









-
Using chemical pesticides and fertilizers has promoted the development of modern agriculture. However, the irrational use of pesticides and fertilizers not only leads to the imbalance of soil nutrients, low yield of crops, poor quality and reduced stress resistance, but also causes environmental pollution[1]. Biostimulants can boost plant growth, improve crop quality, and alleviate abiotic stress, which mainly includes humic acids, seaweed extracts, amino acids, chitin, chitosan, and microbial agents[2]. Biostimulants can improve the utilization rate of fertilizers or enhance the efficacy of pesticides, and regulate the damage caused by the unreasonable use of fertilizers and pesticides to the soil microenvironment, thereby improving the growth of crops[3,4]. For instance, humic acid promotes plant growth via regulating the activity of H+-ATPase to enhance the absorption and transport of nutrients in roots[5]. Seaweed extracts increase plant photosynthetic capacity by enhancing phosphatase and nitrate reductase accumulation. Chitooligosaccharides increase the content of secondary metabolites related to low-temperature resistance in wheat[6]. Microbial agents increase crop yield by enhancing respiration and increasing microbial biomass in soil[7,8]. Biostimulants have diverse origins and distinct functions in regulating crop growth and development.
Chinese herbal medicines are taken from nature and have the advantages of being green, having no residue, and having low toxicity. Chinese medicinal herbs have only been studied in detail for fungal inhibition, medical treatment, and addition to livestock feeds[9]. Previous studies have shown that secondary metabolites derived from Chinese medicine herbs play an important role in regulating plant growth and development[10]. Artemisia argyi extracts significantly inhibit the stem and root length of Setaria viridis and Portulaca oleracea, thereby reducing their biomass[11]. Caffeic acid is an important allelochemical extracted from Artemisia argyi, which down-regulates multiple genes involved in gibberellin, phytoalexin biosynthesis, and mitogen-activated protein kinase signaling pathways to inhibit weed growth[12]. The allelopathy of Artemisia argyi extracts is manifested in inhibiting seed germination and seedling growth of Chinese cabbage, lettuce, and rice[13]. As traditional Chinese herbal medicine, Scutellaria baicalensis Georgi ('Huangqin' in Chinese) has a wide range of pharmacological effects and high medicinal value in clinical practice. Freezing injury reduced the content of baicalin, wogonoside, and scutellarin and increased the content of baicalein, wogonin, and scutellarein in S. baicalensis[14]. FNSII, GUS, and UBGAT as key enzyme genes play important roles in baicalin biosynthesis[15]. Its root is an important medicinal organ of S. baicalensis[16]. Baicalin, belonging to flavonoids and is an important bioactive component in S. baicalensis, which can effectively inhibit the reproduction and growth of Aspergillus niger, Aspergillus oryzae, Aspergillus fumigatus, Candida albicans, Candida glabrata, and Candida tropicalis[17]. To explore new ways to promote crop growth via the application of new biostimulants, a comprehensive study on the regulatory effects of herbal extracts on crop growth and quality was carried out to provide theoretical and data support for practical production.
Biostimulants have been widely applied in crops to increase production and income[18]. However, the study of active compounds in S. baicalensis extracts as biological stimulants in regulating plant growth and development has not been reported. In the present study, the LC-MS/MS method was first utilized to analyze the crude extracts of S. baicalensis and it was verified that baicalin was the major active compound in the S. baicalensis extract. Next the S. baicalensis extracts and the actual standard of baicalin were applied to tomato seedlings at the vegetative growth stage via spraying foliar (S), root irrigation (R) or spraying foliar and root irrigation (SR) modes. Both S. baicalensis extracts and baicalin had very similar effects in promoting seeding growth and leaf photosynthetic efficiency. Overall, S. baicalensis extracts and baicalin were more effective on tomato seedings growth and development.
-
Seeds of tomato (Solanum lycopersium L.) cultivar Ailsa Craig germinated on nutrient agar medium for three days in incubator at 28 °C. When the radicle of the seed began to grow, these seeds were sown in nutrient substrates that contained vermiculite, perlite, and soil (1:1:1, v/v/v) under normal conditions. At the two-leaf stage, the uniform seedlings were selected and transferred to the nursery pot (10 cm × 10 cm × 10 cm, one plant per pot), and Hoagland's nutrient solution was used to water them every 3 d. All seedlings were cultured in a light incubator. The incubator conditions were set to a temperature of 25 °C, and a light-dark cycle of (16 h light:8 h dark), and a light intensity of 100 μmol/m2/s.
Experimental treatment of S. baicalensis crude extracts and baicalin
-
The dry slices of S. baicalensis roots were purchased from Taobao web (
www.taobao.com ). The S. baicalensis roots were placed in an oven at 50−60 °C for 2−3 h with forced-air drying. In the process of forced-air drying, the wind speed is 2.0−2.5 m/s. For experimentation of S. baicalensis crude extracts, dry slices of S. baicalensis were first washed with flowing water and then immersed in water for 1 h mixing every 10 min under normal lighting conditions. The slices of S. baicalensis were boiled in water at 100 °C for 1 h to obtain the active compounds. According to our pre-experiment, the primitive extracts of S. baicalensis were cooled down and diluted to a working concentration (6 g dry S. baicalensis slices per liter) for subsequent treatment experiments. We utilized the same batch of S. baicalensis dry slices, and the concentration of baicalin was determined using LC-MS/MS. The amount of 1 g S. baicalensis dry slices contained about 7.7 mg baicalin. The baicalin purchased from Shanghai Yuanye Bio-Technology Co., Ltd (Shanghai, China) was dissolved in ultrapure water and diluted to 0.046 g/L. The six uniform tomato plants with four leaves were tested for each experiment. In this experiment, four treatments (application modes) were conducted as follows: (1) S: spraying S. baicalensis extracts or baicalin on leaves and irrigating water on roots. (2) R: spraying water on leaves and irrigating S. baicalensis extracts or baicalin on roots. (3) SR: spraying S. baicalensis extracts or baicalin on leaves and irrigating S. baicalensis extracts or baicalin on roots. (4) CK represents spraying water on leaves and irrigating water on roots (Fig. 1). Plastic sprinklers with 1 cm diameter round nozzles were used for the spraying. Plants were treated once every 3 d, a total of five treatments. Then, the physiological indexes of the treated plants were measured. Each treatment included six biological replicates.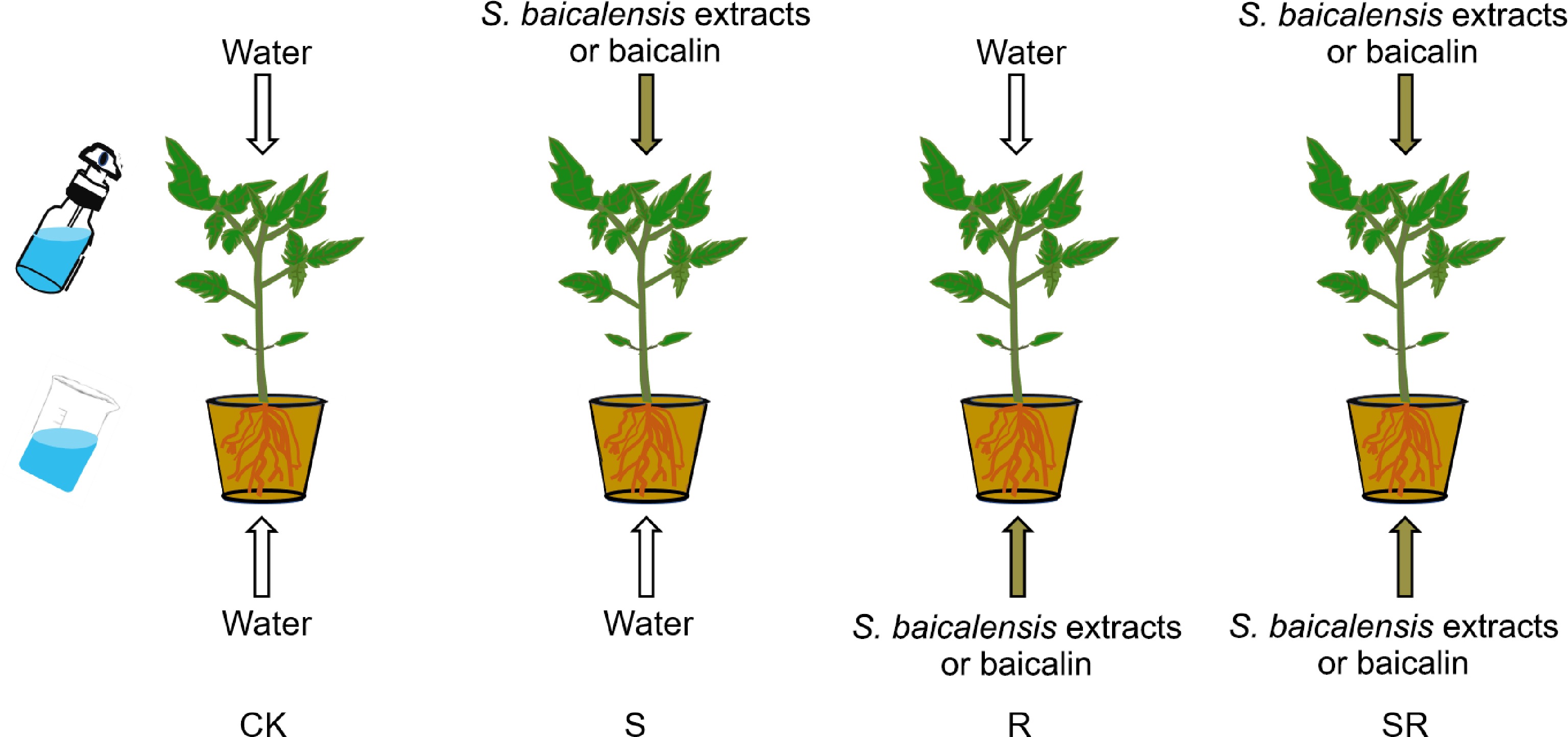
Figure 1.
Four treatment modes were performed as follows: (1) CK: spraying water on leaves and irrigating water on roots. (2) S: spraying S. baicalensis extracts or baicalin on leaves and irrigating water on roots. (3) R: spraying water on leaves and irrigating S. baicalensis extracts or baicalin on roots. (4) SR: spraying S. baicalensis extracts or baicalin on leaves and irrigating S. baicalensis extracts or baicalin on roots. In this study, CK was the control group.
Identification of the main active compound in S. baicalensis extracts
-
The major active compound baicalin in S. baicalensis crude extracts was determined by LC-MS/MS (LCMS-8050) using the actual standard of baicalin (cas#: 21967-41-9; purity: 90%) (Xu et al.[19]). Based on the standard curve generated for baicalin via LC-MS/MS, the concentration of baicalin in the original S. baicalensis extract solution was calculated to be 0.77 g/L (Supplemental Fig. S1).
Plant growth and photosynthetic pigment measurement
-
The tomato plants with treatments were photographed. ImageJ software was used to determine plant height, stem diameter, and leaflet area after treatments. The length from the surface of the substrate to the top of the stem was used as the plant height. The average plant height was obtained from six individuals. Leaf area was measured from individual leaves. The fourth fully developed leaves were quick-frozen with liquid nitrogen and then ground into a fine powder. Then, photosynthetic pigments were extracted from 100 mg powder with 80% acetone[20]. Six plants of each treatment were analyzed.
Photosynthetic capacity index and chlorophyll fluorescence of leaves
-
The net photosynthetic rate (Pn), stomatal conductance (Gs) and transpiration rate (E) of the fourth leaf of tomato seedlings with treatments were measured on a sunny day from 9:00 a.m. to 12:00 noon using the CIRAS-3 Portable Photosynthesis System. The maximum photochemical efficiency (Fv/Fm) and actual quantum efficiency (PSII) of tomato leaves after dark treatment was measured for 30 min using an FMS-2 portable fluorometer. Six plants of each treatment were analyzed.
Root scanning experiment
-
The WINRHIZO multi-parameter plant root analysis system was used to scan the roots of tomato seedlings after S. baicalensis extracts and baicalin treatment then the total root length, surface area, volume, diameter, number of nodes, and number of root tips were counted. Six plants of each treatment were analyzed.
Measurement of tissue fresh weight and dry weight
-
The aboveground and underground tissues were separated from the plants, and then the fresh weight (FW) of tissues was quantified using balance. The aboveground and underground tissues were placed in envelopes and then dried at 105 °C for 0.5 h, cooled to 60 °C and dried for an additional 2 d in an oven.
Nuclear DNA ploidy level analysis
-
Leaf and stem tissues of tomato seedlings were used for extracting the nucleus. About 50 mg of fresh tissue was collected, added with 1 mL cell lysis buffer, and then chopped with a blade. The buffer contained 10 mM MgSO4·7H2O, 50 mM KCl, 5 mM 4-Hydroxyethylpiperazine ethane sulfonic acid (HEPES), 2.5% Triton X-1006, and 5 mM Dithiothreitol (DTT). The nuclei were stained with propidium iodide (PI) at a concentration of 50 μg/mL. The cell cycle was determined and calculated by BD flow cytometry (BD Biosciences, San Jose, California, USA) and FlowJo_v10 software (Tree Star, Inc., Ashland, OR, USA), respectively, according to the previously reported methods[21].
RNA isolation and expression analysis
-
Tomato seedlings were treated with S. baicalensis extracts to detect the expression of cyclin genes. Total RNA was extracted with the RNAiso Plus (TaKaRa, Otsu, Japan, cat. #108-95-2) from stems. The first-strand cDNA synthesis was conducted as described in the manufacturers instructions. qRT-PCR were conducted using the ABI QuantStudio 3 (Applied Biosystems, USA). ACTIN (SGN-U580609) was used as the internal control[22]. Three biological replicates were detected for each treatment.
Statistical analysis
-
The differences among experimental groups were assessed using DPS software. Principal component analysis (PCA) and clustering analysis were performed in R Studio Version 1.1.456 (
www.rstudio.com ) with R version 3.5.2 using ggplots (www.cran.r-project.org/web/packages/gplots ) and factoextra (www.cran.r-project.org/web/packages/factoextra ) packages, respectively. Six plants of each treatment were analyzed. -
LC-MS/MS was used to determine whether baicalin, major active compound could be identified in the S. baicalensis extracts using the chemical baicalin as the actual standard. In negative mode, the detected parent ion for the actual standard of baicalin was 445 ([M-H]−) and its predominant product ion was m/z 269 (C15H9O5−) with optimized collision energy (CE) at 25 V (Fig. 2a). Therefore, the optimized mass pair for baicalin was 445/269 (Fig. 2b), which was then used in the multiple reaction monitoring (MRM). According to the retention time (RT) of baicalin standard (RT = 4.0 min) and the LC chromatogram of S. baicalensis extracts, two large peaks (RT = 3.97 min; RT = 4.67 min) in S. baicalensis extracts were obtained and it was confirmed that the first peak (RT = 3.97 min) was baicalin and the second peak (RT = 4.67 min) was probably an isomer of baicalin (Fig. 2c).
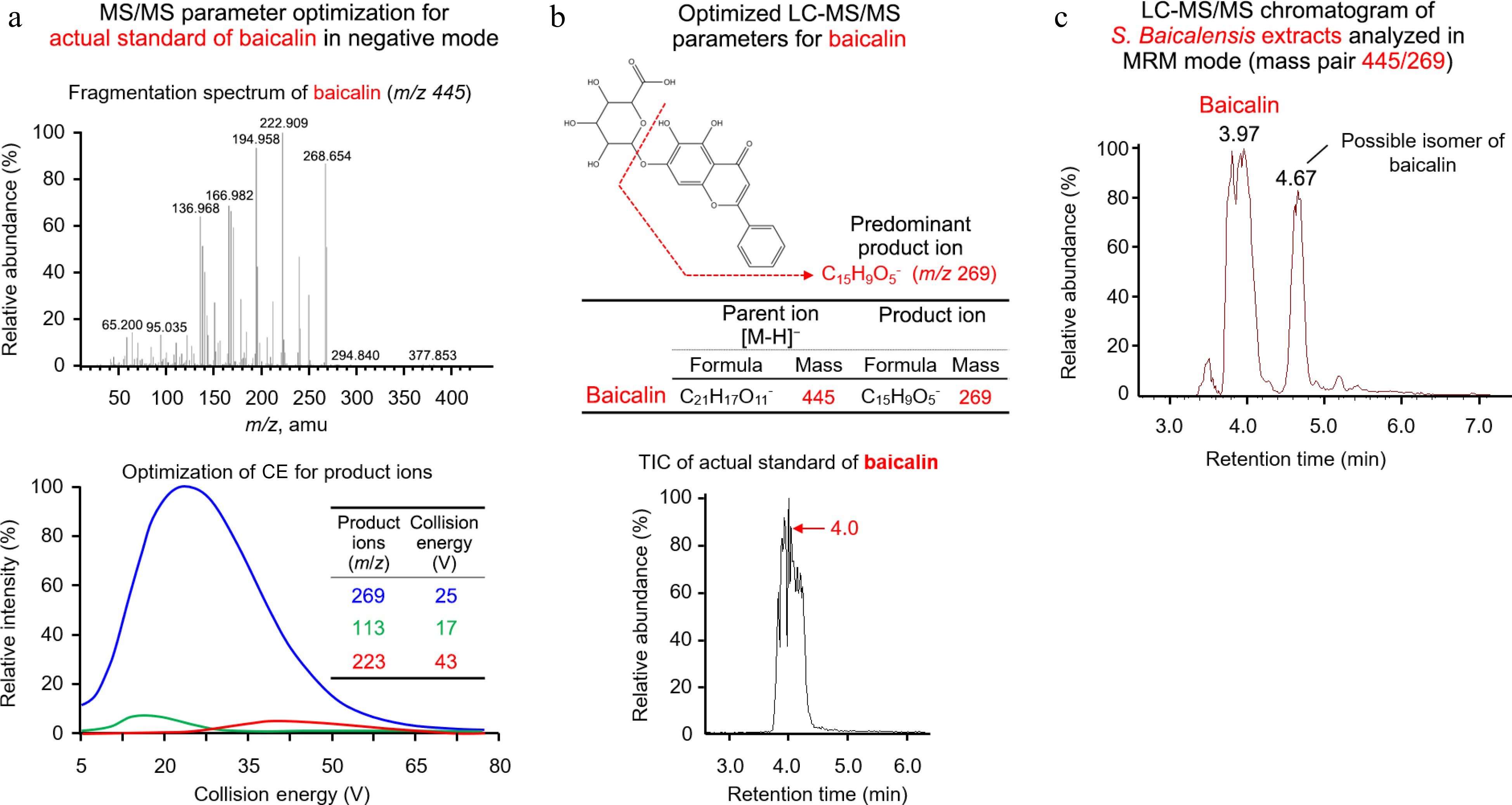
Figure 2.
Mass spectrometry analysis showed that baicalin was the main active ingredient in S. baicalensis extract. (a), (b) Direct infused baicalin standard was used to optimize compound-dependent MS/MS parameters and create a multiple reaction monitoring (MRM) method on LC-MS/MS. (c) The predominant active compound in S. baicalensis extract was confirmed to be baicalin. The two large peaks at 3.97 min and 4.67 min were isomers of baicalin.
Both S. baicalensis extracts and baicalin increase tomato plant height and stem diameter
-
Several runs of preliminary experiments were performed to confirm the suitable concentrations of working solutions of S. baicalensis extracts and the actual standard of baicalin. It was determined that the suitable working solution concentration of S. baicalensis extracts was 6 g/L. According to the standard curve of baicalin, the concentration of baicalin in S. baicalensis extracts was 0.77 g/L. According to the dilution ratio of S. baicalensis extracts, the working concentration of baicalin was calculated to be 0.046 g/L. So, it was finally decided to use S. baicalensis extracts (6 g dry slices/L) and baicalin (0.046 g/L) to treat tomato seedlings and compare their effects on plant growth (Supplemental Fig. S1). Plant height, stem diameter, and leaflet area of tomato seedlings after five treatments were measured. It was found that S. baicalensis extracts promoted tomato growth compared with control (Fig. 3a). Specifically, S. baicalensis extracts in SR treatment enhanced plant height, stem diameter, and leaflet area by 8.00%, 9.22% and 21.84% compared to the control, respectively (Fig. 3b−d). Baicalin as the main active ingredient of S. baicalensis extracts can also significantly promote the growth of tomato seedlings (Fig. 3e). For example, the plant height, stem diameter, and leaflet area of the SR group under baicalin treatment were 18.72%, 11.32%, and 1.81% higher than those of the control group, respectively (Fig. 3f−h). The cyclin genes can affect cell growth, division, and differentiation by affecting the cell cycle, thereby regulating plant development[23]. The cyclin genes transcription in stems of tomato seedlings treated with S. baicalensis extracts and water indicate that S.baicalensis extracts may enhance cyclin expression to promote tomato seedling growth via increasing endoreduplication (Fig. 3i). Hereby, these data suggest that S. baicalensis extract and baicalin have similar effects on tomato development. Meaningfully, SR treatment is more conducive to tomato development.
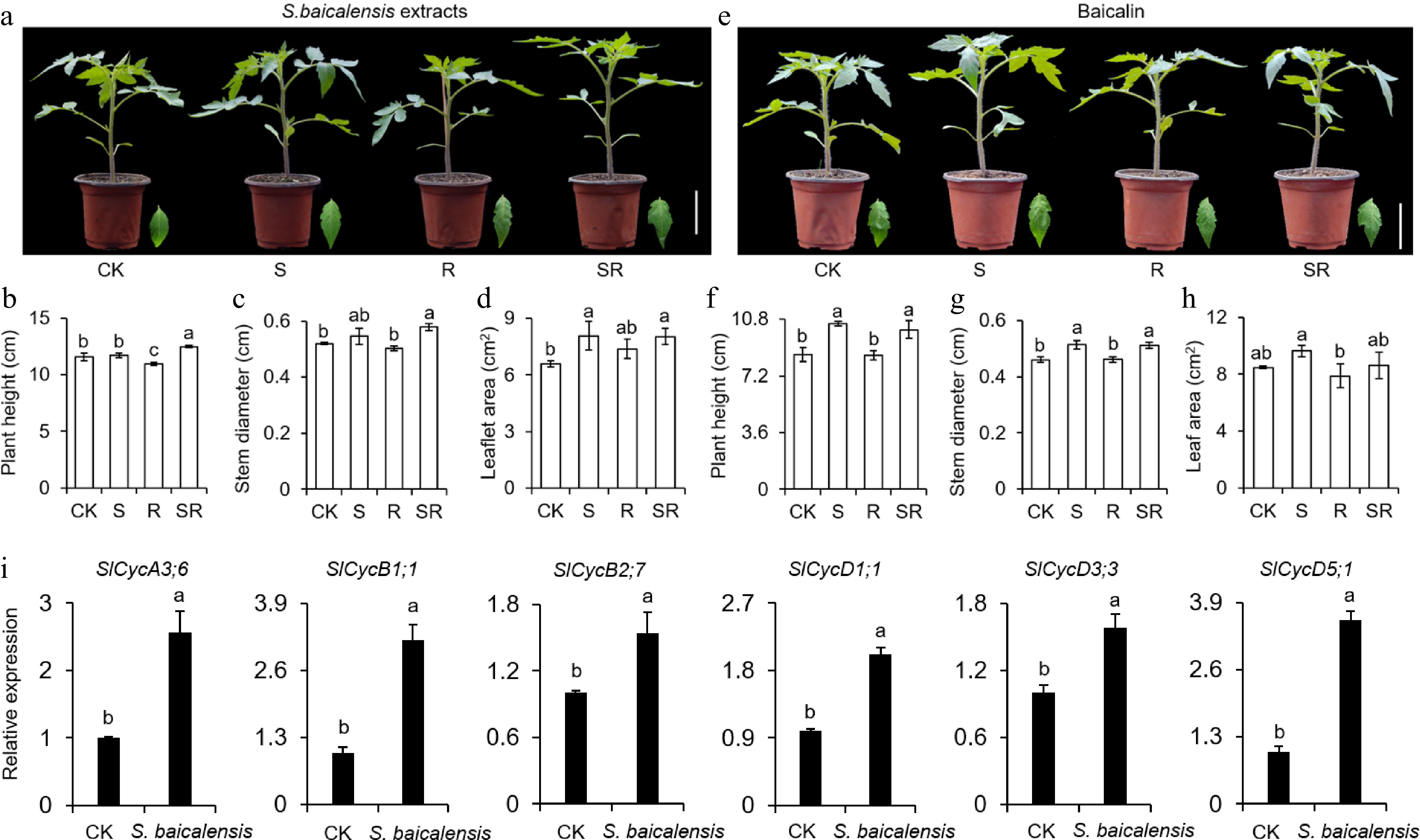
Figure 3.
Scutellaria baicalensis Georgi (S. baicalensis) extracts and baicalin affect height, stem diameter and leaflet size of tomato seedlings after treatments. (a), (e) Morphology of tomato seedlings after application of S. baicalensis extracts and baicalin. Scale bar = 5 cm. Treatments: Water control (CK), Spray (S), Root irrigation (R) and Spray and root irrigation (SR). (b), (f) Tomato plant height after application of S. baicalensis extracts and baicalin. (c), (g) Tomato stem diameter after application of S. baicalensis extracts and baicalin. (d), (h) Tomato leaflet size after application of S. baicalensis extracts and baicalin. (i) Analysis of cyclin genes expression in stems of tomato seedings treated with S. baicalensis extracts. Results are shown as means ± SD (n = 6). Values marked with the same letter within a sampling date are not significantly different at p < 0.05 according to Duncan's new multiple-range test.
Both S. baicalensis extracts and baicalin enhance photosynthesis of tomato leaves
-
To test whether S. baicalensis extract and baicalin influence the photosynthetic performance of tomato leaves, tomato seedlings were treated with S. baicalensis extracts and baicalin and determined their photosynthetic pigment content and photosynthetic rate. Chlorophyll a content, chlorophyll content, photosynthetic rate, and Fv/Fm of tomato seedlings in S group treated with S. baicalensis extracts were 1.05 times, 1.05 times, 1.29 times and 1.03 times higher relative to CK, respectively (Fig. 4a−f). Furthermore, the chlorophyll a content, total chlorophyll content, photosynthetic rate, PSII, and Fv/Fm were increased by 5.69%, 4.54%, 18.75%, 138.17%, and 8.68% in S, 11.15%, 8.49%, 14.77%, 18.69%, and 6.77% in SR, respectively, compared with CK under the baicalin treatment (Fig. 4g−l). To determine the mechanism of S. baicalensis extracts enhancing photosynthesis, qRT-PCR was used to assess the expression of photosynthetic efficiency-related genes in leaves of tomato seedings treated with S. baicalensis extracts. Compared with the control, S. baicalensis extracts significantly activated expression of light-harvesting chlorophyll a/b-binding factor (SlLhcb1;1 and SlLhcb1;2)[24], rubisco activase gene (SlRca1)[25], and photosystem subunit (SlPsaE and SlPsaQ)[26] (Fig. 4m). In summary, S. baicalensis extracts and baicalin improved photosynthesis of tomato seedlings by increasing chlorophyll and carotenoid contents.
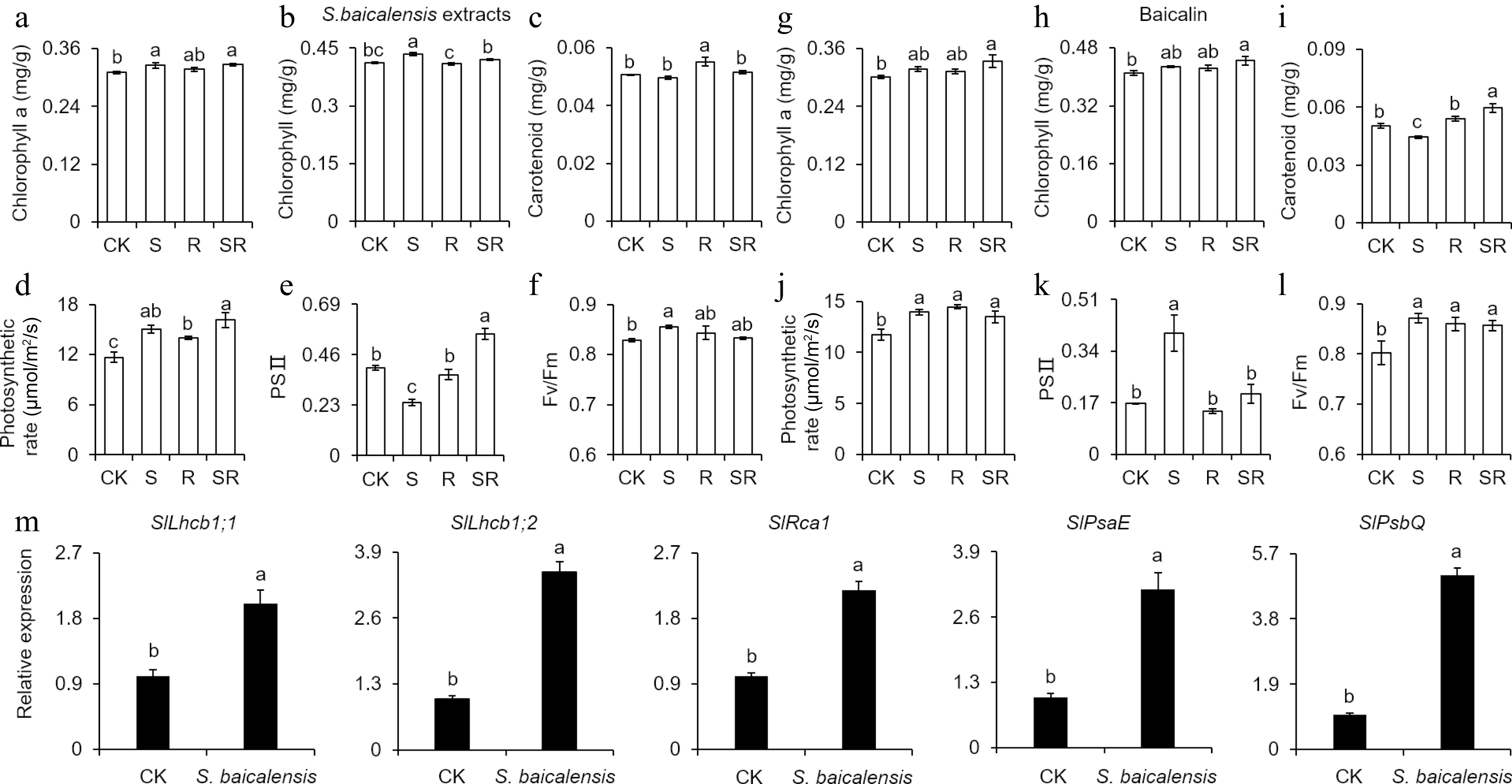
Figure 4.
Leaf chlorophyll content and photosynthetic rates of tomato seedlings in response to Scutellaria baicalensis Georgi extracts and baicalin treatments. (a), (g) Leaf chlorophyll a contents of tomato seedlings after application of S. baicalensis extracts and baicalin. (b), (h) Leaf chlorophyll contents of tomato seedlings after application of S. baicalensis extracts and baicalin. (c), (i) Leaf carotenoid contents of tomato seedlings after application of S. baicalensis extracts and baicalin. (d), (j) Photosynthetic rates of tomato seedlings after application of S. baicalensis extracts and baicalin. (e), (k) Actual quantum efficiency (PSII) of tomato seedlings after application of S. baicalensis extracts and baicalin. (f), (l) Maximum quantum efficiency (Fv/Fm) of tomato seedlings after application of S. baicalensis extracts and baicalin. (m) Expression of the photosynthesis-related genes in the CK and tomato seedings treated with S. baicalensis extracts. Results are shown as means ± SD (n = 6). Values marked with the same letter within a sampling date are not significantly different at p < 0.05 according to Duncan's new multiple-range test.
Both S. baicalensis extracts and baicalin boost tomato root growth
-
The morphological indexes of tomato roots were measured to investigate the effects of S. baicalensis extracts and baicalin on underground tissues. Tomato roots were scanned and it was found that S. baicalensis extracts promoted the root development (Fig. 5a). Under S. baicalensis extracts treatment, total root length, average root diameter, root surface area, root volume, root node number, and root tip number in the SR group increased by 29.91%, 3.39%, 27.48%, 23.03%, 42.31%, and 22.78%, respectively, compared with CK. On the contrary, R treatment significantly inhibited root growth. Furthermore, it was found that baicalin can also significantly improve root development of tomato seedlings (Fig. 5h). Total root length, average root diameter, root surface area, root volume, root node number, and root tip number were significantly increased in SR-treated tomato seedlings than in CK group. (Fig. 5i−n). These data provided evidence that S. baicalensis extracts and baicalin significantly strengthen root system architecture.
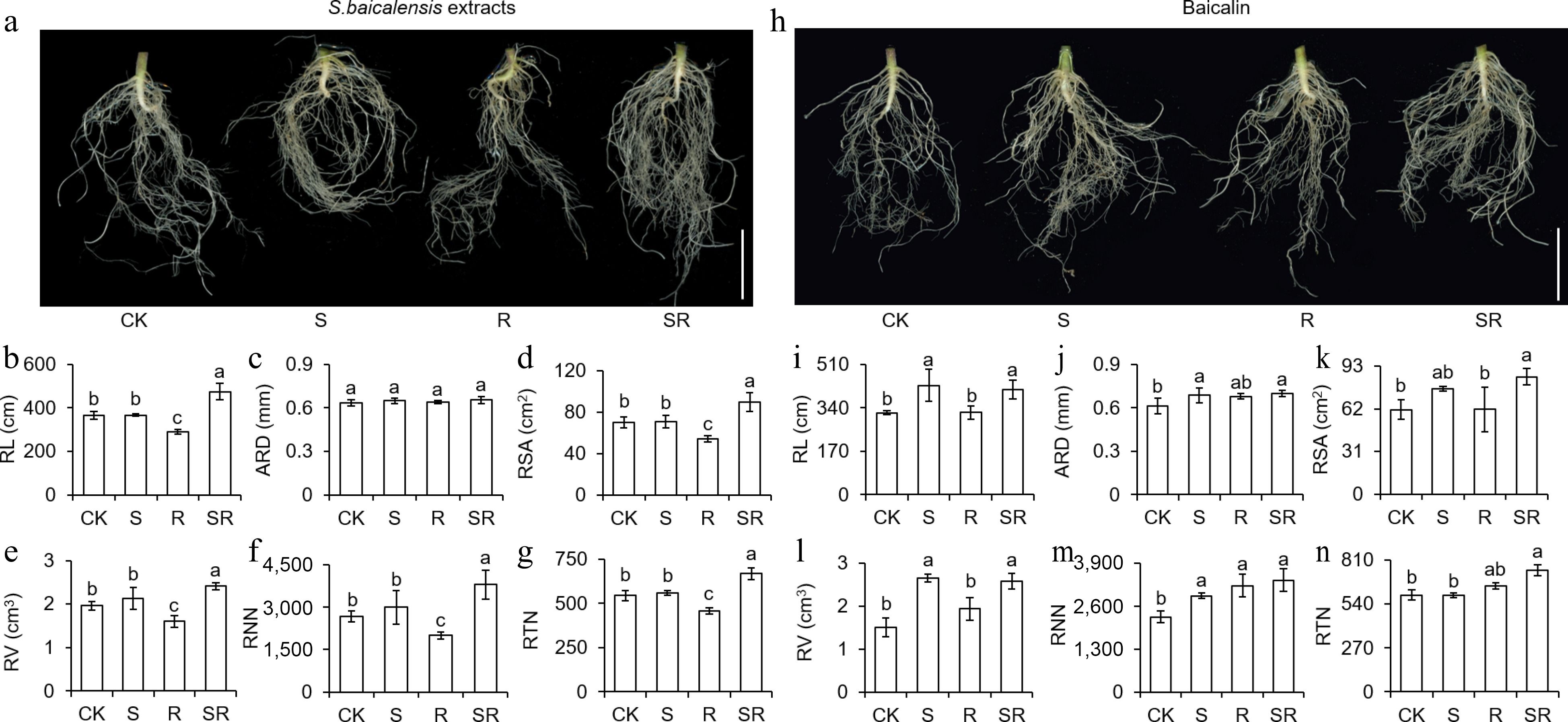
Figure 5.
Scutellaria baicalensis Georgi extracts and baicalin affect root physiological indicators after treatments. (a), (h) Morphology of tomato seedling roots after application of S. baicalensis extracts and baicalin. Scale bar = 2.5 cm. Total root length (RL), average root diameter (ARD), root surface area (RSA), root volume (RV), root node number (RNN) and root tip number (RTN) were measured after (b)−(g) application of S. baicalensis extract and (i)−(n) baicalin via WINRHIZO multi-parameter plant root analysis system. Results are shown as means ± SD (n = 6). Values marked with the same letter within a sampling date are not significantly different at p < 0.05 according to Duncan's new multiple-range test.
Both S. baicalensis extracts and baicalin increase tomato seedling biomass
-
The dry weight of tomato seedlings after S. baicalensis extracts and baicalin treatment were examined. Under S. baicalensis extracts treatment, compared with CK, SR treatment increased the dry weight of aboveground tissues by 26.65% (Fig. 6a). However, SR treatment slightly increased the fresh weight of aboveground tissues. Compared with the CK group, SR treatment increased the dry weight of underground tissue by 28.66% (Fig. 6b). Under baicalin treatment, S treatment increased the dry weight of aboveground tissues by 27.03% relative to CK. The dry weight of aboveground tissues in the S treatment were increased by 27.03% compared with CK, respectively (Fig. 6c). The S treatments increased dry weight of underground tissues by 32.92% compared with CK (Fig. 6d). Meanwhile, S. baicalensis extracts and baicalin also significantly increased the dry weight of tomato seedlings (Supplemental Fig. S2). Therefore, S. baicalensis extracts and baicalin significantly enhanced the biomass of tomato seedlings.
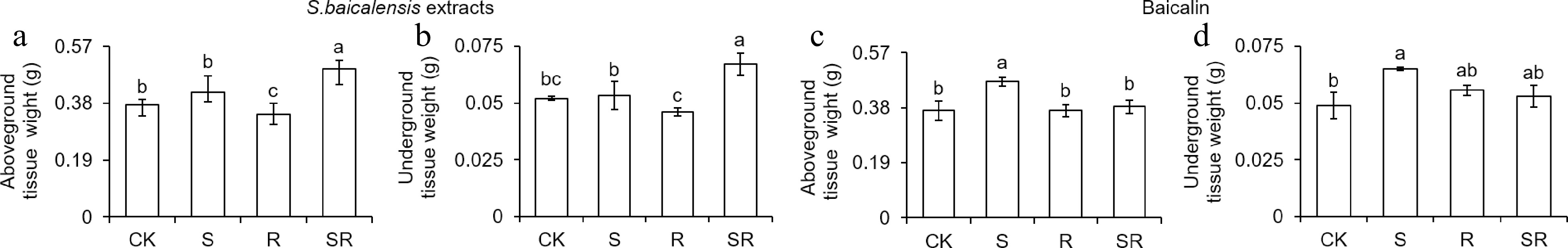
Figure 6.
Scutellaria baicalensis Georgi extracts and baicalin affect the aboveground and underground tissue dry weight of tomato seedlings. (a), (c) Dry weight (DW) of tomato aboveground tissues after S. baicalensis extracts and baicalin treatments. (b), (d) DW of tomato underground tissues after S. baicalensis extracts and baicalin treatments. Results are shown as means ± SD (n = 6). Values marked with the same letter within a sampling date are not significantly different at p < 0.05 according to Duncan's new multiple-range test.
Both S. baicalensis extracts and baicalin have an essential role in cultivating strong tomato seedlings
-
Root-shoot ratio and strong seedlings index (SSI) can be used to objectively evaluate the robustness of seedlings[27]. Therefore, the root-shoot ratio and SSI of tomato seedlings treated with S. baicalensis extract and baicalin were determined. Under S. baicalensis extracts treatment, the root-shoot ratio and SSI of the SR group were 1.62 times and 1.47 times that of CK, respectively (Fig. 7a, b). Under baicalin treatment, SR treatment significantly increased the root-shoot ratio and SSI of aboveground tissues, which were 58.60% and 18.50% higher than CK, respectively (Fig. 7c, d).
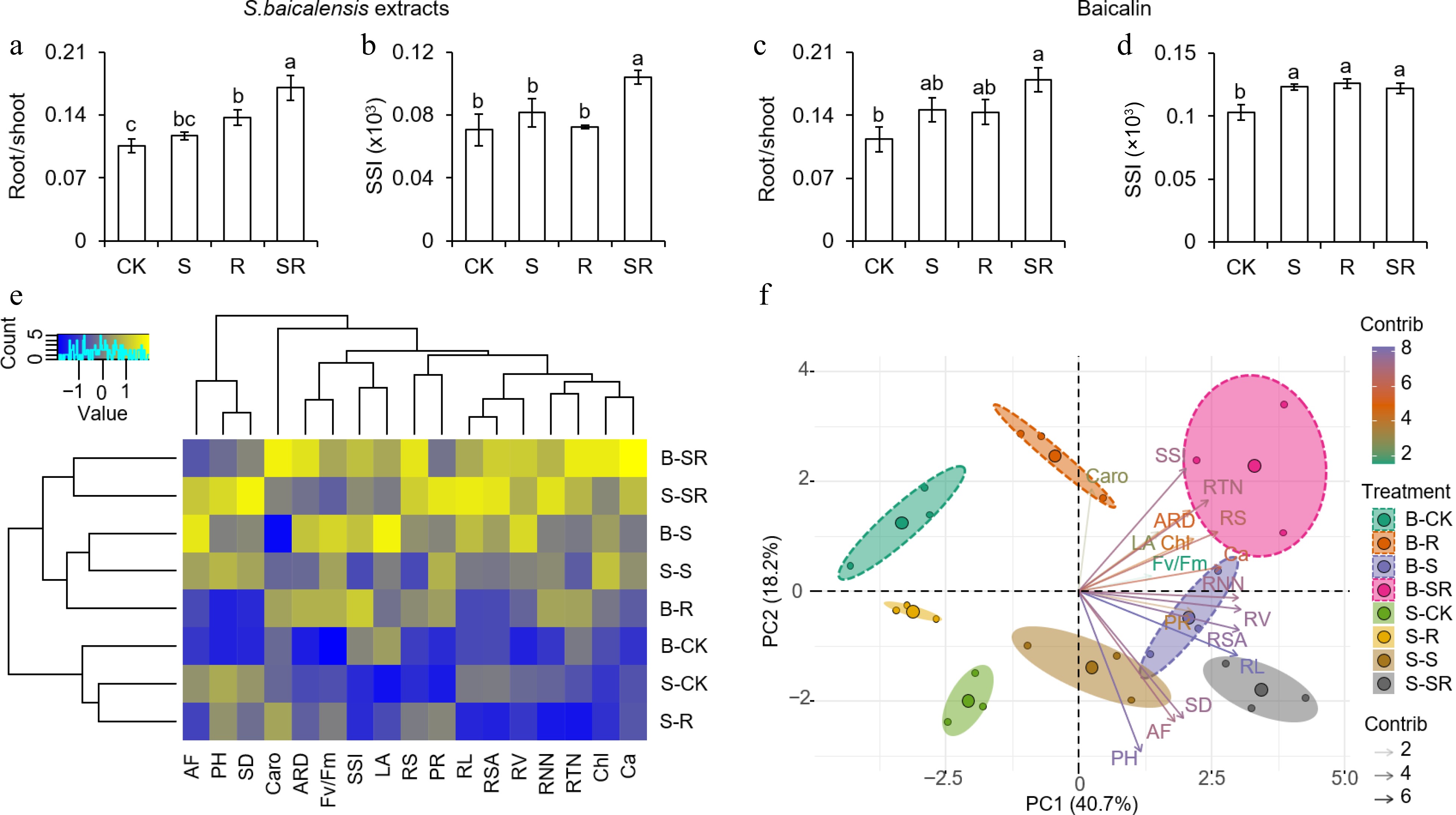
Figure 7.
Effects of different treatments of S. baicalensis extracts and baicalin on physiological data of tomato seedlings. (a), (b) The root: shoot ratio and strong seedling index (SSI) of tomato seedlings after application of S. baicalensis extracts. (c), (d) The root: shoot ratio and SSI of tomato seedlings after application of baicalin. Results are shown as means ± SD (n = 6). Values marked with the same letter within a sampling date are not significantly different at p < 0.05 according to Duncan's new multiple-range test. (e) Clustering analysis and (f) principal component analysis were used to analyze physiological indicators related to tomato seedling development. AF, aboveground fresh weight; PH, plant height; SD, Stem diameter; Caro, carotenoid content; ARD, average root diameter; SSI, strong seedling index; LA, leaflet area; RS, root: shoot ratio; PR, photosynthetic rate; RL, root length; RSA, root surface area; RV, root volume; Chl, chlorophyll content; Ca, chlorophyll a. B and S represent baicalin and S. baicalensis, respectively. Baicalin treating is marked using dotted lines. R programming was used for data analysis.
Cluster analysis and principal component analysis were conducted based on physiological data after S. baicalensis extracts and baicalin treatments. Compared with the control, SR treatment dramatically enhanced tomato seedling growth and development via cluster analysis. More meaningfully, SR treatment can be clustered together (Fig. 7e). Principal component analysis showed that the effect of SR treatment on tomato seedlings was the farthest from that of CK (Fig. 7f). These data showed that the effects of S. baicalensis extract and baicalin treatment on most physiological indexes of tomato seedlings were generally consistent, and showed the best effect under SR application. In general, the effect of S. baicalensis extract treatment was slightly better than that of baicalin treatment, which may be because S. baicalensis extract is a mixture and may contain other active compounds.
-
Biostimulants are organic compounds, inorganic compounds, or microorganisms that can improve crop resistance and improve crop quality. When applied to plant leaves or rhizosphere, biostimulants regulate physiological processes and nutrient uptake in plants[28]. Biostimulants have been widely used in horticulture and agricultural crops to increase yields. Humic acid increases the germination rate of tomato, wheat, and rice seeds and promotes the elongation of lateral roots, which in turn increases the yield and improves the quality of crops[4]. Chitooligosaccharides with foliar spraying can significantly accumulate proline in wheat and increase chlorophyll content in leaves to improve photosynthesis[29]. Chitosan can also inhibit the growth of pathogens in soil and improve soil aggregate structure, thereby increasing crop yield and quality[30]. In summary, the application of biostimulants in agriculture has achieved some important results.
Currently, biostimulants are of great value in efficient agricultural production and are therefore widely used to enhance crop productivity capacity[31]. The application of biostimulants with alginate and chitooligosaccharides as the main components are becoming more and more extensive[32]. Chinese herbal medicines possess the advantages of being green, pollution-free, and environmentally friendly[33]. However, application of Chinese herbal medicine extracts as plant-derived biostimulants in agriculture is rarely described. It was observed that the S. baicalensis extracts and baicalin significantly promote stem and root growth, increase tomato biomass, enhance leaf photosynthetic capacity, and eventually lead to the robust growth of tomato seedlings (Fig. 8). This is consistent with previous reports that the application of Boosten, Megafol, and Isabion significantly promoted plant growth and increased plant biomass[34].
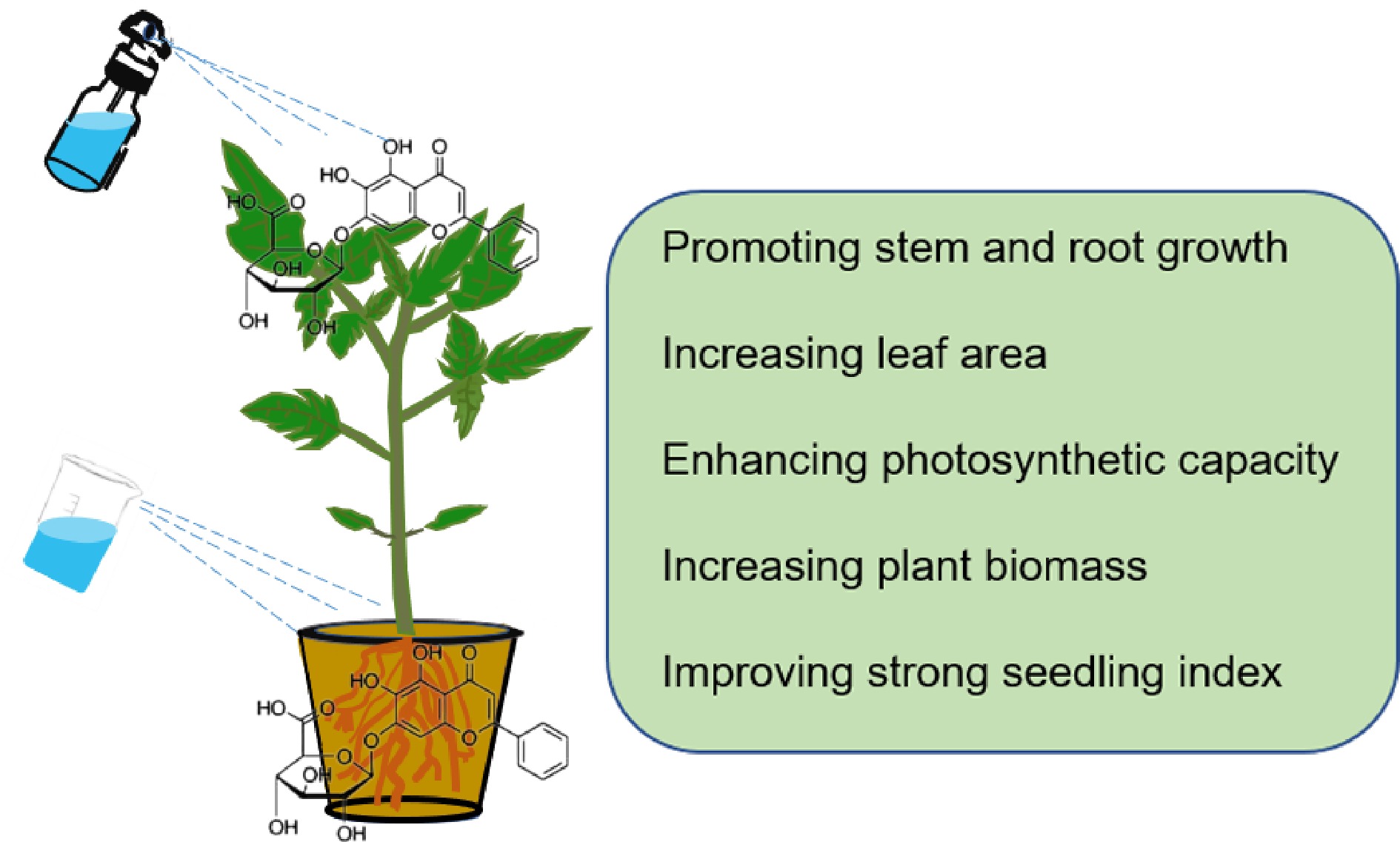
Figure 8.
Schematic illustration of a proposed model showing effects of S. baicalensis extracts and baicalin on growth and development of tomato seedlings.
The manner of biostimulant treatment is an important factor influencing plant development[35]. The main function of plant leaves is to carry out photosynthesis to produce organics, and can also absorb exogenous nutrients. Foliar spraying can make plants absorb and utilize biostimulants quickly and evenly, which can effectively improve the photosynthesis efficiency and ultimately promote the rapid growth of plants[36]. Plant roots are the foundation of life. Reasonable irrigation can directly provide proper biostimulants for plant roots, which significantly enhances the vitality of roots, thereby further improving the absorption capacity of plants for water and fertilizer[37]. Therefore, the combination of spraying and root irrigation can have an overall better effect on plant growth. Cluster analysis and PCA were carried out based on all phenotypic data with three treatments and it was found that the SR treatment was better in this study.
S. baicalensis, a natural herb, is rich in active substances that do not pollute the environment and cause pesticide residues[38]. Some studies have reported that Chinese herbal extracts affect plant hormone biosynthesis and their gene expression to regulate plant development[12]. However, there are few studies on the role of S. baicalensis in plant growth. Spraying and root irrigation with S. baicalensis extracts and baicalin can significantly promote the growth, stomatal conductance, and transpiration rate of tomato seedlings (Supplemental Fig. S3). Plants store energy through photosynthesis, which is conducive to the formation of crop yield and quality[39]. Chlorophyll and carotenoids of leaves participate in light absorption and transmission. Therefore, the content and composition of photosynthetic pigments play an important role in the photosynthetic rate of leaves[40]. The reason may be that S. baicalensis extracts and baicalin improve photosynthesis in leaves and ultimately make plants grow robustly. The root system has the function of absorbing, transporting, and storing nutrients, which determines the vigorous growth of plants[41]. S. baicalensis extract treatment can expand the root system (Fig. 5a). The accumulation of biomass is an important characterization of plant growth and metabolism[42]. S. baicalensis extracts treatment dramatically increased tomato seedling biomass (Fig. 6a, b). Chinese herbal medicine extracts play an active role in plant growth. As a plant-derived biostimulant, it does not produce drug residues on plants. Therefore, Chinese herbal medicine extract as a biostimulant has great prospects in agriculture. The photosynthesis of plants directly influences the synthesis and accumulation of organic matter, which is a direct factor affecting crop yield[43]. Interestingly, the expression of photosynthesis-related genes (SlLhcb1;1, SlLhcb1;2, SlRca1, SlPsaE, and SlPsaQ) was verified by qRT-PCR (Fig. 4). The results showed that the treatment of S. baicalensis extracts increased the expression of multiple photosynthesis-related genes, which was consistent with the increased photosynthetic rate of tomato after treatment with S. baicalensis extracts. Moreover, cyclins can regulate the division and differentiation of cells by affecting the cell cycle process, and ultimately modulate the biomass of plants[44]. Several Chinese herbal extracts have been confirmed to be involved in plant photosynthesis[19]. S. baicalensis extracts recognize a set of genes involved in cell cycle progression, by the increased endoreduplication of tomato stems (Fig. 3). The mechanism of S. baicalensis extracts enhancing photosynthetic capacity and increasing biomass requires further research.
As an exogenous non-nutritive chemical, biostimulants can be absorbed and transferred to different tissues of plants, affecting their growth and metabolic processes[45]. The effect of biostimulants are affected by many factors[46]. The low concentration also promoted tomato seedling growth and development without statistical significance, while the high concentration of the extract can inhibit the growth of tomato seedlings possibly due to overdosage. In conclusion, biostimulants are not equivalent to conventional fertilizers, but improve the physiological state of plants to affect plant growth and development[47,48]. Here, mass spectrometry was used to detect the possible active compounds in S. baicalensis extracts, which is baicalin. We then applied a certain dosage of baicalin on plants to compare its effects on plant development with S. baicalensis extracts and confirmed that baicalin indeed is the predominant active compound in the extract. The effect of S. baicalensis extracts on tomato seedling growth were slightly better than that of baicalin, which may be due to the fact that S. baicalensis extracts as a mixture may contain other active components. At the vegetative growth stage of the tomato seedlings, both S. baicalensis extracts and baicalin treatments promoted seedling growth, improved leaf photosynthetic efficiency and increased seedling index. These data suggest that the Chinese herbal extracts and their active compounds are important sources of biostimulants and play an important role in regulating plant growth and development, which may contribute to the development of sustainable agriculture.
-
The application of biostimulants in agricultural production may solve a series of problems such as environmental pollution caused by the excessive use of chemical fertilizers[49]. These results showed that S. baicalensis extracts and baicalin had very similar effects in boosting stem and root development, increasing leaf photosynthetic capacity, improving tomato biomass, and eventually contributed to the tomato seedling vigorous growth (Fig. 8). Therefore, S. baicalensis has the potential to serve as a source of biostimulants that can enhance resistance to abiotic stress and promote plant nutrient absorption.
-
The authors confirm contribution to the paper as follows: software, formal analysis, bioinformatics analysis: Liu G; conceptualization: Li J, Bao Z; investigation: Xin J, Li C, Ma M, Fan J, Xu C, Ma F; methodology, data curation: Xin J; funding acquisition: Liu G, Bao Z; resources: Fang D, Bao Z; supervision: Liu G, Ma F, Bao Z; writing–review & editing: Liu G, Bao Z. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This research was supported by the Taishan Scholar Foundation of Shandong Province (tsqn201812034), Agricultural Seed Project of Shandong Province (2020LZGC005), China Postdoctoral Science Foundation (2022M711967), Key research and development program of Shandong Province (2021LZGC017), and the National Natural Science Foundation of China (31872951).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Genzhong Liu, Jinyang Xin
- Supplemental Table S1 Primers used in this study.
- Supplemental Fig. S1 Preparation of crude extract solutions for application.
- Supplemental Fig. S2 Scutellaria baicalensis Georgi extracts and baicalin affect the aboveground and underground tissue fresh weight of tomato seedlings.
- Supplemental Fig. S3 The stomatal conductance and transpiration rate of tomato seedling leaves in response to Scutellaria baicalensis Georgi extracts and baicalin treatments.
- Supplemental Fig. S4 The endoreduplication level in tomato stem cells (A) and leaf cells (B) after Scutellaria baicalensis Georgi extracts treatments.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu G, Xin J, Li C, Ma M, Fan J, et al. 2024. Scutellaria baicalensis Georgi extracts and its active compound baicalin promote tomato seedling growth. Vegetable Research 4: e023 doi: 10.48130/vegres-0024-0023 

Scutellaria baicalensis Georgi extracts and its active compound baicalin promote tomato seedling growth
- Received: 27 December 2023
- Revised: 29 April 2024
- Accepted: 08 May 2024
- Published online: 16 July 2024
Abstract: Excessive use of chemical fertilizers and pesticides causes pollution of soil, water, and the atmosphere. Biostimulants are derived from natural sources, which can help plants absorb nutrients and promote plant development. Chinese herbal medicine extracts are enriched with bioactive compounds and therefore hold great potential for developing novel biostimulants. In this study, the predominant active compound, baicalin, was detected in Scutellaria baicalensis Georgi ('Huangqin' in Chinese) extracts via LC-MS/MS. To explore their effects, we used three different methods to treat tomato seedlings with Scutellaria baicalensis Georgi (S. baicalensis) extracts or baicalin, including foliar spraying (S), root irrigation (R), and the combination of foliar spraying and root irrigation (SR). Both S. baicalensis crude extracts and commercial baicalin promoted stem and root development, enhanced the photosynthetic capacity, and increased tomato seeding biomass, eventually making tomato seedlings grow vigorously. Clustering analysis and principal component analysis showed that S. baicalensis extracts and baicalin had very similar effects and showed the best effects in SR treatment. S. baicalensis extracts and its active compound baicalin could promote tomato seedling growth, suggesting that S. baicalensis is a potential source of biostimulants.
-
Key words:
- Biostimulants /
- Scutellaria baicalensis extracts /
- Tomato seedlings /
- Growth /
- Photosynthetic capacity /
- Biomass