









-
Saline-alkali stress poses a severe threat to crop growth and development, representing a major abiotic stress factor that constrains sustainable agricultural development worldwide[1−3]. Soil salinization is a severe ecological problem confronting arable lands worldwide[4]. Saline-alkali soils, characterized by high levels of sodium bicarbonate and sodium carbonate, are marked by high salinity and elevated pH (above 8.0)[5]. Neutral salts damage plants through ionic toxicity, osmotic stress, and secondary stress. More critically, saline-alkali stress causes more severe damage through the combination of high pH, ionic toxicity, osmotic stress, and secondary stress[6−8]. Compared with the knowledge about plant responses to neutral salts, the mechanism underlying plant resistance to saline-alkali stress is still far behind.
The evolutionarily conserved SOS (salt overly sensitive) pathway serves as a hub in maintaining ion homeostasis[9]. Under salt stress, intracellular Ca2+ signals are elevated. SOS3 and SCaBP8, the Ca2+-binding proteins, perceive Ca2+ signals and activate the kinase activity of SOS2 through physical interaction. SOS2 activates the Na+/H+ efflux carrier activity of SOS1 by phosphorylation, thus promoting the efflux of sodium ions. In addition, SOS2 can also activate SOS1[10,11], NHX (a possible vacuolar K+/H+ transporter), CAX1 (vacuolar Ca2+/H+ antiporter), and VHA (vacuolar H+-ATPase), thereby sequestering Na+ in the vacuole[9]. Recent studies have found that the function of CIPK8, a protein in the same family as SOS2, is similar to that of SOS2 and can mediate salt homeostasis in Arabidopsis thaliana[12]. Besides SOS3/SCaBP8, GRIK1 (geminivirus Rep-interaction kinase 1) and GRIK2 proteins are also capable of phosphorylating and activating SOS2, thereby enhancing plant salt resistance[9]. Natural variations in components of the SOS pathway, such as SlSOS1, SlSOS2, SlSCaBP8, ZmSOS1, and ZmCBL8, lead to the compromised salt tolerance in cultivars of tomato and maize[13−16]. Transcription of SOS1 is regulated by the CycC1;1-WRKY75 module during salt stress[17].
The activation of the SOS signaling pathway depends on the perception of salt signals. Glycosyl inositol phosphorylceramide (GIPC) may be involved in this perception process. It binds to extracellular salt ions, triggering changes in cell-surface potential and the opening of calcium ion channels, which in turn leads to an increase in the intracellular calcium ion concentration, thereby activating the SOS signaling pathway[18]. The receptor-like kinase FERONIA can sense the integrity of the cell wall and participate in root growth recovery under salt stress conditions[19]. The cell wall-localized signaling pathway, consisting of leucine-rich repeat extensin proteins (LRX) 3/4/5, rapid alkalinization factors RALF22/23, and FERONIA, regulates cell wall growth and thereby mediates plant growth responses to salt stress[20].
To deal with osmotic stress, plants adjust their metabolism to synthesize osmotic regulators such as proline and sorbitol. OSCA1, MSL8, and KEA1/2/3 are three potential osmotic stress sensors, but their functions remain to be further verified[21−23]. The cascade signaling pathways mediated by MKK4-MPK3 and MKKK20-MPK6 are also implicated in osmotic regulation[24,25]. Leaf starch is degraded by salt-mediated ABA signaling, releasing sucrose to relieve osmotic stress[25].
Under salt stress, intracellular reactive oxygen species (ROS) need to be scavenged. Lectin receptor kinase SIT1 exerts its effect by phosphorylating and activating MPK3/6, which sets off ethylene production as well as salt-induced ethylene signal transduction, yet it acts to negatively regulate salt tolerance in plants[26]. The B'κ subunit of protein phosphatase 2A negatively regulates SIT1 through dephosphorylation[27]. The receptor-like cytoplasmic kinase STRK1 boosts the activity of catalase C, which helps balance H2O2 levels and, in turn, enhances plant tolerance to salt stress[28].
Compared with neutral salts, saline-alkali stress imposes much more complex and severe damage to plants, and the underlying mechanisms remain poorly understood. Protein kinase PKS5 exerts an inhibitory effect on PM-H+-ATPase activity via phosphorylation, thus functioning as a negative regulator of plant resistance to high-pH stress[29]. The calcium-binding protein SCaPB3 interacts with PKS5 and PM-H+-ATPase AHA2, respectively, phosphorylating AHA2. High-pH stress activates the calcium ion signal and inhibits the phosphorylation of AHA2, promoting AHA2-mediated proton efflux and resistance to high-pH stress[30]. In maize, saline-alkali stress strengthens the interaction between calcium ions and their binding protein ZmNSA1, promoting the protein degradation of ZmNSA1, increasing the activities of PM-H+-ATPase and SOS1, and thus promoting the efflux of sodium ions. The natural variation in ZmNSA1 leads to a decrease in the translation efficiency of its mRNA, thereby enhancing resistance to saline-alkali stress[31]. Natural variation in SlSCaBP8 accounts for the impaired saline-alkaline tolerance during tomato domestication[16]. In some monocot crops, such as wheat, sorghum, maize, rice, and millet, the Gγ subunit AT1 (alkaline tolerance 1), acts to negatively regulate the phosphorylation of PIP2 aquaporins. This, in turn, diminishes their H2O2 export activity, resulting in H2O2 overaccumulation and subsequent sensitivity to alkaline stress[32,33]. ATT2, which encodes GA20-oxidases, emerges as a potential candidate for precisely modulating GA concentrations to optimal thresholds. This modulation, in turn, orchestrates a balance between reactive oxygen species (ROS) and H3K27me3 methylation, thereby fortifying plant tolerance to combined alkali and thermal stress[34]. The SlWRKY42-SlMYC2 module works synergistically to boost saline-alkali tolerance in tomato, by activating both the jasmonic acid (JA) signaling pathway and spermidine biosynthesis pathway[35]. Melatonin improves tomato plant tolerance to saline-alkali stress through two key mechanisms: alleviating nitrosative damage and modulating S-nitrosylation of plasma membrane H+-ATPase 2[36].
Heterotrimeric G proteins (G proteins) are important components modulating plant growth and stress response[37,38]. The tomato genome encodes one canonical Gα, four noncanonical extra-large Gα (XLGs: Solyc08g005310, Solyc08g076160, Solyc03g097980, and Solyc02g09016), one Gβ (Solyc01g109560), and four Gγ subunits (Solyc09g082940, Solyc12g096270, Solyc08g005950, and Solyc07g041980)[39−41]. The Gγ subunit AT1 (alkaline tolerance 1) is the homologue of SlGGC1 (Solyc07g041980) in tomato. Due to the important role of AT1 against alkaline stress in wheat, maize, sorghum, millet, and rice[32,33], whether and how SlGGC1 regulates saline-alkali tolerance is not known. In this study, SlGGC1 was cloned, and a slggc1-cr mutant was constructed by the CRISPR/Cas9 gene editing technology. The results demonstrate that SlGGC1 is an important positive regulator in saline-alkali stress response.
-
The cultivar 'AC' was obtained from TGRC (Tomato Genetic Resources Center, University of California at Davis, US), and the slggc1 mutant in 'AC' background was produced using CRISPR/Cas9 gene-editing technology[42]. The planting substrate was formulated by mixing nutrient soil with 2–4 mm vermiculite at a 1:1 volume ratio (v/v), followed by uniform distribution into 7 cm × 7 cm square seedling pots. Hoagland's nutrient solution, supplemented with or without 75 mM NaHCO3, was applied to each pot until soil water saturation was achieved, thereby simulating saline-alkaline and normal soil conditions, respectively. For sowing, uniformly sized 'AC' and slggc1 tomato seeds with consistent radicle emergence were selected and planted in the corresponding soil without or with NaHCO3 as required. Seedlings were cultivated in a climate chamber with 60% relative humidity under a 16 h light period at 26 °C and an 8 h dark period at 20 °C.
Root length measurement
-
The seeds were sterilized with 50% NaClO for 10 min and washed with sterile water to eliminate residual NaClO. Subsequently, the sterilized seeds were germinated on 1/2 MS medium with or without 10 mM NaHCO3. The root length was quantified after a 5 d incubation period using ImageJ software.
SlGGC1 sequence assessment
-
GGC1 protein sequences of tomato, potato, and pepper were obtained from NCBI. The phylogenetic analyses using the neighbor-joining approach were conducted using MEGA 11 (
www.megasoftware.net ). Bootstrap sets the initial value 1,000 times. DNAMAN10 software was utilized to align the GGC1 sequences. The GGC1 sequences are listed in Supplementary Table S1.RNA extraction and qRT-PCR
-
The sterilized seeds were grown on 1/2 MS medium for 5 d, and subsequently, the seedlings were transferred to 1/2 MS medium without or with 10 mM NaHCO3. Root samples were collected at 0, 6, 12, 24, and 48 h, and total RNA was extracted using TRIzol® reagent (TransGen Biotech). Total RNA (1 μg) was reverse-transcribed into cDNA using an all-in-one kit (TransGen Biotech, Beijing). Quantitative PCR was subsequently performed using the SYBR Green kit (TransGen Biotech) on a Bio-Rad CFX96 instrument. Primers are listed in Supplementary Table S2.
Subcellular localization assays
-
The 35S:SlGGC1-GFP construct was generated by cloning the coding sequence of SlGGC1 into the pK7FWG2 vector. The confirmed positive clone was subsequently transferred into the Agrobacterium tumefaciens strain GV3101. 35S:SlGGC1-GFP was co-expressed with the plasma membrane marker PM-cherry in N. benthamiana leaves and visualized by confocal microscopy. 35S:GFP was used as a positive control. Primers used for these constructs are listed in Supplementary Table S2.
Measurement of Na+ and K+ contents
-
The method for determining Na+ and K+ contents was described previously[16]. Briefly, 21-day-old seedlings grown under either water (control) or 75 mM NaHCO3 conditions were harvested. The collected plants were rinsed three times with double-deionized water to remove surface contaminants, then dried at 80 °C for 24 h and ground into a fine powder. A 10 mg aliquot of the tissue powder was digested with 1 mL of nitric acid for 2 h using a Microwave 3000 digestion system (Anton Paar, Austria). Germanium was employed as the internal standard, and Na+ and K+ concentrations were quantified by inductively coupled plasma mass spectrometry (ICP-MS; ICAPQ, Thermo Fisher Scientific).
Quantification of reactive oxygen species
-
The determination of H2O2 content, O2− content, MDA content, GSH activity, SOD activity, POD activity, and CAT activity followed the manufacturer's protocols (Jiangsu Aidisheng Biological Technology Co., Ltd). Detailed procedures for key measurements are as follows:
For H2O2 content determination (potassium iodide method): 1 g of fresh tomato leaf tissue was weighed and transferred to a pre-chilled mortar. Under ice-bath conditions, the tissue was homogenized with 10 mL of ice-cold 0.1% (w/v) trichloroacetic acid (TCA). The homogenate was centrifuged at 12,000 rpm for 15 min at 4 °C. A 1 mL aliquot of the supernatant was transferred to a 10 mL centrifuge tube, followed by sequential addition of 1 mL PBS (10 mM, pH 7.0) and 2 mL 1 M KI solution. The mixture was vortexed vigorously and incubated at 28 °C for 30 min. After adding 0.5 mL of 0.1% starch solution, absorbance at 620 nm was measured immediately using a spectrophotometer.
For superoxide anion (O2−) content determination (hydroxylamine hydrochloride oxidation method), 1 g of fresh tomato leaf tissue was weighed and placed into a pre-chilled mortar, then homogenized thoroughly with 10 mL of ice-cold 0.05 M PBS (pH 7.8) under ice-bath conditions. The homogenate was centrifuged at 10,000 rpm for 20 min at 4 °C. A 2 mL aliquot of the supernatant was transferred to a tube, and 1 mL of 10 mM NH2OH·HCl solution was added. The mixture was gently vortexed and incubated at room temperature in the dark for 1 h to allow O2− to convert to NO2−. Subsequently, 2 mL of chromogenic reagent was added, and the reaction mixture was shaken vigorously and left to stand at room temperature for 15 min. Absorbance at 530 nm (A530) was then measured using a spectrophotometer.
Statistical analysis
-
All experimental data are presented as the mean ± standard error (SE) from at least three independent biological replicates. Statistical analyses were conducted using IBM SPSS Statistics software. Significant differences between treatment groups were assessed via two-way analysis of variance (ANOVA), followed by Dunnett's multiple comparison test, with the control group serving as the reference. A p value < 0.05 was regarded as statistically significant.
-
SlGGC1 encodes 210 amino acids. The GGC1 protein sequences (Supplementary Table S1) from Solanum lycopersicum and other plants were compared using DNAMAN software, revealing that SlGGC1 showed high similarity with other GGC1 proteins (Fig. 1a). A phylogenetic tree was constructed using GGC1 sequences from ten species by MEGA 11 software. SlGGC1 and StGGC1 from Solanum tuberosum clustered in the same clade, suggesting the evolutionary conservation of GGC1 among plant species (Fig. 1b). SlGGC1 is composed of a Gγ-like domain and a cysteine-rich domain, indicating that SlGGC1 is a member of the Gγ family (Fig. 1c).
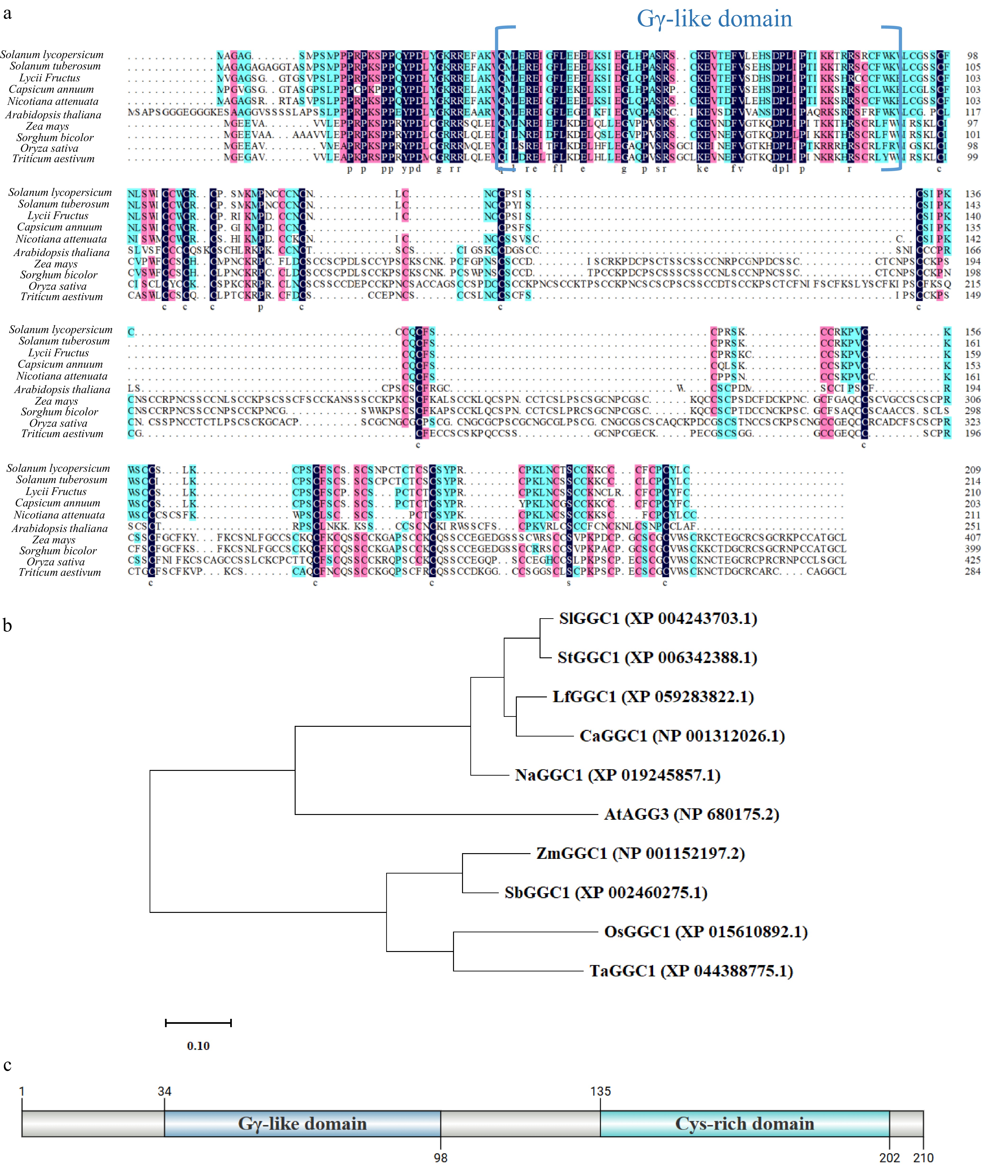
Figure 1.
SlGGC1 is highly conserved in Solanaceae. (a) Amino acid sequence alignment of GGC1 in Solanum lycopersicum, Solanum tuberosum, Lycii Fructus, Capsicum annuum, Nicotiana attenuata, Arabidopsis thaliana, Zea mays, Sorghum bicolor, Oryza sativa, and Triticum aestivum. (b) Phylogenetic relationship between tomato SlGGC1 and GGC1 of other plant species. The phylogenetic tree of multi-species GGC1 proteins was constructed using MEGA11, with an evolutionary distance scale set to 0.1. The protein sequences used for the construction, including AtAGG3 (NP_680175.2), SlGGC1 (XP_004243703.1), OsGGC1 (XP_015610892.1), ZmGGC1 (NP_001152197.2), SbGGC1 (XP_002460275.1), TaGGC1 (XP_044388775.1), StGGC1 (XP_006342388.1), LfGGC1 (XP_059283822.1), CaGGC1 (NP_001312026.1), and NaGGC1 (XP_019245857.1), were obtained from GenBank. (c) Structure of SlGGC1 protein. The SlGGC1 protein represents an atypical class of Gy proteins, characterized by a C-terminal G protein gamma subunit-like domain (GGL) and an N-terminal cysteine-rich domain.
Subcellular localization of SlGGC1
-
To investigate the subcellular localization of SlGGC1, the gene was cloned into the pK7FWG2 vector to generate the 35S:SlGGC1-GFP construct. Transient expression assays in Nicotiana benthamiana leaves were then used to analyze the localization pattern of SlGGC1. Additionally, co-localization experiments were performed with SlGGC1-GFP and a plasma membrane marker (pm-rk; CD3-1007). Green fluorescence from SlGGC1-GFP was detected at the cell membrane and showed overlap with the red fluorescence of the mCherry-tagged marker. These results indicate that SlGGC1 is localized to the cell membrane (Fig. 2)
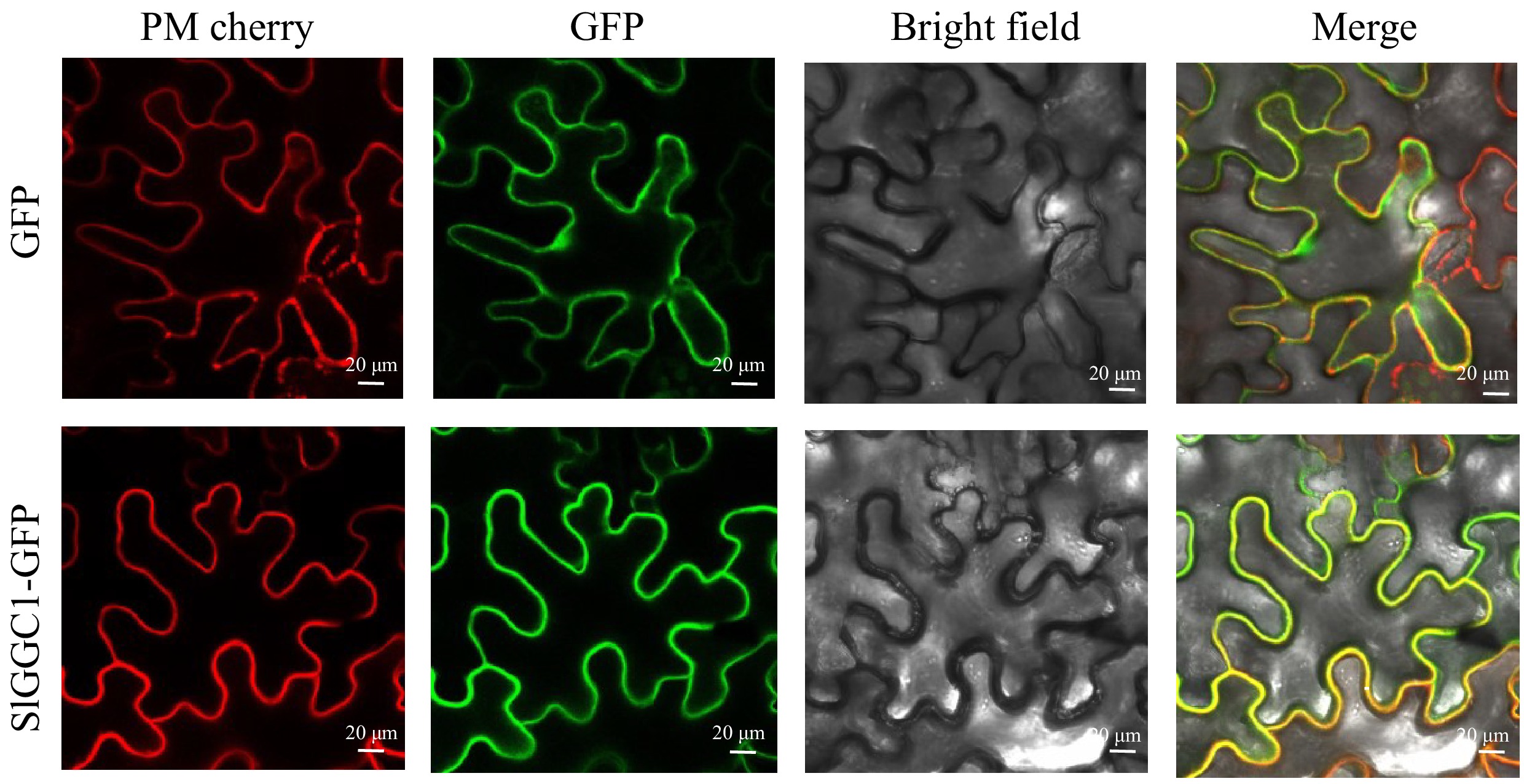
Figure 2.
SlGGC1 is localized in the plasma membrane. Subcellular localization of SlGGC1. The GFP fusion of SlGGC1 was co-expressed with the plasma membrane marker PM-mCherry in N. benthamiana leaves and visualized by confocal microscopy. 35S::GFP is used as a positive control. The GFP and mCherry signals are represented in green and red, respectively. Bars = 20 μm.
SlGGC1 expression is induced by saline-alkali stress
-
To analyze SlGGC1 expression in different tissues of tomato, transcription was assessed in root, stem, leaf, flower, and fruit from the cultivar 'AC' by qRT-PCR. The results indicated that SlGGC1 was expressed at a relatively high level in roots, at a moderate level in leaves, flowers, and fruits, and at a lower level in stems (Fig. 3a). To further characterize the response of SlGGC1 to saline-alkali stress, transcription in seedlings treated with NaHCO3 for different time points was investigated by qRT-PCR. The results showed that SlGGC1 was induced by saline-alkali stress at 6 h, and the induction peaking at 24 h (Fig. 3b). These data suggest that SlGGC1 is a potential modulator in the plant's response to saline-alkali stress.
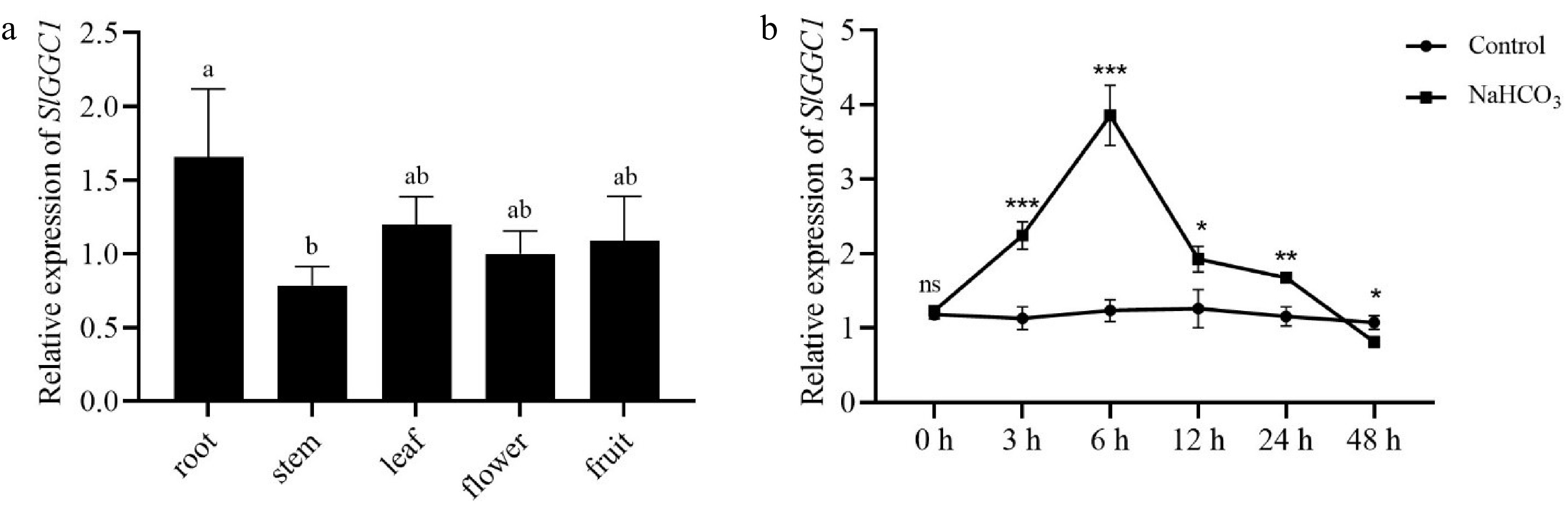
Figure 3.
SlGGC1 expression pattern in tomato and its response to saline-alkali stress. (a) Relative expression levels of SlGGC1 in root, stem, leaf, flower, and fruit detected by qRT-PCR. Statistical significance was determined by one-way ANOVA. Significant differences are indicated by different lowercase letters. (b) The expression of SlGGC1 with or without NaHCO3 treatment. Total RNA was extracted from 5-day-old seedlings treated with or without 10 mM NaHCO3 for 0, 3, 6, 12, 24, and 48 h. The experiments were performed with three biological replicates with similar results. Asterisks indicate significant differences between control and NaHCO3 treatment (* p < 0.05, ** p < 0.01, *** p < 0.001).
SlGGC1 is a positive regulator of saline-alkali tolerance in tomato
-
To further investigate whether SlGGC1 is involved in response to saline-alkali stress, the slggc1-cr mutant was constructed (Supplementary Fig. S1). Saline-alkali stress impedes plant growth, and one of its manifestations is the inhibition of primary root elongation. Under saline-alkali stress, the reduction of primary root elongation of AC, slggc1-cr-1 and slggc1-cr-2 were reduced by 61.4%, 72.8%, and 78.5% respectively (Fig. 4a, b). Similarly, the fresh weight of 'AC', slggc1-cr-1, and slggc1-cr-2 was reduced by 34.8%, 46.1%, and 43.3%, respectively, when exposed to saline-alkali stress (Fig. 4c, d). Given that salt stress inhibits seed germination, the germination tolerance of 'AC' and slggc1-cr under saline-alkali stress was investigated. In the absence of NaHCO3 treatment, no significant difference in germination rate was observed between 'AC' and slggc1-cr. By contrast, upon NaHCO3 exposure, the germination rate of slggc1-cr was markedly lower than that of 'AC' (Supplementary Fig. S2). These findings strongly indicate that the slggc1-cr mutants display reduced tolerance to saline-alkali stress.
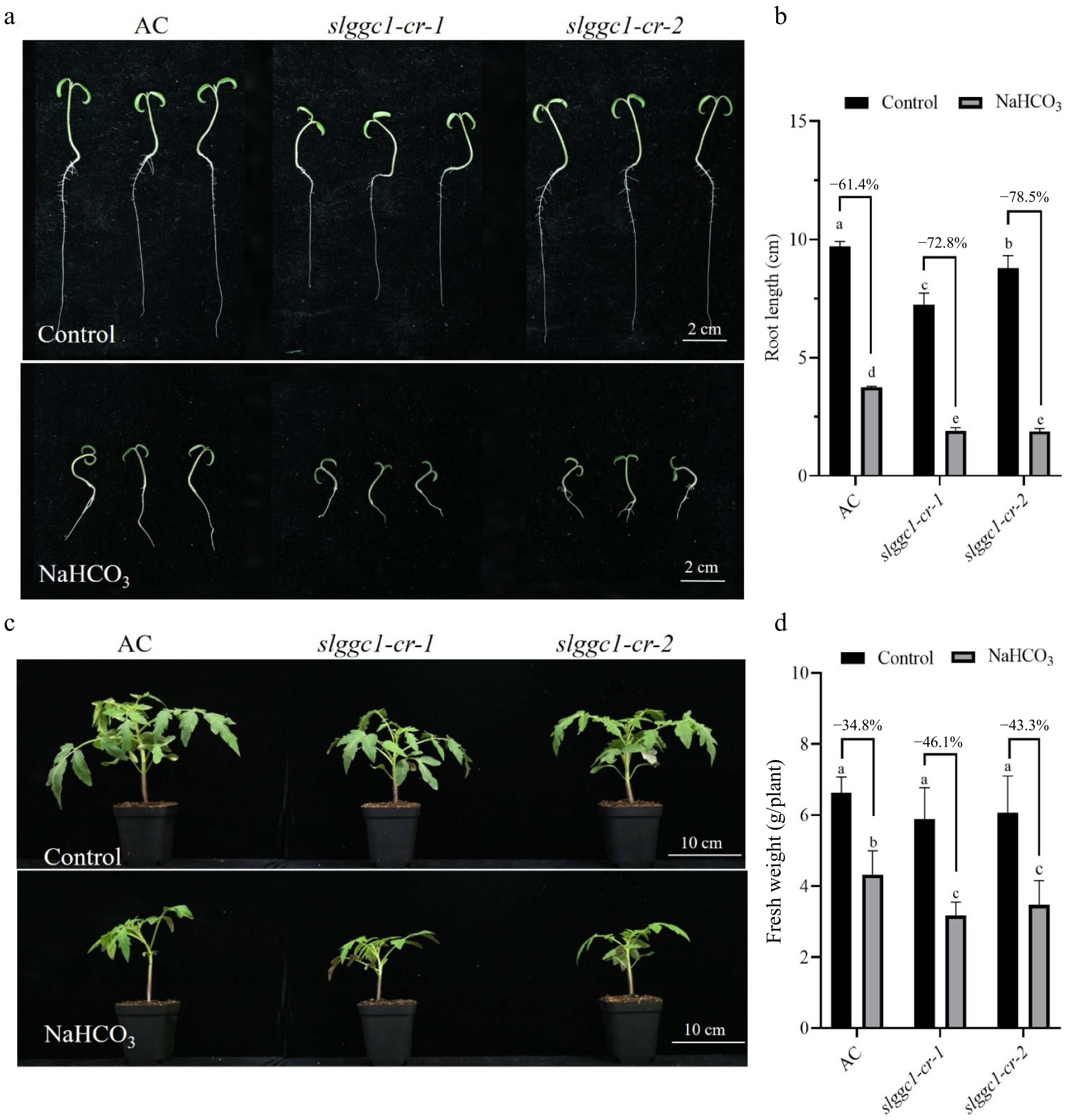
Figure 4.
slggc1 is more sensitive to saline-alkali stress. (a) Phenotype analysis of 'AC' and slggc1 grown on Murashige and Skoog (MS) medium without or with 10 mM NaHCO3. (b) The root length of 'AC' and slggc1 under saline-alkali stress for 5 d. Seedlings were cultivated in a climate chamber with 60% relative humidity under a 16 h light period at 26 °C and an 8 h dark period at 20 °C. (c) Phenotype analysis of three-week-old 'AC' (wild-type) and slggc1 grown in soil without or with 75 mM NaHCO3. (d) The biomass of AC and slggc1 grown under saline-alkali stress for 21 d. The experiments were performed with three biological replicates with similar results. Statistical significance was determined by two-way ANOVA, p < 0.05. Significant differences are indicated by different lowercase letters.
SlGGC1 regulates Na+ and K+ accumulation during saline-alkali stress
-
The balance of Na+ and K+ is important for plants in response to salt stress, and excess Na+ accumulation greatly damages plant growth. Hence, the compromised saline-alkali tolerance in slggc1-cr could be caused by overaccumulation of Na+. To test this hypothesis, the contents of Na+ and K+ in shoots and roots of 'AC' and slggc1-cr exposed to saline-alkali stress were analysed. Compared with 'AC', shoots of slggc1-cr showed a higher level of Na+ and a similar level of K+ with NaHCO3 treatment, which led to a higher Na+/K+ ratio in slggc1-cr shoots (Fig. 5a–c). Roots of slggc1-cr-1 showed a higher level of Na+, and slggc1-cr-2 showed a lower level of K+ in comparison with 'AC', which also led to a higher Na+/K+ ratio in slggc1-cr roots with NaHCO3 treatment (Fig. 5d–f). These lines state that the homeostasis of Na+ and K+ is disturbed in slggc1-cr, and SlGGC1 is a regulator of Na+ and K+ accumulation during saline-alkali stress.
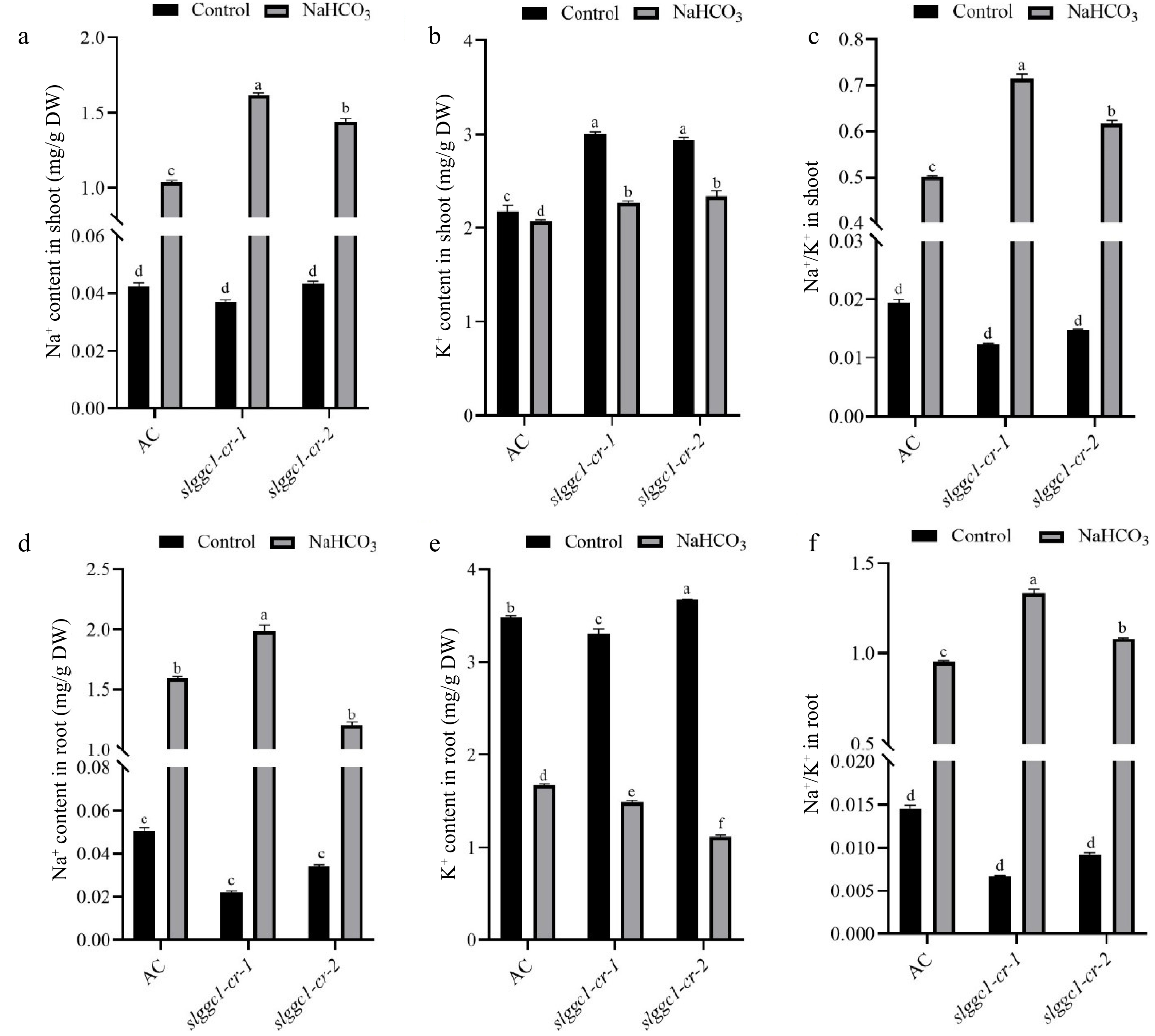
Figure 5.
SlGGC1 regulates Na+ and K+ accumulation under saline-alkali stress. The Na+ and K+ contents in AC and slggc1 grown under control and saline-alkaline (75 mM NaHCO3) conditions. (a) Na+ content, (b) K+ content, and (c) Na+/K+ ratio in the shoots of AC and slggc1 were measured after three weeks of treatment with 75 mM NaHCO3. (d) Na+ content, (e) K+ content, and (f) Na+/K+ ratio in the roots of AC and slggc1 were measured after three weeks of treatment with 75 mM NaHCO3. The Na+ and K+ contents were measured by inductively coupled plasma-mass spectrometry (ICP-MS). The experiments were performed with three biological replicates with similar results. Statistical significance was determined by two-way ANOVA, p < 0.05. Significant differences are indicated by different lowercase letters.
SlGGC1 modulates ROS accumulation in response to saline-alkali stress
-
ROS is an important secondary messenger and cytotoxic substance when plants are subjected to biotic and abiotic stresses. Due to the reduced saline-alkali tolerance in slggc1-cr, it was hypothesized that ROS were overaccumulated in slggc1-cr. H2O2 and superoxide anions were investigated using DAB and NBT staining, and H2O2 and superoxide anions were quantitatively measured. 'AC' and slggc1-cr showed similar levels of H2O2 and superoxide anions without NaHCO3 treatment. Saline-alkali stress promoted the accumulation of H2O2 and superoxide anions, but the promotion was greater in slggc1-cr than in 'AC' (Fig. 6a–d). The slggc1-cr mutant had significantly higher malondialdehyde (MDA) contents and lower GSH contents under saline-alkali stress than 'AC' (Fig. 6e, f), suggesting that slggc1-cr suffered more oxidative damage than 'AC'. Antioxidant enzyme activities, including those of SOD, POD, and catalase (CAT), are subject to changes when plants respond to stress. Under normal growth conditions, no differences in SOD, POD, or CAT activities were observed between 'AC' and slggc1-cr. Saline-alkali stress activated the enzyme activities, but the activation in slggc-cr was weaker than that in 'AC' (Fig. 6g–i). All these findings demonstrate that SlGGC1 is involved in the modulation of ROS homeostasis to counteract saline-alkali stress.
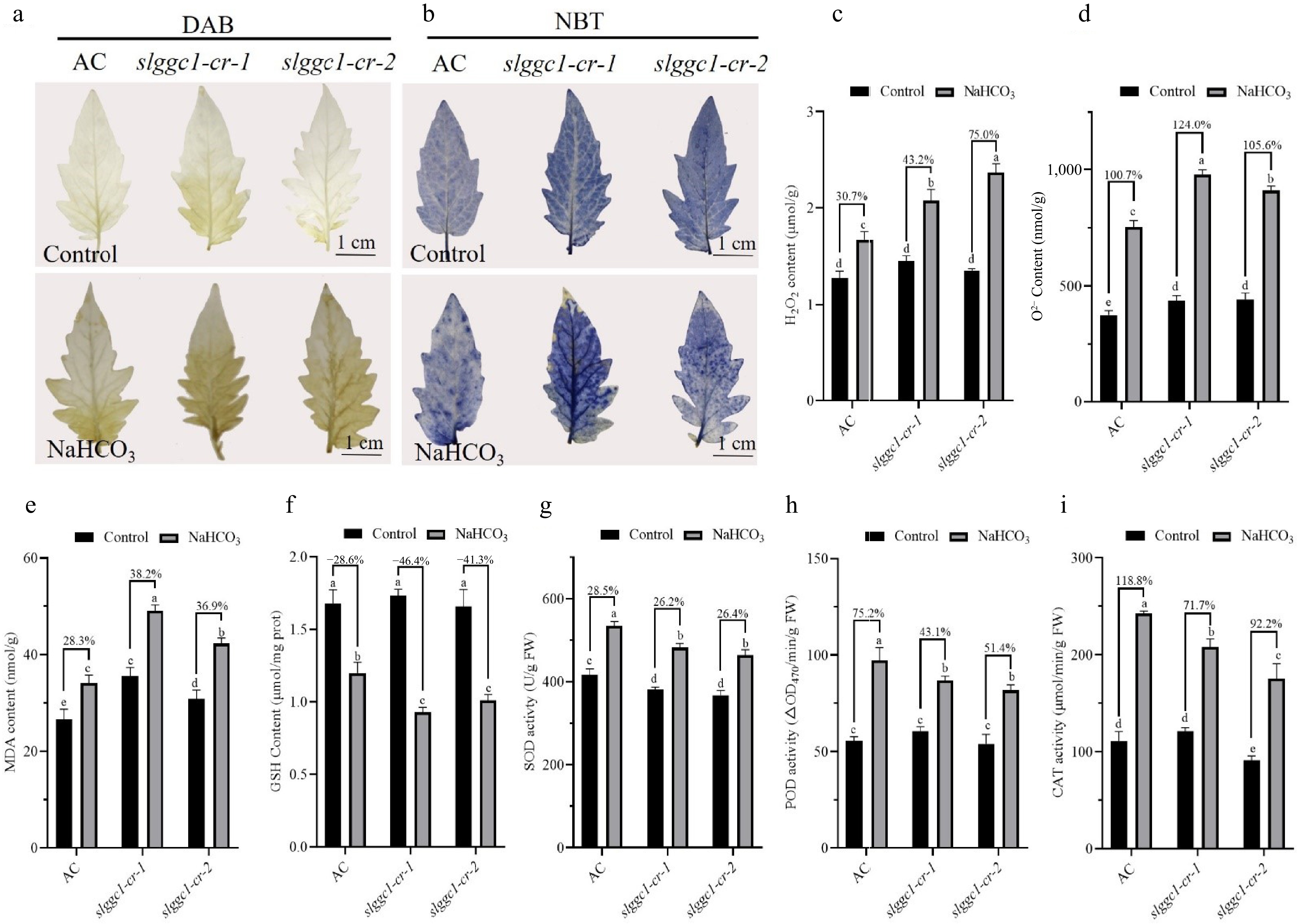
Figure 6.
SlGGC1 regulates ROS homeostasis under saline-alkali stress. (a) 3,3′-Diaminobenzidine (DAB) staining. (b) Nitrotetrazolium chloride blue (NBT) staining. (c) H2O2, and (d) superoxide anion content of 'AC' and slggc1 cultivated in saline-alkaline conditions for 21 d. (e) MDA, (f) GSH content of 'AC' and slggc1 cultivated in saline-alkaline conditions for 21 d. The enzymatic activities of (g) SOD, (h) POD, and (i) CAT in 'AC' and slggc1 were measured after 8 h of saline-alkali treatment. The experiments were performed with three biological replicates with similar results. Statistical significance was determined by two-way ANOVA, p < 0.05. Significant differences are indicated by different lowercase letters.
SlGGC1 modulates ABA signaling in response to saline-alkali stress
-
Abscisic acid (ABA) is a crucial plant hormone implicated in abiotic stress responses. To determine whether the impaired saline-alkali tolerance in the slggc1-cr mutant is due to defects in the ABA signaling pathway, the expression of several ABA signaling-associated marker genes was checked under saline-alkali stress conditions. It was observed that the expression of all these genes was upregulated by saline-alkali stress in both WT and slggc1-cr. However, the upregulation was significantly attenuated in slggc1-cr compared to that of AC (Fig. 7a–d). These results suggest that the compromised saline-alkali tolerance in slggc1-cr could be partially ascribed to defects within the ABA signaling pathway.
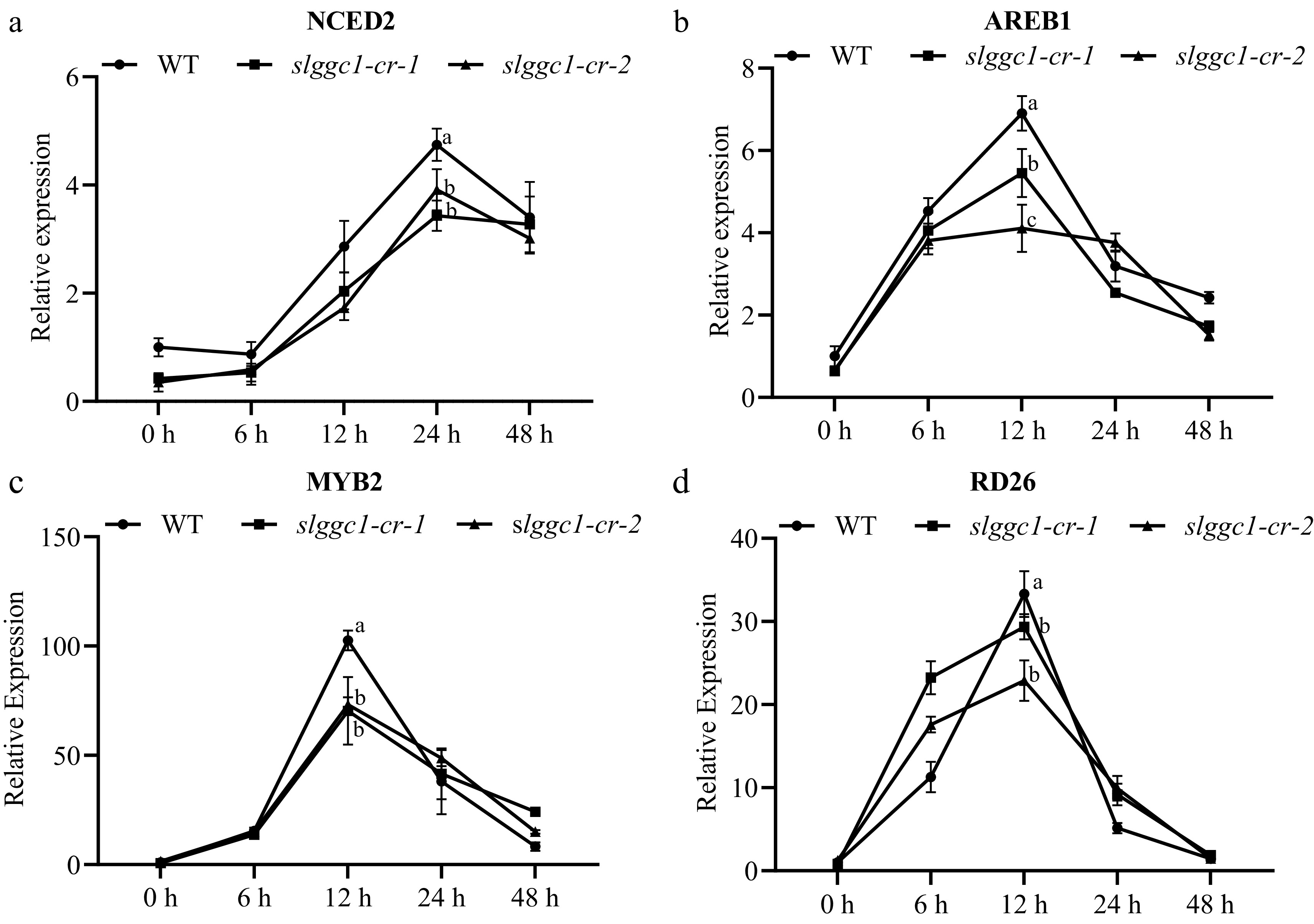
Figure 7.
SlGGC1 affects ABA signaling in response to saline-alkali stress. The expression levels of (a) NCED2, (b) AREB1, (c) MYB2, and (d) RD26 in seedlings of WT and slggc1-cr were assessed at 0, 6, 12, 24, and 48 h following treatment with 10 mM NaHCO3 on MS Petri dishes. The expression level of WT at 0 h was set to 1. The EF1α gene was used as the internal control for qRT-PCR. The experiments were performed with three biological replicates with similar results. Statistical significance was determined by two-way ANOVA, p < 0.05. Significant differences are indicated by different lowercase letters.
-
In this study, the role of the G protein γ subunit SlGGC1 in tomato's response to saline-alkali stress was systematically investigated, revealing several critical aspects of its function and underlying mechanisms.
Bioinformatics analysis revealed that SlGGC1 shares high sequence similarities with GGC1 proteins from other plant species and clusters closely with StGGC1 from Solanum tuberosum, indicating evolutionary conservation of this protein. The conserved domain structure, including the Gγ-like domain and cysteine-rich domain, further confirms its classification within the Gγ family. This conservation suggests that GGC1 proteins likely play similar and evolutionarily important roles in different plant species, potentially related to stress-response regulation. Subcellular localization results showed that SlGGC1 is localized to the cell membrane, consistent with the typical membrane association of G protein subunits. This membrane-bound localization is crucial for its potential interaction with other membrane-associated proteins or receptors, which may be essential for signal transduction processes in response to saline-alkali stress. The expression pattern of SlGGC1 provides important clues about its function. The relatively high expression in roots, where plants first encounter soil-borne stresses such as salinity and alkalinity, suggests that SlGGC1 may play a key role in the initial perception and response to these stresses. Additionally, the induction of SlGGC1 transcription by saline-alkali stress, peaking at 24 h, indicates that it is an integral part of the plant's adaptive response mechanism to such environmental challenges.
Analysis of the slggc1-cr mutants demonstrated that SlGGC1 acts as a positive regulator of saline-alkali tolerance in tomato. Under saline-alkali stress, the mutants showed more severe growth inhibition, as evidenced by greater reduction in primary root elongation and fresh weight compared to the wild type. This finding strongly supports the role of SlGGC1 in maintaining plant growth and development under adverse conditions.
One of the key mechanisms by which SlGGC1 contributes to salin-alkali tolerance is through the regulation of ion homeostasis. The slggc1-cr mutants exhibited disturbed Na+/K+ balance, with higher Na+ accumulation and altered K+ levels in both shoots and roots, leading to a higher Na+/K+ ratio. Since maintaining a proper Na+/K+ balance is essential for plant survival under salt and alkali stress, the disruption of this balance in the mutants likely accounts for their reduced tolerance. This indicates that SlGGC1 may be a potential regulator of ion transporters or channels to control Na+ influx and K+ retention.
Another important aspect of SlGGC1 function is its modulation of reactive oxygen species (ROS) accumulation. The Gγ subunit AT1 (alkaline tolerance 1) exerts a negative regulatory effect on the phosphorylation of PIP2 aquaporins, which in turn diminishes the H2O2 efflux capacity of these PIP2 proteins. Such impairment results in excessive H2O2 accumulation, ultimately rendering the crops susceptible to alkali stress[32,33]. Given that AT1 is the tomato homologue of SlGGC1, it is reasonable to speculate that SlGGC1 also modulates saline-alkali stress responses by maintaining reactive oxygen species (ROS) homeostasis. Under saline-alkali stress, the slggc1-cr mutants showed excessive accumulation of H2O2 and superoxide anions, increased oxidative damage (higher MDA content and lower GSH content), and weaker activation of antioxidant enzymes. ROS overaccumulation can cause oxidative stress and damage cellular components, and the inability of the mutants to properly regulate ROS levels likely exacerbates the negative effects of saline-alkali stress on plant growth. This suggests that SlGGC1 shares functional similarity with AT1 and may be involved in ROS scavenging pathways or the activation of antioxidant defense systems.
Finally, SlGGC1 was also found to modulate the abscisic acid (ABA) signaling pathway in response to saline-alkali stress. The expression of ABA signaling-related marker genes was significantly less upregulated in the slggc1-cr mutants compared to the wild type under saline-alkali stress. As ABA is a key hormone in plant stress responses, defects in ABA signaling in the mutants may contribute to their reduced tolerance. SlGGC1 may interact with components of the ABA signaling pathway to enhance the plant's adaptive response to saline-alkali stress, which should be further investigated.
In conclusion, this study demonstrates that SlGGC1 is a multifunctional positive regulator of tomato's response to saline-alkali stress. It functions by maintaining ion homeostasis, regulating ROS accumulation, and modulating ABA signaling. These findings not only expand our understanding of plant responses to saline-alkali stress but also provide potential targets for genetic engineering to improve plant tolerance to such challenging environments. Future studies could focus on elucidating the specific molecular interactions of SlGGC1 with other components in these regulatory pathways to further understand its mode of action.
This work was supported by the National Natural Science Foundation of China-Shandong Joint Fund (Grant No. U22A20459 to Chuanyou Li and Qian Chen), the National Natural Science Foundation of China (Grant No. 32272701 to Qian Chen), the National Key Research and Development Program of China (Grant No. 2022YFD1201700 to Xianwen Meng), Agricultural Seed Project of Shandong Province (Grant No. 2021LZGC017 to Chuanyou Li and Qian Chen).
-
The authors confirm contributions to the paper as follows: study conception and design: Chen Q, Li C; experiments: Zhang YR, Liu J, Sun H, Zang Y, Zhang Y, Wang H; data analysis: Sun C, Deng L, Meng X, Chen Q, Li C; manuscript preparation: Zhang YR, Liu J, Chen Q, Li C. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated and/or analyzed in this study can be obtained from the corresponding author upon reasonable request.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yanru Zhang, Jian Liu, Heyao Sun, Yinqiang Zang
- Supplementary Table S1 List of SlGGC1 and GGC1 of other plant species.
- Supplementary Table S2 Primers used in this study.
- Supplementary Fig. S1 Generation of SlGGC1 mutants via CRISPR/Cas9-based approach.
- Supplementary Fig. S2 SlGGC1 positively regulates seeds germination against saline-alkali stress in tomato.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang Y, Liu J, Sun H, Zang Y, Zhang Y, et al. 2025. The G protein γ subunit SlGGC1 regulates saline-alkali stress response in tomato. Vegetable Research 5: e033 doi: 10.48130/vegres-0025-0027 

The G protein γ subunit SlGGC1 regulates saline-alkali stress response in tomato
- Received: 30 April 2025
- Revised: 18 June 2025
- Accepted: 02 July 2025
- Published online: 12 September 2025
Abstract: Saline-alkali stress poses a severe threat to crop yield and quality. While significant progress has been achieved in elucidating the mechanisms of plant resistance to salt stress, the molecular underpinnings governing plant responses to saline-alkali stress remain largely unclear. This study investigated the role of the G-protein γ-subunit, SlGGC1, from Solanum lycopersicum, in modulating the plant's response to saline-alkali stress. The findings revealed that SlGGC1 was predominantly localized in the cell membrane, and its transcription was induced by saline-alkali stress conditions. Analysis of SlGGC1 mutants demonstrated heightened sensitivity to saline-alkali stress, manifested as severe growth inhibition, excessive accumulation of reactive oxygen species (ROS), disruption of Na+/K+ homeostasis, and dysregulation of abscisic acid (ABA) signaling pathways. Collectively, these results provide compelling evidence that SlGGC1 acts as a positive regulator of the saline-alkali stress response in tomato plants.
-
Key words:
- G protein /
- SlGGC1 /
- Saline-alkali stress /
- Tomato