









-
Leaves are the principal organs of photosynthesis and transpiration in plants. Their size and morphological traits directly affect photosynthetic efficiency and nutrient distribution, thereby playing a critical role in determining crop yield[1]. Among horticultural crops, leaf length, width, and area are key agronomic traits, as approximately 95% of the dry matter in these crops originates from leaf photosynthesis[2]. The leaf exhibits three axes of polarity: basal–apical, medial–lateral, and proximal–distal, across which cell morphology and size vary significantly. During plant growth and development, multiple genes act in coordination along these axes to ultimately shape distinct leaf forms[3]. Both leaf length and width are quantitative traits regulated by various hormones, including auxin, gibberellin, and methyl jasmonate. For instance, Chen et al.[1] found that the R2R3-MYB transcription factor CsRAXs modulates cucumber leaf size through the auxin pathway and also influences fruit set. In tomato, leaf morphology is a fundamental component of plant architecture, with leaf length and width directly affecting planting density and yield potential[4]. Therefore, elucidating the genes and molecular mechanisms underlying leaf size regulation is of great significance for optimizing pepper plant architecture and improving crop productivity. The LBD (lateral organ boundaries domain) transcription factor family, also known as the ASL (asymmetric leaves 2-like) gene family, constitutes a group of transcription factors unique to plants[5,6]. Members of this family contain a conserved LOB domain at their N-terminus and are typically divided into two classes—Class I, and Class II—based on structural differences in their sequences[7]. Class I proteins feature a highly conserved zinc-finger-like motif (CX2CX6CX3C, where C represents conserved cysteine residues and X denotes non-conserved amino acids), that enables specific binding to the GCGGCG DNA sequence. They also contain a glycine-alanine-serine (GAS) region, which modulates DNA-binding activity, as well as a leucine-zipper motif (LX6LX3LX6L) involved in protein dimerization. Additionally, a conserved proline residue at the C-terminus is thought to play a key functional role in LBD proteins[8−10]. In contrast, Class II members possess only a single conserved zinc-finger-like motif[7]. LBD transcription factors were first identified in Arabidopsis. Since then, genome-wide analyses have revealed 44, 43, 35, and 46 LBD proteins containing the LOB domain in maize[11], Arabidopsis[12], rice[13], and tomato[14], respectively.
LBD proteins exhibit considerable functional diversity in regulating plant organ development and metabolism. In Arabidopsis, for example, Class Ia member AtLBD12, Class Ib member AtLBD36 (AtASL1), and Class Id member AtLBD28 contribute to leaf development and polarity establishment[15,16]. Class Ic members AtLBD16 and AtLBD18 are involved in auxin signaling[17,18], while AtLBD18 and AtLBD29 further regulates lateral root emergence and formation[19,20]. Overexpression of the Class Ib member AtAS2 can induce alterations in leaf morphology[21]. The Class Ia member AtLBD11 promotes meristem cell proliferation by activating transcription of ROS metabolism genes, such as peroxidase 71 and respiratory burst oxidase homolog D/F, thereby triggering maximal ROS production in the meristem region[22]. Meanwhile, Class IIb member AtLBD41 is involved in controlling the specialized differentiation of apical meristem cells; its ectopic expression in Celosia leads to leaf curling[12,23]. Beyond Arabidopsis, genes such as MdLOB11 in apple, LjLOB1 and LjLOB3 in Lycopodium, and OsAS2 in rice have also been shown to regulate leaf growth and development[24−26]. Moreover, LBD transcription factors are implicated in a wide range of processes, including pollen development, plant regeneration, stress responses, photomorphogenesis, secondary growth, and metabolic regulation. In tomato, for instance, SlLBD1 and SlLBD2 display light-dependent expression patterns and influence plant architecture through transcriptional networks[27]. In Arabidopsis, Class IIa members LBD37, LBD38, and LBD39 act as central repressors that integrate nitrogen signaling with the anthocyanin biosynthesis pathway, thereby coordinating primary and secondary metabolism[28]. Similarly, ZmMS1/ZmLBD30, a recently identified repressor in maize, is essential for normal development of the pollen exine[29].
The LBD function is closely related to hormones. For instance, SlLBD40 contributes to tomato drought tolerance through jasmonic acid signaling, while ectopic expression of Gossypium GhLBD41 enhances disease resistance in Arabidopsis by promoting JA accumulation and activating JA-mediated defense responses[14,30]. A dual regulatory mechanism involving 'transcriptional regulation and protein interaction' has been observed in ginseng to optimize root plasticity synergistically. Specifically, PgLBD18 regulates lateral root development via auxin-dependent pathways, whereas PgLBD49 functions independently of auxin. PgLBD49 forms heterodimers with PgARF to stabilize the transcriptional complex, thereby enhancing downstream gene expression, and promoting lateral root development[31].
The genus Capsicum includes 43 species and thousands of cultivars, among which substantial variations in leaf size and morphology have arisen due to long-term selection. Nevertheless, the physiological, cellular, and molecular mechanisms underlying these differences remain poorly understood. Based on LBD genes previously identified through GWAS as being associated with leaf length, this study further examines expression changes of LBD36 under exogenous hormone treatments and explores the effects of its silencing on leaf size, photosynthetic parameters, endogenous hormone levels, and cellular morphology. This work aims to clarify the potential regulatory roles of LBD36, offering a theoretical foundation for the targeted improvement and efficient breeding of high-yield, high-quality pepper varieties.
-
The experiment utilized 182 chili pepper germplasm accessions supplied by the Chili Pepper Research Group of Hainan University. This collection included 107 wild chili pepper (Capsicum frutescens) accessions collected from various cities and counties across Hainan Province, along with 75 cultivated varieties of C. annuum originating from other provinces. All materials were sown in September 2022 at the experimental base in Sanjia Town, Dongfang City, Hainan Province. A completely randomized design was adopted, with 10 plants cultivated per accession. Uniform cultivation and management practices were applied across all accessions. Individual plants from each accession were labeled and sampled, and these samples served as the materials for subsequent genome-wide association study (GWAS) of chili pepper leaf length.
In addition, large-leaf varieties (BD-HDL, Cc268, and ZY5), and small-leaf varieties (45-E, 37-Y, and 65-Y) were planted at the experimental base of Qionghai Zhongyuan Soil and Fertilizer Station. Each variety was arranged in three replicates, with labeled single-plant samples collected from each replicate for further analysis. Three biological replicates were set for each cultivar, and samples were collected from individually tagged plants for each material.
Measurement and statistical analysis of leaf phenotypes
-
For the expression analysis of the LBD36 gene, leaves of uniform age were collected from the middle section of selected plants. From each line, three plants were randomly sampled, and leaf measurements were performed using ImageJ software. Data processing and statistical analyses were conducted with Excel 2016 and GraphPad Prism 9.
Genome-wide association study
-
Using the Capsicum annuum 'Zunla-1' genome (NCBI accession: GCF_000710875.1) as the reference, high-quality single nucleotide polymorphism (SNP) loci were identified. These SNP data were integrated with the leaf length phenotypic data, and a genome-wide association study (GWAS) was performed based on the fastlmm model.
Gene cloning and bioinformatics analysis
-
Following the genome-wide association study, LBD36 was identified as a candidate gene involved in the regulation of pepper leaf length. Total RNA was extracted from plant tissues using the Novazene RNA Extraction Kit, and first-strand cDNA was synthesized using the Novazene Reverse Transcription Kit. Based on the reference sequence of LBD36, full-length gene-specific primers were designed (Supplementary Table S1) and used for PCR amplification.
The genome sequence and annotation file of Capsicum chinense was retrieved from the NCBI database (
www.ncbi.nlm.nih.gov ). Using the hmmer 3.0 software and the LBD family domain (PF03195) as a query, a genome-wide homology search was performed. Candidate genes were initially screened with an E-value threshold of < 0.00001. Corresponding protein sequences were then extracted and further verified through the SMART and Pfam databases to confirm the presence of a complete LBD domain. Sequences lacking a complete LBD domain were discarded.Chromosomal localization of the CcLBD family was analyzed with TBtools. Putative cis-acting elements in promoter regions were predicted using PlantCare. A phylogenetic tree was constructed with MEGA 12, and visualized using ITOL.
Exogenous hormone treatment
-
BD-HDL pepper seedlings were treated with exogenous hormone solutions, including 100 μmol/L ABA, MeJA, and GA3, as well as 50 μmol/L IAA. After 24 h of treatment, leaves at the 40-day-old stage were collected, and the expression of the LBD36 gene under different hormone treatments was analyzed using real-time quantitative PCR (qPCR). Three biological replicates were performed for each experiment.
Expression analysis
-
Based on phenotypic results, leaves of comparable developmental stages were collected from different pepper varieties. cDNA was synthesized from leaf samples of the BD-HDL cultivar harvested at 12, 19, 26, and 33 d post-treatment, as well as from hormone-treated leaves. qPCR primers specific to LBD36 were designed as listed in Supplementary Table S1. Quantitative real-time PCR was conducted using the SYBR Green I fluorescent dye method. Each sample was analyzed with at least three biological replicates and three technical replicates. Gene expression levels were calculated by the 2−ΔΔCᴛ method using Excel 2016, and final data visualization was performed with GraphPad Prism 9. Three biological replicates with three technical replicates were performed for each experiment.
Construction of silent expression vectors
-
Using cDNA from the BD-HDL cultivar as template, an interference fragment was amplified by PCR. The fragment was then inserted into the silencing expression vector pTRV2 via homologous recombination. After heat-shock transformation into E. coli, positive recombinant plasmids were screened and extracted. These plasmids were subsequently introduced into Agrobacterium tumefaciens competent cells by heat-shock transformation for use in plant genetic transformation. Using BD-HDL as the recipient material and following the genetic transformation system established by Zhang[32], transgenic pepper plants carrying the pTRV2-LBD36 silencing construct were obtained.
Paraffin sectioning
-
Based on the effective duration of VIGS, leaves of 40-day-old BD-HDL pepper plants from both wild-type and silenced lines were selected for paraffin sectioning to observe differences compared with the control. Three independent biological replicates were performed. For each paraffin section, three non-overlapping fields of view were randomly selected, and leaf cell images were captured using NDP.view2 software. All images were uniformly magnified to 40× for cell morphological observation. Cells in each field were counted, and the average cell area per field was calculated using ImageJ software for statistical analysis.
Determination of chlorophyll content
-
Chlorophyll contents were determined using the Suzhou Grace G0601F kit. Functional leaves of 40-day-old pepper plants were sampled. After removing the midrib, 0.1 g of shredded leaf tissue was added to 1 mL of 95% ethanol-distilled water extraction buffer and 50 mg of Reagent 1. The mixture was ground in the dark, and the volume was adjusted to 10 mL, followed by dark incubation until the residue turned white. A 1-mL aliquot of the extract was used, with extraction buffer as the blank control. Absorbance at 665 and 649 nm was measured using a spectrophotometer. The contents of chlorophyll a, chlorophyll b, and total chlorophyll were calculated according to the kit's formula. Three independent biological replicates were performed for each experiment.
Hormone content analysis
-
Healthy, intact leaves from the middle section of pepper plants were collected as samples, with three biological replicates per group. Hormone analysis was performed by Wuhan Punes Testing Technology Co., Ltd. using liquid chromatography. Standard solutions of IAA and GA3 were prepared at a concentration of 500 µg/mL and serially diluted to a concentration range of 0.1−200 ng/mL, which included 20 ng/mL of deuterated internal standard, to generate a calibration curve. For sample preparation, 0.2 g of leaf tissue was homogenized with 2 mL of acetonitrile containing an internal standard, and extracted overnight at 4 °C. After centrifugation, the supernatant was collected, and the pellet was re-extracted. Combined supernatants were purified using a C18 solid-phase extraction column, concentrated, and redissolved in methanol prior to filtration. Chromatographic separation was achieved using a gradient elution of 0.1% formic acid in water and acetonitrile. Detection was performed in multiple reaction monitoring (MRM) mode with the following settings: curtain gas 35 psi, and ionization temperature 500 °C. Quantification was based on the established standard curve.
Photosynthetic parameter measurement
-
Leaf photosynthetic parameters, including net photosynthetic rate (Photo), stomatal conductance (Gs), intercellular CO2 concentration (Ci), and transpiration rate (Trmmol), were measured using an LI-6400XT portable photosynthesis system (Beijing Ligeta Technology Co., Ltd.). For different pepper varieties, measurements were taken from healthy leaves of the same nodal position at 80 d after planting. For gene-silenced plants, healthy leaves of the same nodal position at 40 d after treatment were selected. Three target leaves per plant were measured, with three replicate readings recorded for each leaf. Three target leaves were selected per plant, with three technical replicates per leaf.
-
Genomic resequencing and leaf length measurements were conducted on 182 natural pepper accessions to identify single nucleotide polymorphisms (SNPs) associated with leaf length. Using a significance threshold of 9.99 × 10−6, GWAS revealed 881 significant SNPs (Supplementary Fig. S1a). A quantile–quantile (QQ) plot showed that observed leaf length distribution in the natural population deviated from the expected distribution, indicating that leaf length variation is largely influenced by major-effect SNP mutations (Supplementary Fig. S1b).
Further candidate gene analysis was performed on reliable SNPs within associated genomic regions. Within a 100 kb interval centered on the most significant SNP, the candidate gene LBD36 was preliminarily identified as related to pepper leaf length, based on p-value and phenotypic contribution rate (Supplementary Table S2). Primers were designed based on the CcLBD36 reference sequence (NCBI accession: PHU25347.1). The CcLBD36 cDNA was successfully cloned from Hainan huangdenglong pepper. Its open reading frame (ORF) spans 900 bp and encodes a protein of 299 amino acids (Supplementary Fig. S2).
Identification, chromosomal distribution, and synteny analysis of the CcLBD gene family, and characterization of Its promoter cis-acting elements
-
A total of 39 LBD genes were identified in the C. chinense genome. Through Pfam and NCBI conserved domain searches, all 39 CcLBD proteins were confirmed to contain the conserved LOB domain. According to their chromosomal locations, these genes were systematically named from CcLBD1 to CcLBD39. Among them, CcLBD1 to CcLBD37 are distributed across 10 chromosomes (Chr1–6, 8, 9, 11, and 12), with Chr6 harboring the highest number of genes (11), while Chr5 and Chr9 each contain only one gene (Fig. 1a). Synteny analysis revealed five orthologous LBD gene pairs between C. chinense and A. thaliana, including CcLBD36/AtLOB and CcLBD35/AtLBD10 (Fig. 1b). Prediction of cis-acting elements in the promoter regions of CcLBD genes indicated the presence of multiple hormone- and light-responsive elements, such as auxin-, gibberellin-, and abscisic acid-related motifs (Fig. 1c). These results suggest that LBD36 transcription may be regulated by photoperiod or light quality, and could coordinate leaf growth in pepper through the integration of plant hormone signaling.
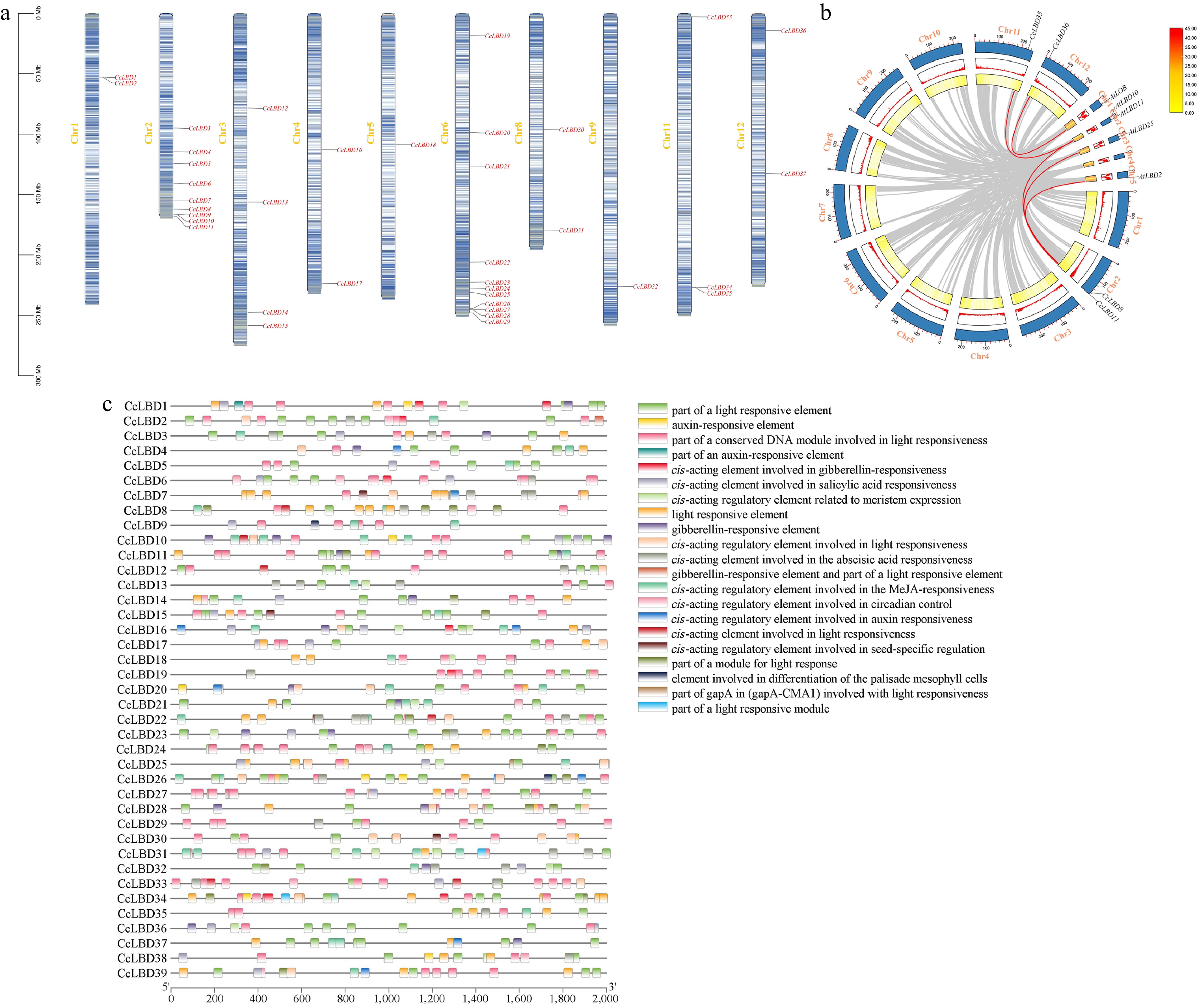
Figure 1.
Chromosomal localization of the LBD gene family and prediction of cis-acting elements in the promoter.
Phylogenetic analysis of the CcLBD36 sequences
-
A phylogenetic tree was constructed using the CcLBD family from pepper, along with LBD families from Arabidopsis and tomato to clarify their evolutionary relationships. The 39 pepper LBD genes were divided into two major classes: Class I and Class II. Class I was further classified into four subclasses (Ia, Ib, Ic, Id), while Class II comprised two subclasses (IIa, IIb). Within Class Ia, CcLBD18, 20, 25, and 35 clustered with SlLBD1-2, SlLBD4-1, AtLBD11, and AtLBD12. Notably, CcLBD36 grouped with SlLBD36-1 and AtLBD36 in Class Ib, suggesting its potential role in regulating pepper leaf size (Fig. 2).
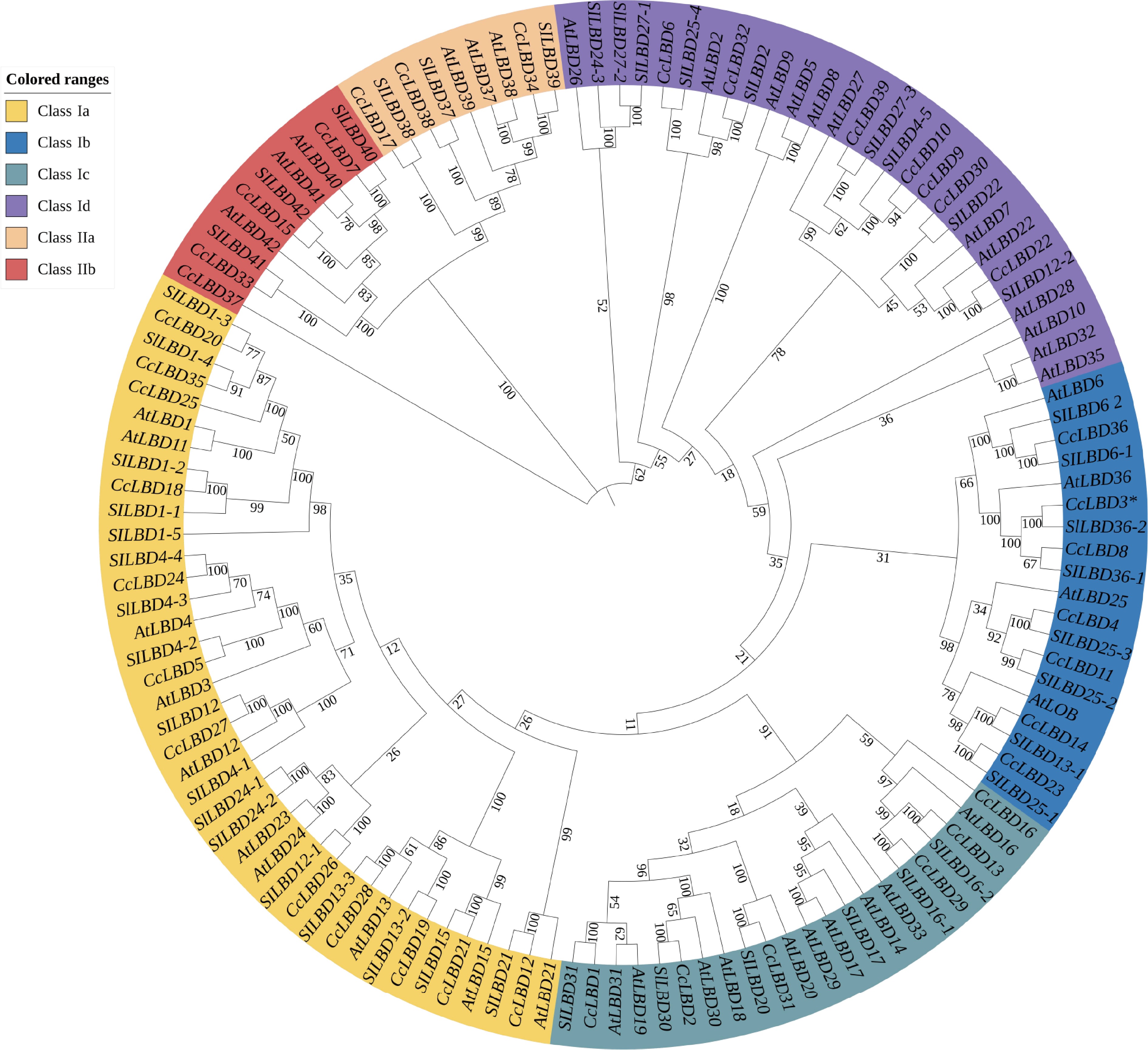
Figure 2.
Phylogenetic tree of the LBD gene family sequence source (Supplementary Table S3).
Analysis of the relationship between CcLBD36 expression patterns and leaf phenotypes and physiological indicators
-
To explore the potential role of CcLBD36 in leaf development, we systematically examined the correlation between its expression patterns and leaf phenotypic and physiological indicators. Phenotypic analysis revealed that leaf length, width, perimeter, and area were significantly greater in large-leaf varieties than in small-leaf varieties. Specifically, the largest leaf had a length of 17.189 cm, width of 9.038 cm, perimeter of 42.450 cm, and an area of 107.073 cm2, whereas the smallest leaf had a length of 2.280 cm, width of 1.069 cm, perimeter of 6.200 cm, and an area of 1.365 cm2 (Fig. 3a, b). Moreover, chlorophyll a, chlorophyll b, and total chlorophyll content were significantly higher in large-leaf varieties (Fig. 3c), which corresponded with a significantly elevated net photosynthetic rate. In contrast, no significant differences were observed in intercellular CO2 concentration, stomatal conductance, or transpiration rate between the two groups (Fig. 3d), suggesting that the enhanced photosynthetic capacity is likely driven mainly by increased mesophyll cell activity rather than stomatal regulation.
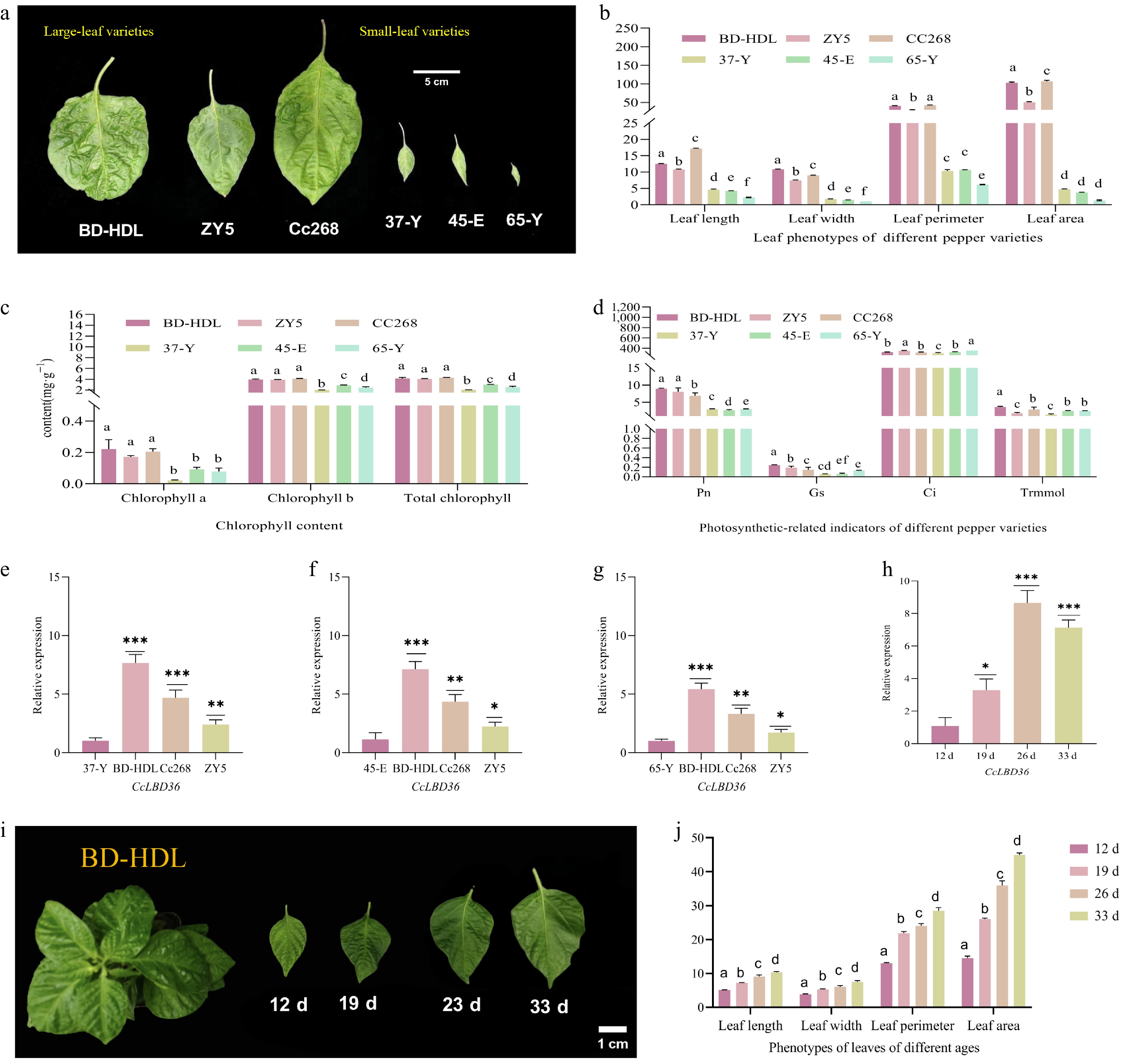
Figure 3.
The expression levels of CcLBD36 genes in different varieties of pepper. Note: All data is presented as the mean values of five biological replicates. Different lowercase letters denote significant differences at the 0.05 probability level. * Indicates p ≤ 0.05, ** indicates p ≤ 0.01, *** indicates p ≤ 0.001, and **** indicates p ≤ 0.0001. Abbreviations and units of the measured physiological and morphological parameters. Photo: Net photosynthetic rate (μmol CO2 m−2·s−1); Gs: Conductance to H2O (mol H2O m−2·s−1); Ci: Intercellular carbon dioxide concentration (μmol CO2 mol−1); Trmmol: Transpiration rate (mmol H2O m−2·s−1). Morphological indicators: leaf length, leaf width, leaf circumference (unit: cm), and leaf area (unit: cm2). Photosynthetic pigments: chlorophyll a, chlorophyll b, and total chlorophyll content (unit: mg·g−1).
qPCR analysis showed that CcLBD36 expression was significantly higher in large-leaf varieties than in small-leaf varieties. Compared with the small-leaf variety 37-Y, expression levels in large-leaf varieties BD-HDL, Cc268, and ZY5 were 765.80%, 469.30%, and 240.50%, respectively (Fig. 3e). Relative to variety 45-E, expression reached 711.40%, 435.90%, and 223.40% (Fig. 3f), and compared to variety 65-Y, levels were 542.30%, 332.30%, and 170.30% (Fig. 3g). These results imply that high expression of CcLBD36 may contribute to large-leaf formation and promote photosynthetic performance.
Leaves from the BD-HDL variety, which exhibited pronounced phenotypic changes at different developmental stages, were selected for analysis. Measurements of average leaf length, width, perimeter, and area showed that leaf size increased progressively with age, with statistically significant differences observed across stages (Fig. 3i, j). qPCR analysis of CcLBD36 expression at these developmental stages indicated that transcript levels gradually rose with leaf age, peaked at day 26, and then declined (Fig. 3h).
The effects of exogenous hormones on CcLBD36 expression
-
To determine whether CcLBD36 is hormone-responsive, its expression in pepper leaves was examined by qPCR following exogenous hormone treatments. The results showed that after application of ABA, IAA, MeJA, and GA3, CcLBD36 transcript levels increased significantly, reaching 383.80%, 376.30%, 451.40%, and 688.40% of the control level, respectively (Fig. 4).
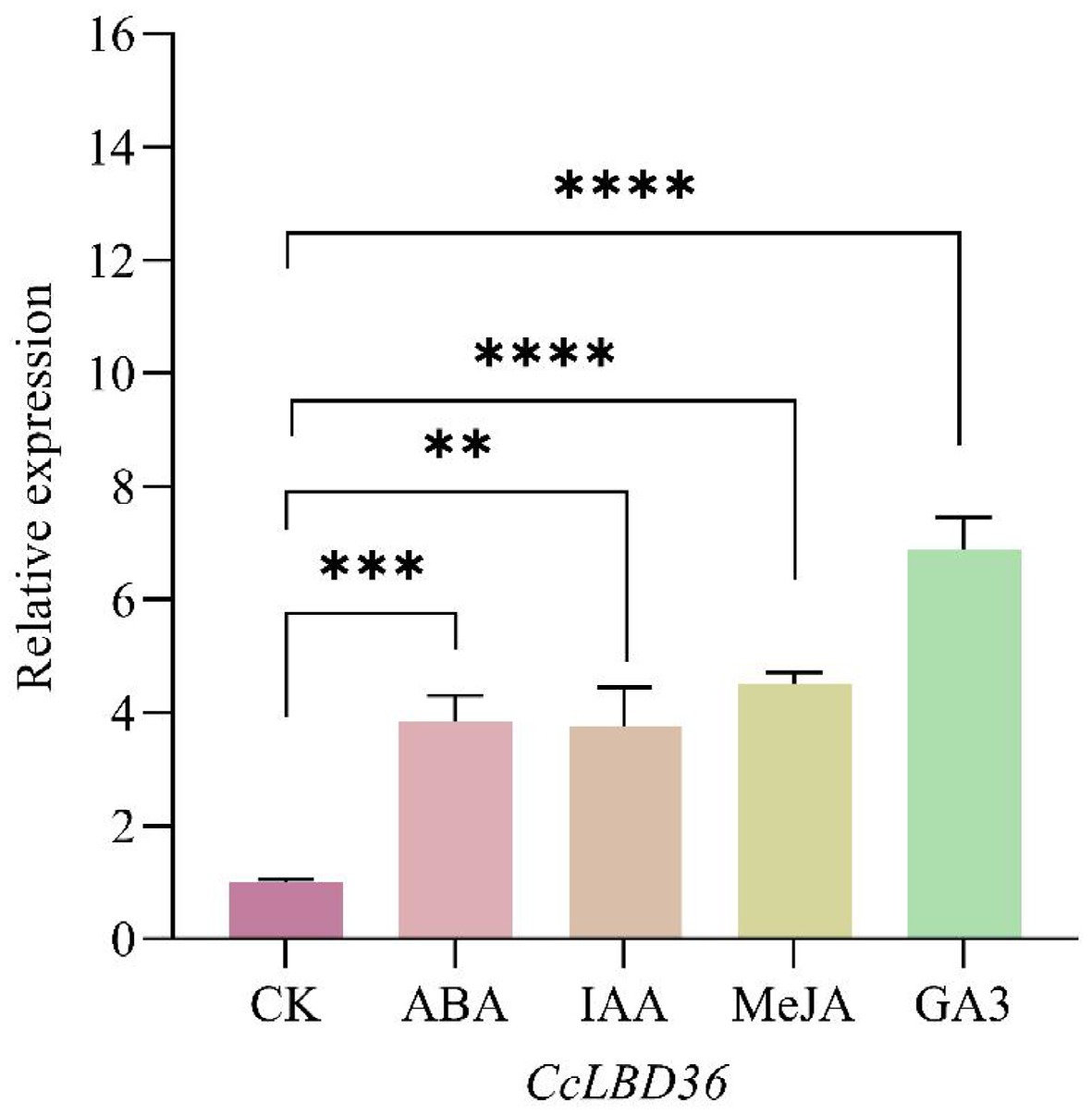
Figure 4.
The expression levels of CcLBD36 genes in capsicum under different hormone treatments. Note: ** indicates p ≤ 0.01, *** indicates p ≤ 0.001, and **** indicates p ≤ 0.0001.
Functional validation of CcLBD36
-
Using the large-leaf variety BD-HDL as the experimental material, a CcLBD36-silencing vector was constructed via virus-induced gene silencing (VIGS). In positive control plants (pTRV2-PDS), a distinct white-leaf phenotype was observed (Fig. 5a), confirming the efficiency of the silencing system. Phenotypic comparisons showed that, relative to the wild-type (WT), leaves of CcLBD36-silenced lines were significantly shorter (Fig. 5b, c) and narrower (Fig. 5b, d), with reduced leaf perimeter and area (Fig. 5e, f). Plant width was also significantly smaller in silenced lines (Fig. 5h), while plant height remained unchanged (Fig. 5g). Chlorophyll content was significantly lower in silenced plants compared to the WT. Chlorophyll a levels in the three silenced lines (PTRV2-LBD36-1, -2, and -3) decreased by 64.67%, 63.15%, and 60.86%, respectively (Fig. 5i). Chlorophyll b content was reduced by 25.72%, 18.79%, and 13.60% (Fig. 5j), and total chlorophyll content declined by 37.46%, 32.11%, and 27.82%, respectively (Fig. 5k).
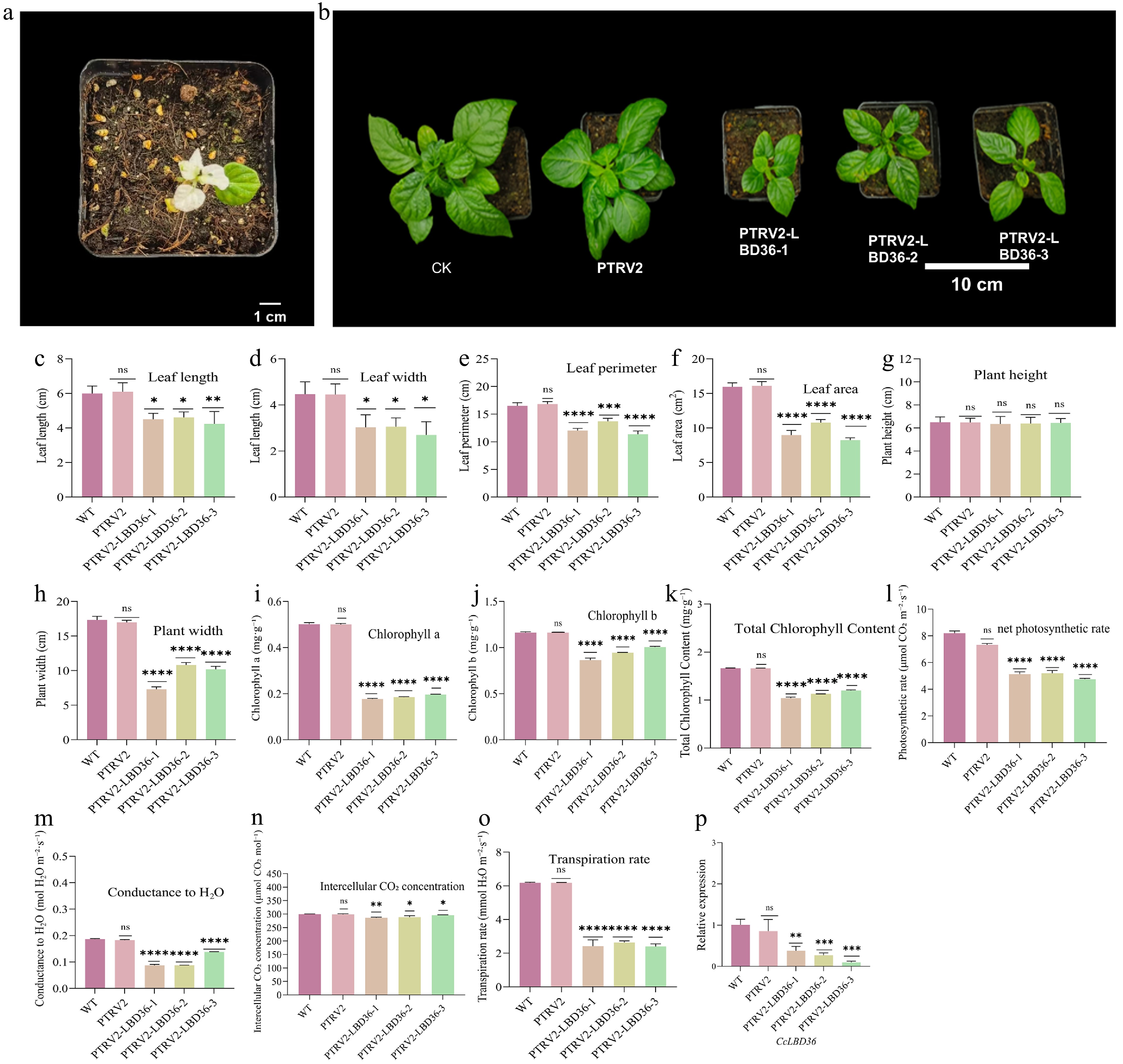
Figure 5.
The albinism map of the positive control (pTRV1-PDS) plants, and the phenotype, chlorophyll content, photosynthetic indicators, and expression level of the CcLBD36 gene of peppers under gene silencing treatment. Note: Different lowercase letters indicate significant differences in levels when p < 0.05. * Indicates p ≤ 0.05, ** indicates p ≤ 0.01, *** indicates p ≤ 0.001, and **** indicates p ≤ 0.0001. Abbreviations and units of the measured physiological and morphological parameters. Photo: Net photosynthetic rate (μmol CO2 m−2·s−1); Gs: Conductance to H2O (mol H2O m−2·s−1); Ci: Intercellular carbon dioxide concentration (μmol CO2 mol−1); Trmmol: Transpiration rate (mmol H2O m−2·s−1). Morphological indicators: leaf length, leaf width, leaf circumference (unit: cm), and leaf area (unit: cm2). Photosynthetic pigments: chlorophyll a, chlorophyll b, and total chlorophyll content (unit: mg·g−1).
Compared with the WT, CcLBD36-silenced lines showed significantly lower net photosynthetic rates, with reductions of 37.39%, 36.66%, and 42.15% in pTRV2-LBD36-1, -2, and -3, respectively (Fig. 5l). Stomatal conductance decreased even more markedly, declining by 53.31%, 53.05%, and 25.93% in the three silenced lines (Fig. 5m). Transpiration rates were also strongly suppressed, falling by 60.92%, 57.36%, and 61.24%, respectively (Fig. 5n). In contrast, the decrease in intercellular CO2 concentration (Ci) was relatively small, with reductions of only 4.46%, 3.45%, and 1.11% (Fig. 5o). Subsequent qRT-PCR analysis of leaf tissue from the silenced lines confirmed that CcLBD36 expression was significantly down-regulated by 62.3%, 73.1%, and 90.4%, respectively (Fig. 5p).
Effects of CcLBD36 on leaf hormone content
-
Silencing of CcLBD36 led to significant changes in hormone levels. Compared with the WT, all three silenced lines exhibited a pronounced reduction in growth-promoting hormones: GA3 decreased by 53.94%, 75.30%, and 97.85% (Fig. 6a); IAA by 60.50%, 51.05%, and 45.02% (Fig. 6b); tZ by 57.85%, 69.16%, and 75.31% (Fig. 6c); and IP by 88.57%, 44.00%, and 37.50% (Fig. 6d). In contrast, the stress-related hormone MeJA increased substantially by 190.80%, 242.53%, and 543.68% (Fig. 6e). The level of the leaf senescence-associated hormone ABA decreased by 76.15%, 17.98%, and 58.43%, respectively (Fig. 6f).
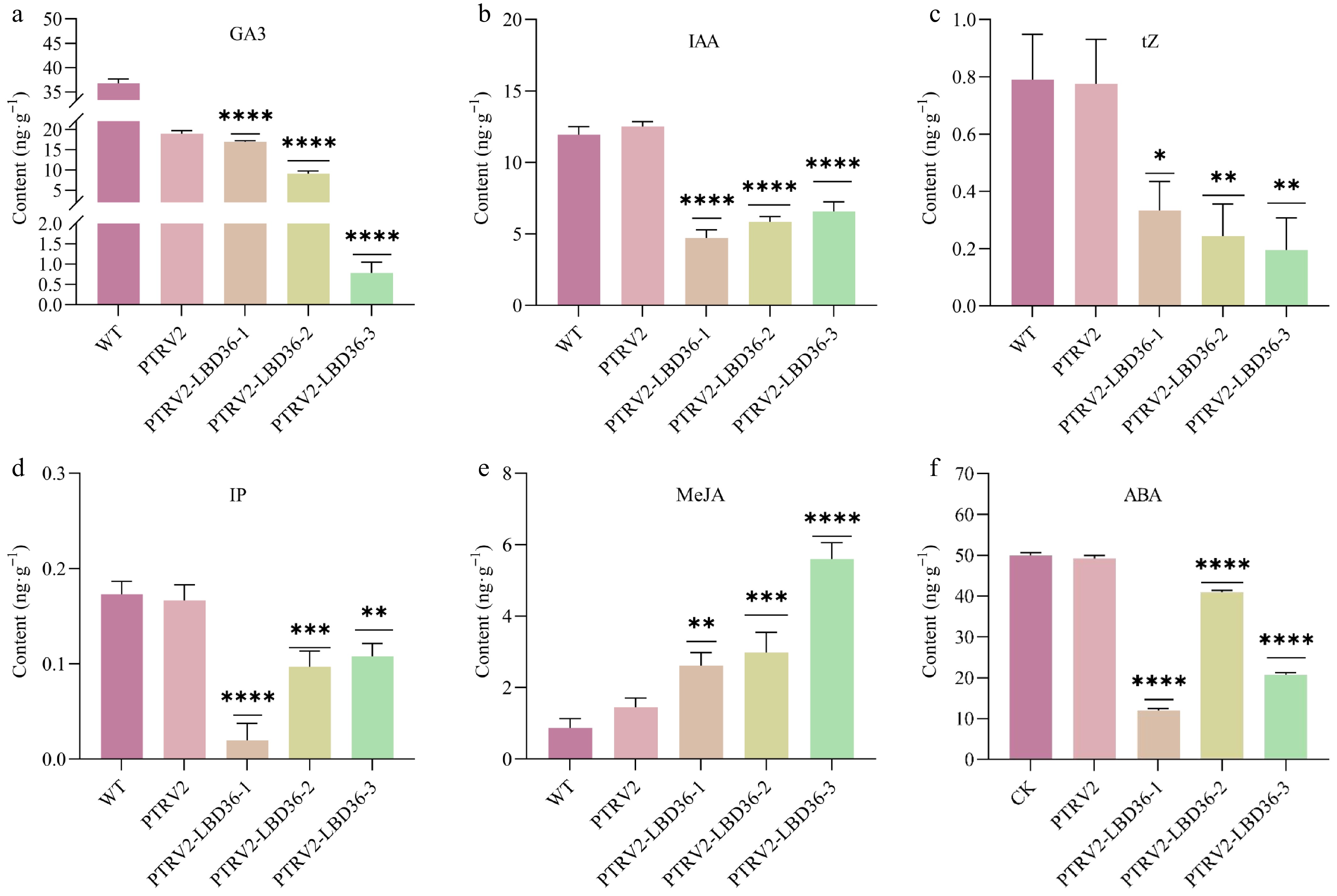
Figure 6.
Hormone content analysis of pepper leaf. Note: * indicates p ≤ 0.05, ** indicates p ≤ 0.01, *** indicates p ≤ 0.001, **** indicates p ≤ 0.0001.
Effects of CcLBD36 on leaf cell morphology
-
To further elucidate the cytological basis of leaf size differences between silenced and control plants, paraffin sections of 40-day-old leaves from the WT, and three CcLBD36-silenced lines (pTRV2-LBD36-1, -2, and -3) were examined. The results showed that CcLBD36 silencing significantly inhibited cell expansion in the palisade tissue, upper epidermis, and lower epidermis (Fig. 7).
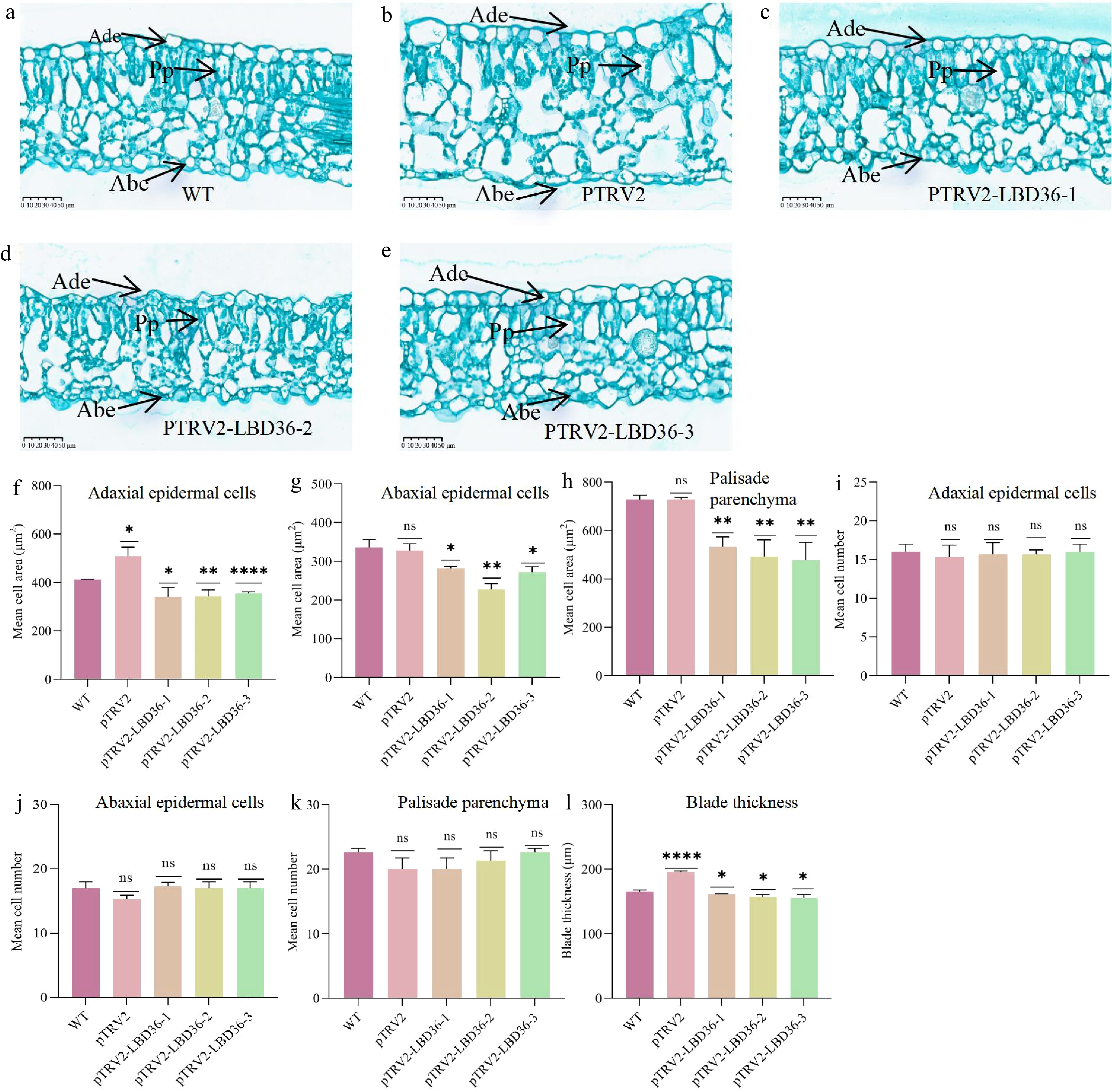
Figure 7.
Leaf anatomical structure and cellular phenotype analysis. Note: Ade indicates Adaxial epithelial cells; Abe indicates Abaxial epithelial cells; Pp indicates Palisade epithelial cells. * Indicates p ≤ 0.05, ** indicates p ≤ 0.01, *** indicates p ≤ 0.001, and **** indicates p ≤ 0.0001.
Compared with the WT, CcLBD36 silencing led to a marked reduction in upper epidermal cell area. The average cell area per unit area in the upper epidermis decreased by 17.4% in pTRV2-LBD36-1 (Fig. 7c, f), 16.7% in pTRV2-LBD36-2 (Fig. 7d, f), and 13.6% in pTRV2-LBD36-3 (Fig. 7e, f). Similarly, the average cell area per unit area in the lower epidermis declined by 15.8%, 32.2%, and 19.0% in the three silenced lines, respectively (Fig. 7g). In the palisade tissue, the average cell area per unit area decreased by 26.9%, 32.4%, and 34.2% in pTRV2-LBD36-1, -2, and -3, respectively (Fig. 7h). Moreover, leaf thickness per unit area was significantly reduced by 2.5%, 5.2%, and 6.2% in the silenced lines compared with the WT (Fig. 7l). In contrast, no significant differences were observed in the average number of cells per unit area in the upper epidermis, lower epidermis, or palisade tissue between silenced and WT plants (Fig. 7i–k). These findings indicate that CcLBD36 influences leaf growth primarily by regulating cell enlargement rather than cell proliferation. Combined with the hormone assay results, it is speculated that the elevation of growth-promoting hormone contents may promote cell expansion by facilitating cell wall loosening and enhancing the expression of genes related to cell elongation, thereby increasing cell area. In contrast, the reduction of growth-inhibiting hormone contents can attenuate their inhibitory effects on cell expansion. Therefore, changes in hormone contents can regulate cell area, which in turn affects leaf thickness and overall leaf area.
-
Pepper is a globally important vegetable consumed both fresh and as a seasoning. Pepper leaves are essential nutritional organs, providing the primary energy source for both vegetative and reproductive growth[33], and thus play a critical role throughout plant development. Leaf length and width in pepper are complex quantitative traits governed by genetic and environmental factors[34]. Through GWAS analysis of 182 pepper accessions—including Hainan wild pepper and varieties from diverse geographic origins—we identified 16 candidate genes associated with leaf length and width, among them the transcription factor gene LBD36. These findings further support the role of specific genes in regulating leaf size and subsequently influencing photosynthetic performance.
Arabidopsis AtLBD36 and C. chinense CcLBD36 exhibit conserved core functions in regulating leaf development, while also displaying species-specific regulatory divergence. Both belong to the AS2/LOB family, share conserved family domains, and participate in leaf development by governing key cellular processes, thereby representing essential components of the leaf developmental regulatory network. Specifically, AtLBD36 acts synergistically with AS2 to mediate leaf morphogenesis and safeguard normal leaf development. In contrast, CcLBD36 functions autonomously and significantly promotes leaf elongation and expansion, leading to increased leaf size. Evolutionary homology and shared core regulatory roles between the two orthologs underscore the evolutionary conservation of gene function. Comparative analysis of their regulatory similarities and differences, combined with the well-characterized function of AtLBD36 in leaf development, firmly establishes that CcLBD36 plays a prominent regulatory role in leaf development, representing a direct functional manifestation of this conserved transcription factor in Chinese pepper[16,35]. Overexpression of the apple MdLBD11 gene in Arabidopsis induces leaf curling[24], and overexpression of the MeLOB36 gene significantly enlarges rosette leaves in Arabidopsis[36]. In this study, the CcLBD36 gene cloned from 'BD-HDL' pepper leaves was shown to positively regulate leaf size, supported by multiple lines of evidence: promoter cis-element analysis, phylogenetic clustering with AtLBD36, differential expression across varieties and leaf ages, and phenotypic alterations in gene-silenced plants.
Extensive research has demonstrated that, within the effective leaf area range, a larger leaf area provides greater physical space for light capture, thereby improving light-use efficiency. Shipley et al.[37], in a meta-analysis of 160 leaf-level datasets from woody and herbaceous plants, reported that specific leaf area (SLA)—closely correlated with leaf area—and leaf nitrogen content jointly determine net photosynthetic rate. Similarly, Li et al.[38] found that reed leaves in waterside habitats had significantly larger leaf areas than those in drier sites, with net photosynthetic rate, actual photosynthetic efficiency, and photochemical quenching coefficient all showing strong positive correlations. Large leaves thus enhance photosynthetic efficiency under favorable water conditions by expanding light-intercepting area and reducing hydraulic resistance. Lei et al. further observed that cotton increases leaf area and chlorophyll content under low-light conditions[39], thereby improving photosynthetic performance and carbon assimilation, which in turn elevates above-ground biomass and productivity.
Leaf internal structure, particularly mesophyll and epidermal anatomy, also significantly influences photosynthesis. In general, palisade tissue exhibits higher photosynthetic capacity than spongy tissue[40]. In the present study, the large-leaf pepper variety with high CcLBD36 expression displayed significantly higher chlorophyll a, chlorophyll b, total chlorophyll content, and net photosynthetic rate compared with small-leaf varieties. Conversely, CcLBD36-silenced lines (PTRV2-LBD36) showed marked reductions in leaf length, width, perimeter, area, plant width, epidermal, and mesophyll cell area, chlorophyll content, and net photosynthetic rate relative to the WT. These results indicate that CcLBD36 acts as a positive regulator that enhances net photosynthesis through two complementary mechanisms: first, by promoting cell expansion to increase leaf area and light-capturing surface; second, by modulating mesophyll tissue structure to create more space for chlorophyll accumulation. Together, these effects synergistically upregulate photosynthetic performance.
Plant hormones coordinately regulate leaf size through modulating cell elongation and division. Studies have shown that auxin response factors OsARF6 and OsARF17 can directly bind to the rice leaf angle enlargement 1 gene (ila1) either individually or synergistically, altering secondary cell wall composition to influence flag leaf angle, which in turn shapes plant architecture and photosynthetic yield in rice[41]. Overexpression of the giant eucalyptus LBD37 resulted in a 61% increase in leaf size, whereas overexpression of LBD29 led to a reduction exceeding 72%. These transgenic plants exhibited significant changes in the expression of key genes involved in gibberellin, ethylene, and auxin signaling pathways, as well as genes related to cell expansion[42]. In Arabidopsis, LBD15 mutants grown on MS medium supplemented with 0.2 μM ABA showed a significantly higher green cotyledon ratio compared to WT plants, while LBD15 overexpressors displayed ABA hypersensitivity and enhanced tolerance to water deficit, highlighting the role of LBD15 in balancing growth and stress adaptation[43]. Further research indicates that GA4 content in rice is highly correlated with leaf length, promoting leaf elongation primarily by stimulating cell division and increasing cell number[44]. In maize, ZmLBD5 expression is strongly induced by ABA, GA3, and methyl jasmonate (MeJA), but responds less distinctly to indole-3-acetic acid (IAA). Overexpression lines of ZmLBD5 exhibit larger leaf area, which is closely associated with increased cell length[45]. In the present study, CcLBD36-silenced lines (PTRV2-LBD36) showed significant reductions in growth-promoting hormones such as IAA, tZ, and IP, whereas levels of the growth-inhibiting hormone MeJA were markedly elevated. Furthermore, exogenous application of IAA, GA3, and MeJA significantly upregulated CcLBD36 expression. These findings suggest that CcLBD36 may participate in plant hormone signaling pathways and cooperate with multiple hormones to jointly regulate leaf growth and development. Furthermore, we speculate that CcLBD36 exerts a negative regulatory effect on MeJA. Silencing of CcLBD36 abolished its negative regulatory function on MeJA biosynthesis or metabolism, thereby relieving the inhibitory effect on endogenous MeJA, and leading to its accumulation in plants. Conversely, exogenous application of MeJA can activate intracellular signal transduction pathways, which in turn induces upregulation of the transcription of the negative regulator CcLBD36.
In summary, we propose that CcLBD36 acts as a key regulator of leaf size and photosynthetic capacity in chili pepper (Fig. 8). CcLBD36 upregulates its own expression in response to growth-promoting hormones (IAA, tZ, IP), while concurrently suppressing the accumulation of the growth-inhibiting hormone MeJA, thereby maintaining a hormonal balance conducive to leaf development. The resulting high expression of CcLBD36 promotes cell expansion in the upper and lower epidermis as well as the mesophyll tissue. This enlarges the leaf area available for light capture and provides increased space for chlorophyll accumulation. In contrast, silencing CcLBD36 (PTRV2-LBD36) disrupts hormone homeostasis—reducing IAA, tZ, and IP, while elevating MeJA, which inhibits cell expansion, leads to epidermal and mesophyll cell shrinkage, lowers chlorophyll content, and reduces net photosynthetic rate. This decline in photosynthetic performance can be attributed to reduced mesophyll area, which diminishes both light interception and carbon assimilation efficiency (Fig. 8).
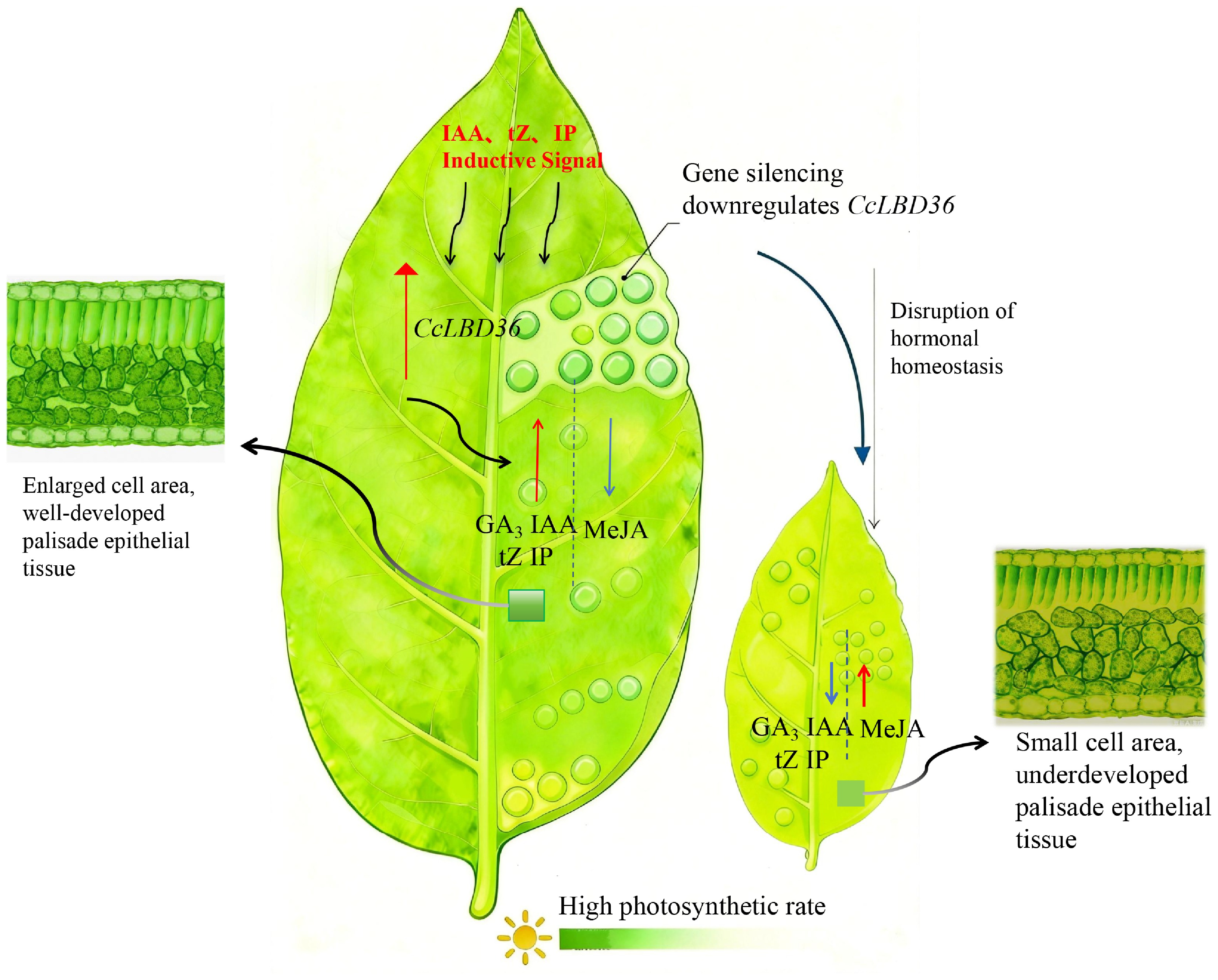
Figure 8.
Schematic model of GA3, IAA, tZ, IP and MeJA crosstalk regulating leaf photosynthetic efficiency upon CcLBD36 gene silencing. Note: Red upward arrows indicate up-regulated gene expression and increased hormone contents; blue downward arrows indicate decreased hormone contents; darker green indicates stronger leaf photosynthesis.
This work was supported by grants from the research Project of Hainan Academician Innovation Platform (YSPTZX202206),National Natural Science Foundation of China (32160717, 32460768) and Key Laboratory for Quality Regulation of Tropical Horticultural Crops of Hainan Province (202505).
-
The authors confirm their contributions to the paper as follows: conceptualization, funding acquisition, software, validation, investigation, project administration, writing − original draft preparation, writing − review and editing: Cheng S, Cai J; methodology, visualization: Cai J, Cheng S, Zeng Q; formal analysis, data curation: Cai J, Zeng Q, Yang M, Wu K, Huang R; resources: Cheng S, Wang Z, Fu H, Lu X, Li M; supervision: Cheng S, Wang Z, Fu H, Lu X. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study is included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper online at: https://doi.org/10.48130/tp-0026-0005.
-
Received 23 December 2025; Accepted 27 February 2026; Published online 16 March 2026
- Supplementary Fig. S1 Manhattan plot and QQ plot of genome-wide association analysis of leaf length in 182 core germplasms of pepper.
- Supplementary Fig. S2 The cDNA sequence and amino acid sequence of the CcLBD36 gene.
- Supplementary Table S1 Primers for the genes related to Capsicum leaf length.
- Supplementary Table S2 Prediction of candidate genes LBD36 related to pepper leaf length.
- Supplementary Table S3 Phylogenetic tree sequence information.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Cai J, Zeng Q, Yang M, Wu K, Huang R, et al. 2026. Physiological and cellular mechanisms of the transcription factor CcLBD36 positively regulating leaf size in pepper. Tropical Plants 5: e005 doi: 10.48130/tp-0026-0005 

Physiological and cellular mechanisms of the transcription factor CcLBD36 positively regulating leaf size in pepper
- Received: 23 December 2025
- Revised: 11 February 2026
- Accepted: 27 February 2026
- Published online: 16 March 2026
Abstract: A genome-wide association study of leaf length in 182 pepper accessions, combined with gene annotation and p-value screening, identified LBD36 as a key candidate gene associated with leaf size. Sequence characteristics, phylogenetic relationships, chromosomal localization, and promoter cis-acting elements of CcLBDs in the genome of Capsicum chinense were analyzed. CcLBD36 was cloned from Hainan huangdenglong pepper. Real-time quantitative PCR analysis showed that CcLBD36 expression was significantly higher in large-leaf pepper varieties than in small-leaf varieties, and was positively correlated with chlorophyll a, chlorophyll b, total chlorophyll content, and net photosynthetic rate. Exogenous application of IAA, GA3, and MeJA induced its upregulation, while its expression exhibited an initial increase, followed by a decline during leaf development. Virus-induced gene silencing (VIGS) targeting CcLBD36 resulted in significantly reduced leaf area, leaf length, leaf width, and plant width, along with decreased chlorophyll a, chlorophyll b, total chlorophyll content, net photosynthetic rate, stomatal conductance, intercellular CO2 concentration, and transpiration rate. Cytological analysis revealed significantly smaller cell areas in the leaf tissues of silenced plants, while hormone assays indicated a concurrent reduction in growth-promoting hormones, and a marked increase in the inhibitory hormone MeJA in these lines. In summary, CcLBD36 positively regulates pepper leaf size.
-
Key words:
- Capsicum /
- Leaf size /
- Gene silencing /
- Cytological analysis