by Nguyen et al. showed that covering with UV-absorbing plastic effectively reduced the efficiency of the thrips Ceratothripoides claratris to invade the greenhouse interior by 77%[63].
According to the behavior characteristic of thrips, we are looking for a more ecologically oriented control approach based on the specific ecological contexts such as their vision and other senses that drive their behaviors. In this study, scanning and transmission electron microscopy were used to observe the external morphology and internal ultrastructure of the compound eyes of adult thrips provide the basis for further exploring the visual mechanism of thrips sensitivity to different light waves.
In this study, the yield and quality of cowpea and Hami melon were improved by UVa-FG. The occurrence of pests was also reduced compared with C-FG. This study is the first to show the effectivity of UV-FG on M. usitatus and T. palmi, which may provide a theoretical basis for further study on the behavioral mechanism of these pests. The present results agree with those of the western flower thrip, F. occidentalis, where Kigathi and Poehling found a significant reduction in thrips populations in UV-deficient environments[25].
Many insects are sensitive to UV light, but the degree of sensitivity is different. C. phaseoli was most sensitive to 290–330 nm UV[64]. This could be related to insect opsin type and vision[3,65−67]. The sensitivity of thrip photoreceptors was detected by the retinal potential to sense the direction and intensity of light. The ocellus of the thrips is used to distinguish between day and night and sense the change of seasons[68]. In addition to the different sizes of ommatidia, species, sex, living habits, and living environment of thrips all affect morphological and structural characteristics of thrip compound eyes[3]. The ommatidia in DRA are responsible for receiving polarized light signals[3]. In the present research, under different wavelengths of light, some of the ommatidia in each of the compound eyes had different color responses compared with the surrounding ommatidia. These special ommatidia have UV-shielding pigments and are not sensitive to UV light. In contrast, the lack of UV-shielding pigment in surrounding ommatidia means that the ultraviolet light can normally pass through the semper cell and reach the rod cell where the light is accepted by the corresponding UV-sensitive opsins, appearing dark under the fluorescence microscope. The result obtained by combining with computer vision method for assisted analysis also indicates that thrips indeed have some ommatidia which have significant differences in fluorescence luminance value compared with the surrounding ommatidia. We speculate that there are more shielding pigments in the ommatidia in the marginal region of the back so that the interference of the ambient color is blocked. This ommatidia only accepts the polarized light signal.
It has also been found that visual genes regulate a variety of physiological functions in insects apart from visual perception[69]. It was found that long-wave photosensitive proteins were involved in the development of body color plasticity of N. lugens through the mediation by tyrosine. Other studies identified that Drosophila Rh7 opsin in the midbrain of D. melanogaster played an important role in regulating Drosophila circadian rhythm[68]. Heliconiidae have evolved dual UV-sensitive visual pigments, which are used as signals to other animals and predators through changes of wing color[70−72]. Many insects are very sensitive to UV and have a special visual loop system, which increases their adaptability in tropical and high UV environments[73]. In this study, the cloning and expression analysis revealed that opsin and UV-sensitive genes might be involved in visual communication in thrips. This could be a possible mechanism of UVa-FG in controlling thrips and improving crop yields. More in-depth visual mechanisms, especially the response of thrips to ultraviolet light, need to be further studied. This could reduce the use of insecticides and delay the development of insect resistance to insecticides.
-
This study clarifies the development of thrips resistance to several insecticides in the cowpea and melon-producing regions in Hainan and provides a theoretical basis for green and sustainable management of thrips. It demonstrated that UVa-FG has a high control effect on the thrips and could be used to improve crop growth and yield as an efficient green control measure. The mechanisms that the thrips use to respond to UV lights and adapt to environmental changes were explored in this study by the analyses of the compound eye structure and vision-related genes in thrips. The upregulation of the vision-related genes was demonstrated under UV light conditions. This study lays a foundation for its field application evaluation.
-
The authors confirm contributions to the paper as follows: study conception and design: Li F, Wu S; experiments: Jin H, Yao Z, Xian L, Zhang K, Shi X; data collection: Jin H, Xian L, Liu K; analysis and interpretation of results: Jin H, Yao Z, Xian L, Liu K; draft manuscript preparation: Li F, Jin H, Xian L, Zhang K, Wang L, Jiang W, Wu S. All authors reviewed the results and approved the final version of the manuscript.
-
The transcriptome data generated by this research are deposited in NCBI (Sequence lookup URL:
www.ncbi.nlm.nih.gov/Traces/sra_sub ) and other data needed to evaluate the conclusions are present in this paper and/or the supplementary materials. Original scripts used in the analysis are available upon request (https://zenodo.org/record/8333255 ). This work was supported by Hainan Major Science and Technology Project (ZDKJ2021007), National Key Research and Development Program of China (2022YFD1401200 and 2022YFD1400900), Hainan Province Key Research and Development Project (ZDYF2021XDNY190), The Project of Sanya Yazhou Bay Science and Technology City (SCKJ-JYRC-2023-15) and The 111 project (grand no. D20024).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0024-0014)
-
Received 14 February 2024; Accepted 18 April 2024; Published online 8 July 2024
-
Thrips have developed resistance due to numerous mutations on neuroreceptors.
Ultraviolet -absorbing film in greenhouses can control the number of thrips.
The photosensitive areas in their compound eyes are identified.
Ultraviolet light can affect thrips’ vision.
-
# Authors contributed equally: Fen Li, Haifeng Jin, Zhiye Yao, Limin Xian
- Supplemental Fig. S1 Mutation detection of voltage-gated sodium channel and nicotinic acetylcholine receptors in Megalurothrips usitatus and Thrips palmi.
- Supplemental Table S1 Collection details for field populations of Megalurothrips usitatus and Thrips palmi.
- Supplemental Table S2 Primers used in detection mutation sites.
- Supplemental File S1
- Supplemental Table S3 Primers used in RT-qPCR.
- Supplemental Table S4 Detection of mutation site in the thrips neuroreceptor gene.
- Supplemental File S2
- Supplemental Table S5 Information on the specie names of Fig. 4.
- Supplemental Video S1 Video for the python code of auxiliary analysis.
- Supplemental File 1 The python code of auxiliary analysis (Fig. 3).
- Supplemental File S3
- Supplemental File 2 NCBI download sequence of phylogenetic analysis (Fig. 4).
- Supplemental File 3 Open reading frame sequence of the thrips neuroreceptor gene.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li F, Jin H, Yao Z, Xian L, Liu K, et al. 2024. A new optical practice as an effective alternative to insecticides for controlling highly resistant thrips. Tropical Plants 3: e021 doi: 10.48130/tp-0024-0014 

A new optical practice as an effective alternative to insecticides for controlling highly resistant thrips
- Received: 14 February 2024
- Revised: 10 April 2024
- Accepted: 18 April 2024
- Published online: 08 July 2024
Abstract: Insecticide resistance is one of the most critical problems for the control of thrips in agriculture systems. Thus, sustainable technologies are urgently needed. Thrips (Thysanoptera), a group of small fringed-winged insects, are destructive pests and cause severe economic losses to various crops worldwide. Among thrips, Megalurothrips usitatus (Bagnall) and Thrips palmi (Karny) are the dominant pests of melons and vegetables in tropical regions of China. In this study, it was found that field populations of these two thrips have developed a high level of resistance to multiple insecticides, and many mutations were detected in major insecticide targets (voltage-gated sodium channel and nicotinic acetylcholine receptors) by next-generation sequencing technologies. These results revealed the difficulties in their controlling. To overcome these difficulties, ultraviolet-absorbing film in greenhouses (UVa-FG) was evaluated for its control efficiency of the thrips and crop protection. The results showed that using UVa-FG achieved 96.67% and 97.69% of the control rate for these two thrips with an increase rate in crop yield of 22.5% and 14.3% for cowpea and Hami melon, respectively. Furthermore, the microscopic observation identified the responsive ommatidium specific to UV light in thrips' compound eyes. At the molecular level, five visual genes were cloned and the expression levels of three visual genes were found significantly upregulated under ultraviolet light environments, suggesting they could be the key genes in the regulation of visual changes in different light environments in thrips. The present research demonstrated that the thrips visual system could be used as a new environmentally friendly approach to control thrips.
-
Key words:
- Thrips /
- Insecticide resistance /
- Ultraviolet light sensitive /
- Visual system /
- OpenCV