








-
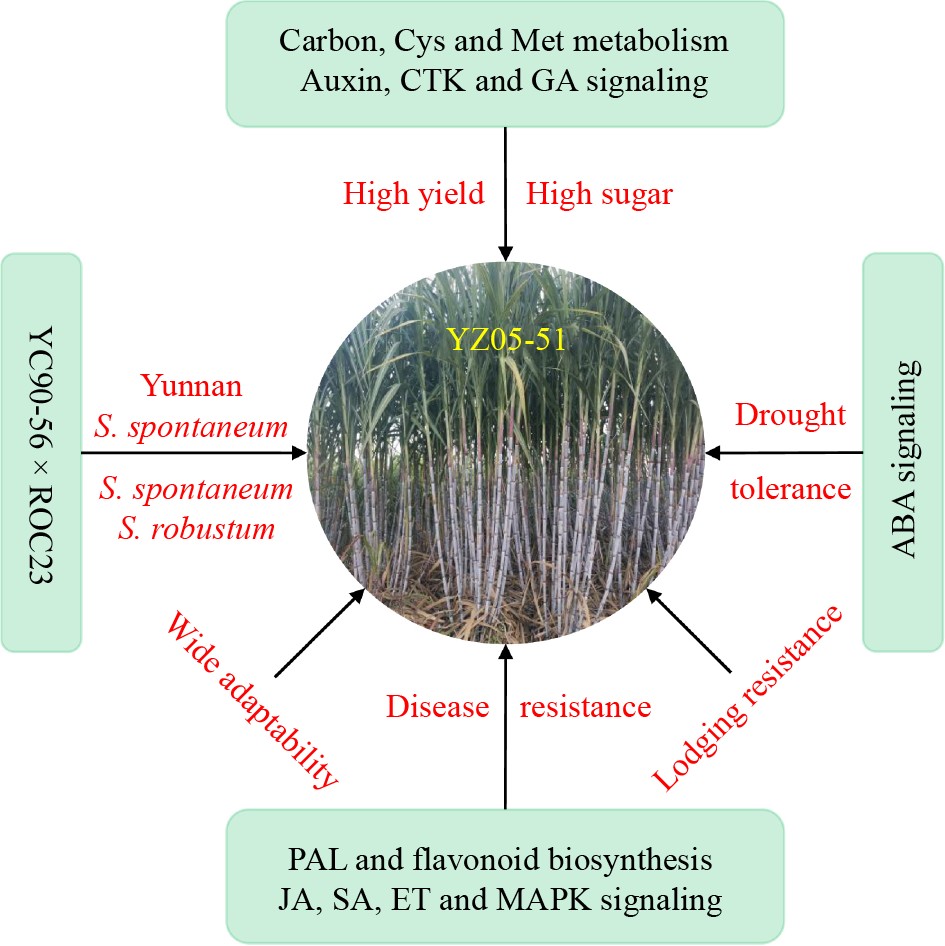
Sugarcane (Saccharum spp) is a significant cash crop in the tropics and subtropics and accounts for 80% of sugar and 40% of bioethanol production in the world. It is a prominent sugar crop in China, with cane sugar contributing to over 85% of sugar production[1, 2]. Saccharum is categorized into cultivated and wild species, encompassing six species, including S. sinense, S. edule, S. barberi, S. officinarum, S. spontaneum, and S. robustum. Notably, S. spontaneum and S. robustum are wild species, while the other four are cultivated[3]. Sugarcane is cultivated in 18 provinces across China, with the primary areas in the tropical and subtropical regions below the latitude line 24° N. These regions are predominantly situated in the southern and southwestern parts of China, particularly in Guangxi, Yunnan, Guangdong, and Hainan provinces[4]. In recent years, there has been a notable shift towards drier slopes in Chinese sugarcane cultivation, resulting in significant ecological changes. Surprisingly, from 2000 to 2013, approximately 85% of sugarcane cultivation was occupied by ROC series varieties, especially ROC22[5]. This has raised concerns about the sustainable development of the sugarcane industry. There is thus a pressing need to prioritize the selection and adoption of new sugarcane varieties.
High yield and high sugar content are essential for a desirable sugarcane variety. As the economy continues to evolve and agricultural science advances, new standards are continually being established for sugarcane varieties. Ideally, they should possess favorable traits such as high yield, high sugar, drought tolerance, disease resistance, resistance to lodging, and wide adaptability[4,6−8]. Recently, researchers have developed the concept of high-heterogeneous composite high-yield and high-sugar breeding, resulting in the breeding of fifth-generation (2013 to now) sugarcane varieties, mainly including Yunzhe 08-1609 (YZ08-1609), YZ05-51, Guitang 42 (GT42), GT44, and Liucheng 05-136 (LC05-136), now accumulatively accounting for over 83.4% of cultivation in China, mostly in Guangxi, Yunnan, Guangdong, and Hainan provinces[4,6,9]. Among them, YZ05-51, developed by Sugarcane Research Institute (SRI) of Yunnan Academy of Agricultural Sciences, stands out as the one bred in China and first registered in the United States. This variety, arising from crossing Yacheng 90-56 (YC90-56) with ROC23, showcases high yield, high sugar, and strong stress resistance, particularly excelling in dry slope conditions[4,10,11]. Notably, in the Gengma county of Yunnan province, YZ05-51 achieved a record-breaking yield of 138 t/ha in non-irrigated areas[4]. Besides, Luo et al.[7,8] utilized GGE biplot to identify that YZ05-51 had relatively high yield and wide stability. In the present review, detailed insights into the breeding logic and genetic characteristics of YZ05-51 are provided. Furthermore, the biological mechanisms for the formation of desirable agronomic traits are explored. Finally, the challenges and prospects are addressed, laying the groundwork for cultivation and breeding of sugarcane.
-
During sugarcane breeding, sexual hybridization, transgene, molecular marker-assisted selection and mutagenesis are frequently used[3], with hybrid breeding being the most effective[4,12,13]. Parental material is crucial for selecting and breeding elite sugarcane varieties. Furthermore, germplasm enhancement plays a key role in the development of breakthrough varieties. In sugarcane hybrid breeding, significant advancements have consistently emerged from the discovery and utilization of novel parental materials throughout the history.
In 1991, the National Sugarcane Germplasm Resource Nursery was established as a national-level gene bank managed by SRI of Yunnan Academy of Agricultural Sciences. At present, it houses more than 5,900 sugarcane germplasm resources, making it the largest one in the world[4]. This nursery has exploited key parent varieties like YC71-374, YC90-56, Yunrui 05-292 (YR05-292), and YR05-770, known for their genetic impact on sugar content, resistance, and other elite agronomic traits. Three sugarcane varieties, namely, YZ05-51, YZ08-1609 and YZ01-1413, have been registered in the United States. Among them, YZ05-51, is an early-maturing variety resulting from the cross of YC90-56 and ROC23 (Fig. 1). The female parent YC90-56 contains the bloodline of Yunnan S. spontaneum (YN57-11-35), and the male parent ROC23 is an elite cultivar developed by Taiwan Sugar Research Institute (Fig. 1). YZ05-51, inheriting a pedigree from POJ, F, Co, and NCo series[14,15] (Fig. 1), achieved a significant breakthrough in high yield on dry slopes, with a record yield of 138 t/ha in the Gengma county of Yunnan province, surpassing the previous record for sugarcane yield in non-irrigated areas[4,10,11]. In China, the introduction of specific bloodlines from the high yield of ROC-type and high resistance of S. spontaneum , along with the bloodlines fusion of large-stemmed wild S. robustum, has expanded the sugarcane breeding bloodlines from four to five. This has enabled the mixing of domestic and foreign parentage, leading to significant progress in sugarcane breeding[4] (Fig. 1).
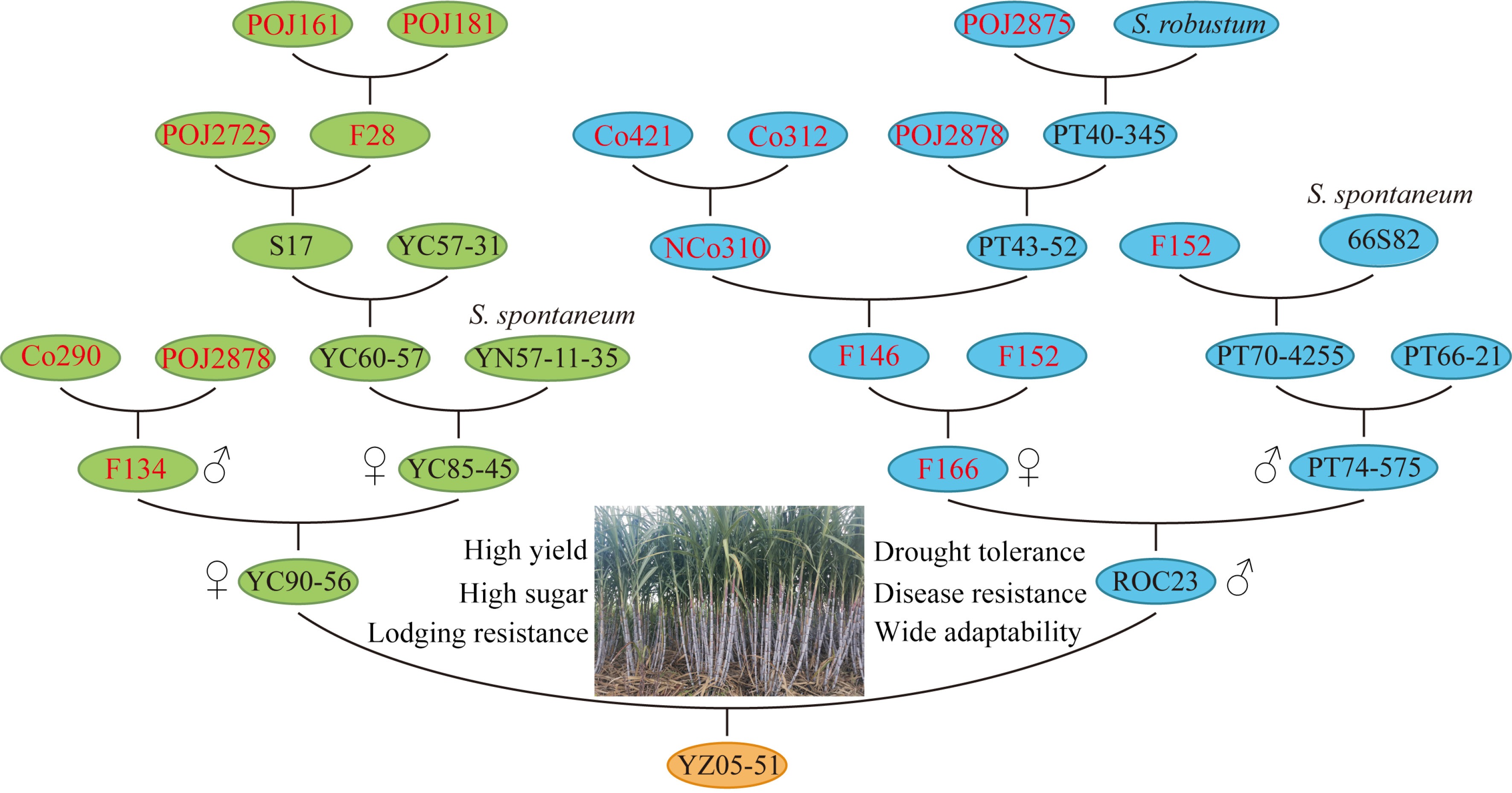
Figure 1.
Pedigree and elite traits of sugarcane variety YZ05-51.
-
YZ05-51 is an early-maturing, high-yielding, and high-sugar variety with several other outstanding traits. It exhibits fast seedling emergence, strong tillering, uniform and neat stems, strong lodging, and easy defoliation. The plants are tall and erect, with medium to large stems, solid, medium-length, cylindrical internodes, thick wax powder, no water splitting, and no aerial roots. Its internodes are initially yellowish-green but turn purple upon exposure. The prismatic buds have medium bodies, shallow and inconspicuous grooves, medium wings, tips that extend beyond the growth zone, bases level with the leaf scars, and moderate root zones. This variety features drooping leaf tips, few or no group 57 hairs, triangular inner leaf auricle, and absent outer leaf auricle. Interestingly, it is highly resistant to smut disease, and highly tolerance to drought stress, but moderately susceptible to mosaic disease (Fig. 2). It demonstrates rapid growth in the initial and middle stages, requiring early fertilization in the seedling period and sufficient fertilization for stem boosting during growth.

Figure 2.
Genetic characteristics of sugarcane variety YZ05-51. (a) seedling stage, (b) elongation stage, (c) maturity stage, (d) plant type, (e) bud, (f) stem, (g) stem longitudinal section, (h) stem transverse section, (i) leaf, (j) leaf sheath, (k) leaf auricle. The original photos were provided by the co-author Dr. Jiayong Liu.
Due to the strong growth potential, it is important to cultivate it in appropriately high soil to prevent late collapse. After harvest, it is necessary to promptly clean up the cane field, ensure early irrigation and root and stem loosening, and to use ground film cover to promote early plant development. It is also important to cover with sugarcane leaves or mulch to utilize soil moisture and promote plant sprouting on dry slopes. Additionally, it is crucial to strengthen pest, disease, and weed control measures. During the growth stage, attention should be given to preventing withered heart in the seedling stage and controlling thrips[10]. In a national regional trial, the average yield is 100.8 t/ha, with a sucrose content of 14.10% in November to December, 15.67% in January to March, and an overall average of 15.00%[11]. YZ05-51 is recommended for row spacing of 1.1~1.2 m, suitable for planting in autumn, winter, and spring, with an ideal density of 112,500~135,000 buds per hectare. It is particularly well-suited for cultivation in Yunnan, Guangxi, Guangdong, and Fujian, especially in locations with ample water and fertilizer resources, offering potential for increased production and raised sucrose content[10]. Since 2017, the planting acreage of YZ05-51 has been rising each year in Yunnan province. Notably, in 2023, the cultivation of this variety reached a nominal value of 31,947 hectare, accounting for 12.13% of the total cultivated area in Yunnan province (Table 1).
Table 1. Information on the trend in promotion acreage for YZ05-51 in Yunnan province.
YZ05-51 2017 2018 2019 2020 2021 2022 2023 Planting area (hectare) 1,380 3,047 5,673 14,640 24,333 27,513 5,673 Percentage of total area (%) 0.48 1.05 1.96 5.08 8.91 10.20 12.13 Percentage of plant-cane area (%) / / / 10.71 17.25 13.67 12.13 The data was provided by the Yunnan Provincial Department of Agriculture and Rural Development. -
During the eighth national sugarcane regional trial at 14 locations in China (2011-2011, two plant-cane crops and one ratoon crop), except for Zhanjiang (ZJ) of Guangdong and Lingao (LG) of Hainan, YZ05-51 exhibited a higher average cane yield than the control ROC22 (Fig. 3a). Specifically, at Chongzuo (CZ) of Guangxi, Kaiyuan (KY) and Lincang (LC) of Yunnan, the average cane yield of YZ05-51 was significantly higher than that of ROC22, with an increasing of 6.65%, 8.51%, and 34.35%, respectively (Fig. 3a). Except for ZJ, Baise (BS) and Laibin (LB) of Guangxi, and LG, the average sucrose content of YZ05-51 was higher than ROC22 (Fig. 3b), while at Fuzhou (FZ) of Fujian, KY, and LC, the average cane yield of YZ05-51 was 4.80%, 15.39%, and 38.76% higher than ROC22, respectively[9,10] (Fig. 3b). Previous studies revealed that tillering ability and well development, controlled by cysteine and methionine metabolism, have a significant impact on sugarcane yield[16, 17]. In the present study, transcriptomics integrated with metabolomics (unpublished) identified three differential accumulated metabolites (DAM; L-aspartic acid, L-homocystine, and 3P-D-glycerate) and six differentially expressed genes (DEGs) including cysE, hom, TAT, MTAN, mtnA, and mtnD (Fig. 3c). Carbon fixation and starch/sucrose metabolism provides energy for plant growth and dry matter accumulation are vital for plant development and biomass accumulation[18−20]. In the present study, L-aspartic acid was also discovered, along with eight DEGs such as Rubisco[21], PGK, GAPDH, MDH2, β-Glu, endo-Glu, TPS, and BAM (Fig. 3d). Phytohormones are acknowledged for their impact on plant growth and development[22]. In the present study, six DEGs including AUX/IAA, ARF, GH3, SAUR, A-ARR, and GID1 involved in Auxin[23], CTK[24] and GA[25,26] signaling pathways, were identified (Fig. 3e). These above DEGs and DAMs jointly contributed to the high yield and high sugar for YZ05-51.
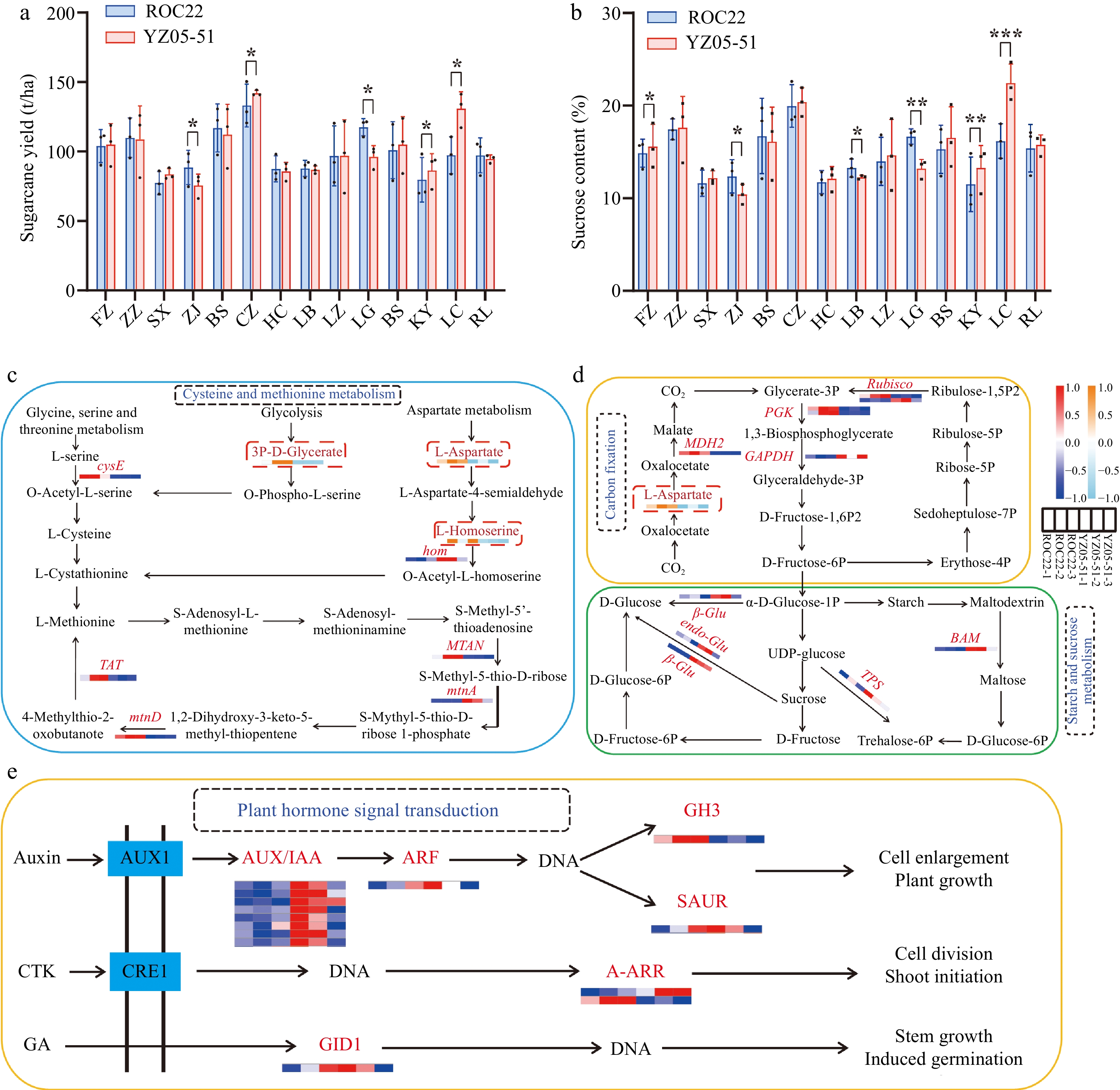
Figure 3.
Biological basis of high yield and high sugar in YZ05-51. (a), (b) Cane yield and sucrose content of YZ05-51 and ROC22 in the eighth national sugarcane regional trial (2011−2012, two plant-cane crops and one ratoon crop)[10,11]. t/ha: tons per hectare. Significance (* for p < 0.05, ** for p < 0.01, and *** for p < 0.001) and standard deviation (SD) were calculated using two-way analysis of variance (ANOVA) followed by Duncan's new multiple range test of GraphPad Prism 8.0 software. FZ, Fuzhou of Fujian; ZZ, Zhangzhou of Fujian; SX, Suixi of Guangdong; ZJ, Zhanjiang of Guangdong; BS, Baise of Guangxi; CZ, Chongzuo of Guangxi; HC, Hechi of Guangxi; LB, Laibin of Guangxi; LZ, Liuzhou of Guangxi; LG, Lingao of Hainan; BS, Baoshan of Yunnan; KY, Kaiyuan of Yunnan; LC, Lincang of Yunnan; RL, Ruili of Yunnan. (c) Cysteine and methionine metabolism. (d) Carbon fixation and starch/sucrose metabolism. (e) Plant hormone signal transduction. The heat maps represent the expression of DEGs and DAMs in YZ05-51 compared to ROC22. Red/orange and blue/sky-blue colors indicate up-regulated and down-regulated DEGs/DAMs, respectively. cysE, serine O-acetyltransferase; hom, homoserine dehydrogenase; TAT, tyrosine aminotransferase; MTAN, 5'-methylthioadenosine nucleosidases; MtnA, methylthioribose-1-phosphate isomerase; MtnD, 1,2-dihydroxy-3-keto-5-methylthiopentene dioxygenase; Rubisco, ribulose-bisphosphate carboxylase; PGK, phosphoglycerate kinase; GAPDH, glyceraldehyde 3-phosphate dehydrogenase (phosphorylating); MDH2, malate dehydrogenase 2; β-Glu, beta-glucosidase; endo-Glu, endoglucanase; TPS, trehalose 6-phosphate synthase. BAM, beta-amylase; AUX/IAA, auxin; ARF, auxin response factor; GH3, gretchen hagen 3; SAUR, small auxin up-regulated RNA; A-ARR, type-A Arabidopsis response regulator; GID1, GIBBERELLIN-INSENSITIVE DWARF1.
-
Since the 1950s, the global temperature has been increasing by approximately 0.13 °C every decade, causing global water scarcity due to rising temperatures and declining precipitation[27]. The frequent occurrence of droughts in agricultural areas has significantly jeopardized agricultural production, resulting in substantial reductions in crop yields and severe impacts on its quality. In plants, abscisic acid (ABA) signaling pathway is the most studied signaling pathway involved in plant drought stress response[28]. In the present study, several ABA-related DEGs including PYR/PYL, PP2C, SnRK2 and ABF were identified, most of which had a higher expression level in YZ05-51 compared to ROC22 (Fig. 4a). Besides, six ABA-related genes (PYL8, two PP2C9, SAPK8, SAPK4, and ABF9) were up-regulated under drought stress in sugarcane (Fig. 4c). Plants frequently generate high levels of reactive oxygen species (ROS) in response to stress. To mitigate this, they utilize enzymes like ascorbate peroxidase (APX), catalase (CAT), peroxidase (POD) and superoxide dismutase (SOD) to remove excess toxic ROS[29]. In the current dataset, five ROS-related DEGs (three APX, CAT, and SOD) were detected (Fig. 4b), among which four were up-regulated under drought stress (Fig. 4c). Transcription factors (TFs), including MYB, WRKY, ERF, NAC, and bZIP, have been confirmed as key players in drought stress tolerance[30−34]. Here, it was observed that the expression of several DREB (2), WRKY (7), and bZIP (8) were induced by drought stress (Fig. 4b, c). We can thus deduce that, those DEGs involved in ABA signaling pathways, ROS-related enzymes and DREB, WRKY, and bZIP TFs collectively enhanced the tolerance to drought stress in sugarcane.
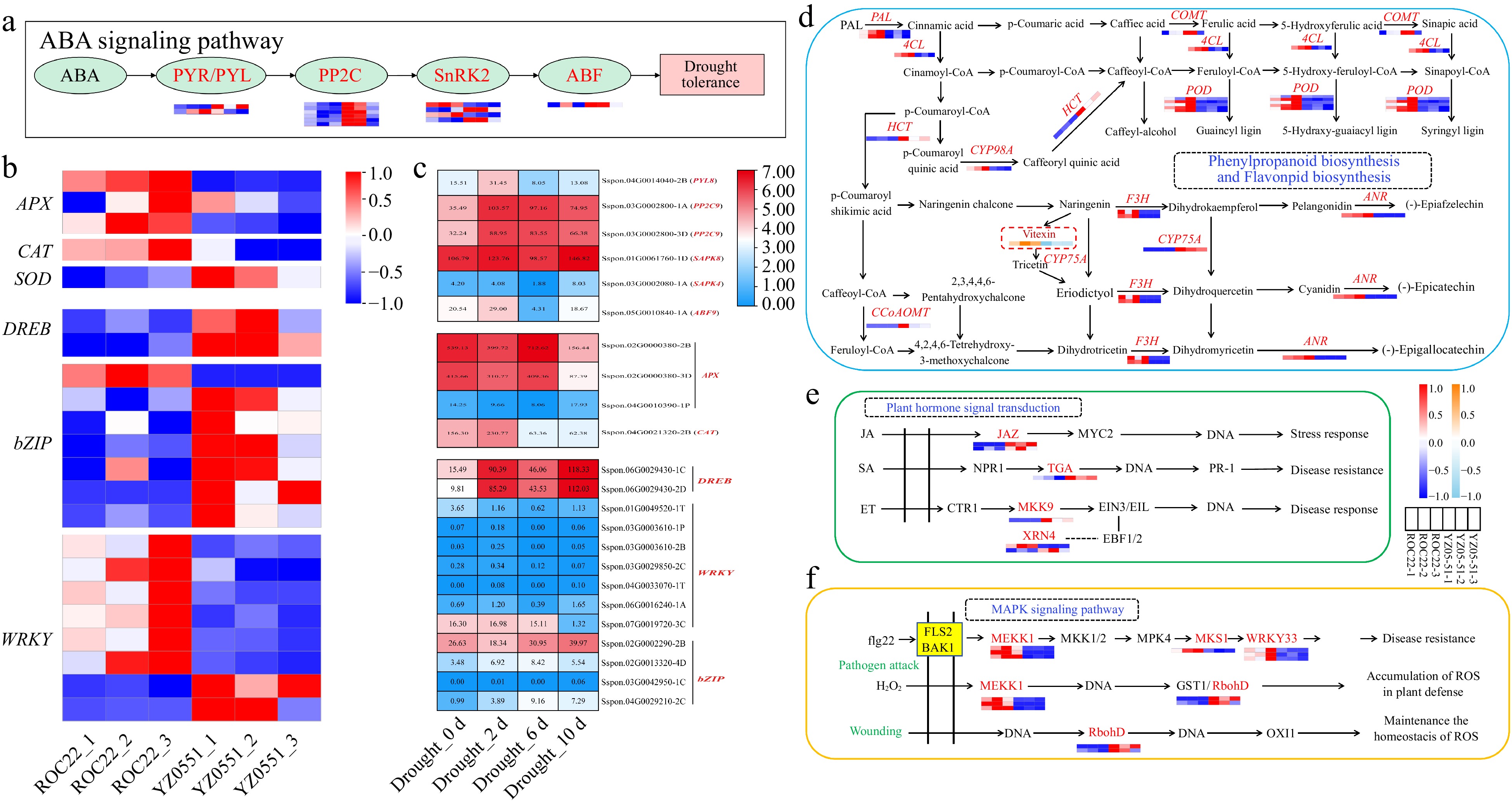
Figure 4.
Biological basis of drought tolerance and disease resistance in YZ05-51. (a) ABA signaling pathway. (b) ROS-related genes and transcription factors (TFs) in YZ05-51 compared to ROC22. (c) Expression patterns of ABA-related, ROS-related, DREB, WRKY, and bZIP genes. (d) Phenylpropanoid and flavonoid biosynthesis. (e) JA, SA, and ET signaling pathways. (f) MAPK signaling pathway. Red/orange and blue/sky-blue colors indicate up-regulated and down-regulated DEGs/DAMs, respectively. The source data of S. spontaneum subjected to drought treatment was obtained from NCBI website under project PRJNA636260[35,36]. PYR/PYL, abscisic acid receptor PYR/PYL family; PP2C, protein phosphatase 2C; SnRK2, serine/threonine-protein kinase 2; ABF, ABRE binding factors; APX, ascorbate peroxidase; CAT, catalase, SOD, superoxide dismutase. PAL, phenylalanine ammonia-lyase; 4CL, 4-coumarate-CoA ligase; COMT, caffeic acid 3-O-methyltransferase; POD, peroxidase; HCT, shikimate O-hydroxycinnamoyltransferase; CYP98A, 5-O-(4-coumaroyl)-D-quinate 3'-monooxygenase; F3H, naringenin 3-dioxygenase; ANR, anthocyanidin reductase; CYP75A, flavonoid 3',5'-hydroxylase; CCoAOMT, caffeoyl-CoA O-methyltransferase; JAZ, jasmonate ZIM domain-containing protein; TGA, transcription factor TGA; MKK9, mitogen-activated protein kinase kinase 9; XRN4, 5'-3' exoribonuclease 4; MEKK1, mitogen-activated protein kinase kinase kinase 1; MKS1, MAP kinase substrate 1; WRKY33, WRKY transcription factor 33; RbohD, respiratory burst oxidase D.
In plants, phenylpropanoid and flavonoid effectively regulate the accumulation of ROS in cells. They serve as the primary source of plant defensive secondary metabolites[37−39]. Here in our study, one DAM (Vitexin) and 10 DEGs (PAL, 4CL, COMT, POD, CYP98A, CYP75A, HCT, F3H, ANR, and CCoAOMT) were determined in the phenylpropanoid and flavonoid biosynthesis pathways (Fig. 4d). As shown in Fig. 4e, JAZ, TGA, MKK9, and XRN4 involved in the jasmonate acid (JA)[40,41], salicylic acid (SA)[42], and ethylene (ET)[43] signaling pathways, were identified. When the plant is infected with pathogens, to promote resistance, pathogen-associated molecular proteins (PAMPs) bind to receptor-like proteins (RLKs) or disease resistance proteins such as Cf9, CNGCs, and FLS2 to activate MAPK cascade signaling and pathogen secretion system[44]. Based on our RNA-Seq data, MEKK1, MKS1, WRKY33, and RbohD enriched in MAPK pathways, were also discovered (Fig. 4f). Collectively, the DEGs and DAMs involved in phenylpropanoid and flavonoid biosynthesis, JA, SA, ET, and MAPK signaling pathways improved the resistance of sugarcane to pathogen infection, which is the potential reason why YZ05-51 is highly disease-resistance.
-
Currently, sugarcane breeding faces several challenges, including a limited genetic pool, low breeding efficiency, and the struggle to develop innovative sugarcane varieties. Besides, resource sharing between different breeding units is challenging, information exchange, and communication are lacking, and collaboration between industry and academia is not as robust as needed. Additionally, mutagenic and transgenic breeding have not seen adequate advancement, and sugar industry enterprises are considered to be more actively involved in sugarcane breeding. To address these challenges, it is essential to enhance the introduction of sugarcane varieties and germplasm resources to broaden the genetic diversity. This involves identifying and selecting superior germplasm through rigorous testing and evaluation. Efforts should also be made to collect, study, and utilize wild sugarcane germplasm resources to generate innovative germplasm for further crossbreeding initiatives, with a focus on crossbreeding and backcrossing S. spontaneum and S. arundinaceum. Furthermore, research on sugarcane hybridization needs to be intensified, especially flowering induction for difficult-to-flowering parents. Moreover, refinement of variety selection and breeding procedures, along with comprehensive evaluation for disease and drought resistance in hybrid progeny, is indispensable. Continued efforts in gene cloning, transgenic breeding, and gene editing are urgently needed to cultivate superior sugarcane lines with huge potential. Lastly, the industrialization of healthy seedlings is imperative for expediting the dissemination of a sugarcane variety, which facilitates the adoption of sugarcane varieties and enhances the standards for seed production, leading to cost reduction and increased profitability.
Taken together, YZ05-51, resulting from a cross between YC90-56 and ROC23, demonstrate high yield, high sugar, strong stress resistance, and wide adaptability, particularly thriving in dry slope conditions. With a combination of high-yield characteristics from ROC-type and high-resistance traits from S. spontaneum, along with a pedigree from POJ, F, Co, and NCo series, it represents a significant advancement in sugarcane production. This variety has surpassed previous records for cane yield in non-irrigated areas, achieving a remarkable yield of 138 t/ha in Yunnan province. We can reasonably deduce that the widespread use of healthy seedlings has contributed to the promotion of YZ05-51, approximately 31,947 hectare, constituting 12.13% of the total cultivated area in Yunnan province. Hopefully, the successful breeding and widespread adoption of YZ05-51 establish a solid foundation for the development of sugarcane varieties with desirable agronomic traits.
-
The authors confirm contribution to the paper as follows: formal analysis, methodology: Wu Q; visualization: Wu Q, Li A, Liu J, Zhao Y, Zhao P; funding acquisition: Wu Q, Que Y; writing – original draf: Wu Q, Li A; writing – review & editing: Liu J, Zhao Y, Zhao P, Zhang Y, Que Y; data curation: Liu J, Zhao Y, Zhao P; conceptualization, project administration, resources, supervision: Zhang Y, Que Y. All authors reviewed the results and approved the final version of the manuscript.
-
The raw RNA-Seq data of this study were deposited in the Genome Sequence Archive at the China National Center for Bioinformation (
www.cncb.ac.cn ). The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding authors. This work was funded by the Project of National Key Laboratory for Tropical Crop Breeding (NKLTCB20230305, Hainan; NKLTCB-YAAS-2024-S01, Yunnan), China Agriculture Research System of MOF and MARA (CARS-17), Central Public-interest Scientific Institution Basal Research Fund (1630052024003 and 1630052024020), Guangxi Key Laboratory of Sugarcane Genetic Improvement (21-238-16-K-02-03), and Yunnan Key Laboratory of Sugarcane Genetic Improvement (2023KFKT001).
-
The authors declare that they have no conflict of interest.
-
Received 20 April 2024; Accepted 29 April 2024; Published online 5 June 2024
-
The breeding logic and agronomic features of YZ05-51 are described in detail.
The biological basis of the elite traits of YZ05-51 is dissected.
The challenges and prospects of sugarcane breeding are discussed.
-
# Authors contributed equally: Qibin Wu, Aomei Li
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wu Q, Li A, Liu J, Zhao Y, Zhao P, et al. 2024. Sugarcane variety YZ05-51 with high yield and strong resistance: breeding and cultivation perspectives. Tropical Plants 3: e019 doi: 10.48130/tp-0024-0019 

Sugarcane variety YZ05-51 with high yield and strong resistance: breeding and cultivation perspectives
- Received: 20 April 2024
- Revised: 26 April 2024
- Accepted: 29 April 2024
- Published online: 05 June 2024
Abstract: Sugarcane (Saccharum spp), a significant cash crop in the tropics and subtropics, accounts for 80% of sugar and 40% of bioethanol in the world. In China, it is also a prominent sugar crop, with cane sugar contributing to over 85% of sugar production. In recent years, there has been a notable shift towards drier slopes in Chinese sugarcane cultivation, resulting in significant ecological changes. Over the past two decades, scientists have developed the theory of high-heterogeneous composite high-yield and high-sugar breeding, leading to the successful breeding of the fifth-generation sugarcane varieties, among which YZ05-51 is a representative one. It was developed by the Sugarcane Research Institute (YSRI) of Yunnan Academy of Agricultural Sciences. Originating from a cross between YC90-56 and ROC23, YZ05-51 demonstrates several excellent agronomic traits, such as high yield, high sugar, strong stress resistance, and wide adaptability, particularly thriving in dry slope conditions. By 2023, cultivation of YZ05-51 has reached approximately 31,947 hectare, constituting 12.13% of the total cultivated area in Yunnan province. This variety has even surpassed previous records for sugarcane yield in non-irrigated areas, achieving a remarkable yield of 138 tons per hectare (t/ha). To summarize experience and inspire innovation, here the insights into the breeding logic and genetic characteristics of YZ05-51 are detailed. Further, the biological mechanisms underlying its agronomic traits were investigated through the integration of transcriptomics and metabolomics analysis. The challenges and prospects were also discussed, setting up the basis for the breeding and promotion of newly bred sugarcane varieties.
-
Key words:
- Sugarcane /
- Hybrid breeding /
- YZ05-51 /
- Agronomic trait /
- Biological basis