









-
The family Brassicaceae includes various vegetable crops that are widely grown across the world[1]. Broccoli is the most often consumed brassica vegetable and contains a wide variety of bioactive secondary compounds, such as glucosinolates[2]. Broccoli is harvested as immature inflorescences (and their stems) and has a relatively short shelf life when stored at room temperature[3] mainly due to premature senescence. Harvesting suddenly interrupts the supply of energy, nutrients, and hormones, leading to rapid senescence. Yellowing of the inflorescence tissues during postharvest senescence is one of the major negative impacts on the economic value of broccoli heads and can occur at any step during harvest, transport, and storage.
Our current understanding of biotic and abiotic stress responses in broccoli before and after harvest is limited to transcriptomic studies[4−6]. Although RNA-sequencing transcriptomics provides robust and high-throughput data about gene regulation, spatio-temporal mRNA expression does not always overlap with protein expression or reflect protein levels[7]. Moreover, poor coordination between transcript and protein levels has been observed in some studies[8]. Thus, it is important to identify and characterize the proteins active during broccoli senescence through proteomic approaches to reveal protein dynamics during cellular senescence. Changes in protein levels during senescence can reflect how plants acclimatize to the environment while maintaining homeostasis. Protein functions are not just restricted to their identities or structures, but also depend on location and post-translational modifications. Therefore, it is essential to discover the protein dynamics at the tissue and cellular levels during the onset and progression of senescence, which can potentially lead to the development of effective treatments for the inhibition of postharvest senescence and thus improvement of postharvest shelf life.
In recent years, omics technologies have made tremendous contributions to the study of molecular and cellular mechanisms in crop plants. Diverse proteomic approaches have been employed to investigate postharvest quality and shelf life in vegetables and fruits[3]. Labelling techniques such as isobaric Tags for Relative and Absolute Quantitation (iTRAQ) and Tandem Mass Tags (TMT) have been used to quantify protein profiles in fresh produce. The principles underlying iTRAQ and TMT are essentially the same, with differences in labeling specificity and in detecting the molecular structure of proteins. TMT can label a broader range of proteins and peptides[3]. A TMT method has been applied to broccoli sprouts to monitor the protein changes induced by jasmonic acid (JA)[9]. Previous transcriptomic analyses have reported some key genes and transcription factors (TFs) involved in color changes of broccoli and the cytokinin response in broccoli based on proteomics and transcriptomics[10]. Additionally, exogenous application of JA increases indolic glucosinolate in Brassica rapa[11] through transcript regulation of the indole glucosinolate pathway[12]. Broccoli sprouts showed that JA plays a role in plant defense by regulating protein production related to various metabolic pathways, such as carbohydrate catabolism and amino acid biosynthesis, inhibiting the photosynthesis process[9]. A proteome-level study based on iTRAQ and TMT labelling was carried out on postharvest tobacco leaves when they were exposed to high temperatures of 38 and 42 °C, showing that 1,150 up-regulated and 1,074 down-regulated proteins were differentially expressed during senescence and that many proteins were related to sugar and energy metabolism, with decreases in the photosynthetic machinery and increased nutrient recycling through degradation of older cellular constituents[13]. While already proven that carbohydrate- and energy-related metabolic pathways (like glycolysis and TCA cycle, and starch and sucrose metabolism) play a crucial role in the senescence process by regulating the metabolic energy in the cells[14], mass spectrometry (MS) has been used to more recently show that key protein complexes are highly conserved across different plant species[15]. Such high throughput analysis identified two million abundant proteins that are significantly expressed in different tissues across 13 plant species. Additionally, co-fractionation MS was used to newly identify conserved protein subunits involved in vernalization, pathogen defense, and certain other agricultural traits that could reveal ways to improve crop yield and biofuel production[15].
The present study was conducted to test the hypothesis that loss of freshness in harvested produce is divided into discrete processes within senescence that can be defined by diagnostic patterns of the protein profile. The proteins from harvested broccoli during storage under different temperatures were quantitively analyzed using Tandem Mass Tag technology. The protein profiles were linked to specific stages of storage under each treatment. The results provide a high throughput quantification of the proteome on a global level.
-
Fresh broccoli (Brassica oleracea) florets were obtained from commercial farms in Hastings, Florida, USA. All broccoli samples were manually harvested at the farms to minimize mechanical damage and were transported approximately 80 min in an air-conditioned automobile to the Postharvest Laboratories in Gainesville, FL, USA. Broccoli florets that were of the same shape and size and harvested at the same physiological age were used for protein profiling. The heads were stored in darkness either in a cold room at 4 °C or in a plant growth chamber at 25 °C, both with a relative humidity (RH) of 95% to 100%. Three biological replicates of broccoli florets were used for RNA sequencing and proteomics at each sampling time. Subsamples of inflorescence tissue were collected from the broccoli florets on days 1, 3 and 5 and wrapped in aluminum foil. The samples were immediately frozen in liquid nitrogen and stored at −80 °C until further use.
RNA extraction and transcriptome sequencing
-
Broccoli floret tissues were chosen for total transcriptome analysis. There were 18 samples in total, three replicates each sampled on days 1, 3, and 5 from the two storage temperatures. Total RNA was isolated from broccoli tissue using Trizol (Ambion, Life Technologies, Norwalk, CT, USA) as per the manufacturer's protocol. Extracted RNA was subjected to DNase treatment (Turbo DNA free, Thermo Fisher Scientific, Waltham, MA, USA). The absorption ratios at 260/280 nm and 260/230 nm were tested on a Nanodrop (Applied Biosystems, Waltham, MA, USA) to determine concentration and purity.
Protein extraction from stored broccoli florets
-
Samples (2.0 g) of broccoli floret tissue were ground in a mortar and pestle using liquid nitrogen. Finely powdered tissue was solubilized in 2.0 mL of phenol and 1.0 mL of protein extraction buffer (10 mM Tris pH 7.2, 10 mM EGTA, 150 mM NaCl, 10 mM KCl, 1% Sigma plant protease inhibitor cocktail, 20 mM dithiothreitol). Samples were agitated for 1 h followed by centrifugation (Beckman Coulter, Brea, CA, USA) at 11,000× gn for 30 min at 4 °C. The supernatant was discarded, and the pellet was suspended in a 1:1 ratio of phenol and protein extraction buffer and allowed to gradually resuspend. This extraction was repeated thrice for all processed samples. Samples in 15-mL tubes were shaken overnight in a top to bottom shaker at −20 °C. The next day, the samples were centrifuged at 11,000× gn, for 30 min at 15 °C. Aliquots of extracted proteins were frozen in liquid nitrogen followed by long-term storage at −80 °C. The samples were digested with 50 μL trypsin (V5111 Promega, Fitchburg, WI, USA) at 37 °C for overnight incubation. The digested peptides were incubated with TMT 11-plex labelled kit (Sigma Aldrich, Burlington, MA, USA) for 2 h at room temperature. The protein concentration was analyzed using the PierceTM BCA Protein Assay Kit (Thermo Fisher Scientific).
Protein sample preparation, digestion, and TMT labeling
-
Proteins were being diluted seven-fold with 100 mM TEAB, and then reduced by 10 mM Tris (2-carboxyethyl) phosphine at 37 °C for 1 h, followed by alkylation by 20 mM iodoacetamide in the dark for 30 min. Trypsin (Promega), solubilized in 100 mM TEAB, was added for digestion (w/w for enzyme:sample = 1:100, overnight at 37 °C). The digested peptides were desalted on MacroSpin Column Silica C18 columns (The Nest Group, Ipswich, MA, USA), and then labeled using the TMT11-plex tagging kit (Thermo Fisher Scientific) according to the manufacturer's instructions to label all the samples. To generate a universal control sample, equal aliquots of each sample were pooled, mixed and labeled with the TMT-tag. The labeling was quenched by adding 5% hydroxylamine to the reaction mixture, and the samples were subsequently pooled and desalted by C18-solid phase extraction. Labeled peptides were then dissolved in a strong cation exchange (SCX) solvent [Solvent A: 25% (v/v) acetonitrile, 10 mM ammonium formate, and 0.1% (v/v) formic acid, pH 2.8].
Strong cation exchange fractionation and reverse-phase LC-MS/MS
-
The peptides were fractionated using an Agilent HPLC 1260 (Agilent Technologies, Santa Clara, CA, USA) with a polysulfoethyl A column (2.1 mm × 100 mm, 5 µm, 300 Å; PolyLC, Columbia, MD, USA). Peptides were eluted using a linear gradient of 0%–20% Solvent B [25% (v/v) acetonitrile and 500 mM ammonium formate, pH 6.8] over 50 min followed by ramping up to 100% Solvent B over 5 min. The absorbance at 280 nm was monitored, and a total of 12 fractions were collected. The fractions were lyophilized and resuspended in LC solvent C [0.1% formic acid in 97% water (v/v), 3% acetonitrile (v/v)]. A hybrid quadrupole Orbitrap (Q Exactive Plus) MS system (Thermo Fisher Scientific, Bremen, Germany) was used with high energy collision dissociation (HCD) in each MS and MS/MS cycle. The MS system was interfaced with an automated Easy-nLC 1000 system (Thermo Fisher Scientific, Bremen, Germany). Each sample fraction was loaded onto an Acclaim PepMap 100 pre-column (20 mm × 75 μm; 3 μm-C18, Thermo Fisher Scientific) and separated on a PepMap RSLC analytical column (250 mm × 75 μm; 2 μm-C18) at a flow rate at 350 nL/min during a linear gradient from LC Solvent C [0.1% formic acid (v/v)] to 30% Solvent D [0.1% formic acid (v/v) and 99.9% acetonitrile (v/v)] for 95 min, to 98% Solvent D for 15 min, and held at 98% Solvent D for additional 30 min. Full MS scans were acquired in the Orbitrap mass analyser over a range of 400–2,000 m/z with a resolution of 70,000 at 200 m/z. The top ten most intense peaks with charge state ≥ 2 were isolated (with a 2-m/z isolation window) and fragmented in the high energy collision cell using a normalized collision energy of 28%. The maximum ion injection time for the survey scan and the MS/MS scans were 250 ms, and the ion target values were set to 3e6 and 1e6, respectively. The selected sequenced ions were dynamically excluded for 60 s.
Proteomics data analysis
-
The raw MS/MS data files were processed by a thorough database search with consideration to biological modification and amino acid substitution against a green plant database using ProteinPilot v5.0 with the Fraglet and Taglet searches under the ParagonTM algorithm. The following parameters were considered for all searches: fixed modification of methylmethane thiosulfonate-labeled cysteine, fixed TMT modification of amine groups in the N-terminus lysine, and variable TMT modifications of tyrosine. The false discovery rate at the peptide level was estimated to be 1.0% using the integrated PSPEP tool in the ProteinPilot Software (SCIEX, Framingham, MA, USA). For quantification, only MS/MS spectra that were unique to a particular protein and had a sum > 9 of the signal-to-noise ratios for all the peak pairs were used. The ratios with p-values less than 0.05 and that were present in at least two replicates were considered significant. Only the significant ratios from the replicates were used to calculate the average ratio for a protein. For a protein to be determined as differentially expressed, it must have been identified and quantified with at least three unique peptides. All the differentially expressed proteins (DEPs) are listed in Supplemental Tables S1 & S2.
RT-PCR validation of the differentially expressed proteins in the proteomics study
-
Expression studies were performed for the differentially expressed proteins observed during the protein profiling in the broccoli stored for 1, 3, or 5 d after harvest. Relative transcript expression was investigated using three technical replicates from each of the biological replicates from these three time points. The Actin2 gene was used as an internal reference control (Supplemental Table S3). The total reaction volume was 12 µL for each gene using the reagents recommended for the Bio-Rad Thermocycler (Bio-Rad, Hercules, CA, USA) and the following conditions: 95 °C for 10 min, 45 cycles at 95 °C for 30 s, 60 °C for 30 s. Relative gene expression was calculated by the ΔΔCᴛ method[16].
Tobacco transient assay
-
For the selected DEPs, cDNA corresponding to Arabidopsis homologs were obtained from the Arabidopsis Biological Resource Center (ABRC;
https://abrc.osu.edu ). The protein coding sequence (CDS) for each gene was fused into the pMDC43 vector, which contains a 35S promoter and a green fluorescent protein (GFP) tag. Each plasmid was introduced into the Agrobacterium strain GV3101. An overnight culture of bacteria containing each gene of interest was prepared, the pelleted culture was dissolved in a 100 mL MS liquid solution containing N-morpholino-ethanesulfonic acid (MES) (10 mM), magnesium chloride (10 mM), and acetosyringone (100 μM) and then adjusted to an optical density (OD) of 0.5 at A600. A 5-mL syringe was used to gently inject the culture into the abaxial side of 28-day-old tobacco leaves. We repeated the experiment three times with three biological replicates. Photographs were taken every 3 d to track phenotypic changes in the leaves. -
We observed distinct differences between broccoli stored at 4 and 25 °C. At 4 °C, the broccoli maintained its green color and firmness for a longer period compared to those stored at 25 °C. In contrast, broccoli stored at 25 °C exhibited faster yellowing and wilting, indicative of accelerated senescence. We have performed physiological parameters such as water loss, chlorophyll measurement in our previous research on transcriptomics of stored broccoli[4]. To understand the proteomic responses of storage temperatures, we conducted proteomics analysis. Farm-grown broccoli in three biological replicates was stored at two different temperatures, 25 and 4 °C, for 1, 3, and 5 d before proteomic profiling was conducted to understand the onset and progression of senescence using a TMT-based method. The total protein was isolated from broccoli florets and digested into peptides that were subsequently labelled with isobaric reagents to be run and analyzed using LC-MS/MS.
Two sets of internal controls were used to normalize the data in order to determine the differentially expressed proteins (DEPs). Sample name descriptions are detailed in Supplemental Tables S1 & S2. For instance, to identify the DEPs between refrigerated and room temperature storage, all samples from day 1 stored at 4 °C were pooled for use as the control (Supplemental Table S1). We then used a fold-change cut-off (cut-off score = 2) to select 1714 DEPs from the broccoli stored as 25 °C for 1 d and identified them using sequences available from Uniprot (
www.uniprot.org ) (Supplemental Table S2). Similarly, we pooled all samples from day 1 stored at 25 °C and selected 2,276 DEPs from the broccoli stored at 4 °C. There were a total of 2,395 proteins expressed only at either 25 or 4 °C (Supplemental Tables S1 & S2). To determine the upregulated and downregulated DEPs for a specific storage condition, we conducted Venn Diagram analysis (Fig. 1a, b). Among the upregulated DEPs, there were 156 proteins detected at 4 °C and 221 proteins at 25 °C. A higher number of proteins were upregulated on day 3 and day 5 of storage at 25 °C in comparison to storage at 4 °C, suggesting that a larger number of proteins related to stress, growth, and development were induced during the more rapid senescence that occurred at the higher temperature. In contrast, the samples stored for 5 d at 4 °C had a larger number of down-regulated DEPs than those stored at 25 °C. There were 231 DEPs identified during storage at 25 °C, out of which 71 were up-regulated only on day 3 and 67 were up-regulated only on day 5; however, 93 proteins were shared between days 3 and 5 (Fig. 1c, Supplemental Table S1). On the other hand, 169 DEPs were identified during storage at the lower temperature, of which only 57 were downregulated on day 3 and 29 on day 5. Additionally, the higher number of DEPs following storage at 25 °C suggests that protein expression significantly increased during the 5 d of storage, resulting in an accumulation of protein, leading to faster progression of senescence at the higher temperature.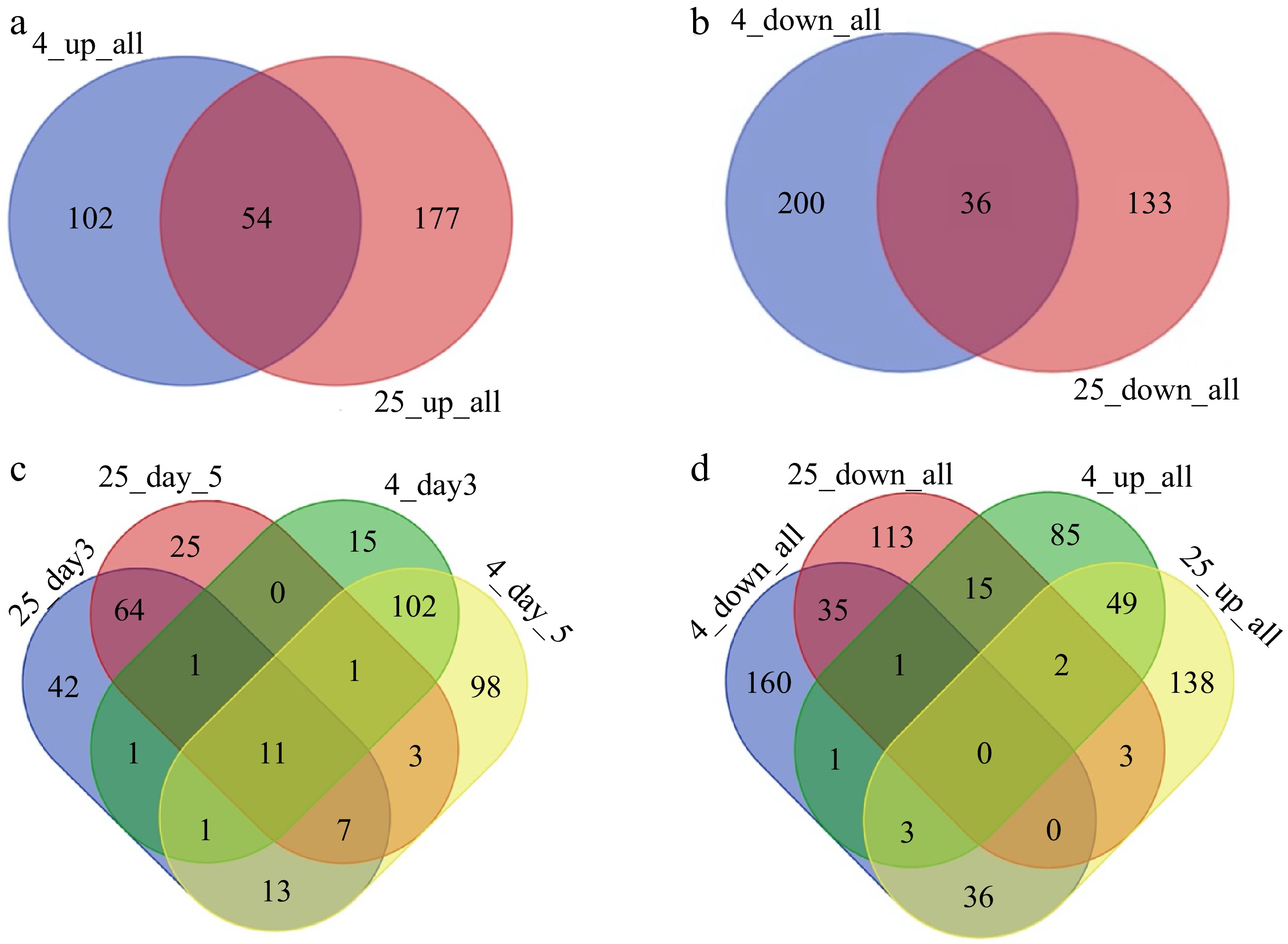
Figure 1.
The number of differentially expressed proteins under different storage conditions. (a) Venn diagram illustrating all up-regulated DEPs between 25 and 4 °C after storage for 3 or 5 d. (b) Venn diagram illustrating all the down-regulated DEPs between 25 and 4 °C. (c) The number of DEPs at different timepoints (days 3 and 5, compared to day 1) in the 25 and 4 °C treatments. (d) Venn diagram of the DEPs between the warm and cold storage conditions for all points of time.
There were some proteins that were differentially regulated on both day 3 and day 5 under both 25 and 4 °C, suggesting that certain proteins underwent biological programs during senescence regardless of the temperature (Fig. 1a, b). There were 54 upregulated DEPs that were present in both warm and cold-stored samples. These included proteins encoded by the ARGONAUTE4 (AGO4), DICER-LIKE3 (DCL3), and SUPPRESSOR OF GENE SILENCING3 (SGS3) genes. Several DEPs in this group were also involved in silencing transcriptional genes as components of the RISC complex associated with the small interfering RNA (siRNA) pathway. These results suggested that siRNA-involved molecular processes modulate tissue senescence. Proteins encoded by 36 genes were downregulated under both warm and cold storage. Among them, 30% were ribosomal proteins, suggesting that ribosome biosynthesis was reduced during senescence.
There were 36 DEPs that were upregulated at 25 °C, but downregulated at 4 °C (Fig. 1d). Many known proteasome components were present in this group, for example, Embryo Defective 2719 (EMB2719), 20S Proteasome Beta Subunit (PBG1), Regulatory Particle Non-ATPASE 12A (RPN12a), ATS9, and Regulatory Particle Non-ATPASE (RPN10). The 26S proteasome mediates ubiquitin-dependent protein degradation, and the 20S proteasome mediates ubiquitin-independent protein degradation[17]. These studies confirmed that both 26S and 20S proteasome activities were inhibited at low temperature, which would likely slow down the progression of senescence. A further 15 DEPs were downregulated at 25 °C but upregulated at 4 °C (Fig. 1d). These included several Nascent polypeptide-Associated Complex subunit (NAC) proteins. NAC proteins are involved in the autophagy pathway and in regulating protein homeostasis during tissue senescence[9]. This indicated that high temperature influenced the autophagy pathway and inhibited protein homeostasis under stress conditions.
Functional categorization of the DEPs
-
To determine the properties and functional classifications of the DEPs in postharvest broccoli, the proteins were linked to gene ontology (GO) terms within the database, and the GO was analyzed to categorize the DEPs into cell components, molecular functions, and related biological processes (Fig. 2, Supplemental Table S4). At room temperature, several GO terms related to biological activities, such as stress responses, abiotic stimuli, chemical responses, and transport mechanisms, were enriched. The DEPs were mainly involved in organellar and cytoplasmic development processes responsible for RNA and protein binding, which could lead to structural modifications during postharvest senescence.
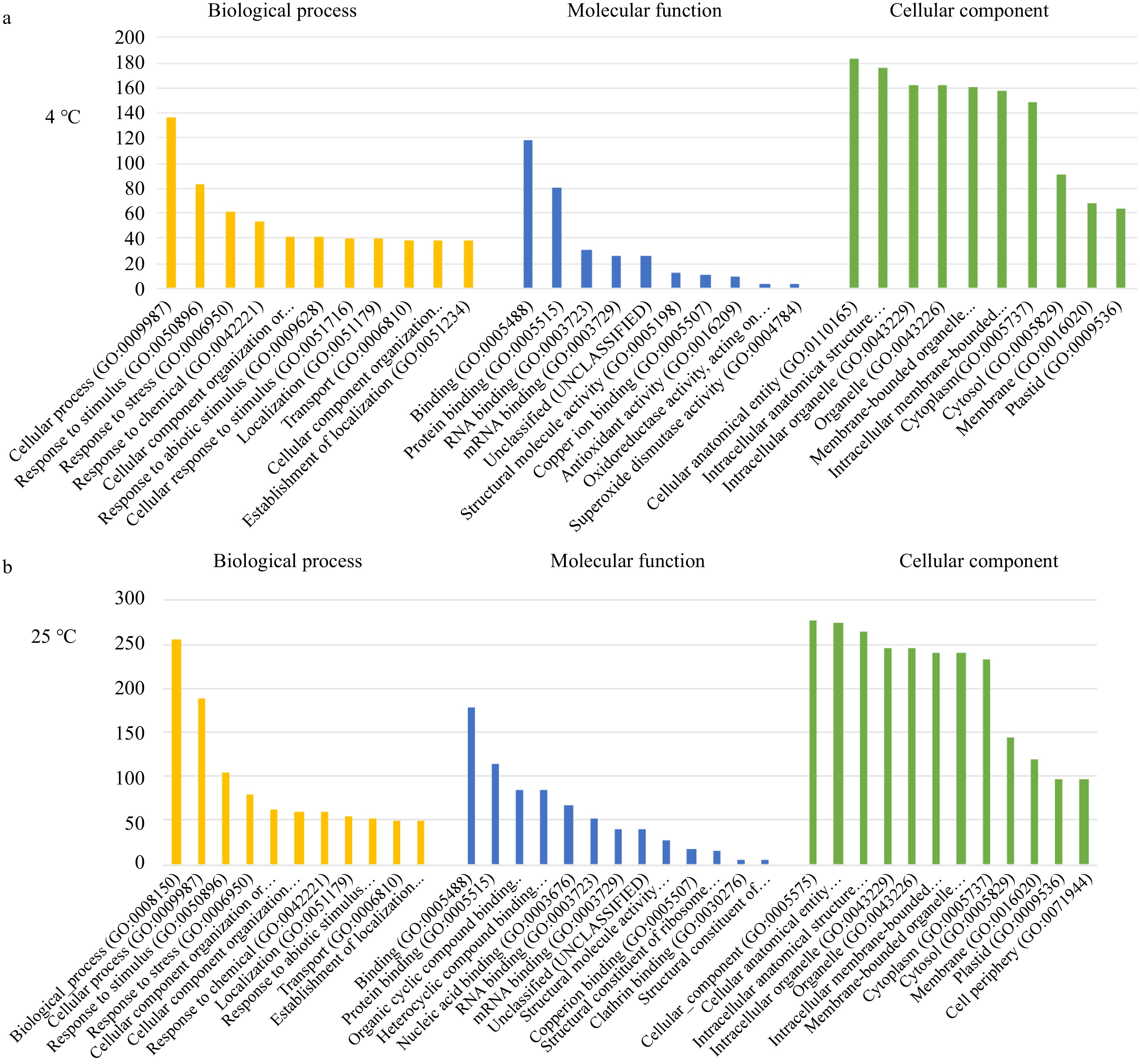
Figure 2.
Gene Ontology (GO) enrichment analysis of broccoli DEPs identified by comparing day 5 to day 1 of storage at (a) 4 °C and (b) 25 °C. The y-axis is the number of genes enriched in each individual GO term categories.
The GO enrichment studies from cold storage identified similar processes, yet with lower rates of enrichment for GO terms related to signal processing compared to those related to metabolic processes, cellular components, and anatomical development, with an additional increase in antioxidant activity and oxidoreductase activity. Overall, the percentage response to the functions related to room temperature storage of broccoli was higher relative to cold storage. Therefore, the rate of progression of postharvest senescence was higher at room temperature due to higher biological activity, with a steep decline in energy related to these biological processes.
Identification of oppositely regulated proteins during cold and room temperature storage
-
To discover regulators of temporal postharvest stress responses, we examined postharvest proteins that could lead to a better understanding of distinct gene expression over storage time. Such genes could possibly serve as indicators of the different stages of senescence and be used to monitor the physiological status of broccoli. We sought candidate transcripts associated with fundamental physiological processes or biochemical pathways likely to be highly conserved over a wide range of conditions. We identified proteins that were oppositely regulated under room temperature and cold storage and that also showed stage-specific expression patterns, including proteins upregulated after 5 d of storage at room temperature but downregulated under the other conditions and at the other time points (Fig. 3a, b). These criteria identified transcripts that showed distinctive, repeatable patterns of change during the period of senescence before senescence symptoms such as yellowing were visible and also groups of transcripts that changed in contrasting ways. A heat map of protein expression patterns illustrated the proteins that were oppositely regulated at room temperature or cold storage (Fig. 3c). Our analysis revealed four indicators of senescence: calmodulin-domain protein kinase 5 (CDPK5, LOC106307255), mitogen-activated protein kinase 8 (MPK8, LOC106325233), a growth regulating factor (GRF5, LOC106266836), and a RING/PHD zinc finger transcription factor (LOC106295767) as proteins induced by storage under higher temperature for 5 d (Fig. 3c). These protein kinases and transcription factors are known to play crucial roles in switching genes on or off during developmental transitions due to their ability to regulate various cellular processes, including gene expression, signal transduction, and protein activity.
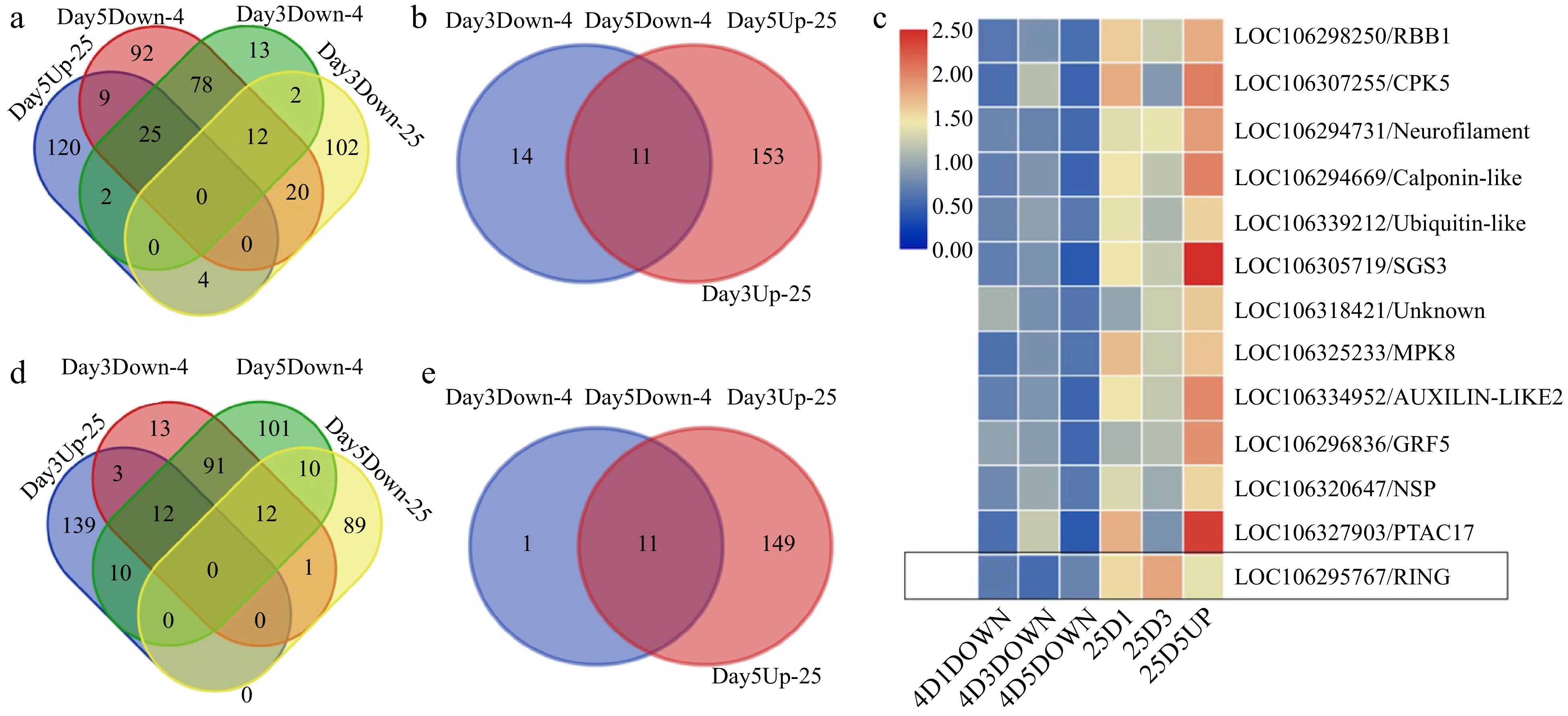
Figure 3.
Proteins showing oppositely regulated expression patterns in different temperature treatments. (a) Proteins that were up-regulated at day 5 during room temperature storage while being down-regulated at day 3 during room temperature storage as well as during cold storage. (b) Proteins that were up-regulated at day 3 and day 5 during the room temperature storage but were down-regulated during the cold storage. (c) A heat map illustrating the oppositely regulated expression patterns of proteins during room temperature or cold storage. The LOC106295767, a ring finger gene, was highlighted in the black box showing the down regulation in cold storage while up regulation during room temperature storage. (d) Proteins that were up-regulated at day 3 during room temperature storage while being down-regulated at day 3 during room temperature storage as well as during cold storage. (e) Proteins that were up-regulated at day 3 and day 5 during the room temperature storage but were down-regulated during the cold storage.
It is worth noting that protein kinases can also trigger a cascade of post-translational modifications, such as ubiquitination, SUMOylation, and acetylation. These modifications can further influence protein interactions and localization, impacting gene regulation and developmental transitions. Several proteins related to these steps of signal transduction were identified among those upregulated after 3 d of storage at 25 °C (Fig. 3d, e; Supplemental Table S5), including an adenylate kinase (ADK1, LOC), a dehydrin (HIRD11), an early nodulin-like protein (ENODL 15), and a tetraketide alpha-pryone reductase (TKPR2). Those protein kinases and dehydrin proteins act as intermediaries in signal transduction pathways, receiving and transmitting these signals to the nucleus, where they can influence gene expression. We also found proteins up-regulated at day 3 during room temperature storage while down-regulated at day 3 and day 5 during cold storage. Those proteins can be used as marker genes for day 3 during room temperature storage. A list of those protein-associated genes are included in Supplemental Table S5.
Validation of DEPs through Real-Time PCR
-
Changes in mRNA expression levels may correlate with changes in protein expression. Therefore, comparing qPCR results with proteomics data can provide a more comprehensive understanding of gene expression regulation. To validate the proteomics results, we investigated the transcript levels for the top 12 DEPs in broccoli over time during storage. The genes that encoded the 12 DEPs were identified, and their transcript levels were examined during storage by qRT-PCR (Fig. 4). It's crucial to acknowledge that mRNA levels do not always correlate perfectly with protein levels due to post-transcriptional and translational modifications, and protein stability factors.
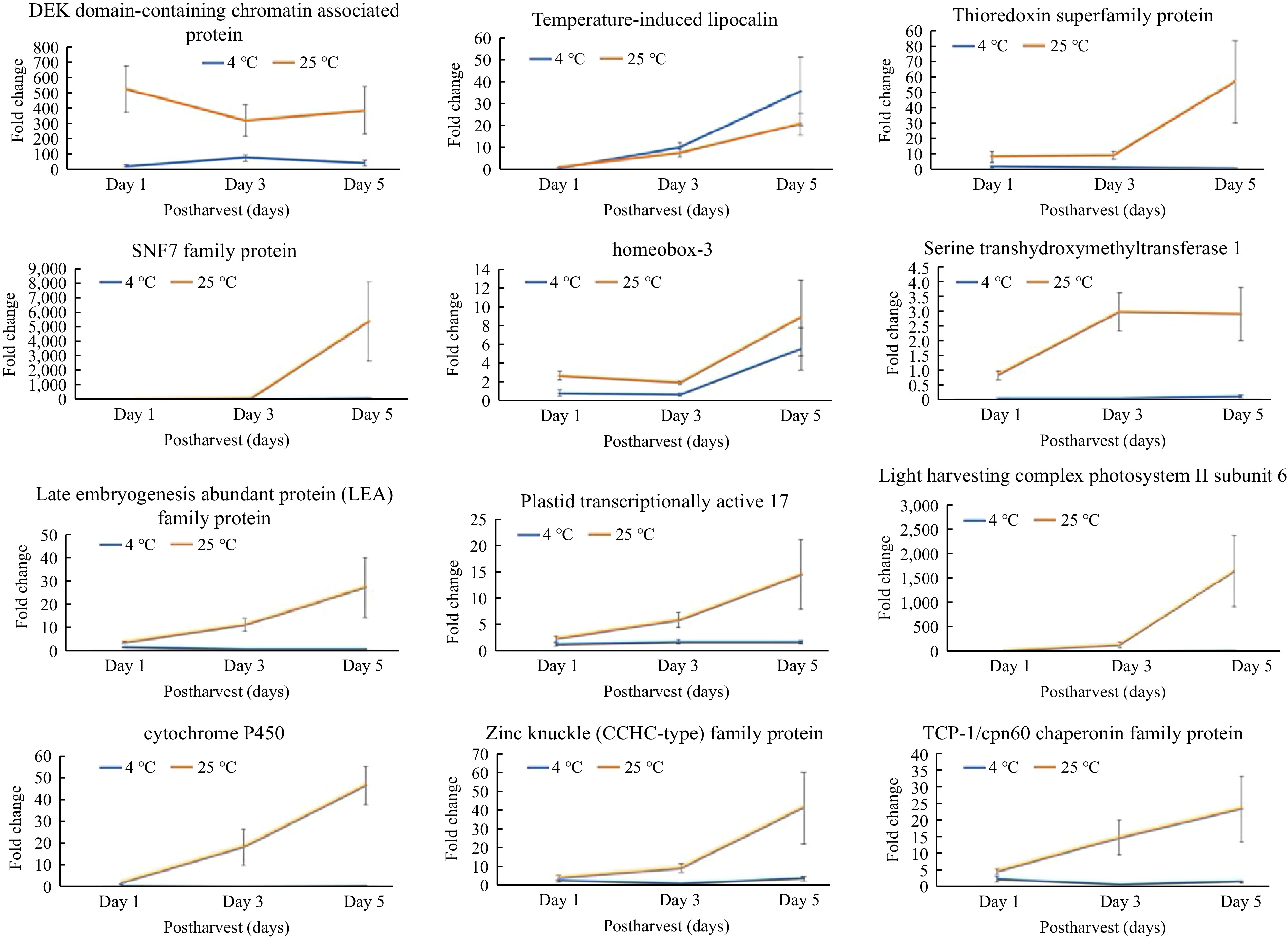
Figure 4.
Relative transcript levels of genes encoding differentially expressed proteins (DEPs) observed during postharvest storage of broccoli. Relative transcript data from qRT-PCR were normalized to actin as an endogenous control using the formula of ΔΔCᴛ. The y-axis is relative expression of fold change to the housekeeping genes BoACTIN. Values of transcript expression are shown as average ± SE.
The 12 DEPs selected for qRT-PCR evaluation included a thioredoxin, cytochrome P450, a subunit from the light harvesting complex PS II, a plastid protein, and a serine methyltransferase. Consistent with the proteomics data, the transcript levels of those genes were higher after 3 and 5 d of storage at room temperature (25 °C) in comparison to the control (day 1), but were unchanged during 5 d of cold storage (4 °C). Several of the DEPs are predicted to be part of the nucleosome complex, namely the DEK domain chromatin-associated protein, homeobox-3, and a zinc finger protein, but their transcript levels varied under both storage temperatures. For instance, transcript levels of the DEK domain protein decreased from day 1 to day 3 but increased on day 5 at 25 °C, while at 4 °C, the levels increased by day 3 but declined by day 5. Additionally, the homeobox gene exhibited relatively higher expression levels on day 5 in comparison to day 1 at both temperatures. The transcript levels of a few genes involved in developmental regulation, including embryogenesis-related genes, a TCP chaperonin gene, a temperature-induced lipocalin, and SNF7, were also validated (Fig. 4). The transcript levels of lipocalin were lower on day 5 at room temperature compared to cold storage. The chaperonin and LEA transcripts were high at room temperature and showed a significant transition from day 1 to day 5, but remained unchanged in cold storage.
Identification of the key pathways involved in postharvest senescence
-
To further identify the key pathways and crucial proteins during tissue senescence, we utilized the Kyoto Encyclopedia of Genes and Genomes (KEGG) database and conducted pathway enrichment analysis (Supplemental Table S5). We found several networks that are directly related to senescence, for example, ribosome synthesis, the spliceosome, and carbon metabolism.
Ribosomal protein changes
-
Our pathway enrichment analysis suggested that ribosome proteins were downregulated during storage at the lower temperature, resulting in slower protein synthesis during postharvest senescence (Fig. 5). Ribosomal proteins are involved in translation efficiency, ribosome stability, and other cellular processes, including DNA repair, apoptosis, and regulation of gene expression[16]. Our study identified numerous ribosomal proteins associated with both the smaller and larger subunits that were differentially regulated (Fig. 5). This finding suggested that ribosomal protein synthesis is inhibited at the lower temperature. However, determination of the precise functions of these small and large ribosomal protein subunits in postharvest senescence requires further research. Additionally, photosynthesis and plant-pathogen interaction pathways were also down-regulated in the lower temperature storage conditions (Supplemental Table S5).
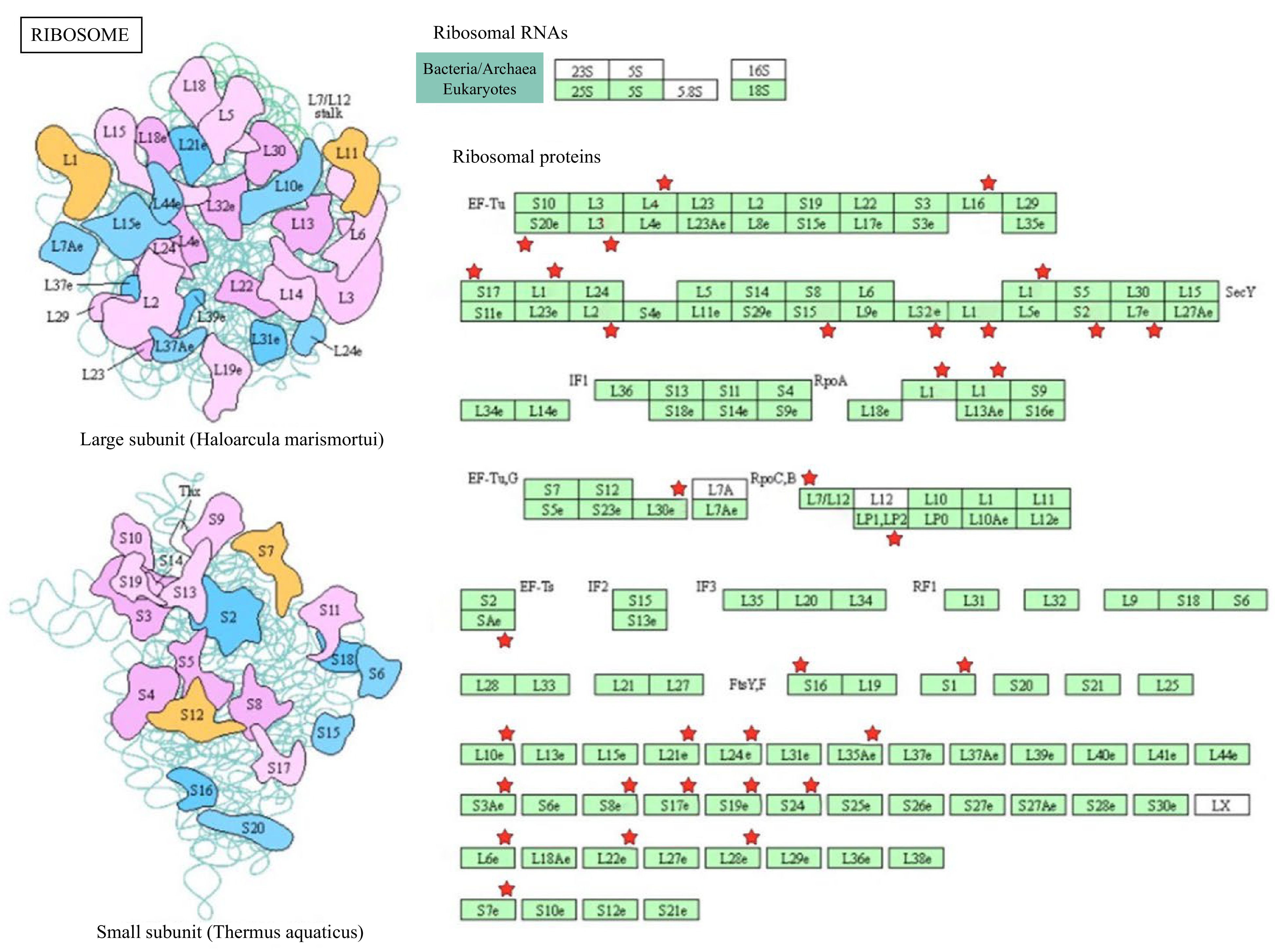
Figure 5.
Differential expression and KEGG pathway analysis of proteins related to ribosome biosynthesis in broccoli during postharvest storage. The DEPs are indicated by red stars.
We speculated that the lower temperature inhibited photosynthesis and the activities of protein-proteins interaction to slow down senescence and to reduce pathogen infection. We also found that spliceosome proteins were up-regulated on day 3 of storage at the higher temperature. Recently, alternative splicing has been recognized as a critical contributor to senescence and aging[18]. The RNA-protein complexes that combine with unmodified pre-mRNA and various other proteins to form a spliceosome are known as snRNPs. The spliceosome consists of five snRNPs and over 200 additional proteins (Supplemental Fig. S1a). The spliceosome controls splicing of introns and requires the recognition and subsequent cleavage of genes at the 5' donor and 3' acceptor sites[19]. Splicing is one of the post-transcriptional, pre-processing steps of mRNA maturation, allowing translation into proteins. Splicing of pre-mRNAs is a key step in the regulation of gene expression, transcriptome plasticity, and proteome diversity in eukaryotes[20]. Several spliceosome components, including pre-mRNA splicing proteins, were up-regulated during storage at 25 °C, suggesting up-regulation of splicing during senescence (Supplemental Table S6). These proteins could be used as potential markers for the early stages of senescence. Several enriched genes were also involved in translation elongation factor activity, GTPase activity, GTP binding, and RNA transport (Supplemental Table S6).
Carbon metabolism
-
Several proteins associated with carbon metabolism and carbon fixation in photosynthetic organisms and protein processing in the endoplasmic reticulum (ER) were up-regulated during storage at 25 °C (Supplemental Fig. S1b, Supplemental Table S5). Most of the protein synthesis machinery occurs in the ER, a localized subcellular organelle where protein folding occurs via chaperones. Our results indicated that misfolded proteins were retained within the ER lumen during senescence. It also suggested that protein production during stress-induced senescence produces proteins that are terminally misfolded and directed toward degradation through the proteasome. This accumulation of misfolded proteins in the ER activated the unfolded protein response signaling pathway, leading to protein degradation. Additionally, we observed in our data that most of the carbon sequestering and fixation-associated genes were differentially regulated (Supplemental Fig. S1b). These DEPs were involved in several energy-requiring processes, suggesting the active processes speed up the progression of senescence. The affected metabolic pathways included carbon metabolism, glycolysis, and the Kreb's cycle. During the ongoing senescence process, when subjected to prolonged storage at room temperature, fresh broccoli florets underwent a series of complex physiological processes, allowing the activation of stress-associated genes and proteins, leading to degradation of chloroplasts and reduction in other accessory pigments[13].
ROS signaling
-
There were several DEPs related to oxidative phosphorylation pathways that were downregulated during storage at 25 °C. Senescence involves accumulation of reactive oxygen species (ROS), increasing energy metabolism and carbohydrate metabolism during storage. Accumulation of ROS in the cells generates free radicals, which cause damage to cells and macromolecules, including DNA, proteins, and nucleic acids. Under stress, pathways associated with protein modification are upregulated to counter any loss or damage that occurs during the changing environmental conditions. Most of the proteins associated with protein metabolism belong to the thioredoxin family, which are responsible for maintaining the redox status of proteins targeted during development[21]. Thioredoxins function in developing plant resistance toward stress conditions. Differential expression of certain proteins, including Rubisco, suggested that most of the carbohydrate pathways were involved in the senescence process. We observed certain DEPs involved in carbohydrate-related metabolism during biosynthesis of glycolysis components and pyruvate metabolism, and in the functioning of photosystems during energy biosynthesis (Supplemental Table S5). In this study we found that the cellular machinery involved in stress metabolism was differentially expressed during room temperature storage.
Differentially expressed proteins and altered chemical interaction networks observed during postharvest storage
-
We predicted potential relationships among the DEPs in broccoli stored under the two different storage temperatures using the STRING web tool (
https://string-db.org/ ), which provides an integrated, differential network of proteins from metabolic pathways as well as chemical-protein and protein-protein interactions. Relationships observed during postharvest senescence were mapped (Fig. 6). Cysteine was the most prominent amino acid observed to interact with diverse DEPs (Bra000159, Bra34356, Bra032828, Bra002411, and Bra40980), forming a strong network and suggesting that cysteine regulates a wide variety of metabolic processes, including protein biosynthesis. It is also responsible for regulating Coenzyme A, which is an important intermediate for the electron transport chain, photosynthetic complexes, and the C3 cycle.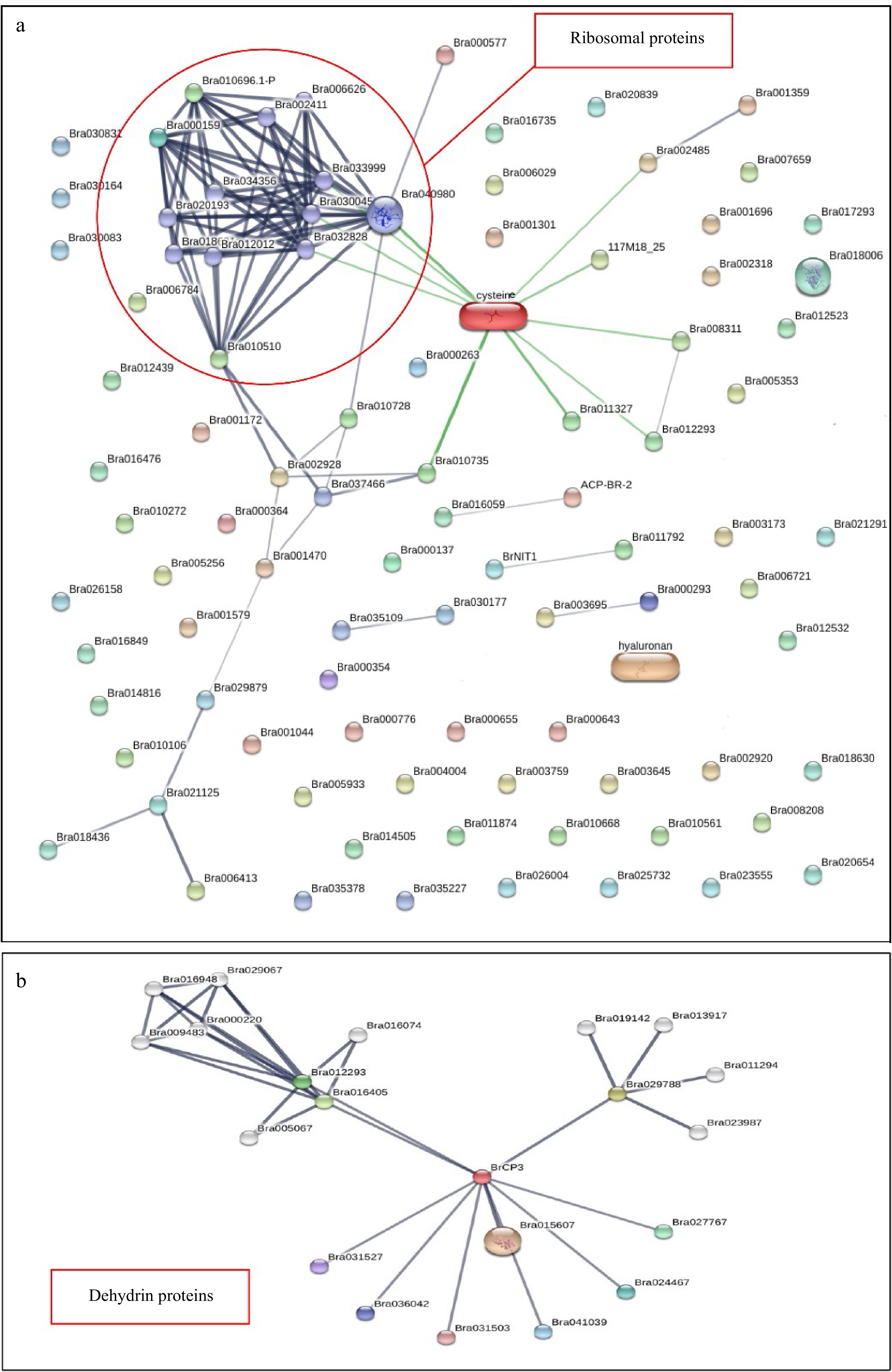
Figure 6.
Investigating the protein-protein interaction network of differentially expressed proteins in broccoli during postharvest storage at 25 and 4 °C. Protein interaction networks generated using the STRING software showed that (a) ribosomal proteins and (b) dehydrin proteins were hubs during postharvest senescence in broccoli.
Several key protein networks were identified from this analysis (Fig. 6a). For instance, Bra040980, which encodes a chloroplast ribosomal protein that is a constituent of the large subunit of the ribosomal complex, was a hub for protein interactions. Furthermore, some stress-responsive proteins, such as the antioxidant proteins superoxide dismutase, peroxidase, and thioredoxin, were also included in this interaction network. Another protein, Bra021125, which encodes Glutaredoxin 4 (GRX4), a metal ion binding protein that is involved in iron homeostasis, had several contacts with several antioxidant proteins including Bra029879, which encodes a manganese superoxide dismutase 1; Bra018436, a hydrogen peroxide catabolic gene; and Bra006413, a redox stress related gene (Fig. 6a). In addition, some proteins involved in RNA binding and translation initiation were also part of this network.
Some dehydrin proteins were also activated during room temperature storage (Fig. 6b). Dehydrins are late embryogenesis abundant (LEA) proteins involved in environmental stress responses in higher plants[22]. The expression of dehydrins occurs in response to diverse stresses such as cold, drought, high salinity, and pathogen infection[23,24]. These are mostly plant-specific proteins, although some LEA proteins have been found in other kingdoms accumulating in the cytoplasm, nucleus, plastid, mitochondrion, ER, and plasma membrane[25]. Functional genomics studies have shown the participation of dehydrins in promoting stress tolerance. Dehydration-responsive proteins such as Brassica rapa cysteine protease 3 (BrCP3), a cysteine-type peptidase involved in various stress response mechanisms, was also present in this network (Fig. 6b). BrCP3 was a hub for this protein interaction network. Within this network, we found BrCP3 interacts with a programmed cell death-related protein, Bra015607 (a homolog of AtSERPIN1, At1g47710), and a serpin family Bra031527. These proteins are parts of intricate regulatory networks that showcase the remarkable adaptability of plants to their environment[26]. Understanding their functions and interactions can provide insights into the molecular pathways that plants use to counteract the negative effects of dehydration and other abiotic stresses (Fig. 6b).
Overexpression of dehydrins promotes tissue senescence
-
To test the function of those dehydrin proteins that are activated during broccoli storage, we identified the Arabidopsis homologs of the broccoli gene HIRD1 (AT1G54410) and harpin (HrpN)-interacting protein (a homolog of Arabidopsis gene AT1G15395). Both of those proteins were found in this study to be significantly differentially expressed (Supplemental Table S5). The genes were overexpressed through transient transformation of tobacco leaves, and the resulting phenotypes were analyzed to determine if any of the genes were involved in tissue senescence. We hypothesized that overexpression of the identified genes would accelerate leaf senescence in tobacco epidermal tissue (Fig. 7). In this assay, the negative control did not show obvious changes until 9 d after injection. We selected as a positive control little zipper 3 (ZPR3), which showed a strong senescence pattern in our previous study[4]. Similarly, we found that the dehydrin protein HIRD11 and the HrpN-interacting protein caused bleaching of leaf color after infiltration, suggesting that these two genes are involved in tissue senescence (Fig. 7). Dehydrins, specifically the Arabidopsis dehydrin HIRD11, exhibit cryoprotective activities, protecting freeze-sensitive enzymes such as lactate dehydrogenase (LDH) from damage during freezing conditions, and this cryoprotective ability is attributed to the presence of hydrophobic amino acids and certain conserved segments within the dehydrin protein[27]. One of the typical phenomena observed in transgenic plants expressing dehydrins is the reduction of lipid peroxidation under stress conditions[28]. While dehydrins protect the plants against cold stress[29], their in vivo mechanisms remain mostly unknown. Mutation of hrpN caused a significant decrease in virulence of the plant pathogen Erwinia amylovora[30]. When externally applied to Arabidopsis, a non-host plant, partially purified HrpN promoted plant growth[31].

Figure 7.
Tobacco transient assay. Images of tobacco leaves injected with the empty pMDC43 vector as a negative control; with ZPR3, a senescence-inducing protein, which caused color bleaching in tobacco leaves after injection; and with AtHIRD11 or a HrpN-interacting protein, both of which showed color bleaching in tobacco leaf, demonstrating induction of senescence symptoms.
-
The onset of yellowing in broccoli florets during postharvest storage was investigated through proteome profiling using peptide-tagging mass spectrometry to reveal dynamic changes at the global proteome levels. Use of proteomics can provide crucial information for understanding the physiological processes and metabolic pathways associated with the storage of horticultural crops. Insights gained from such studies can help optimize efficient postharvest management practices, potentially leading to significant reductions of economic losses due to crop deterioration. This proteomics study identified protein kinases, transcription factors, and dehydrin proteins that contribute to postharvest senescence in broccoli. Degradation of structural and functional components, as observed through pathway enrichment analysis, contributed to the more rapid senescence mechanism under higher temperature. Identification of key biological markers, such as kinases and dehydrin proteins, will be helpful in providing insights about stress responses and quality traits and could be used to assess quality during storage and handling of biological commodities. This information is crucial for controlling the timing of senescence and for preventing premature deterioration. Our findings provide key insights into the molecular mechanisms underlying temperature-related storage treatments in broccoli. Incorporating proteomics into postharvest biology research enables a comprehensive understanding of the molecular changes that influence the quality characteristics of broccoli during storage and transportation. This knowledge can lead to improved postharvest management practices, reduced food waste, and better utilization of agricultural resources.
-
The authors confirm contribution to the paper as follows: study conception and design: Liu T, Brecht JK, Pliakoni ED; protein extraction and qPCR: Ahlawat YK; proteomics data analysis: Timilsena P, Li S; tobacco assay and some of the data analysis: Ghimire U; figure preparation and drafting the manuscript: Liu T, Brecht JK, Pliakoni ED, Ghimire U, Ahlawat YK. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
We thank Dr. Jin Koh at the Proteomics & Mass Spectrometry Core Research Facility of the UF Interdisciplinary Center for Biotechnology Research (ICBR) for performing the proteomics and raw data analyses. This work was supported by the USDA National Institute of Food and Agriculture, AFRI GRANT 2021-67021-34484 to U.G. and T.L. The contents do not necessarily reflect the views or policies of the USDA nor does mention of trade names, commercial products, services, or organizations imply endorsement by the US Government.
-
The authors declare that they have no conflict of interest. Tie Liu is an Editorial Board member of Vegetable Research who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
- Supplemental Tables S1 DEPs during cold treatment.
- Supplemental Tables S2 DEPs during room temperature treatment.
- Supplemental Tables S3 The primers used for validating expressions of DEPs observed in the proteomics study.
- Supplemental Tables S4 GO-term analysis of DEPs.
- Supplemental Tables S5 Oppositely DEPs during cold and room temperature treatment.
- Supplemental Tables S6 KEGG analysis of pathways among DEPs.
- Supplemental Fig. S1 Spliceosome proteins and protein processing in endoplasmic reticulum are differentially expressed in this cold storage. A. Spliceosome proteins pathway; B. Protein processing in endoplasmic reticulum.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ahlawat YK, Ghimire U, Li S, Timilsena P, Pliakoni ED, et al. 2024. Postharvest senescence profiling in broccoli using Tandem Mass Tag-based proteomics. Vegetable Research 4: e010 doi: 10.48130/vegres-0024-0009 

Postharvest senescence profiling in broccoli using Tandem Mass Tag-based proteomics
- Received: 01 December 2023
- Revised: 24 January 2024
- Accepted: 09 February 2024
- Published online: 02 April 2024
Abstract: Harvesting fresh produce induces countless changes in different metabolic processes, initiating senescence of the plant tissue. Understanding the changes in protein expression and activity during postharvest senescence can help identify the networks that contribute to quality loss and shelf-life reduction in produce. Identifying these proteins can lead to strategies for extending fruits' and vegetables' freshness and shelf life. In this study, we investigated the proteome-level changes in harvested broccoli florets stored at either room temperature (25 °C) or in refrigerated conditions (4 °C) using Tandem Mass Tag technology (TMT). A lower storage temperature reduced the levels of proteins involved in ubiquitin-dependent and ubiquitin-independent protein degradation pathways mediated by the 20S and 26S proteasomes and in the RNA-Induced Silencing Complex (RISC) associated with the small interfering RNA (siRNA) pathway. Moreover, we identified protein kinases, transcription factors, and dehydration-related proteins, as well as mapped chemical interaction networks and key pathways active during broccoli senescence. Pathways related to autophagy, ribosome biosynthesis, and carbon metabolism were significantly downregulated under cold temperatures. Proteins related to chlorophyll and reactive oxygen species (ROS) pathways were differentially expressed in harvested broccoli stored at room temperature. The current report facilitates a comprehensive understanding of the complex regulatory orchestration that builds a protein network that could be used for future endeavors in improving the postharvest shelf life of broccoli.
-
Key words:
- Postharvest /
- Senescences /
- Broccolis /
- Proteomics