









-
Tropical high mountain ecosystems host important biodiversity hotspots[1,2]. These precious ecosystems are under threat of rapid biodiversity loss which is a global concern[2−4]. Globally, 20% of plant species are on the brink of extinction due to both anthropogenic and natural factors[4]. Climate change[3,5], habitat fragmentation[3,4], invasion of alien species[6−10], pathogens[4], overexploitation[3,4,11], and a booming human population[3,4] are major risk factors contributing to species extinction.
Tree heath (Erica arborea L.), referred to hereafter as Erica, (family Ericaceae) is a much-branched evergreen shrub or small tree up to 12 m tall[12,13]. It is native to East and North Africa, Southern and Eastern Europe and a few countries in West Asia[14,15]. It grows on dry rocky areas with shallow soils[15]. In the African highlands, it is found between 1,900 to 4,000 m usually as a distinct vegetation belt above Afromontane forests (Ericaceous Belt), and also in clearings within forested areas. Above 3,500 m, it is found as a small bush[1,16] while in the Mediterranean it is found in evergreen shrublands or maquis along the coasts up to more than 1,100 m[13].
Erica is used for various purposes. For instance, it produces copious pink pollen and abundant nectar throughout most of the year. This makes Erica one of the most important honey sources at higher altitudes[17] as it fetches premium prices and higher monetary returns, especially for the Irob communities in Tigrai, Ethiopia (personal communication). Erica also produces lignotubers that are highly treasured for making briar pipes[13] and dice[17]. In traditional medicine, the smoked parts of the species are used to fumigate houses against contagious diseases[17]. Branches and wood are popular for firewood and charcoal production[18]. It is also used as fodder and for live fences[15].
Erica is experiencing a drastic population decline throughout its native range. Globally, though there is no known population information[14,19], the species is listed as Vulnerable in Andorra and Bulgaria and as a Least Concern within Europe[14]. Likewise, WeForest[20] reported that Erica was extinct in four villages out of 11 villages from the dry Afromontane Desa'a forest in Tigrai. Similarly, in the Peloritani Mountains of Italy, nearly 5,800 ha of Erica-dominated shrublands have vanished[13]. These problems were associated with farmland expansion, wildfire, free grazing, new settlements, illegal logging, and charcoal making[18,20,21]. In addition, climate change has become a clear threat to the distribution, growth, and survival of many Indigenous plant species particularly in recent years[22].
Most tree species distributions are affected by climate change[23−27]. To survive the effects of climate change, species in the tropics may be pushed to higher elevations by effectively shrinking their distributions[28] or migrating to new suitable habitats[25]. Among the species most vulnerable to reductions in suitable habitat due to climate change are those that are adapted to Afroalpine forest ecosystems[2,23]. These ecosystems host many rare, endangered, and flagship species[23].
Previous studies on Erica focused on dendrochronology[21], phylogeography inferred from AFLPs and plastid DNA[12], the effects of fire and grazing[29,30] and tree heath utilization[13]. To the best of our knowledge, studies focusing on the ecological niche (distribution pattern with climate change) of Erica using species distribution models (SDMs) and ArcGIS and how global climate change might affect Erica's distribution in the future are limited. However, to mitigate the effects of climate change, it is possible to effectively target conservation strategies by modelling species distributions and identifying areas where sensitive species exist or are likely to exist[2,26]. Because effective conservation planning requires a comprehensive understanding of the relationships of species to their environment under changing climatic conditions[31,32].
SDMs constitute the most common class of models across ecology, evolution, and conservation[33] in view of supporting species conservation and management interventions. Among the many SDM algorithms, the open-source software Maxent (
https://biodiversityinformatics.amnh.org/open_source/maxent/ ) has proven powerful when modelling rare and endangered species[11,26,34] using presence-only occurrence data[26,35]. Because SDM is valuable for anticipating conservation regions (particularly zones for species protection, restoration, translocation, and reintroductions) and for addressing questions about niche evolution trends[2]. This is the first study to use a niche modelling approach to model the distribution of Erica and the resulting outputs of the study may provide valuable information for conservation, management, and research of the species. Hence, this study aimed to: (1) predict the current potential distribution of Erica in its native distribution ranges; (2) identify the environmental variables associated with the Erica habitat distribution; and (3) assess the potential impact of climate change on the future distribution of the species. -
The study area was located in the mountain chain highlands of Tigrai, Ethiopia (Fig. 1). Tigrai exhibits agroecological diversity, characterized by variations in temperature, rainfall, topography, soil characteristics, vegetation cover, and other natural resources[22]. As such, it is part of the Eastern Afromontane biodiversity hotspot[18,36] due to its natural landscape and diversified agro-ecology[22]. The highlands of Tigrai are home to the remnant dry Afromontane forests of Asimba Natural Forest (14°15'–14°30' N and 39°30'–39°45' E), Desa'a National Forest Priority Area (13°20'–14°10' N and 39°32'–39°55 E), Hugumburda Grat Kahsu National Forest Priority Area (12°33'–12°42' N and 39°30'–39°39' E), Wujig Mahgo Waren Forest (12°47'–13°02' N and 39°26'–39°39' E) and the church forests of Michael Abeda, and Tabotat. The topographies of the study sites are characterised by a mountain plateau with undulating terrain interspersed with low-situated valleys, hills, and flatlands. The amount of rainfall for Asimba and Desa'a ranges from 116.3 to 230.0 mm during the main rainy season while the temperature range is 7.5 to 22.6 °C. Hugumburda Grat Kahsu, Wujig Mahgo Waren, Michael Abeda, and Tabotat receive mean annual rainfall ranging between 350 mm and more than 1,000 mm, mean minimum annual temperature ranges between 6.3 and 20.6 °C and the mean maximum annual temperature ranges from 15.1 to 29.7 °C.
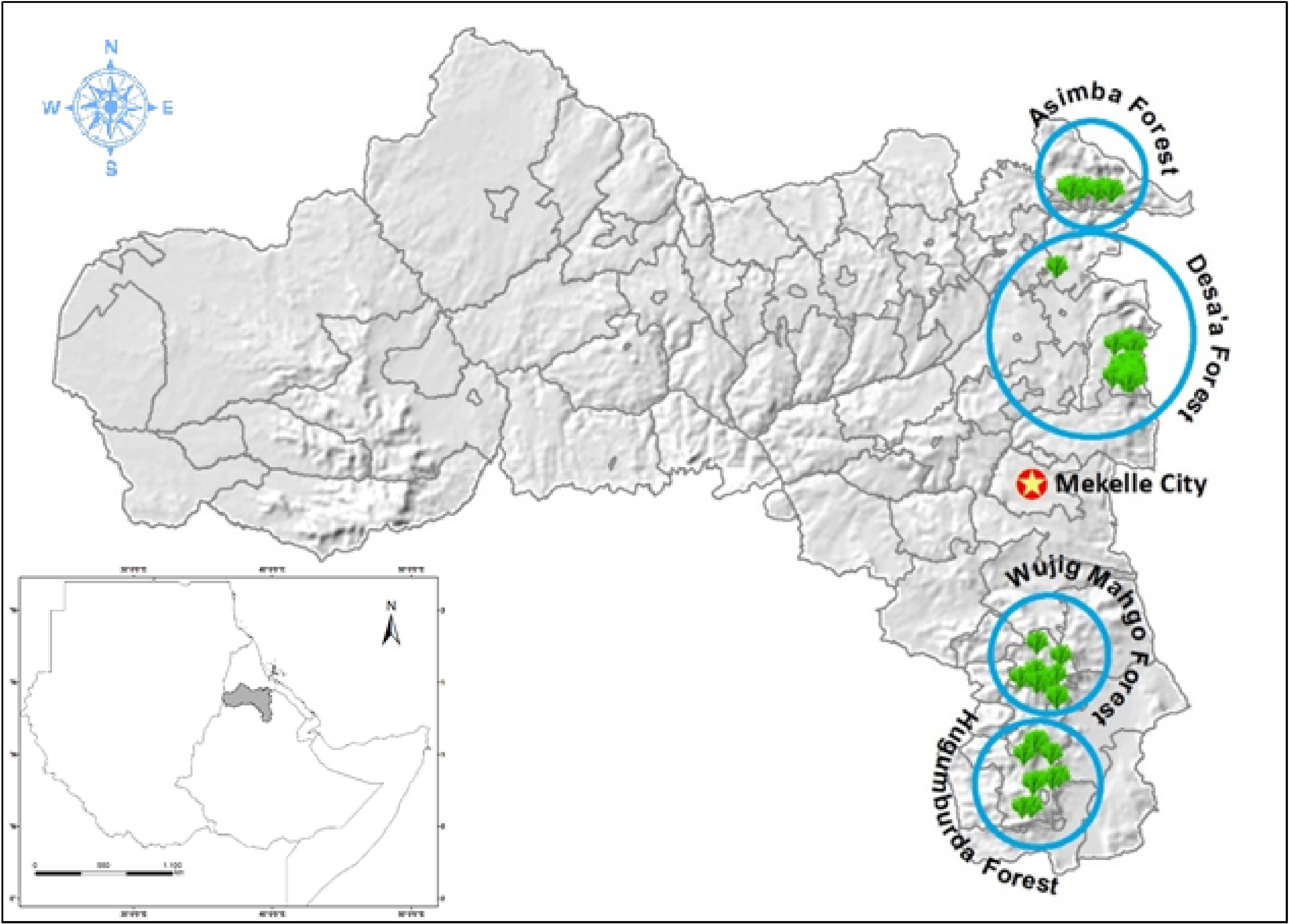
Figure 1.
Location of the study area in relation to the Horn of Africa. The blue circles refer to the regional forest areas where the occurrence points were taken. The green areas are used to indicate forest stands (trees).
Target species and occurrence data
-
The target species for the species distribution modelling simulation was the rare native[18,37] tree Erica (Fig. 2). It is among the rare species (IVI of 0.26−0.58) in the highlands of Tigrai with poor seedling regeneration[38]. The species has experienced local extinction mainly due to logging for its good fuelwood[37] and charcoal production in recent years following the genocidal war and due to climatic factors.

Figure 2.
(a) Erica arborea L. and its landscape around Embahasti – Alaje, and (b) an Erica patch on the northern slopes of the Dabba Selama peak in Dogu'a Tembien.
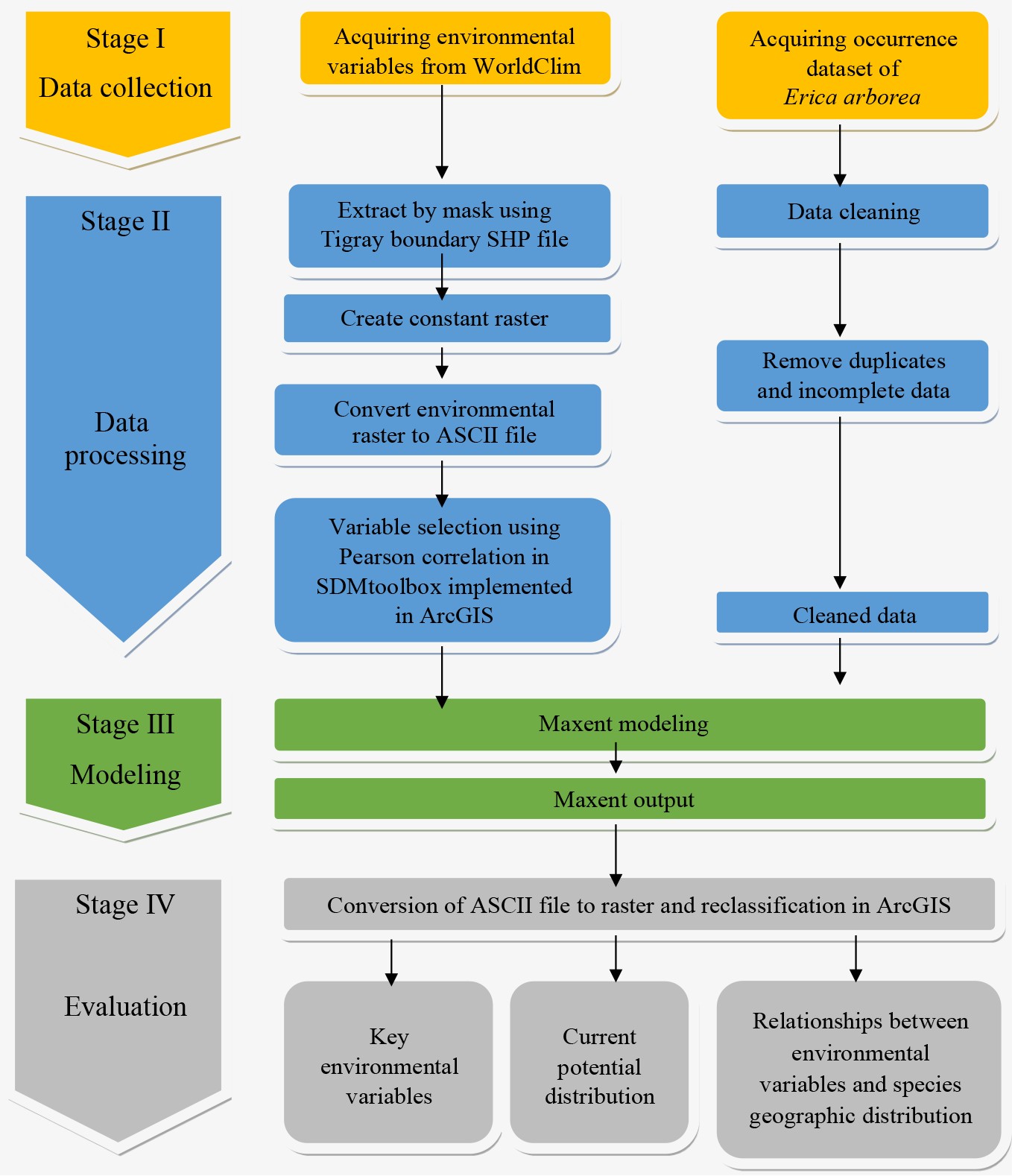
For modelling of the species, presence-only occurrence data was collected for 89 species from the study forests (Fig. 2) and from WeForest records. These occurrence data were converted to decimal degrees and then cleaned by removing outliers and duplicates in each 1 km × 1 km grid at the scale of the bioclimatic variables used[11]. To eliminate duplicated occurrence points and minimize the potential influence of environmental variations on model accuracy, the Python-based GIS toolkit SDM toolbox was used, implemented in ArcGIS[22,39]. Finally, 58 occurrence data points were retained and were used to determine the species distribution using Maxent.
Predictor environmental variables
-
Twenty-three environmental variables (consisting of 19 bioclimatic variables and four topographic variables) were downloaded from different sources to simulate the distributions of the species. The 19 climatic variables and altitude data were retrieved from the World Climate Database (
www.worldclim.org ) at a spatial resolution of 1 km. From the altitude data, aspect and slope were extracted while the distance from the sea was calculated as the shortest distance from the shoreline using ArcGIS 10.8.1.To reduce multicollinearity among the 19 bioclimatic variables and the four topographic variables, highly correlated variables (r ≥ |0.85| Pearson correlation coefficient) were eliminated from further models[40]. This reduction in predictor variables resulted in the inclusion of 11 variables in the models (Table 1 & Supplementary Table S1). The selected variables included temperature annual range, annual precipitation, precipitation in the driest month, precipitation in the wettest quarter, precipitation in the driest quarter, precipitation in the warmest quarter, precipitation in the coldest quarter, altitude, slope, aspect, and distance from the sea (Table 1).
Table 1. Environmental variables used for distribution modelling of Erica in Tigrai.
Label Environmental variables Factor Unit bio1 Annual mean temperature 10 °C bio2 Mean diurnal range (mean of monthly (max temp – min temp)) 10 °C bio3 Isothermality (bio2/bio7)*(100) 100 − bio4 Temperature seasonality (standard deviation*100) 100 % bio5 Max temperature of warmest month 10 °C bio6 Min temperature of coldest month 10 °C bio7 Temperature annual range (bio5−bio6) 10 °C bio8 Mean temperature of wettest quarter 10 °C bio9 Mean temperature of driest quarter 10 °C bio10 Mean temperature of warmest quarter 10 °C bio11 Mean temperature of coldest quarter 10 °C bio12 Annual precipitation 1 mm bio13 Precipitation of wettest month 1 mm bio14 Precipitation of driest month 1 mm bio15 Precipitation of seasonality (coefficient of variation) 100 % bio16 Precipitation of wettest quarter 1 mm bio17 Precipitation of driest quarter 1 mm bio18 Precipitation of warmest quarter 1 mm bio19 Precipitation of coldest quarter 1 mm Altitude 1 m a. s. l Aspect 1 degree Distance from sea 1 km Slope gradient 1 % Those highlighted in bold refer to the selected environmental variable for the modelling. Factor refers to the scale by which the environmental variable is multiplied for ease of storage in the raster. Future climate scenario
-
Climate data obtained from WorldClim was used to model the current and future potential distributions of Erica. The current climate data represent the average for the years 1950–2000. Future climate projections were developed for the years 2050 (2041–2060) and 2070 (2061–2080). Both current and future climate data were obtained from a global atmospheric circulation model, the Community Climate System Model version 4 (CCSM4)[41−43] and two representative concentration pathways (RCPs). RCP4.5 (intermediate GHG emission hypothesis) and RCP8.5 (maximum GHG emission hypothesis) were considered in our study for future species distribution predictions. These RCP scenarios represent climate conditions under which radiative forcing is projected to increase by 4.5 and 8.5 Watts per square meter (Wm−2) by the year 2100[28,41]. In other words, the RCP4.5 scenario represents a pathway in which global greenhouse emissions gradually increase, followed by a decline after 2030 while the RCP8.5 scenario represents a 'baseline' scenario pathway with no climate mitigation target and an increase in emissions throughout the century[44].
Modelling approaches
-
To predict the potential distribution of Erica, Maxent 3.4.1 was used[45,46]. As input, the selected uncorrelated environmental variables and the species occurrence data of Erica were loaded into the Maxent model. To simulate the model, 75% of the occurrence data were used for calibration (training) and the remaining 25% were used for validation[47]. Furthermore, to assess the relative contribution of each variable to the model, the Jackknife test[45] and the area under the curve (AUC) of the receiver operating characteristic curve (ROC) were used[11,40,48]. The AUC values were categorized as invalid (0.5–0.6), poor (0.6–0.7), average (0.7–0.8), good (0.8–0.9), or excellent (0.9–1)[40]. Thus, prediction results can be adopted when the AUC is 0.7–0.9 and when the AUC > 0.9, indicating that the prediction results are very accurate[48].
Finally, using the 10th percentile training presence logistic threshold as the cut-off point, a habitat suitability index divided into five classes was developed. The classes are 0.0–0.2 (unsuitable), 0.2–0.4 (low suitable), 0.4–0.6 (suitable), 0.6–0.8 (highly suitable), and 0.8–1.0 (exceptionally suitable)[26,49]. Based on this classification, an area less than 0.2 is considered as unsuitable, while areas between 0.2 and 1 are considered as suitable and areas above 0.6 are considered as optimally suitable[40]. The total extent of occurrence (EOO) of the species was also calculated based on the procedure outlined in IUCN[50]. This approach considers 0.5 as the threshold above which the species is most likely present[51].
-
The Maxent model performed well in predicting the potential distribution of Erica in its native range (Fig. 3). Except for RCP8.5 in 2070, which had an AUC value of 0.888 ± 0.085, all the AUC values (0.929 ± 0.039 under the current scenario, 0.923 ± 0.031 under RCP4.5 and 0.912 ± 0.036 under RCP8.5 in 2050 and 0.901 ± 0.043 under RCP4.5 in 2070) were > 0.9 indicating that the model's high accuracy and its high discriminative ability.
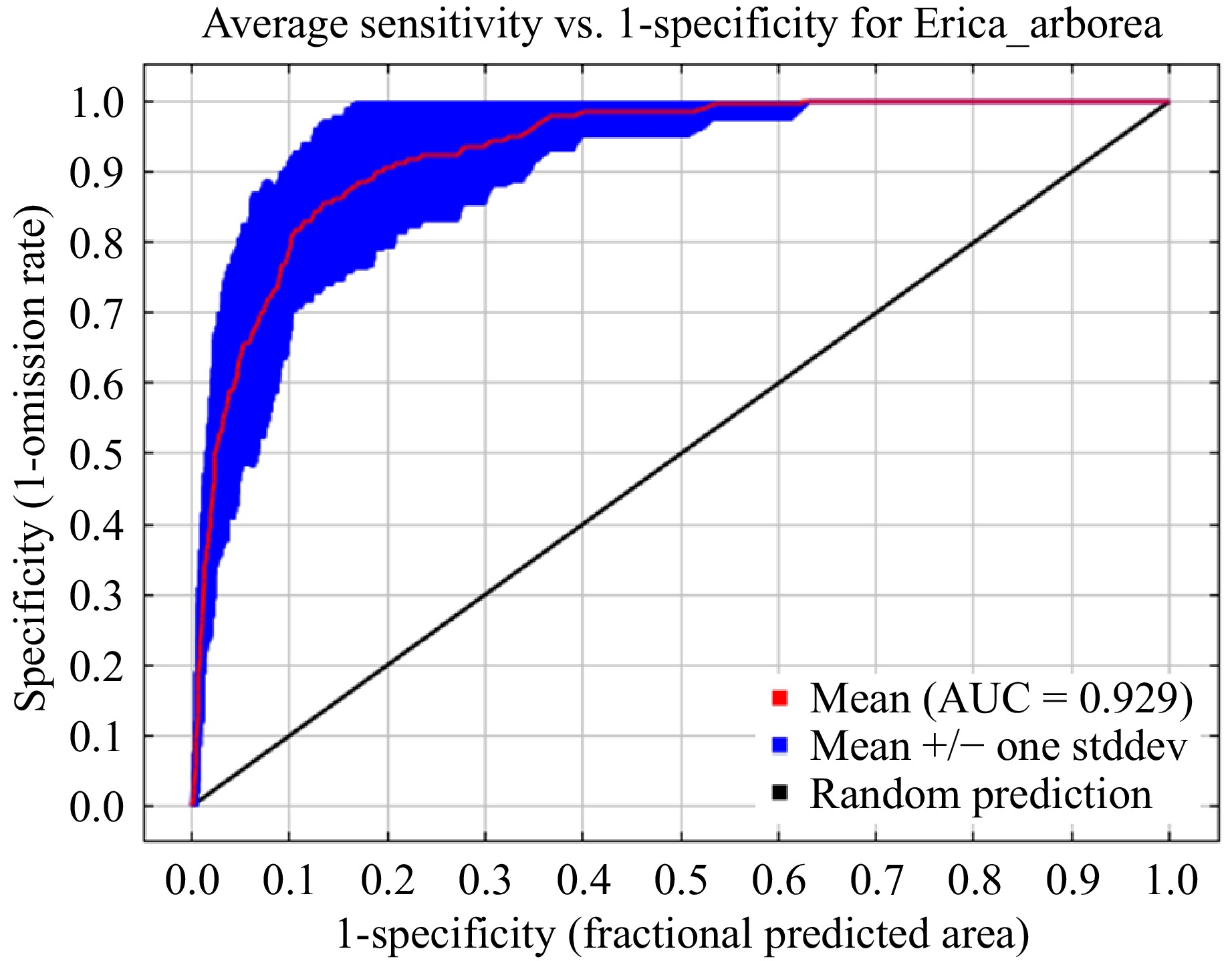
Figure 3.
Model performance ROC curve of the reconstructed Maxent model under current climatic simulation.
Variable importance and contribution
-
The three most important variables for predicting the current potential distribution of Erica, with more than 86% contribution to the model, were altitude, precipitation of the driest quarter and distance from the sea (Fig. 4 & Table 2). Under RCP4.5 in 2050, the variables altitude (56.9%), precipitation of the driest quarter (13.5%), temperature annual range (10.4) and precipitation of the driest month (7.4%) contributed 88.2% to the model. Under RCP8.5 in 2050, altitude (49%), distance from the sea (11.7%), temperature annual range (11.3%) and precipitation of the driest quarter (8.9) combined contributed 80.9% to the model. Under RCP4.5 in 2070, the variables altitude (56.1%), precipitation of the driest quarter (17.7%), precipitation of the warmest quarter (12.3%) and distance from the sea (6.4%) contributed 80.9% to the model. Under RCP8.5 in 2070, the variables altitude (43.7%), precipitation of coldest quarter (15.8%), precipitation of driest month (10.4%), distance from the sea (12.6%) and aspect (5.7%) contributed 88.2% to the model (Table 2).
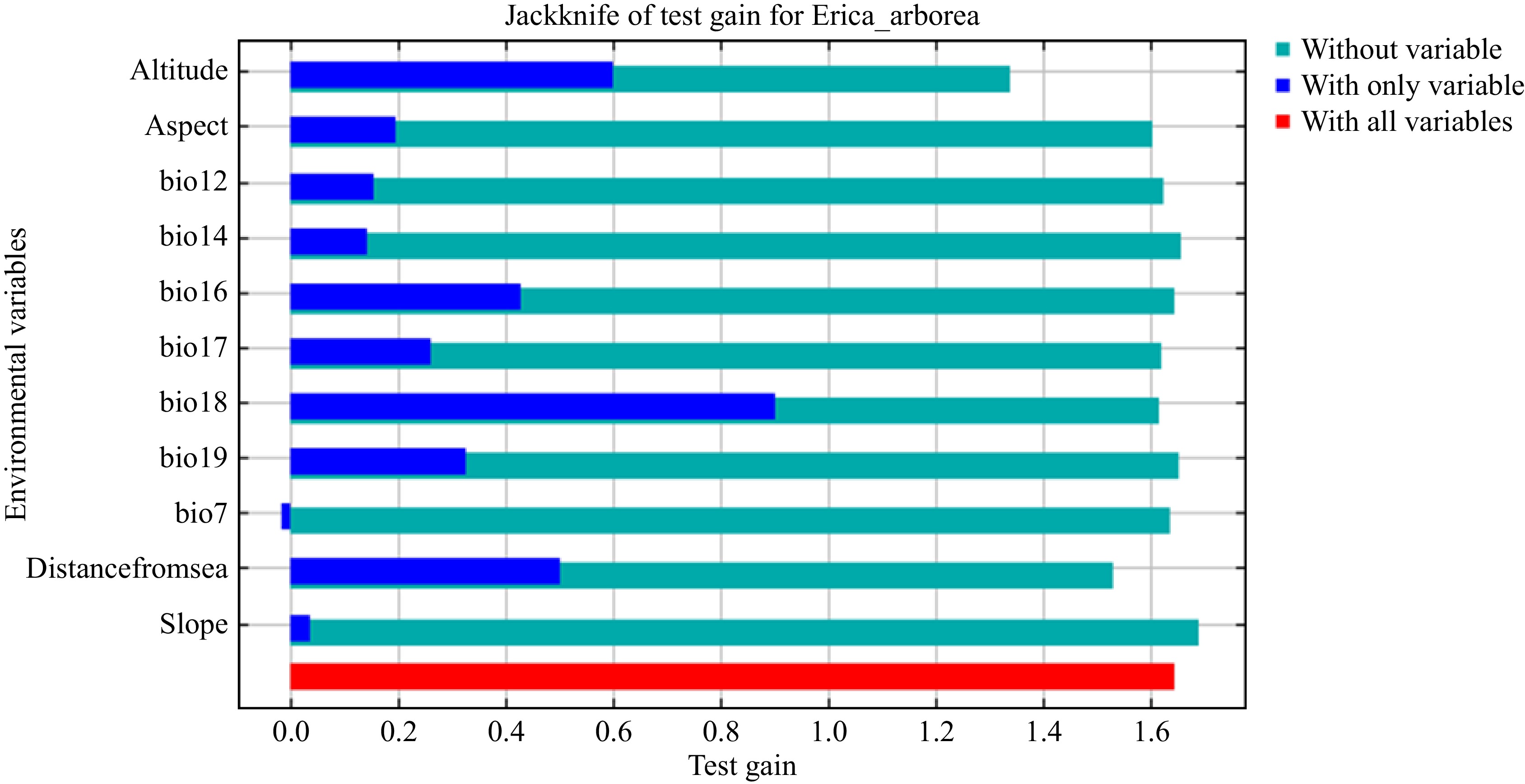
Figure 4.
Jackknife analysis of the importance of environmental variables in modelling the distribution of Erica.
Table 2. Estimates of relative contributions and permutation importance of the predictor environmental variables to the SDMs.
Variables Current Climatic scenario 2050 Climatic scenario2070 RCP4.5 RCP8.5 RCP4.5 RCP8.5 PC (%) PI (%) PC (%) PI (%) PC (%) PI (%) PC (%) PI (%) PC (%) PI (%) Altitude 29.6 45.2 27.3 56.9 32.2 49 33.8 56.1 29.1 43.7 Aspect 9.5 3.3 9.4 3.7 10.1 4.5 10.6 3.1 9 5.7 bio12 2.1 1.9 1.4 1.7 1.6 4 0.5 1 1.7 4 bio14 0.2 0.7 1.2 7.4 0 0.6 0.8 0.9 1.3 10.4 bio16 5.8 0.1 6.7 0.2 3.1 0.1 3.1 0 5.7 1.3 bio17 7.7 33.1 3.7 13.5 2.1 8.9 6.8 17.7 0.1 1 bio18 28.8 3.1 1.8 0.5 22.2 5.7 25.6 12.3 15.8 1.8 bio19 0.5 3 1.2 0.8 1.4 2 1.5 0.2 19.8 15.8 bio7 0.6 0.1 13.2 10.4 12.4 11.3 0.8 0.6 4.3 0.5 Distance 13.7 8.3 1 1.2 12.9 11.7 14.6 6.4 11.3 12.6 Slope 1.5 1.2 33.1 3.7 1.9 2.3 2.1 1.7 1.8 3 PC – Percent Contribution, PI – Permutation Importance, Distance – Distance from the sea. Topography-related variables followed by the precipitation-related variables had the highest (52.8% and 34.6%, respectively) contribution to the model in predicting the current distribution of Erica. Among the topographic variables, altitude (29.6%) contributed the most to the model while precipitation of the warmest quarter (28.8%) contributed the most among the precipitation variables. The temperature-related variable, temperature annual range, contributed the least to the model. The environmental variable with the highest gain when used in isolation is the precipitation of the warmest quarter (Fig. 4), which therefore appears to have the most useful information by itself. The environmental variable that decreases the gain the most when it is omitted is altitude (Fig. 4), which therefore appears to have the most information that is not present (i.e. temperature) in the other variables.
Response of Erica to environmental variables
-
Suitable habitats of Erica increase with an increase in altitude. Suitable altitude for the species was in the range of 2,357–3,750 m with optimal altitude at 3,750 m while habitats with an altitude of less than 2,357 m are unsuitable for Erica. Moreover, both sloppy (10%–70%) and flat (0–10%) and North and northwest-facing areas are suitable habitats for Erica with Northeast facing optimally suitable habitats. Habitats at a distance of 55.5–288.6 km from the sea with the best at 55.5 km are suitable habitats for Erica. Erica revealed habitat suitability inversely proportional to the increase of the temperature annual range, indicating a high sensitivity to large fluctuations in temperature.
Suitable habitats of Erica also increase with increasing annual precipitation. Optimal precipitations were at 600 mm (550–600 mm) for areas around Desa'a and Asimba and at 950 (850–950 mm) for areas in south Tigrai (Tabotat, Michael Abeda, Alaje, and Ofla). Areas with precipitation (3–29 mm) in the driest month, precipitation (200–620 mm) in the wettest quarter, precipitation (16–140 mm) in the driest quarter, precipitation (120–360 mm) in the warmest quarter and precipitation (15–250 mm) in the coldest quarter are suitable habitats for Erica. Habitats with precipitation in the driest month (< 3 mm), precipitation of the driest quarter (< 16 mm) and precipitation (< 120 mm) in the warmest quarter and precipitation (< 15 mm) in the coldest quarter are unsuitable habitats for Erica.
Predicted current potential distribution of Erica
-
The total potential suitable habitat for Erica in Tigrai is 2,097 km2, accounting for 9.12% of Tigrai's highland area (Table 3 & Fig. 5), as Erica is supposed to grow at > 1,800 m. Its EOO is 714 km2. In addition, 21.22% (445 km2) of the total area was optimally suitable habitat (> 0.6), 23.80% (499 km2) was suitable (0.4–0.6) and 54.98% (1,153 km2) was a low suitability (0.2−0.4) habitat. The unsuitable habitat covers 12,855 km2, accounting for 55.97% of the highland area. The exceptionally suitable area is confined to the highlands of Tsegede, Welqait, Irob, Ganta Afeshum, Atsibi, Ofla, and Emba Alaje while the highly suitable areas are found in the districts Tsegede, Welqait, Irob, Subha Saesie, Tsaeda Emba, Atsibi, Tsirae Wenberta, and Ofla districts in Tigrai, Ethiopia. Suitable habitats are found in Tsegede, Irob Subha Saesie, Tsaeda Emba, Atsibi, Tsirae Wenberta, and Ofla. The less suitable habitat covers a large area and is found in the Welqait, Tsegede, Awra, Irob, Gulo Mekeda, Ganta Afeshum, Subha Saesie, Tsaeda Emba, Atsibi, Tsirae Wenberta, Ofla, Raya Alamata, Raya Azebo, and Emba Alaje districts.
Table 3. Predicted changes (km2) in Erica ranges in the 2050s and 2070s under two climatic scenarios (RCP4.5 and RCP8.5) compared with the potential current distribution.
Suitability category Current Future scenario 2050 Future scenario 2070 RCP4.5 RCP8.5 RCP4.5 RCP8.5 Unsuitable 12855 13327 (+472) 13978 (+1123) 11909 (−946) 12473 (−382) Low suitable 1153 1321 (+158) 838 (−315) 1166 (+13) 420 (−733) Suitable 499 588 (+89) 490 (−9) 513 (+14) 171 (−328) Highly suitable 267 315 (+48) 274 (+7) 308 (+41) 142 (−125) Exceptionally suitable 178 186 (+8) 286 (108) 112 (−66) 265 (87) (+) refers to gain and (−) refers to loss in range areas (km2). 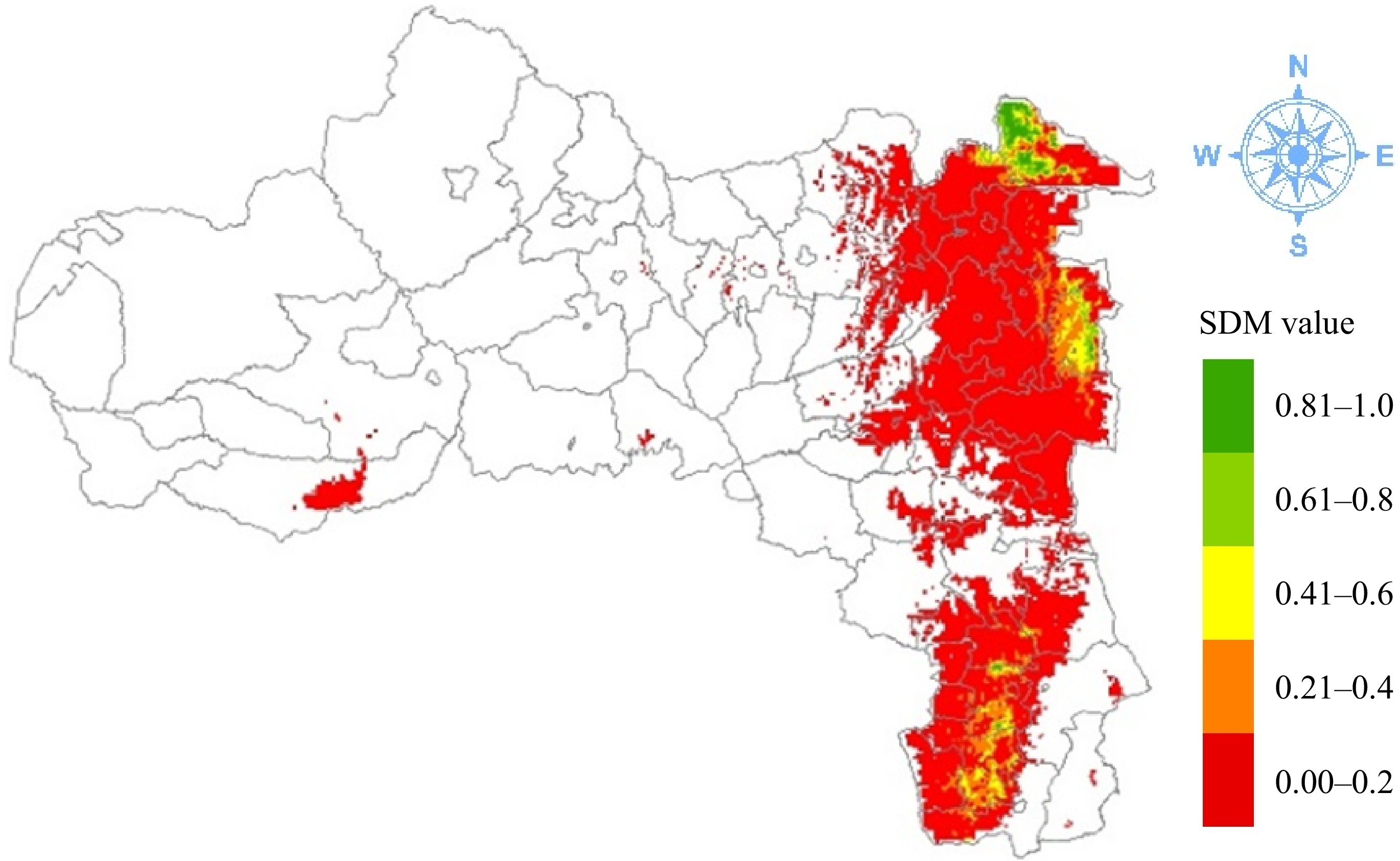
Figure 5.
Predicted current potential distribution of Erica in Tigrai.
Predicted future potential distribution of Erica
-
The potential distribution of Erica under future climate change scenarios revealed various patterns (Fig. 6). The predicted future potential distribution of Erica showed that there was a total gain in both the suitable (+303 km2) and unsuitable (+472 km2) habitats (Table 3 & Fig. 6) under RCP4.5 in 2050 when compared to the current distribution. Under RCP8.5 in 2050, there would be an increase in unsuitable habitat (+1,123 km2), accompanied by a decrease in suitable habitat (–209 km2). Under RCP4.5 and RCP8.5 by 2070, the unsuitable habitat would increase by 946 and 382 km2 respectively, while the suitable habitat would increase by only 0.09% (2 km2) for RCP4.5 and shrink by 1,099 km2 under RCP8.5. In addition, Erica has EOO values of 418, 1,273, 73, and 913 km2 under RCP4.5 in 2050, RCP8.5 in 2050, RCP4.5 in 2070, and RCP8.5 in 2070, respectively.
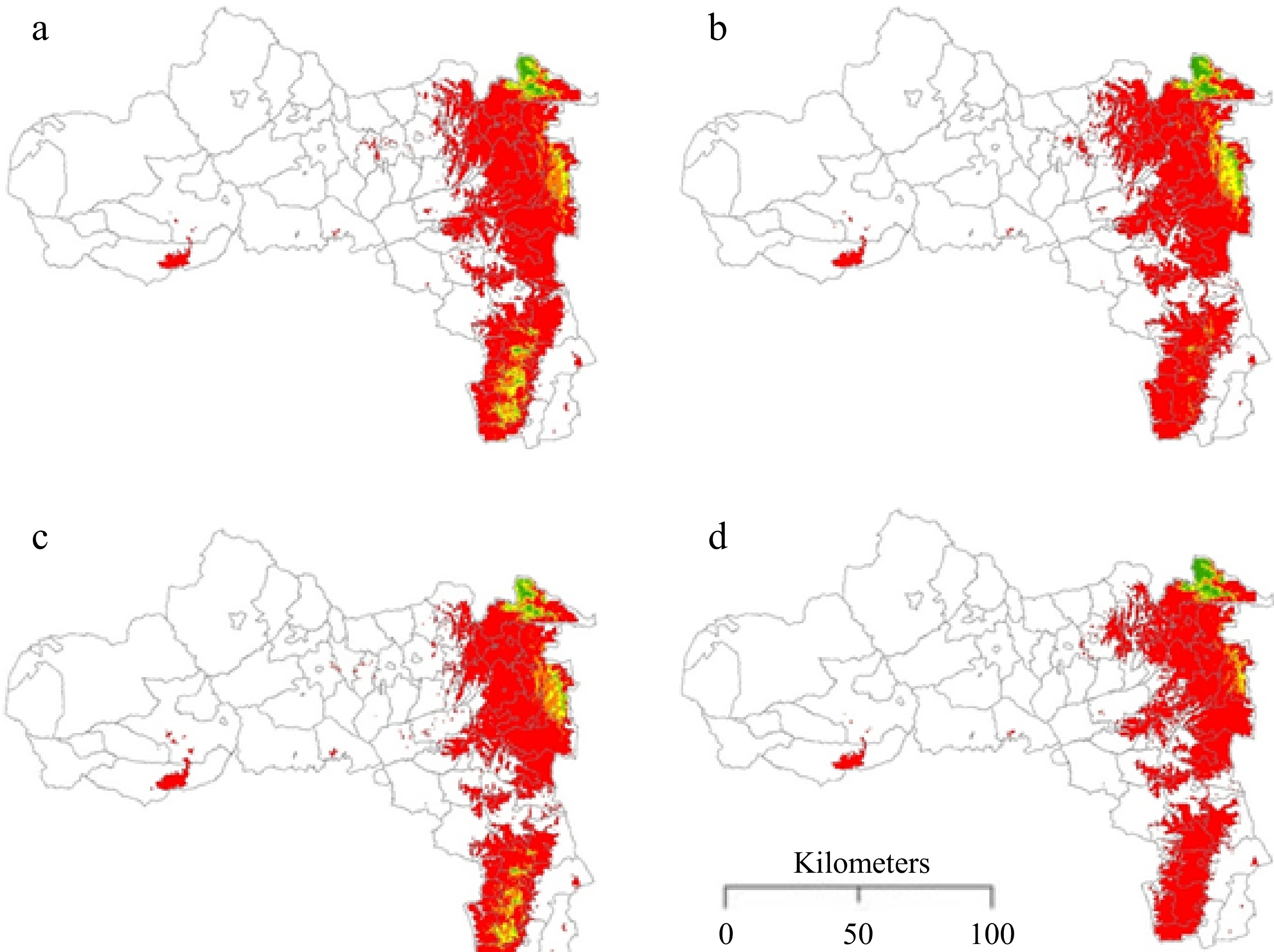
Figure 6.
Predicted future average potential distribution model of Erica (a) 2050 RCP4.5, (b) 2050 RCP8.5, (c) 2070 RCP4.5, and (d) 2070 RCP8.5.
-
The accuracy of the Maxent model outputs were excellent with high (AUC > 0.9 for both training and testing) discriminative ability, except under RCP8.5 in 2070 with good performance, with an AUC of 0.95 for training and AUC of 0.88 for testing. Models with an AUC value above 0.90 are considered to have an excellent performance[9,48] while with an AUC value of 0.8-0.9 is indicative of the high precision[52] and, thus, are considered promising[9,44,52]. Moreover, the present results are comparable to various studies carried out in the region using the Maxent modelling approach[22,42,53−55], and other SDM approaches[23,39,56]. The present simulation results are reliable in terms of model evaluation and thus could be used to simulate other similar highland species.
Species distribution models have proven valuable in filling gaps in the knowledge of species occurrences[57]. Many studies have demonstrated the usefulness of SDMs in determining ideal locations for species reintroduction and relocation[23,39,56,57]. However, there are certain limits to using SDMs, particularly when available data is insufficient for the modelling process to be effective[58]. Other critical shortcomings include sample size, positional uncertainty, sampling bias, and the species ecology[57]. For instance, including solely bioclimatic factors may lead to bias in the results because other factors such as human activities and dispersion restrictions also play important roles in determining such species' future ranges[58]. In this study, to improve the accuracy of the model, additional environmental variables have been added on top of the usual bioclimatic variables.
Response of Erica to environmental variables
-
Environmental factors and climate change have a significant impact on how species disperse, migrate, evolve, adapt, and go extinct[2]. Topographic and bioclimatic variables influenced the distribution range and survival of Erica (Table 2, Fig. 6). The six most influential variables with more than 95% cumulative contribution to the model were altitude, distance from the sea, aspect, precipitation of the warmest quarter, precipitation in the driest quarter, and precipitation in the wettest quarter. The importance of these variables suggests that the association of Erica with cold temperature (i.e. high altitude) and rainfall[12−14,21,29,30,59] and its sensitivity to temperature fluctuations (bio7)[1,12,14,21]. The present findings are in line with earlier studies on the species. In a study on tree ring chronology, Jacob et al., found a statistically significant correlation (p < 0.05) between the growth of Erica and the minimum temperature in March and August and the correlation was able to explain 24% of the variation in tree growth. According to these authors, the minimum temperature in the growing season (August) controlled tree growth and warmer temperatures enhanced the growth of Erica. It was further elaborated that rainfall impeded tree growth under wet conditions and enhanced tree growth under dry conditions[21].
Altitude is an important environmental factor affecting the soil, vegetation, and microclimate of forests, the growth and distribution of plants[60]. Altitude along with aspect and slope are the three main topographic factors that control the distribution and patterns of species in mountain areas of East Africa[39,61]. Altitude was strongly correlated with temperature, and the distribution of Erica was strongly influenced by altitude (Fig. 6). This result reinforces the findings of other species distribution modelling carried out on other plant species on flora[4,11]. In Tigrai, marked variation in altitude results in a distinct spatial distribution of temperature and rainfall[62]. Aspect controls species distribution by inducing local variations in the temperature and precipitation of mountainous topographies[63] through effects on hydrological and solar energy regimes[61,63]. In the present study, Northeast-facing habitats were found to be optimally suitable for Erica presumably due to cooler and wetter conditions in these areas.
Precipitation is the most decisive factor controlling species distribution in the drylands[34,61]. It is also assumed that the dry Afromontane forests trap moisture from the clouds or mist, which usually builds up along the escarpments[36]. Thus, distance from the sea is another factor influencing the distribution of Erica through its influence on the coverage of fog, clouds, and mist, from the Red Sea coast to the North and the Indian Ocean coast to the East, which provides moisture to the mountainous area[34,36] where Erica grows. Moisture, held in cloud droplets form/fog and drifts inland with the prevailing wind inferring the importance of distance from the sea and aspect.
Predicted current potential distribution of Erica
-
The current distribution of Erica and those simulated with the SDMs and with expert knowledge using ArcGIS are consistent (Figs 6 & 7). With the SDM and ArcGIS, additional suitable habitats for Erica were found in the highlands of Tsegede, Welqait, Tselemti, Ganta Afeshum, and Dogu'a Tembien, these habitats need verification. In Tsegede, Welqait, and Tselemti, Erica is expected to be effectively present as these districts are part of the Semien Mountain massif where Erica forms the Erica treeline ecotone[12,21]. In Dogu'a Tembien, a small remnant Erica forest occupies the northern slopes near the top of the Dabba Selama Mountains[64]. The optimally suitable habitats were found around Asimba, Aiga, Desa'a, Emba Alaje, Endamokhoni, and Ofla.
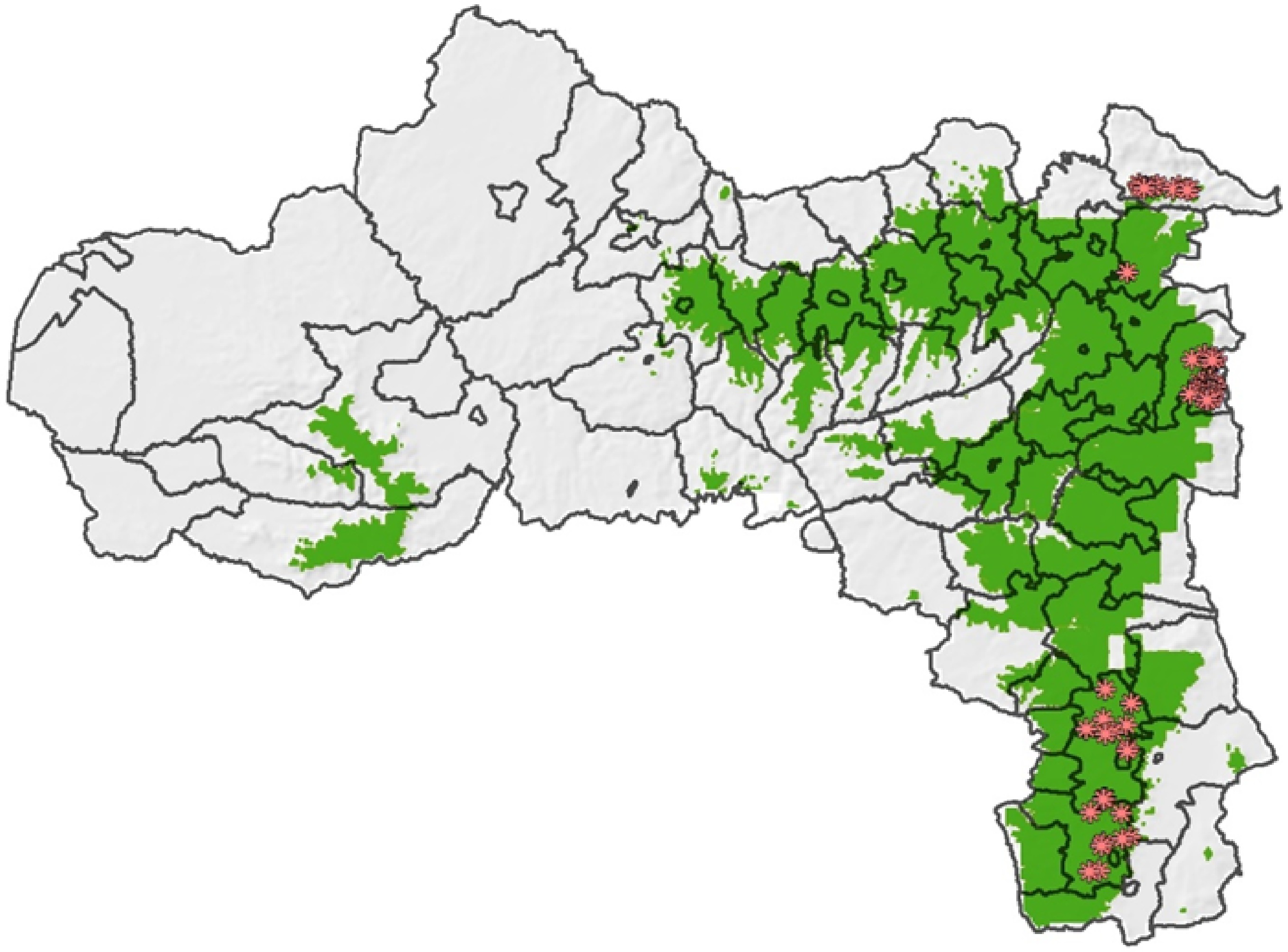
Figure 7.
Suitability model of Erica predicted based on expert knowledge using five environmental variables and the current occurrence points. The green shaded area represents possible suitable areas for the growth of Erica. The red dots represent the actual presence data of Erica.
Predicted future potential distribution of Erica
-
Climate change-induced warming is expected to impact plant diversity, abundance, and distribution[1]. Compared to the extent of suitable habitat under current conditions, the distribution range of Erica showed a tendency to shrink under future climatic scenarios, under RCP8.5 both in 2050 and in 2070 (Fig. 6 & Table 3). Specifically, under RCP8.5 the range will shrink by 209 km2 in 2050 and by 1,099 km2 in 2070. The large range reduction (up to 52.41%) in RCP8.5 suggests that climate change might threaten Erica. In contrast, under RCP4.5, our SDMs show that the range of Erica will expand by 303 km2 in 2050 and 2 km2 in 2070. Much of the range expansion will occur in the eastern and western zones of Tigrai, while the range contraction will occur in the southern zones of Tigrai. Taking the 0.2 logistic threshold as the cut-off point, we found evidence of an altitudinal range shift (from 1,800 to 2,357 m) but we did not find evidence indicating longitudinal or latitudinal range shift. An altitudinal range shift supports the hypothesis that 'climate change is expected to result in altitudinal range shift and range contraction in East Africa'[1] while the absence of latitudinal and longitudinal range shifts support the hypothesis 'Nowhere to go'[28].
Conservation intervention
-
Erica is facing multiple threats including inherent problems, anthropogenic, and climatic. It has a narrow range, the Erica ecotone. The seeds are tiny and difficult to collect. Its natural regeneration is low, and it is hampered by grazing, trampling, and poor precipitation. Seedlings are less successful[15] at establishment. Moreover, the species is not among the priority species for reforestation and afforestation activities. The species is highly logged for charcoal and fuelwood. The habitat where it grows is under constant pressure from increasing population, free grazing, illegal logging, charcoal making, and habitat conversion[53,65]. More importantly, its distribution will be reduced due to the impacts of climate change; putting it at the risk of extinction. Hence, given the drastic population decline and local extinctions, in situ and ex situ conservation of the species demands priority.
For its in-situ conservation, habitat protection, and restoration should be prioritized because suitable habitats are prerequisites for species survival. Habitat restoration includes reintroduction of the species, enrichment planting, enclosing degraded lands to enhance natural regeneration, participatory soil and water conservation, watering, fencing, soil amendments, and awareness creation. In addition, sustainable energy sources and improved stoves should be introduced. For its ex situ conservation, the establishment of botanical gardens, introducing planting in urban landscaping, and use of biotechnology (micropropagation) are important. In numerous countries, also out of the strict territory of the plant (say, The Netherlands), Erica is grown as an ornamental plant[14]. Promoting it as an ornamental plant in private and public gardens in Tigray that fit with the environmental requirements, particularly those that are at least at higher elevations, would further support conservation of the species ex situ while creating job opportunities and generating income for gardeners. Erica is also a good source of nectar for bees and hence an opportunity to plant it would be appreciated in Tigray's home garden agroforestry, or as a benefit from public plantations.
-
The spatiotemporal distribution pattern and suitable habitat of Erica has been succesfully modelled and analysed with SDMs and ArcGIS. Highly suitable habitats were found in the districts of Irob (Aiga and Asimba Mountains), Atsibi (Desa'a), Emba Alaje (Wujig Mahgo Waren), Endamokhoni, and Ofla (Hugumburda) and in the highlands of Tsegede. These districts are areas with stable cold climates where Erica could form the Erica treeline ecotone. Altitude (29.6%), precipitation in the warmest quarter (28.8%), distance from the sea (13.7%), aspect (9.5%), precipitation of the driest quarter (7.7%), and precipitation in the wettest quarter (5.8) were the main environmental variables shaping the distribution and suitable habitat for Erica. The suitable habitat range of Erica will shrink due to climate change. Hence, ex situ and in situ conservation of the plant is crucial.
The authors acknowledge WeForest for provision of occurrence data and the Rufford Foundation for financial support (Code: 10f54f1).
-
All authors have participated in the conception and design of the research, the formal analysis and interpretation of the data; drafting the article and revising it critically for important content and approve the final version of the manuscript for publication.
-
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0024-0045)
-
Received 8 June 2024; Accepted 24 October 2024; Published online 16 January 2025
-
Tree heath is a rare plant species facing a drastic population decline.
Maxent performance was excellent (AUC > 0.9) in predicting the potential distribution of tree heath.
The current potential area coincides with that modeled by Maxent and ArcGIS.
The distribution of E. arborea was highly associated with cold temperature (i.e. high altitude) and rainfall.
Under RCP8.5 in 2070, the suitable habitat range of Erica will shrink by 54%.
- Supplementary Table S1 Pearson correlation coefficients among the predictor environmental variable.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Berhe DH, Tefera AS, Gidey T, Retta AN, Negussie A, et al. 2025. The potential distribution of tree heath (Erica arborea L.) in Tigrai. Tropical Plants 4: e002 doi: 10.48130/tp-0024-0045 

The potential distribution of tree heath (Erica arborea L.) in Tigrai
- Received: 08 June 2024
- Revised: 22 October 2024
- Accepted: 24 October 2024
- Published online: 16 January 2025
Abstract: Tree heath (Erica arborea L.) is a rare and endangered native tree species that is facing an incessant population decline due to climatic and anthropogenic factors. The objective of this study was to analyse the impacts of future climate change on the potential distribution of the species in the alpine areas of Tigrai (Ethiopia). A species distribution modelling using Maxent was employed to predict the potentially suitable habitat for the species under two future climatic scenarios. The model was constructed using 58 sets of presence data collected from five main forest areas of the region and 11 environmental predictors. The results revealed that altitude, precipitation in the warmest quarter, distance from the sea, aspect, precipitation in the driest quarter, and precipitation in the wettest quarter were the main factors influencing the distribution of E. arborea. The distribution of E. arborea was highly associated with cold temperature (high elevation) and rainfall. E. arborea is sensitive to temperature fluctuations (bio7). The current suitable habitats of E. arborea were restricted within 2,097 km2. The highly suitable habitats for future distribution were the Irob, Atsibi, Emba Alaje, Endamokhoni, Ofla, and the Tsegede highlands of Tigrai. Under RCP8.5 in 2070, the suitable habitat range of Erica will shrink by 54%. Therefore, in situ and ex situ conservation interventions should be introduced to save the rare species in the Tigrai highlands.