









-
Hydroxyproline-rich glycoproteins (HRGPs) comprise a major group of structural proteins in the plant cell walls and are essential for plant growth and development[1,2]. The arabinogalactan proteins (AGPs), a subfamily of HRGPs, are highly glycosylated glycoproteins found in the plasma membranes, cell walls, extracellular spaces, and in secretions (e.g., stigma surface)[3]. The β-glucosyl Yariv reagent (β-Glc Yariv) and anti-AGP monoclonal antibodies can specifically interact with AGPs and are used to clarify their functions[4]. β-Glc Yariv binds AGPs through β-1,3-galactan chains[5]. In plants, AGPs participate in fruit ripening[6,7]. Typically, classic AGPs contain three characteristic domains: N-terminal secretion signal, Pro/Hyp-rich domain, and a C-terminal glycosylphosphatidylinositol (GPI) plasma member anchor. Classical AGPs include the Lys-rich classical AGPs, arabinogalactan (AG) peptides, and chimeric AGPs[1].
Fasciclin-like arabinogalactan proteins (FLAs) are classified as chimeric AGPs. Apart from one or two AGP motifs, FLAs possess one or two specific fasciclin (FAS) domains containing conserved regions H1 and H2 regions and contribute to cell adhesion and communication[1,8−11]. Structural analysis of AtFLA4 revealed that the FAS1 domain is required to stabilize plasma membrane localization, while the GPI-modification signal facilitates membrane attachment and endoplasmic reticulum (ER)-exit; N-glycosylated acts for ER-exit, while O-glycosylation affects post-secretory fate[12].
Numerous FLA genes have been identified across various plant species, with examples including 21, 27, 34, and 19 FLAs in Arabidopsis (Arabidopsis thaliana), rice (Oryza sativa), wheat (Triticum aestivum), cotton (Gossypium hirsutum), respectively[1,10,11,13,14]. FLAs play crucial roles in plant growth and development, such as fiber development[4,15−17], sexual reproduction[18−20], cell wall regeneration[21], and responsiveness to environmental conditions[22−24]. Notably, FLA genes have been extensively investigated in Arabidopsis. For instance, AtFLA18 regulates root elongation[25]; AtFLA12 and 16 contribute to stem development and differentiation[26,27]. Additionally, AtFLA3 and AtFLA14 are specifically expressed in pollen grains, playing an essential role in pollen development[18,28]. Ceitain FLA genes respond to stresses and plant hormones. For instance, FLA4, also known as salt overly sensitive 5 (SOS5), is sensitive to salt stress and affects cell-to-cell adhesion[29]. This gene acts within a linear pathway alongside FEI1/2, the leucine-rich repeat receptor-like kinases (RLKs), to modulate cell expansion dependent on ACC-mediated signals[30]. Drought leads to reduced expression of FLA9, and increases seed abortion in maize and Arabidopsis[23]. Consequently, FLAs are likely to play pivotal roles in plant growth and development. However, the exact underlying mechanism remains unclear.
Melon (Cucumis melo L.) is a vital commercial horticultural crop worldwide, belonging to the Cucurbitaceae family. Melon cultivars exhibit diversity, their fruit vary in size, shape, fresh color, rind color, texture, aroma, flavor, and nutritional value[31]. The function of FLAs in melon remains uncertain. Therefore, comprehensive bioinformatics analysis and identification of FLAs should be conducted within the melon genome.
In the present study, we identified 13 putative FLA genes in the melon genome and conducted a comprehensive analysis of CmFLAs using a bioinformatics approach. This analysis contained gene structure, motifs, physiochemical properties, domain analysis, chromosomal localization, subcellular localization, and phylogenetic comparison with other species. Additionally, we determined the expression patterns of these FLAs using publicly available RNA-seq data. Subsequently, we conducted qRT-PCR analysis to validate the expression of CmFLAs in melon fruits and to investigate their response to ABA signaling pathways. Finally, we further investigated the function of CmFLA8 in the regulation of sugar accumulation in melon fruit. This study offers fundamental insights into CmFLAs and may expedite the functional analysis of FLA genes in melon.
-
All the sequence data of Cucumis melo L. cv. DHL92 (.fasta files) was downloaded from the cucurbit genomics database (
http://cucurbitgenomics.org ). Proteins containing FAS domains were identified using several prediction algorithms (http://pfam.sanger.uk , PF02469,http://smart.embl-heidelberg.de , SM00554, andwww.ebi.ac.uk/interpro , IPR000782). Subsequently, the N-terminal signal sequences of these proteins were assessed using SignalP 5.0 (www.cbs.dtu.dk/services/SignalP )[32]. The presence of C-terminal GPI-addition signals was predicted using the big-PI Plant Predictor (https://mendel.imp.ac.at/gpi/plant_server.html )[33]. After filtering out proteins without N-terminal sequences, the remaining proteins with FAS domains were manually inspected for potential AGP-regions. Ultimately, proteins containing both FAS domains and AGP-like glycosylated regions were considered as CmFLA. The proportions of Pro, Ala, Ser, and Thr (PAST) are the putative CmFLAs were calculated using a Perl script[1].We utilized ExPASy (
https://web.expasy.org/protparam ) to analyze the physical and chemical parameters of proteins[34]. The subcellular localization of CmFLAs was determined using WoLF PSORT (www.genscript.com/tools/wolf-psort )[35]. N-glycosylation and O-glycosylation of the putative FLAs were investigated using NetNGlyc (www.cbs.dtu.dk/services/NetNGlyc )[36] and NetOGlyc (www.cbs.dtu.dk/services/NetOGlyc )[37]. Additionally, GO annotation of CmFLAs, describing biological processes, cellular components, and molecular functions, was obtained from the cucurbit genomics database (http://cucurbitgenomics.org ).Chromosomal localization and gene structure analysis
-
All of the gene information (gff3 files) was downloaded from cucurbit genomics (
http://cucurbitgenomics.org ). The chromosomal distribution of CmFLA genes was established using the positions of CmFLA genes from the melon database and visualized using TBtools software[38]. Furthermore, the conserved motifs of these genes were examined using NCBI Batch CD-Search[39] and the MEME program (https://meme.nbcr.net/meme )[40]. The CDS and gene structure were also determined using TBtools software[38].Alignment and phylogenetic analysis
-
The fasciclin-like domain sequences of CmFLAs were generated using ClustalX (2.1) and were manually corrected[41]. A phylogenetic tree was constructed using the neighbor-joining method in MEGA7 software[42] and EvolView version 2 (
www.evolgenius.info/evolview-v2 )[43], with a bootstrap value of 1,000.Plant material and growth conditions
-
Various varieties of melon plants (Cucumis melo L., including cv. Balengcui (BLC), cv. Baicaigua (BCG), cv. Huapicaigua (HPC), cv. Xiaomaisu (XM), cv. Yangjiaosu (YJ), cv. Longtian No.4 (LT4), cv. Gaoshicui (GS), cv. M43) were grown in polytunnels in Wuhan, China, from March to June. Melon fruits were harvested 30 d after anthesis (DAA) for qRT-PCR and sugar quantification. All fruit samples were promptly frozen in liquid nitrogen and stored at −80 °C before usage.
RNA-seq analysis
-
RNA-seq data from the database of cucurbit genomics (
https://cucurbitgenomics.org/ ) were analyzed. PRJNA383830, which contains samples from diverse tissues of melon, and PRJNA286120, covering different developmental phases of green and orange fruit flesh, were used for analysis. The heatmap was created using TBtools software[38].Cis-acting elements analysis
-
The 2 kb upstream region of each CmFLA gene was extracted as the promoter region, and the potential cis-elements were analyzed in PlantCARE database (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html )[44]. The diagrams were generated using the TBtools software[38].Total soluble solids content (TSS) evaluation
-
The TSS of each melon fruit was measured by dropping the extracted juice from the equatorial region of flesh tissue onto a digital refractometer. The value reported for each sample represents the average of three replicates.
ABA treatment
-
Melon fruit of the cv. 'Boyang' at 20 DAA were chosen for ABA treatment, the fruit discs were immersed in a solution of 100 μM ABA, with water used as the control. The discs were shaken at 25 °C for 8 h, with each treatment having three biological replicates. Following three washes with water, the samples were prepared for further analysis.
β-Glc Yariv treatment
-
Melon fruits of the cv. 'BCG' at 20 DAA were sprayed with a solution containing 200 mg/L β-Glc Yariv (Biosupplies, Australia), with water used as the control. Each treatment involved three fruits. Fruit samples were collected from the plants after 5 d of treatments for further analysis.
Sugar content measurements
-
Fruit samples, which were collected and stored at –80 °C, were ground to powder using liquid nitrogen. It was used for measuring the sucrose, fructose, and glucose content by gas chromatography (GC), following the method by Cheng et al.[45]. The ground samples (1 to 2 g) were extracted by ultrasonication in an extraction solution containing 80% ice-cold methanol and 0.1 M imidazole at 70 °C for 30 min. Subsequently, the mixture was centrifuged at 4,000 × g for 30 min. The supernatant was mixed with an internal standard (methyl-a-D-glucopyranoside, Sigma, USA) and was dried using Speed Vac (Eppendorf, Germany) at 45 °C for 2 h. The dried samples were derivatized using hydroxylamine hydrochloride: hexamethyldisilazane: trimethylchlorosilane before analysis by GC (Agilent 7890B).
Standard solutions of fructose, glucose, and sucrose (Sigma, USA) were prepared. Subsequently, standard curves and correlation coefficients were obtained as described above. Thus, we calculated the sugar content of the samples from standard curves.
RNA extraction and cDNA synthesis
-
Total RNA was extracted from melon fruits at 30 DAA using TRIzol. One μg of total RNA was utilized for cDNA synthesis with HighScript III reverse transcriptase after genomic DNA removal.
qRT-PCR analysis
-
The cDNA served as the template for qRT-PCR analysis, conducted on the ABI7500 system (Bio-Rad) using the SYBR green detection protocol (TaKaRa). TUA (α-tubulin) was used as a reference gene. Primers used for qRT-PCR analysis can be found in Supplementary Table S1. Finally, the mean expression level of relevant genes was calculated using the 2−ΔΔCᴛ method[46].
Subcellular localization
-
To determine the subcellular localization of CmFLA8, firstly, we used the primers (F: 5'-ATGGCGCCTTCCAAATCTC-3'; R: 5'-TTAGGACAATAAAAGGGAGG-3' ) to clone the CDS of CmFLA8. Then, the full-length CDS of CmFLA8 without a stop codon was inserted into pH7LIC6.0-ccdB rc-C-eGFP. The vector was linearized by Stu I and assembled through Gibson one-step assembly method. Subsequently, a transient expression method using A. tumefaciens (strain GV3101) was employed to introduce the vector into N. benthamiana leaves[45]. The resulting fluorescence was visualized using a confocal microscope (Leica SP8). GFP was excited with 488 nm, and fluorescence emissions of 495 to 531 nm were collected. Chloroplasts autofluorescence was excited with 552 nm, and fluorescence emissions between 644 and 697 nm were collected.
Agrobacterium-mediated transient transformation
-
For the overexpression of CmFLA8 in cv. 'XM' melon fruit, the full-length CDS of CmFLA8 was cloned into the pHELLSGATE8-HA vector. The vector was linearized by EcoR I and Kpn I, then assembled through Gibson one-step assembly method. Subsequently, the resulting recombinant plasmids were introduced into A. tumefaciens (strain GV3101). The cultures were then resuspended in an infiltration buffer containing 50 mM MES, pH 5.6, 10 mM MgCl2, and 1 mM acetosyringone, reaching an OD600 cell density of 0.3−0.4. This infiltration buffer was then injected into cv. 'XM' melon fruit at 15−20 DAA to a depth of 0.5 cm. An empty vector served as the control. Each fruit underwent three injections, and each treatment was administered to four fruits. The infiltrated fruit was harvested 5 d post-injection, and the fruit flesh surrounding the infiltrated area was sampled for subsequent analysis.
-
A Perl script (amino acid bias) can hardly locate FLAs based on the low proportions of PAST residues[13]. Therefore, we utilized the hidden Markov model (multiple sequence alignments), as described by Schultz et al.[47], to detect proteins with low similarity. Subsequent searches in the Pfam (PF02469), SMART (SM00554), and InterPro (IPR000782) databases, 19 predicted melon proteins containing FAS domains were found. Among these, three genes (MELO3C008283.2, MELO3C005314.2, and MELO3C018783.2) were excluded using SignalP5.0 due to the absence of N-terminal signal peptides. We manually examined the presence of AGP regions. According to Schultz et al.[47], non-contiguous Hyp residues with no more than 11 amino acid residues between consecutive Hyp residues (e.g., [A/S/T]-P-X(0,10)–[A/S/T]-P) are likely to be galactosylated[11,47]. Additionally, contiguous Hyp residues, such as [A/S/T]-P-P and [A/S/T]-P-P-P, indicated arabinosylated[48,49]. Ultimately, 13 putative CmFLAs contained these glycomodules, with the exception of three CmFLAs genes: MELO3C024107.2, MELO3C028604.2, and MELO3C012299.2.
Detailed information about the 13 putative CmFLAs is presented in Table 1. The amino acid sequences consist of 243–466 amino acids, with each FLA containing one or two FAS domains and one or two AGP regions. Among the CmFLAs, five FLAs (CmFLA1, 2, 6, 8, and 9) possess a GPI anchor signal. All CmFLAs exhibit N-terminal secretion signals, with the possible cleavage sites specified in Table 1.
Table 1. Putative FLA genes identified in the genome of Cucumis melo. L.
Protein name Gene ID Chromosome location Strand Size (aa) FASs AGPs GPI SP Possible cleavage site CmFLA1 MELO3C024938.2 Chr01: 10881922, 10883631 − 414 1 2 Y Y 28~29 CmFLA2 MELO3C024192.2 Chr01: 4351699, 4354505 + 421 2 2 Y Y 30~31 CmFLA3 MELO3C015093.2 Chr02: 8392291, 8395918 + 466 2 2 N Y 26~27 CmFLA4 MELO3C009567.2 Chr04: 31139948, 31143044 + 465 2 1 N Y 26~27 CmFLA5 MELO3C019361.2 Chr11: 12828056, 12829226 − 263 1 2 N Y 23~24 CmFLA6 MELO3C018862.2 Chr01: 3459779, 3461694 + 247 1 2 Y Y 18~19 CmFLA7 MELO3C018888.2 Chr01: 3762092, 3763889 − 255 1 2 N Y 23~24 CmFLA8 MELO3C022507.2 Chr11: 34215140, 34216937 − 423 1 2 Y Y 18~19 CmFLA9 MELO3C020643.2 Chr12: 1931250, 1932848 + 352 1 1 Y Y 26~27 CmFLA10 MELO3C014385.2 Chr05: 2950327, 2951358 + 343 1 1 N Y 32~33 CmFLA11 MELO3C014248.2 Chr05: 4832311, 4833298 − 248 1 1 N Y 23~24 CmFLA12 MELO3C014107.2 Chr06: 36961275, 36962122 + 248 1 2 N Y 18~19 CmFLA13 MELO3C005051.2 Chr12: 3927483, 3929729 + 243 1 2 N Y 22~23 SP: Signal Peptide; aa: amino acid. The chromosome distribution of CmFLA genes in Cucumis melo L. was investigated. These genes were distributed on seven chromosomes in the melon genome, with four genes (CmFLA1, 2, 6, and 7) located on chromosome 01 (Fig. 1a). Additionally, the amino acid sequences of these proteins were aligned, and a phylogenetic tree was constructed (Fig. 1b). These genes were categorized into three clades: Clade I includes CmFLA1, 2, 5, 7, 8, 9, and 12; CmFLA6, 11, and 13 were clustered into Clade II; and Clade III comprises CmFLA3, 4, and 10 (Fig. 1b).
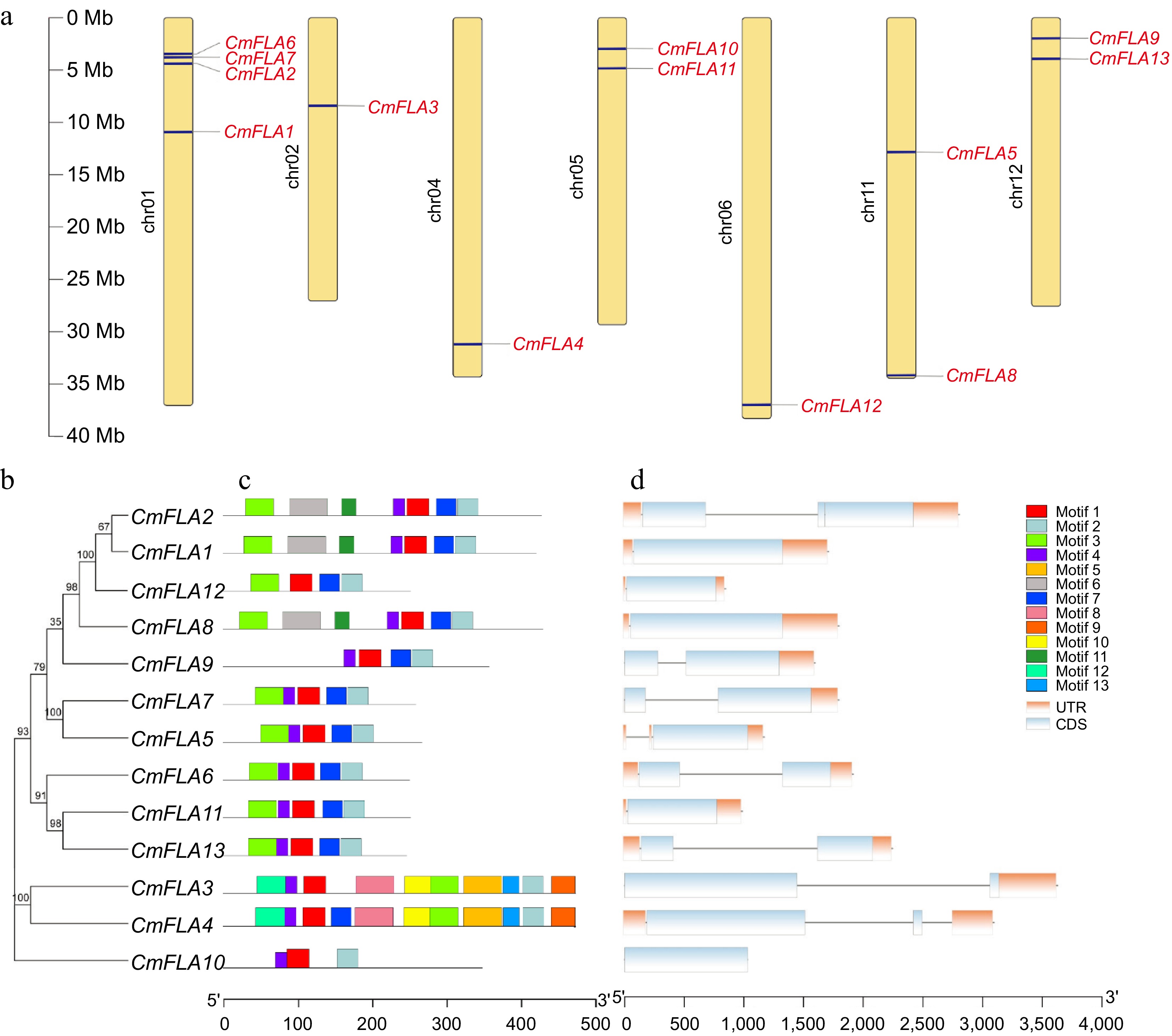
Figure 1.
Gene location, conserved motif, and gene structure analysis of CmFLAs. (a) Distribution of CmFLA genes on melon chromosomes. Thirteen genes were mapped to 7 out of 12 chromosomes. The dark blue lines represent the gene positions on the chromosome. The dark line in the left panel represents the chromosome scale. (b) The phylogenetic relationship among CmFLAs. (c) Conserved motif analysis of CmFLAs. Thirteen predicted motifs are represented by different colored boxes. The sequence information for each motif is provided in Supplementary Table S2. (d) Gene structure analysis of CmFLAs. Exons are denoted by light blue boxes, UTRs by orange boxes, and introns by gray lines.
The characteristic regions of CmFLAs were investigated via conserved motif prediction, leading to the identification of 13 motifs in melon FLA proteins (Fig. 1c; Supplementary Table S2). Motifs 1 and 2 were found in all CmFLAs, indicating their significance for FLA proteins. Moreover, motifs 3, 4, and 7 were present in most CmFLAs, indicating their necessity for FLA. Conversely, motifs 8, 9, 10, 12, and 13 were exclusive to CmFLA3 and CmFLA4, suggesting distinct functions compared to other CmFLAs. Notably, five CmFLAs (CmFLA5, 6, 7, 11, and 13) shared identical conserved motifs, implying similar function (Fig. 1c). Furthermore, CmFLA1, 5, 8, 10, 11, and 12 contained one exon, CmFLA3, 4, 6, 7, 9, and 13 included two exons, while CmFLA2 had three exons (Fig. 1d).
Characteristics of CmFLA proteins
-
CmFLAs were further characterized through an analysis of their molecular weight (MW), pI, GRAVY index, PAST content, putative O-glycosylation sites, putative N-glycosylation sites, and subcellular localization. The data are summarized in Table 2. The MW of CmFLA proteins ranged from 25.81 to 51.48 kD, while the pI value is within the range of 4.23 to 6.18. Among the CmFLAs, CmFLA2, CmFLA3, CmFLA4, CmFLA10, and CmFLA13 were identified as hydrophilic proteins, whereas the rest exhibited hydrophobic proteins.
Table 2. Characteristics of FLA proteins in melon.
Protein name MW (kDa) pI GRAVY PAST Number of
N-glycosylationNumber of
O-glycosylationPredicated subcellular localization CmFLA1 44.15 5.88 0.013 32.6% 6 14 Chloroplast CmFLA2 45.02 5.45 −0.011 31.5% 5 14 Chloroplast CmFLA3 51.48 6.18 −0.312 29.4% 2 15 Vacuolar CmFLA4 51.13 6.03 −0.287 29.4% 2 20 Chloroplast CmFLA5 27.92 5.57 0.197 38.4% 3 16 Cytoplasmic CmFLA6 25.81 5.62 0.071 33.6% 4 5 Chloroplast CmFLA7 26.91 5.32 0.139 40.0% 4 14 Plasma membrane CmFLA8 43.32 5.63 0.218 43.4% 5 35 Plasma membrane CmFLA9 37.47 5.06 0.319 33.0% 5 11 Extracellular CmFLA10 37.41 5.83 −0.041 38.5% 2 25 Chloroplast CmFLA11 25.94 5.14 0.028 35.6% 4 16 Chloroplast CmFLA12 26.27 4.23 0.087 34.8% 1 15 Chloroplast CmFLA13 26.05 4.73 −0.040 33.0% 4 6 Extracellular Notably, the proportion of PAST residues in CmFLAs ranged from 29.4% to 43.4%. All CmFLAs harbored putative N-glycosylation sites and putative O-glycosylation sites (Table 2). Subsequent analysis of putative subcellular localization revealed distinct patterns: CmFLA1, 2, 4, 6, 10, 11, and 12 were localized in the chloroplast, CmFLA9 and 13 in the extracellular domain, CmFLA7 and 8 in the plasma membrane, CmFLA3 in the vacuolar, and CmFLA5 on the cytoplasmic membrane (Table 2).
Protein structure and phylogenetic analysis
-
FAS domains consist of 110–150 amino acids and exhibit low sequence similarity[10]. Putative FAS domains within CmFLA proteins were aligned using the ClustalX program, revealing conserved regions depicted in the smart00554 motif, namely, the H1 and H2 conserved region, and the [Tyr Phe] His ([YF]H) motif situated between H1 and H2 regions. The Thr residue within the H1 region is completely conserved in the FAS -like domain[11]. All FAS domains of CmFLAs contained these conserved regions (Fig. 2a).

Figure 2.
Domain analysis of CmFLAs. (a) Multiple sequences alignment of the CmFLA FAS domains. Two FAS domains present in the protein. The domain closest to the N-terminus is indicated by .1, followed by .2. (b) Schematic representation of the CmFLAs deduced from DNA sequence. The FLAs are grouped into four (I–IV) based on the numbers of AGP-like regions and FAS domains. The indicated regions include the signal peptide (green), FAS-like domains (blue), AGP domain (red), and GPI-anchoring signal (purple box with a white point inside).
Considering the numbers of AGP-like regions and FAS domains as described by Xu et al.[30] with modification, these CmFLAs could be divided into four groups (Fig. 2b). Group I consisted of members with one FAS domain and two AGP regions, including CmFLA1, 5, 6, 7, 8, 12, and 13, while CmFLA9, 10, and 11 belonged to Group II, where one FAS domain was flanked by one AGP-like glycosylated region. Group III comprised CmFLA2 and CmFLA3, each containing two FAS domains and two AGP glycosylated regions. Finally, CmFLA4 was categorized into Group IV, characterized by two FAS domains with one AGP glycosylated region.
A comparison of the FLA protein homoeologous between Arabidopsis and melon reveals the amino acid identities of CmFLAs and AtFLAs, are detailed in Supplementary Table S3. The amino acid identity between AtFLA16 and CmFLA4 was 77.49%, while that between AtFLA16 and CmFLA3 was 79%. Two members exhibit high sequence homology with AtFLA7, namely CmFLA5 (71.71%) and CmFLA7 (77.33%, Supplementary Table S3). To explore the evolutionary relationships among the FLA genes of different species, a phylogenetic tree was constructed using the full-length sequence of 13 CmFLAs, 21 AtFLAs, 27 OsFLAs, and 19 GhFLAs (Fig. 3). These FLAs were divided into nine classes. Notably, CmFLA1, CmFLA2, AtFLA1, AtFLA3, and AtFLA14 belong to the same class. Additionally, CmFLA11, CmFLA13, AtFLA11, AtFLA12, and GhFLA1 are evolutionally close (Fig. 3).
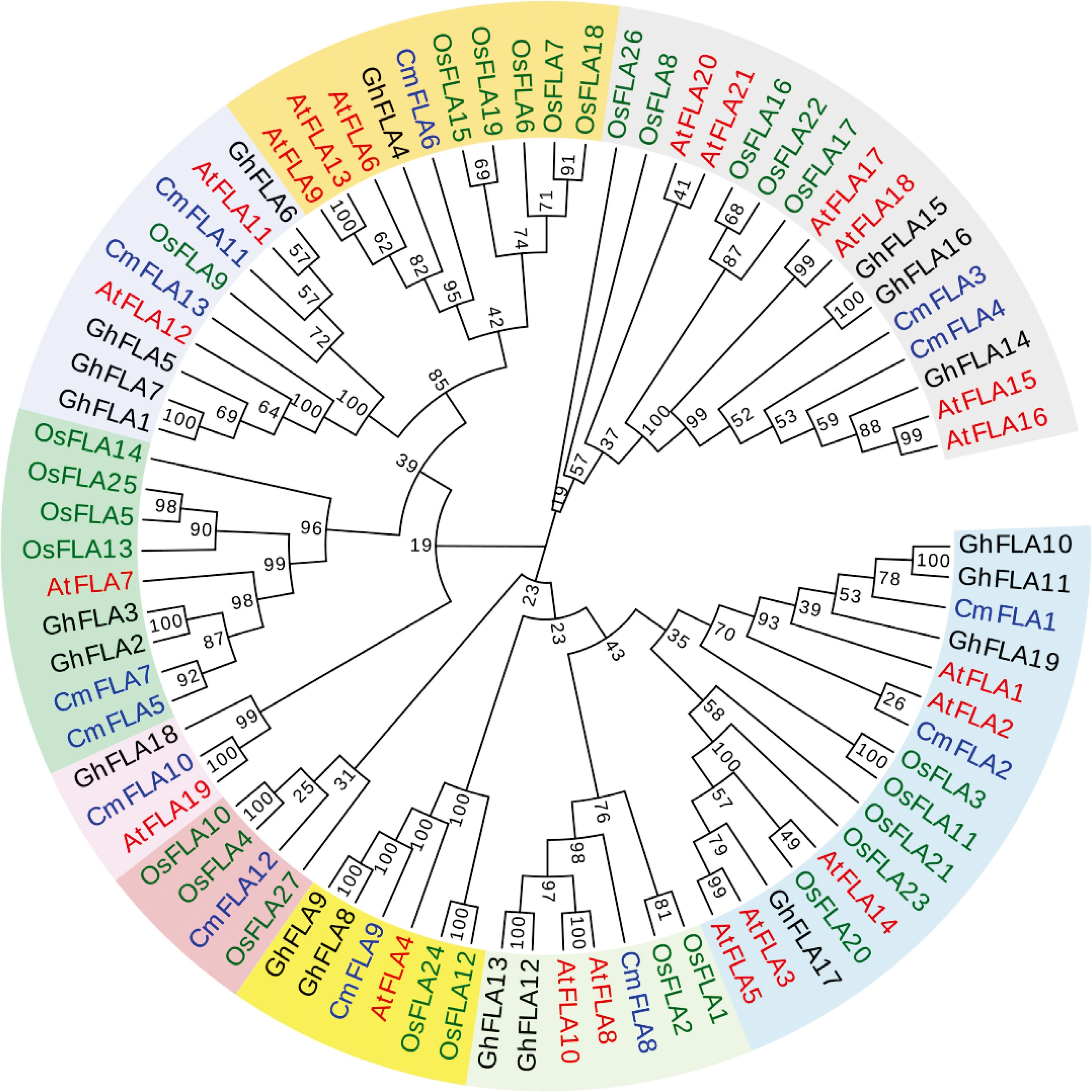
Figure 3.
Phylogenetic relationship of FLAs between melon and other species. Red color represents Arabidopsis thaliana (At); blue color represents Cucumis melon (Cm); green color represents Oryza sativa (Os); and black color represents Gossypium hirsutum (Gh). The phylogenetic tree was constructed using MEGA 7 software with the Neighbor-Joining method, employing a bootstrap value of 1,000 replicates. The GenBank accession numbers of the sequences were used for the analyses, as detailed in Supplementary Table S4.
Expression analysis of CmFLAs genes in various organs and fruit at different developmental stages
-
The expression patterns of CmFLAs genes in different organs of melon were revealed using the public RNA-seq resource in the cucurbit genomic database. It was observed that all CmFLAs showed low expression level in mature fruit (Fig. 4a). CmFLA10 was specifically expressed in male flower, while CmFLA12 showed abundant expression in the root. Interestingly, three genes (CmFLA6, CmFLA11, and CmFLA13) displayed a similar expression pattern across different tissues, with high expression levels in both the root and leaf.
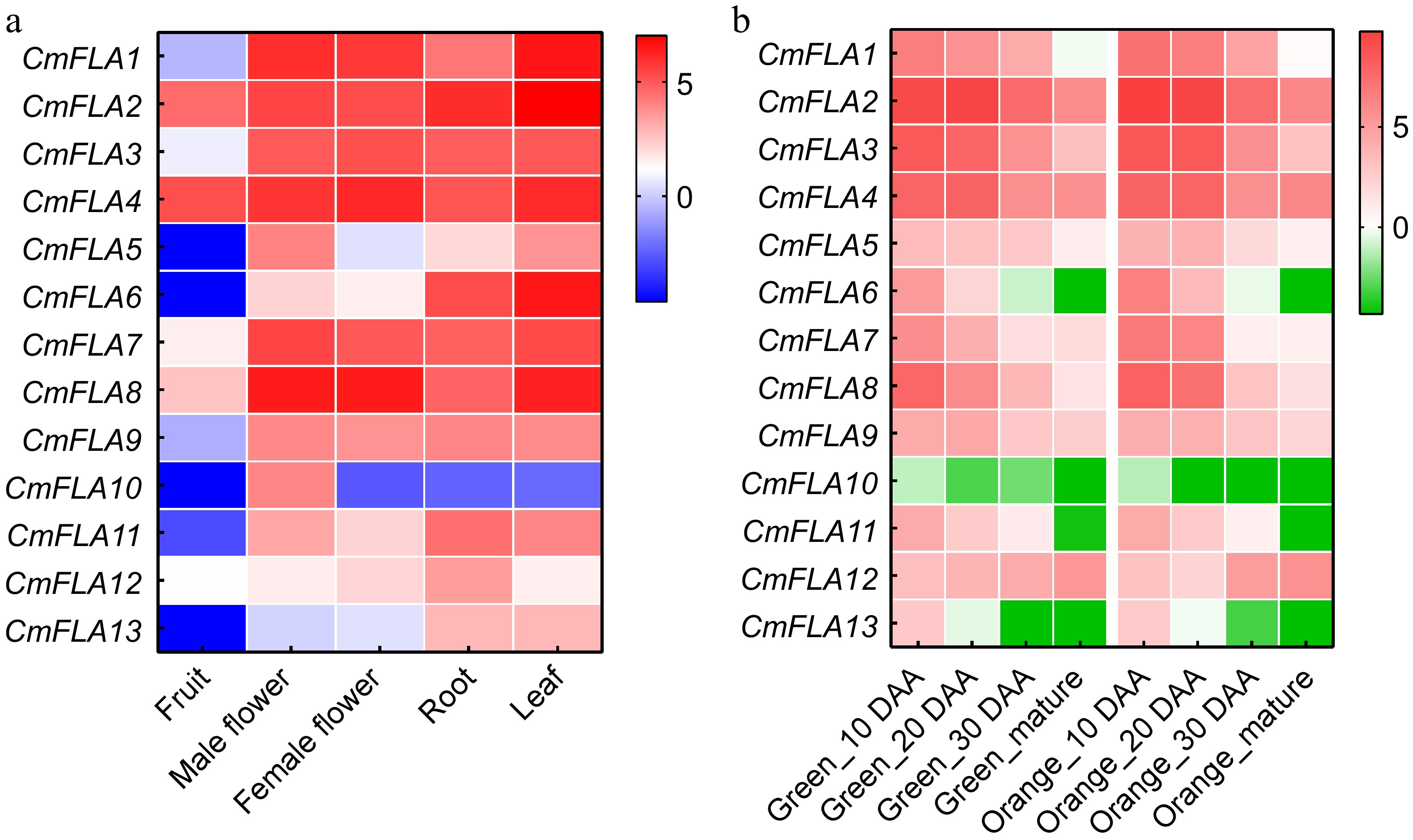
Figure 4.
Heatmaps show the expression analysis of CmFLA gene. (a) Expression profiles of CmFLA genes in different organs. Project: PRJNA383830. (b) Expression profiles of CmFLAs genes in fruit at different developmental stages. Expression was quantified on the indicated day after anthesis (DAA). Project: PRJNA286120. Scaled log2 expression values based on RNA-seq data are shown from blue to red, indicating low to high expression levels. The data were obtained from the Cucurbitgenomics database (
http://cucurbitgenomics.org/rnaseq/home ).To further investigate the gene expression during different development periods of the fruit, we utilized additional RNA-seq data collected from two cultivars of melon. The expression patterns of CmFLA10 were observed to be low in both two cultivars. CmFLA12 demonstrated a positive correlation with fruit maturation, whereas the majority of CmFLAs exhibited a negative relationship (Fig. 4b).
GO analysis of CmFLAs
-
GO annotation is an effective tool for describing gene localizations and functions. Consequently, the GO annotations of CmFLAs were employed to describe biological processes, cellular components, and molecular functions were determined. The results of GO analysis indicated that CmFLAs were enriched in cellular components, integral components of membranes, membrane, and anchored components of plasma membrane of the cellular component categories (Supplementary Fig. S1, Supplementary Table S5). Within the molecular function categories, molecular function, gamma-glutamyltransferase activity, actin binding, and polysaccharide binding were found (Supplementary Fig. S1, Supplementary Table S5). Moreover, biological process, cell adhesion, and response to salt stress exhibited high enrichment (Supplementary Fig. S1, Supplementary Table S5). Additionally, the GO annotation analysis highlighted the involvement of CmFLA2, CmFLA9, and CmFLA12 in response to salt stress (Supplementary Table S5).
Cis-element analysis of CmFLAs promoter
-
Putative cis-elements in the promoter of CmFLAs were predicted using PlantCARE. It was observed that all CmFLA promoters contained light-responsive elements, with some also possessing circadian control elements. Additionally, the promoter regions exhibited an abundance of hormonal or environment-responsive elements, including ABA responsiveness, auxin-responsive element, gibberellin-responsiveness, salicylic acid responsiveness, defense and stress responsiveness, low-temperature responsiveness, and drought induction (Supplementary Fig. S2).
Furthermore, the promoter of CmFLA6 and CmFLA12 contained MYBHv1 binding site. Additionally, CmFLA8 promoter contained a protein binding site, while a cis-acting regulatory element was found in the promoter of CmFLA10 (Supplementary Fig. S2).
Expression analysis of CmFLAs in different melon cultivars
-
To investigate the relationship between the expression of CmFLAs and total soluble solids content (TSS), we examined the expression of CmFLAs in melon fruits from various melon cultivars (Fig. 5a). The qRT-PCR results indicated that the expression levels of CmFLA1, 3, 6, 7, 8, 11, and 13 in BLC, HPC, and BCG (low TSS cultivars) were higher compared to XM, YJ, LT4, GS, and M43 (high TSS cultivars). Correlation analysis between TSS and gene expression indicated most of the CmFLAs were negatively related to TSS (Fig. 5b).
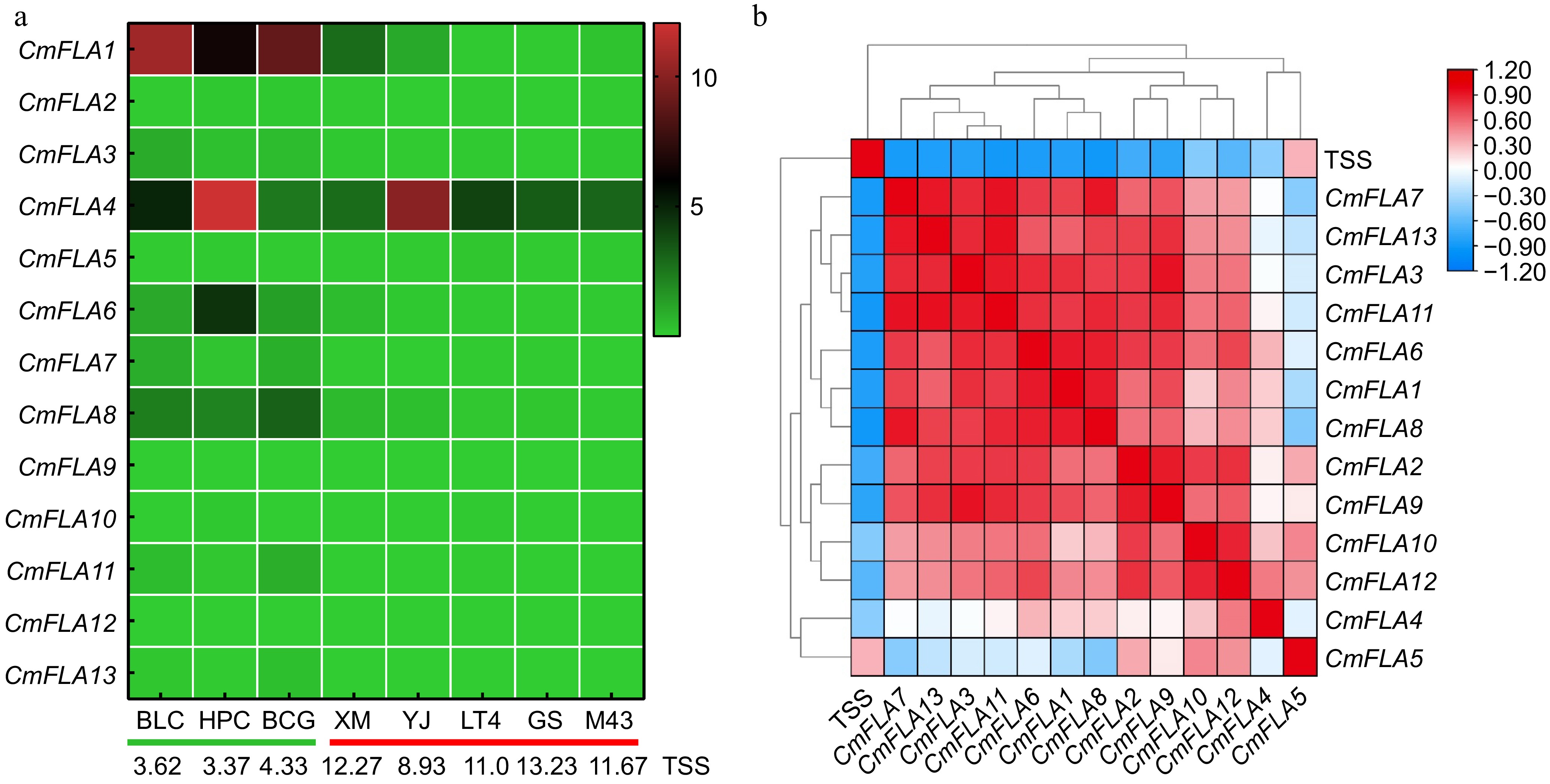
Figure 5.
Correlation analysis between total soluble solid contents (TSS) and gene expression of CmFLAs. (a) Heatmap of the CmFLA genes in melon fruit at 30 DAA from various cultivars obtained using qRT-PCR analysis. TUA (α-tubulin) was used as reference gene. Expression values based on qRT-PCR data are shown from green to red, indicating low to high expression. The values at the bottom of the figure indicate the TSS for each melon cultivars. The green line represents low TSS cultivars, and the red line represents high TSS cultivars. (b) TSS and gene expression correlation analysis in melon fruit at 30 DAA from various cultivars. Data are shown from blue to red, indicating low to high correlation.
ABA inhibited the expression of CmFLA4, 6, and 11 in melon fruit
-
The promoter elements of CmFLA1, 3, 4, 5, and 8 suggest the potential involvement of these genes in ABA responsiveness (Supplementary Fig. S2). Subsequently, we examined the gene expression of CmFLAs after ABA treatment in melon fruit of high TSS cultivar. Our findings indicated that the expression of CmFLA4, 6, and 11 was repressed by ABA, while the expression of other genes remained unaffected (Supplementary Fig. S3).
Sugar content increased after β-Glc Yariv treatment in melon fruit
-
β-Glc Yariv was sprayed on the melon fruit of the cv. 'BCG' (low TSS cultivars) at 20 DAA to investigate its effects on CmFLAs gene expression and sugar content. The results showed that the expression of CmFLA1, 8, 11, and 13 was suppressed after β-Glc Yariv treatment (Fig. 6a). The samples were further analyzed for sugar content by GC. The results revealed a significant increase in sucrose content in the β-Glc Yariv treated samples compared to the control, accompanied by a decrease in fructose content (Fig. 6b).
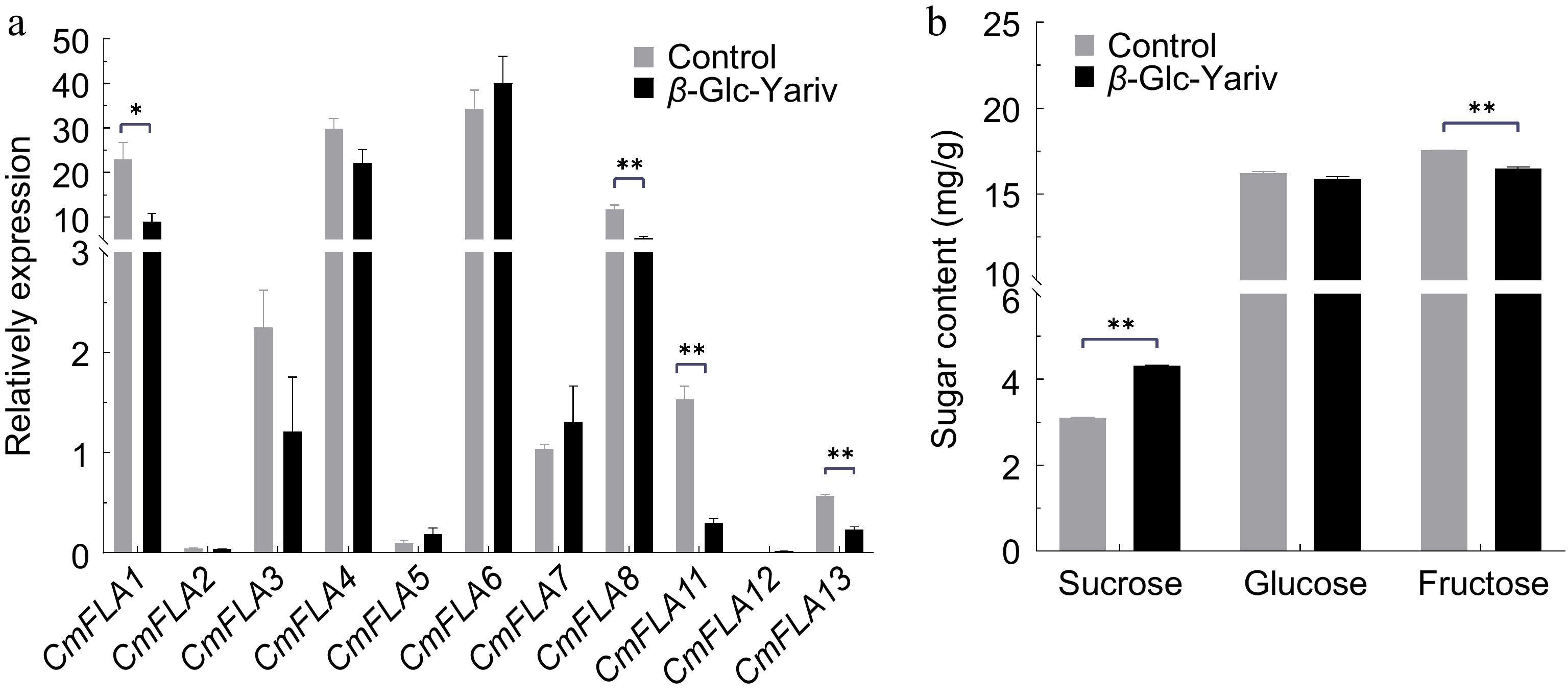
Figure 6.
Sugar content and gene expression of CmFLAs after β-Glc Yariv treatment in melon fruit. Melon fruits of the cv. 'BCG' at 20 DAA were sprayed with β-Glc Yariv solution. (a) Gene expression of CmFLAs after β-Glc Yariv treatment. TUA (α-tubulin) was used as reference gene. (b) Sugar content after β-Glc Yariv treatment. Water was used as the control. Data represent means of three biological replicates ± SE. Statistical significance was denoted as * p < 0.05, ** p < 0.01 by t-test.
To identify genes associated with changes in sugar content, we performed an expression analysis of sugar metabolism-related genes. This analysis revealed a significant increase in the expression of alkaline a-galactosidase/neutral a-galactosidase genes (CmAAG1, CmAAG2; CmNAG1, CmNAG2, CmNAG3), which are associated with the breakdown of RFOs into sucrose[50]. Similarly, the expression of neutral invertase genes (CmNIN1/3/4) was upregulated, while acid invertase gene CmAIN2 was down-regulated, which is responsible for hydrolyzing sucrose into glucose and fructose[51]. Additionally, the expression of CmSPS1 and CmSPS2, responsible for synthesizing UDP-glucose with fructose-6P into sucrose-P[52], was also increased. No significant difference was observed in the expression of CmSUSs genes compared to the control. Furthermore, the tonoplast sugar transporter CmTST2, known for promoting sugar accumulation in the vacuole[45], exhibited elevated expression after β-Glc Yariv treatment (Supplementary Fig. S4).
CmFLA8 localizes to the cytoplasm to inhibit glucose accumulation in melon fruit
-
We then investigated the function of the CmFLA8 gene, which is highly expressed in melon fruit and significantly reduced following inhibitor treatment, to determine its role in regulating fruit sugar accumulation. Localization studies in tobacco protoplasts confirmed cytoplasmic expression of CmFLA8 (Fig. 7a). We then employed a melon transient expression system to validate the function of CmFLA8 in fruit (Fig. 7b). The expression of CmFLA8 in overexpressing fruits (CmFLA8-OE) was significantly higher than that in the control fruits (Fig. 7c). Subsequent assessment of fruit sugar levels revealed a notable decrease in glucose content in CmFLA8-OE compared to control fruit, as shown in Fig. 7d. In Arabidopsis, glucose represses chlorophyllab-binding protein1 (CAB1) expression while inducing the nitrate reductase1 gene (NR1)[53]. Consequently, these genes serve as markers to signify alterations in cytoplasmic glucose levels. We identified melon homologs, CmCAB1 and CmNR1, showing high sequence similarity to these Arabidopsis proteins. Expression analysis revealed an increase in CmCAB1 expression and a decrease in CmNR1 expression in CmFLA8-OE fruits (Fig. 7e), indicating a decrease in glucose in the cytoplasm.
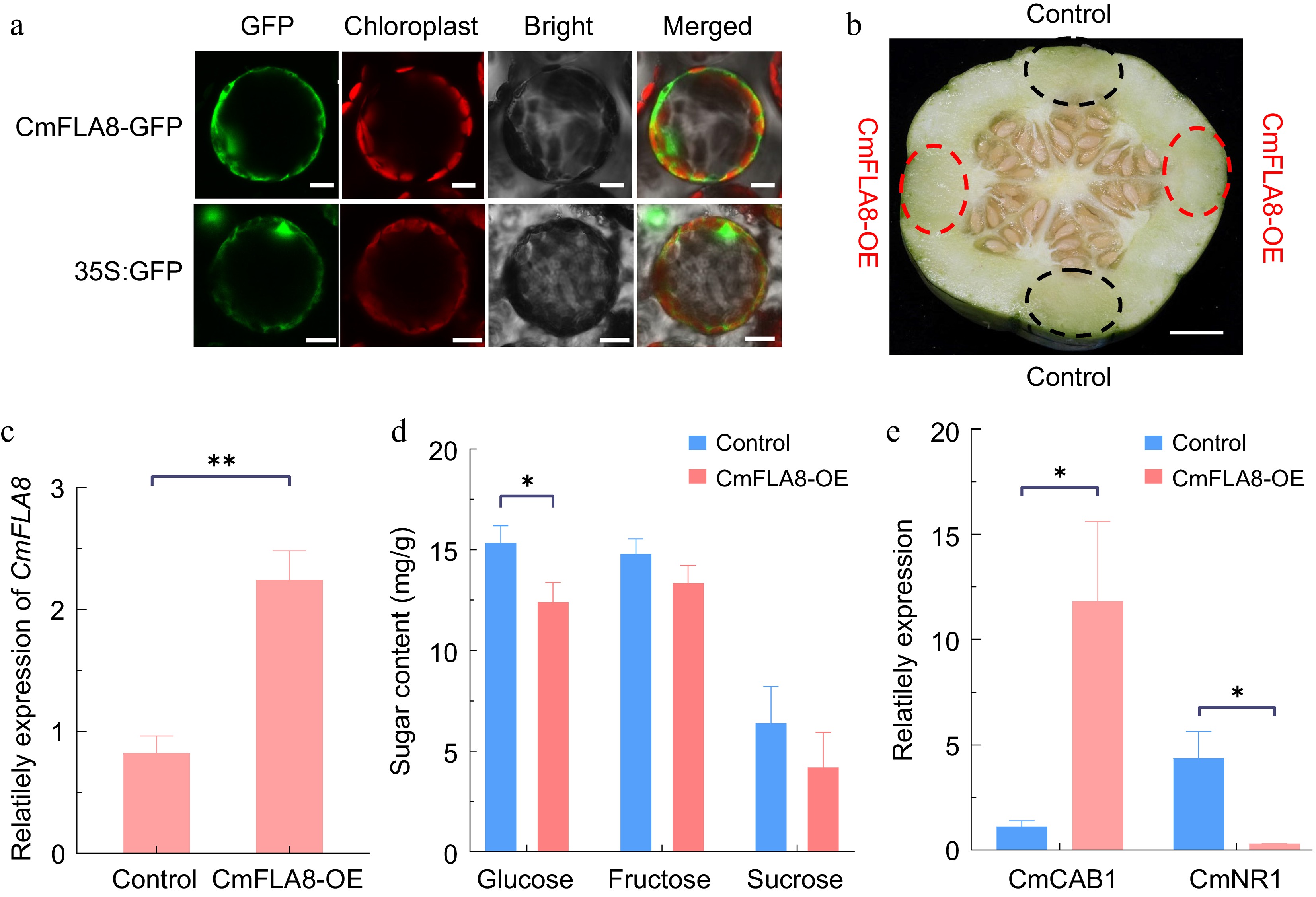
Figure 7.
Gene function verification of CmFLA8. (a) Subcellular localization of CmFLA8 in N. benthamiana protoplasts. Scale bars =10 μm. (b) Transient expression of CmFLA8-OE in melon fruit. CmFLA8-OE agrobacterium cells were injected into the melon tissue, with the pHELLSGATE8-HA vector used as a control. The dotted circles indicate the injection sites, scale bar = 1 cm. (c) Relative expression of CmFLA8 in CmFLA8-OE melon fruit. Reference gene: Tubulin. (d) Sugar concentrations in CmFLA8-OE melon fruit. (e) qRT-PCR analysis of CAB1 and NR1 expression in melon fruit. Reference gene: Tubulin. Empty vector, control fruit expressed the empty pHELLSGATE8-HA vector. Data represent means of four replicates ± SE. * p < 0.05 and ** p < 0.01 by t-test.
-
In this study, we identified 13 CmFLAs in melon and divided them into four groups (Fig. 2b). All CmFLAs recognized as secretory proteins with N-terminal signal peptides and FAS domains containing three highly conserved motifs (H1, H2, and YF), similar to other plant FLAs[10,11]. The GPI-anchor can be cleaved from the protein by endogenous GPI-specific phospholipases or other enzymes, releasing the protein into the extracellular space[54]. The GPI-modification signal is crucial for membrane anchoring, efficient ER exit, secretion, and plasma membrane localization of AtFLA4[12]. Thus, the GPI-anchor signal is vital for maintaining FLA function. Among the 13 CmFLAs identified, five possess the GPI-anchor signal; however, further experiments are needed to ascertain the precise role of the GPI-anchor signal.
Glycosylation involves the post-translational modification of Pro residues to Hyp, and the type of glycosylation relies on the presence of contiguous or noncontiguous Hyp residues[10]. Two types of glycosylation, N-glycosylation, and O-glycosylation, are observed in FLAs. N-glycosylation, occurring on Asn residues, is a characteristic feature, while O-glycosylation, involving the glycosylation of hydroxyl groups on the side chains of amino acids, is more complex[55]. The position of N-glycosylation site is predicted within the FAS domains of FLA[10,47]. N-glycosylation in AtFLA4 plays a vital role in ER-exit, whereas O-glycosylation influences post-secretory fate[12]. Additionally, the O-glycosylation of AtAGP21 is crucial for secretion and cellular targeting[56]. CmFLAs were predicted to contain 1–6 N-glycosylation sites. Furthermore, CmFLA6 and CmFLA13 were found to have fewer than 10 O-glycosylation sites, while the remaining CmFLAs contain 10–35 sites (Table 2), suggesting potential variations in their glycosylation patterns.
Melon FLAs were negatively correlated with fruit ripening and sugar accumulation
-
The expression of most CmFLAs is inversely associated with fruit ripening and TSS content (Figs 4b & 5), indicating a potential role for CmFLAs as negative regulators during melon fruit ripening. In tomato, several SlFLA genes were notably downregulated during fruit ripening stages, and silencing of SlFLA13 accelerates tomato fruit ripening and enhances carotenoid accumulation[57].
In this study, some CmFLAs, such as CmFLA1, 8, 11, and 13, exhibited a negative correlation between their expression and fruit ripening as well as sugar content (Figs 4b & 5). Treatment with β-Glc Yariv led to a reduction in their expression and an increase in sugar content (Fig. 6a). Expression analysis of sugar metabolism-related genes revealed an increase in CmAAG/NAGs after β-Glc Yariv treatment (Supplementary Fig. S4). In watermelon, ClAGA2 is a pivotal factor in controlling stachyose and raffinose hydrolysis, thereby influencing fruit sugar accumulation[50]. According to the model of sugar unloading and post-phloem transport in melon fruit, α-galactosidase represents an essential step in sucrose accumulation[50]. Elevated CmAAG/NAG expression promotes greater sucrose production. Notably, the expression levels of CmNAGs surpassed those of CmAAGs in Supplementary Fig. S4, suggesting the potential heightened importance of CmNAGs compared to CmAAGs in melon fruit.
Invertase enzymes are classified as CWINs, VINs, and CINs based on their cellular localization in the cell wall, vacuole, and cytosol. CINs are categorized as NINs, while CWINs and VINs are classified as AINs[51]. The vacuole can store high concentrations of sugars[58]. Upon sucrose entry into the fruit pulp cells or fruit parenchyma cells, CmAIN2 breaks it down into glucose and fructose. Consequently, reduced CmAIN2 expression leads to a decrease in sucrose hydrolysis (Supplementary Fig. S4), resulting in a reduction in fructose content (Fig. 6b). Moreover, increased expression of CmSPS1/2 gene in β-Glc Yariv treatment facilitated sucrose synthesis in the cytoplasm, with the resulting sugars promptly transported into the vacuole by CmTST2[45]. Consequently, sugar content increased after β-Glc Yariv treatment in melon fruit (Fig. 6b).
Our investigation confirmed that CmFLA8 is localized in the cytoplasm and inhibits glucose accumulation in melon fruit. Additionally, it influences fructose and sucrose levels; however, the observed effect was not statistically significant (Fig. 7). Thus, it is conceivable that CmFLA1, 11, and 13 could also impact sugar levels. Due to the lack of prior research on the impact of FLA proteins on sugar accumulation, we propose a potential role for FLA proteins in modulating sugar-metabolic enzymes or their upstream-regulated transcription factors through post-translational modifications. Further experiments are necessary to elucidate how these CmFLA protein specifically affect the sugar content in melon.
ABA-responsive element was identified in the promoter regions of CmFLA4 (Supplementary Fig. S2). For CmFLA4, 6, and 11, their expression displayed a negative correlation with fruit ripening and was inhibited by ABA (Fig. 4b; Supplementary Fig. S3). ABA plays an important role in promoting the ripening of numerous fleshy fruits[59]. Additionally, the expression of certain FLA genes has been observed to be downregulated by ABA in other plant species[10,25,60]. Consequently, CmFLA4, 6, and 11 may influence the expression of sugar synthesis or sugar transporter-related genes during the early stages of fruit development, resulting in reduced sugar content in the fruit. As melon fruit matures, ABA signaling may suppress the expression of these genes, thereby facilitating the activation of sugar accumulation-related genes and promoting sugar accumulation. The expression of these CmFLAs may be under the regulation of ABA-signal transduction factors, further experiments are needed to verify this hypothesis.
-
In conclusion, 13 CmFLAs were identified in the melon genome and were clustered into four groups based on FAS motifs and AGP-regions. The comprehensive structural, phylogenetic, chromosomal localization, expression analyses, and promoter prediction in this study shed light on the functions of FLAs in melon. CmFLA10 and CmFLA12 exhibited tissue-specific expression patterns. Furthermore, expression analysis revealed a negative correlation between most CmFLAs and fruit ripening as well as TSS accumulation. When melon fruit was treated with β-Glc Yariv, the expression of CmFLA1, 8, 11, and 13 was downregulated. Concurrently, the transcript levels of sugar metabolism-related genes and sugar transporters were upregulated. Consequently, sucrose accumulation was enhanced while the fructose content in the vacuoles was diminished. CmFLA8 is localized in the cytoplasm. Moreover, the transient overexpression of CmFLA8 in melon fruits resulted in a reduction in glucose content compared to the control. Further research, such as the use of CRISPR technology for gene knockout or transgenic technology for overexpression, is necessary to elucidate the function of specific functions of individual FLA genes. Overall, these findings may provide valuable insights for future investigations into the biological function of FLAs in melon.
This work was supported by the Hubei Provincial Funding Programme to Support High-Quality Development of the Seed Industry (HBZY2023B004-5) to Jintao Cheng, and the China Postdoctoral Science Foundation (2022M711276) to Suying Wen.
-
The authors confirm contribution to the paper as follows: study conception and design, experiment conducting, data analysis, manuscript writing: Wen S; data collection: Sun R, Gao M; analysis and interpretation of results: Huang R, Li J, Zhou Y; research supporting, manuscript revision: Cheng J; critical feedback providing and research revision: Bie Z. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Specific primers used for this article.
- Supplementary Table S2 Analysis and distribution of conserved motifs in CmFLA proteins.
- Supplementary Table S3 Analysis of identity among AtFLA and CmFLAs.
- Supplementary Table S4 GenBank accession numbers of the sequences used for the Fig. 5.
- Supplementary Table S5 Gene Ontology annotation details for CmFLA proteins in melon.
- Supplementary Fig. S1 Gene ontology distributions for CmFLAs in melon. The detail information is provided in Table S5.
- Supplementary Fig. S2 Prediction of cis-acting elements in the promoters of CmFLAs.
- Supplementary Fig. S3 Gene expression of CmFLAs after ABA treatment in melon fruit. Melon fruit of the cv. 'Boyang' at 20 DAA were chosen for ABA treatment. TUA (α-tubulin) was used as reference gene. Water was used as the control. Data represent means of three biological replicates ± SE. Statistical significance was denoted as *p < 0.05 by t-test.
- Supplementary Fig. S4 Expression analysis of sugar metabolism-related genes after β-Glc Yariv treatment in melon fruit. Melon fruits of the cv. 'BCG' at 20 DAA were sprayed with β-Glc Yariv solution. TUA (α-tubulin) was used as reference gene. Water was used as the control. Data represent means of three biological replicates ± SE. Statistical significance was denoted as *p < 0.05 and ** p <0.01 by t-test.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wen S, Huang R, Gao M, Li J, Zhou Y, et al. 2025. Genome-wide identification of fasciclin-like arabinogalactan proteins (FLAs) in melon and the role of CmFLA8 in glucose accumulation in fruit. Vegetable Research 5: e009 doi: 10.48130/vegres-0025-0004 

Genome-wide identification of fasciclin-like arabinogalactan proteins (FLAs) in melon and the role of CmFLA8 in glucose accumulation in fruit
- Received: 11 December 2024
- Revised: 13 January 2025
- Accepted: 03 February 2025
- Published online: 27 March 2025
Abstract: Fasciclin-like arabinogalactan proteins (FLAs) play an important role in plant growth and development. Nevertheless, little is known about FLA genes in melon. Here, we conducted a systematic analysis of the FLAs subfamily within the annotated genome of melon (Cucumis melo L.). Thirteen FLAs were identified, and with sequencing analysis categorized into four groups based on the presence of FAS domains and AGP-regions. Notably, five of the CmFLAs proteins contain a glycosylphosphatidylinositol anchor signal. Promoter cis-element prediction analysis indicated that the majority of the CmFLAs family members respond to abiotic stresses and hormones. The RNA-seq and qRT-PCR assays showed that CmFLA10 exhibited specific expression patterns in male flowers, while CmFLA12 was highly expressed in the root tissue. Notably, most of the CmFLAs genes were expressed at high levels during the early stages of fruit development, subsequently declining as the fruit ripened. Their expression levels were negatively correlated with the total soluble solids content. Treatment with β-Glc Yariv inhibited the expression of CmFLA1, 8, 11, and 13, resulting in a significant increase in the sugar content of the fruits. Expression analysis suggested an induced up-regulation of genes associated with sugar metabolism. Subcellular localization studies demonstrated that CmFLA8 was localized in the cytoplasm. Overexpression of CmFLA8 in melon fruit resulted in a decrease in glucose content. In this study, CmFLAs play a negative role in sugar accumulation and fruit ripening in melon, providing new insights into FLA gene functions.
-
Key words:
- Fasciclin-like arabinogalactan proteins /
- Melon fruit /
- Genome-wide /
- Expression analysis