









-
Jasmonate (JA) and its derivatives are generally used as plant growth regulating compounds. These biomolecules have specific functions in plant responses to differing biotic and abiotic stresses[1,2]. The induction in plant immunity post JA application has been previously reported against cold stress[3], drought stress[4], high temperature stress[5] and salt stress[6]. Additionally, JA can regulate various aspects of growth and development, including seed germination[7], root growth[8], hypocotyl growth[9], stamen development[10], and flowering and blooming[11]. In general, JA in plants is synthesized from α-Linolenic acid (ALA) via the octadecane pathway[12]. Firstly, the synthesis of unsaturated fatty acids to 12-oxo-phytadienoic acid (12-OPDA) is completed in the chloroplast, which is oxidized to 13-hydroperoxylinolenic acid (13-HPOTE) under the action of lipoxygenase (LOX), which is then oxidized to 12-oxo-phytadienoic acid under the catalysis of allene oxide synthase (AOS) and allene oxide cyclase (AOC). With the help of transporter proteins, 12-OPDA is transported to the peroxisome for downstream reaction, and JA is finally formed after 12-OPDA reductase and a three-step β oxidation reaction. After JA is transported to the cytoplasm, active JAs (+)-7-iso-JA-L-isoleucine ((+)-7 is formed under the action of JA amino acid synthetase (JAR1) -iso-JA-Ile)[13].
The JAZ (JASMONATE ZIM domain protein) repressor protein is a key stress marker gene, specific to plants. JAZ belongs to the TIFY family, which is characterized by a conserved TIFY motif (also known as ZIM), and there is a TIF[F/Y] XG near the N-terminus, which exhibits homo- and heteromeric interactions[14]. Its C-terminus also contains a Jas motif SLX2FX2KRX2RX5PY (also known as CCT_2). The existence of the Jas motif makes the JAZ protein different from other members of the TIFY protein family[15]. JAZ does not contain a DNA-binding domain and is be functionally dependent on protein interactions[15]. Under normal conditions, the plant endogenous JA content is low, the JAZ inhibitor binds to the transcription factors (TFs), and recruits the co-inhibitor TOPLESS (TPL) by interacting with the adaptor protein NINJA (novel interactor of JAZ, NINJA) and TPL-related protein (TPR) to inhibit the activity of TFs. The JAZ inhibitor protein tightly binds to the MYC2 transcription factor required to inhibit JA-induced gene expression. The JAZ protein targeted by the SCFCOI1 complex plays a negative regulatory role in the JA signaling pathway. In the process of hormone accumulation and perception, the active JA-Ile is identified by SCFCOI1, which promotes the binding of JAZ protein and COI1 (CORONATINE INSENSITIVE 1)[16,17], and facilitates the ubiquitination and degradation of the JAZ protein. Moreover, the 26S proteasome releases transcription factors from NINJA and TPL, leading to the release of MYC2 transcription factor, initiating transcriptional reprogramming of cells, and activating the JA response[18−20].
Cucumber (Cucumis sativus L.) (2n = 2x = 14) belongs to the family Cucurbitaceae and is an important vegetable crop. Cucumber is widely cultivated for its fruit in warm climates around the world. Waterlogging is becoming an increasing threat causing abiotic stress owing to current climate change. Under the background of frequent rainfall and flood disasters caused by global warming, regional seasonal rainfall continues to rise which leads to the upsurge of the water table, resulting in waterlogged soil[21−23]. Cucumber is extremely sensitive to waterlogging stress due to its less developed root system along with poor water absorption capacity[24,25]. The publication of the cucumber genome sequence in 2009[26] made it feasible to carry out extensive molecular work relating to the breeding of cucumber. Further, the availability of the cucumber genome database made it significantly convenient in the identification of genes controlling various agronomic traits and regulating cucumber response to environmental stress.
Since JA plays a key role in enhancing plant tolerance to a variety of stresses, JA could be of great importance to elucidate the transcriptional regulatory mechanism of JAZ protein under waterlogging stress in cucumber. Although many members of the JAZ gene family have been extensively identified and studied in various crops[14,27−31], the function and regulatory mechanisms regarding this gene family in cucumber waterlogging tolerance remain poorly understood. The present study comprised of bioinformatics analysis of the JAZ gene family, a key gene family in the jasmonic acid signal transduction pathway in cucumber. A complete set of 11 CsJAZ genes were identified and characterized. Additionally, the phylogenetic relationship, protein conserved motif, gene structure analysis, chromosomal location, gene duplication, selective evolutionary pressure, cis-acting regulatory analysis, and transcription expression of the cucumber JAZ gene under waterlogging stress were analysed in different tissues under hormone application. We also examined the subcellular localization of CsJAZ8 in tobacco cells and characterized their homologous or heterologous interactions with the JAZ family. This study presents a theoretical foundation for further studies regarding the function of the CsJAZ gene in cucumber, and may provide a new development in cucumber breeding for waterlogging tolerance.
-
In order to identify the CsJAZ genes from the cucumber genome, SMART and NCBI CD-searches were carried out to confirm the JAZ protein from their possession of conserved TIFY and Jas domains in the generated amino acid coding sequence. Next, 11 non-redundant CsJAZ genes were identified in the cucumber genome. According to the chromosomal location of these genes, all CsJAZ genes are systematically numbered, from CsJAZ1 to CsJAZ11. The protein characteristics of the identified JAZ genes were studied using the protparam online server. The length of the protein sequence encoded by these CsJAZ genes ranged from 129aa (CsJAZ7) to 381aa (CsJAZ8). The molecular weight of the proteins were between 14,743.72 kDa (CsJAZ10) to 39,666.79 kDa (CsJAZ8). The isoelectric point ranged from 6.71 (CsJAZ4) to 10.42 (CsJAZ7), with most of them being concentrated at 9. The hydrophilicity and hydrophobicity of 11 CsJAZ protein sequences were predicted, and the results showed that CsJAZ proteins were all hydrophilic proteins (GRAVY < 0) (Table 1).
Table 1. Summary information on the CsJAZ gene family in cucumber.
Gene name Gene identifier Chr Start (bp) End (bp) ORF (aa) MW (KDa) PI GRAVY CsJAZ1 Csa1G042920 1 4573626 4577405 340 36 619.75 9.2 −0.25 CsJAZ2 Csa1G435720 1 15995968 15996717 150 16 930.92 7.21 −0.767 CsJAZ3 Csa1G597690 1 22692966 22694494 231 25 028.26 9.98 −0.455 CsJAZ4 Csa2G222060 2 10711729 10716520 336 36 967.37 6.71 −0.816 CsJAZ5 Csa3G645940 3 25350295 25351576 209 22 874.76 8.94 −0.631 CsJAZ6 Csa4G009880 4 1470022 1476285 200 22 487.17 6.98 −0.553 CsJAZ7 Csa4G062400 4 4986428 4991013 129 14 861.18 10.42 −0.425 CsJAZ8 Csa5G628650 5 25425307 25429296 381 39 666.79 9.3 −0.236 CsJAZ9 Csa6G091930 6 6249153 6251831 184 20 306.27 9.41 −0.317 CsJAZ10 Csa6G523460 6 28126619 28128465 132 14 743.72 9.5 −0.805 CsJAZ11 Csa7G448810 7 18408423 18411214 295 32 092.09 9.31 −0.758 Phylogenetic, gene structure, and conserved motifs analysis of CsJAZ genes in Cucumis sativus
-
To gain insightful knowledge regarding the evolutionary relationship of JAZ genes from cucumber, we utilized MEGA X software to perform ClustalW multiple sequence alignment of the JAZ protein sequences, retrieved from cucumber, Arabidopsis thaliana, Populus trichocarpa, Vitis vinifera, Solanum lycopersicum, and Oryza sativa. An unrooted phylogenetic neighbor-joining tree was constructed on JAZ protein using MEGA X software. It can be seen from the phylogenetic tree that these JAZs are divided into 3 independent branches (Group I − Group III) according to the homology relationship (Fig. 1a). Among these subgroups, there are three CsJAZ genes in Group1 (CsJAZ3, CsJAZ5 and CsJAZ11), accounting for about 27.3% of the total. Group II has four CsJAZ genes (CsJAZ1, CsJAZ6, CsJAZ8 and CsJAZ9). Group3 has four CsJAZ Genes (CsJAZ2, CsJAZ4, CsJAZ7 and CsJAZ10). The results additionally showed that the evolutionary similarity of cucumber JAZ protein with Populus trichocarpa, Vitis vinifera, Arabidopsis thaliana, and Solanum lycopersicum is higher than that of Oryza sativa. This is consistent with the fact that the former four species and cucumber are dicotyledonous plants. The emergence time is later than the differentiation from the lineage of monocots. No species-specific population was found in the phylogenetic tree of the cucumber CsJAZ gene. Although there are differences between each subfamily of different species, members within the same subfamily are similar in nature, which indicates the conservative nature of JAZ genes in plantae.
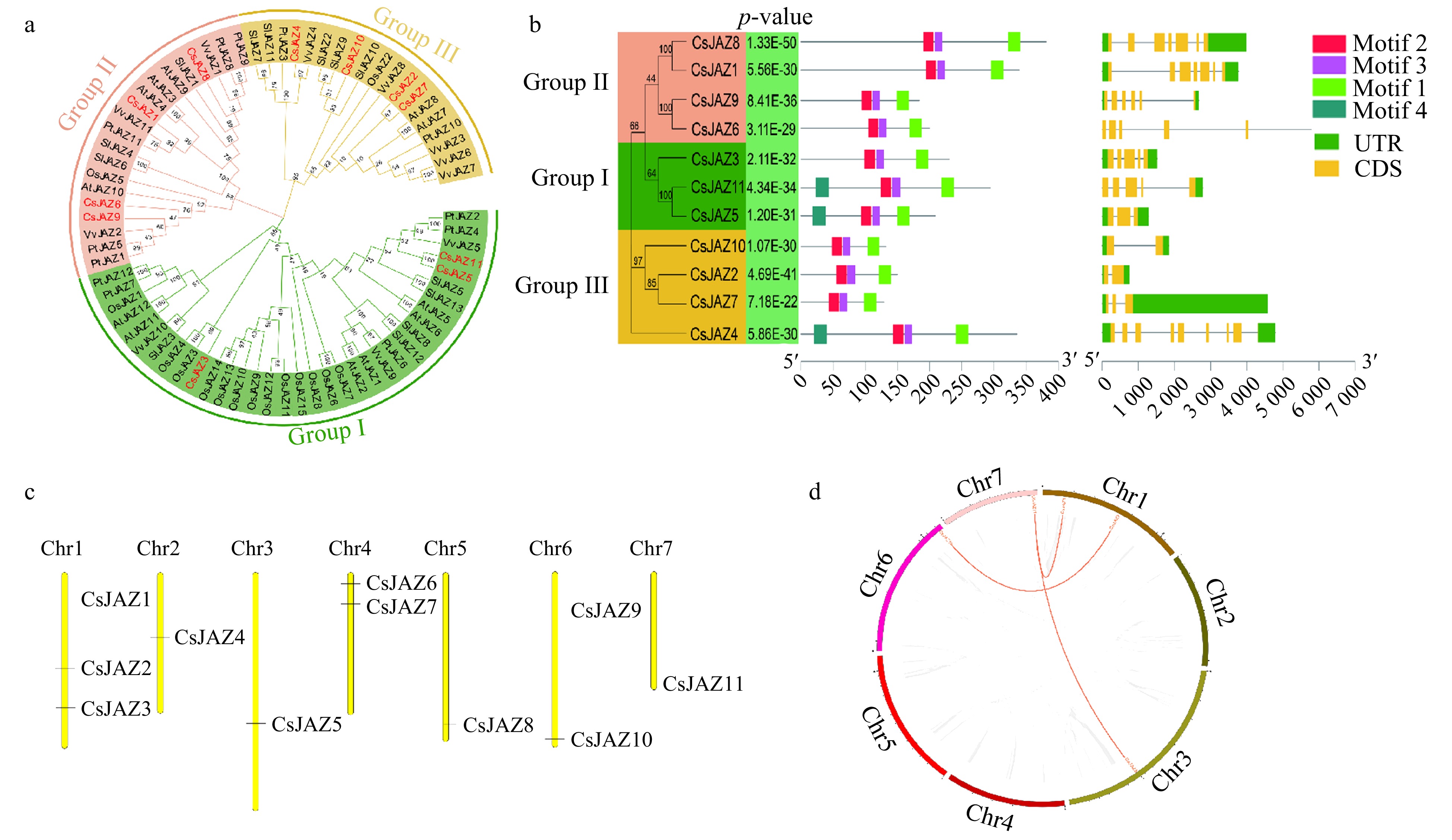
Figure 1.
(a) Phylogenetic analysis of JAZ proteins in 11 C. sativus (Cs), 12 A. thaliana (At), 13 S. lycopersicum (Sl), 15 O. sativa (Os), 11 V. vinifera (Vv), and 12 P. trichocarpa (Pt). ClustalW multi-sequence alignment was performed and an unrooted phylogenetic neighbor-joining tree was constructed fromon JAZ protein by using MEGA X software. Genes IDs are listed in Supplemental Table S3. (b) Phylogeny, gene structure, and motif compositions of the JAZ gene family in cucumber. (c) Chromosome distribution of CsJAZ in the genome of cucumber. The chromosome numbers were demonstrated at the top of each bar and the scale is in megabases (Mb). (d) Gene duplication of CsJAZ genes on 7 chromosomes of cucumber genome. Gray lines represent all syntenic blocks in the cucumber genome. Duplicated gene pairs are identified with red lines inside the circle. Each chromosome number is represented using different colours.
The evolution of gene families is largely manifested in the variation of conserved motifs in family members[32]. The MEME online prediction tool was used to identify the conserved motifs in the CsJAZ protein of cucumber (Fig. 1b and Table 2). All the 11 CsJAZ genes contain either motif 1 (Jas) and motif 2 (TIFY). In the three subgroups, CsJAZ genes contain motifs ranging from 1 to 4. It is worth noting that closely related genes in the phylogenetic tree have similar gene structural components, indicating similar functions within subgroups. The results show that all CsJAZ proteins contain conserved motifs. The evolution of gene families is largely reflected in the diversity of gene structures inside the family members[32]. In order to better understand the structure of the CsJAZ gene of cucumber, the exon-intron structure of the CsJAZ gene was analyzed using the website Gene Structure Display Server (GSDS) (
http://gsds.cbi.pku.edu.cn ) (Fig. 1b). The CsJAZ genes ranged from 2 (CsJAZ2 and CsJAZ10) to 9 (CsJAZ4) exons and 1 to 8 introns. The clustering results with the evolutionary tree showed that genes with similar gene structures clustered together. In Group I, the two genes CsJAZ3 and CsJAZ11 both have 5 exons and 4 introns, whereas CsJAZ5 has 3 exons and 2 introns. Both CsJAZ1 and CsJAZ8 in Group II have 7 exons and 6 introns, the CsJAZ6 and CsJAZ9 in Group II have 6 exons and 5 introns. In Group III, CsJAZ2 and CsJAZ10 both have 2 exons and 1 intron, CsJAZ7 has 3 exons and 2 introns, CsJAZ4 has 9 exons and 8 introns. We observed that the CsJAZ genes are both conservative and variable in the Group, which may be related to the different modes of action and distribution in cucumber.Table 2. Information on conserved motifs in cucumber JAZ proteins.
Motif E-value Sites Width Sequence Logo 1 1.6e-077 11 20 DLPJARKASLQRFLEKRKDR 
2 5.7e-074 11 15 QMTIFYNGKVCVYBD 
3 2.6e-0.13 11 15 PEDKAKAIMALASKG 
4 4.0e-003 3 21 EKSNFAQTCNLLSQYLKEKRT 
Chromosomal location, duplication events, selective evolutionary pressure analysis of CsJAZ genes in Cucumis sativus
-
The distribution of cucumber CsJAZ genes on chromosomes were visualized using MapChart software (Fig. 1c). The results showed that 11 CsJAZ genes were distributed on seven chromosomes of cucumber. There were three CsJAZ genes on the Chr1 accounting for 27.3% of the total. Chr2, Chr3 and Chr5 have only one CsJAZ gene. There were two CsJAZ genes on Chr4, Chr6 and Chr7, accounting for 18.2% of the total. By mapping CsJAZ genes on chromosomes, we found that although the number of CsJAZ genes was small, they were distributed on each chromosome where chromosome 1 possessed the highest number of genes.
To investigate the gene duplication events, we used the Multiple Collinearity Scan toolkit (MCScanX) with default parameters to detect the duplicated blocks in the cucumber genome. The segment duplication events were characterized by copying the whole gene block from one chromosome to another, while adjacent homologous genes with no more than one intervening gene on the same chromosome were considered a tandem duplication event. We analyzed the tandem and segment duplication genes in the whole genome of cucumber. Three gene pairs were generated from five tandem duplicated genes (Fig. 1c and Supplemental Table S1). Meanwhile, five segment duplicated genes were identified and they formed three pairs (Supplemental Table S1). Combined with the phylogenetic tree (Fig. 1a), we found that the majority of duplicated pairs were clustered together in the same subgroups. The ratio Ka/Ks of non-synonymous substitution (Ka) and synonymous substitution (Ks) can reflect the selection criterion during biological evolution. As a result, the Ka/Ks ratios between CsJAZ genes were all greater than one, indicating that CsJAZ genes were mainly subjected to positive selection (Supplemental Table S2).
Putative cis-acting regulatory elements in the promoter regions of CsJAZ genes
-
In order to obtain information on the cis-acting regulatory elements of the CsJAZ gene family, we analyzed the sequence of the promoter region of each CsJAZ gene (Supplemental Table S4). The −1500 bp promoter regions upstream of 11 CsJAZ genes were extracted and the cis-acting elements were analyzed using the Plant CARE website (Fig. 2). The results uncovered the potential regulatory role of CsJAZ genes as they carry hormones, growth and stress related cis-acting elements (Supplemental Table S5). Hormonal responsive cis-elements such as TCA-element and SARE (involved in salicylic acid responsiveness), ABRE (involved in abscisic acid responsiveness), CGTCA-motif and TGACG-motif (involved in MeJA-responsiveness) were found dominantly in the majority of the CsJAZ genes; Meanwhile, auxin responsive cis-acting elements known as TGA-box and TGA-element, gibberellin-responsiveness cis-acting elements named TATC-box and P-box were also identified in the upstream region of some genes. The presence of cis-acting elements involved in light responsiveness named G-box, GATA-motif, GT1-motif, and circadian which are related to circadian control, highlights the diverse roles of CsJAZ genes in regulating growth and providing response to environmental stimuli. In addition, some other stress-responsive cis-acting regulatory elements named LTR (low temperature-responsive), ARE (anaerobic induction cis-elements), TC-rich repeats (involved in defense and stress) were also recognized in the promoter region of CsJAZ genes. Therefore, the presence of these important cis-acting elements in CsJAZ genes confirms their potential involvement in controlling multiple biological functions in cucumber.
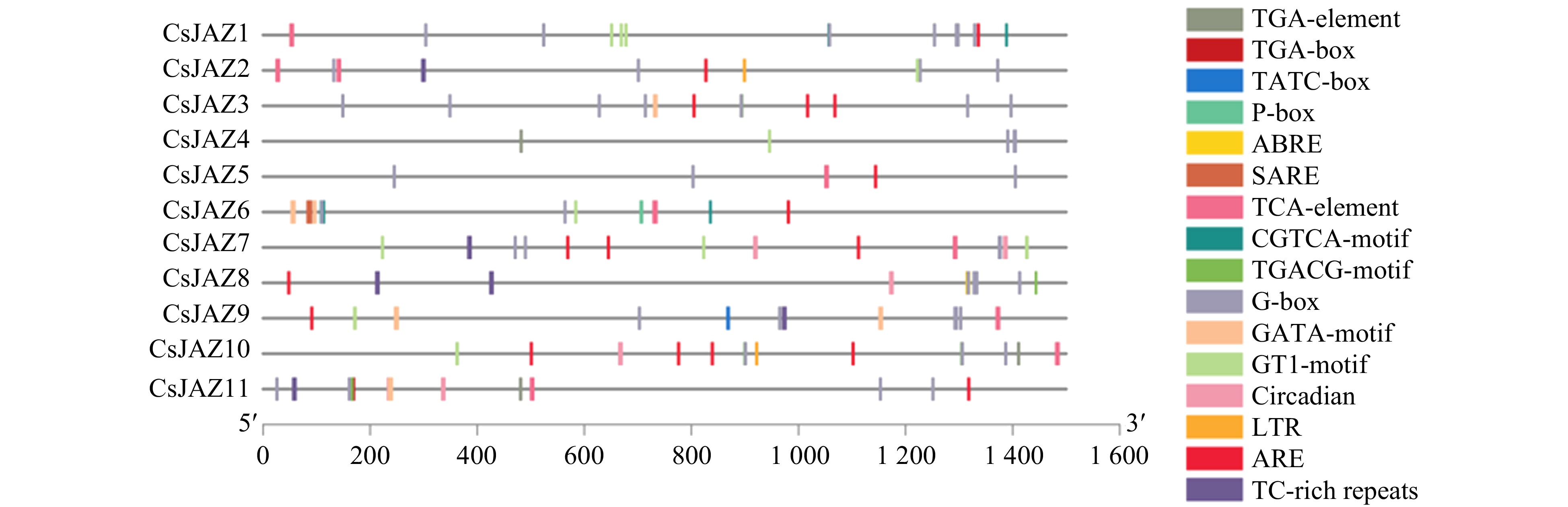
Figure 2.
Prediction of cis-acting elements in CsJAZ promoters associated with plant hormones and stress are shown. The distribution of cis-acting elements in the 1.5 Kbp upstream promoter region of the CsJAZ genes associated with plant hormones, grow and stress are illustrated.
Morphological changes in responses to waterlogging and waterlogging+MeJA
-
Morphological observation of the ARs developed over hypocotyl at 3, 5, and 7 d post waterlogging stress were carried out. Relatively similar phenotypes of control check (CK), Waterlogging (WL), and Waterlogging+MeJA (WL+MeJA) were observed at 3 d. The appearance of ARs on hypocotyl of WL stressed plants was clear at 5 d, whereas no ARs were found on CK and WL+MeJA treated hypocotyl. After 7 d of waterlogging stress, WL treatment produced a large number of ARs, and the number of ARs was higher than that of CK and MeJA treated plants (Fig. 3).

Figure 3.
Morphological changes and statistical collation in response to waterlogging and waterlogging+MeJA. (a) The response of WL and WL+MeJA after 3, 5, 7 d waterlogging treatment. (b) The survival rate of WL and WL+MeJA after 7 d of waterlogging treatment. (c) Comparison of the adventitious root (AR) number capacity of WL and WL+MeJA after 7 d of waterlogging treatment. ** p < 0.01.
The graphical representation of the observed phenotypes revealed that the survival rate of CK treatment was 100%. In contrast, only 50% of the WL+MeJA treated plants survived. Interestingly, the survival rate of WL treated plants was about 64%. The number of ARs in WL treatment was significantly higher than that in MeJA treatment.
Expression of CsJAZ genes in response to waterlogging treatment
-
Following the bioinformatic analysis of CsJAZ genes, we performed quantitative PCR analysis using the cDNA of waterlogging stressed hypocotyl organs of the cucumber plant. The expression of the CsJAZ was investigated for different stress periods, 6, 12, 48 and 96 h respectively (Fig. 4). We found that between the 11 CsJAZ genes, the majority of the CsJAZ genes displayed down-regulated expression trends under waterlogging stress. Among them, CsJAZ1, CsJAZ2, CsJAZ3, CsJAZ4, CsJAZ8, CsJAZ9, CsJAZ10, and CsJAZ11 decreased under waterlogging stress. Similarly, in addition to CsJAZ4, the expression of other CsJAZ were suppressed under waterlogging stress and also in the control (CK), but increased markedly under MeJA application. On the other hand, the transcript level of CsJAZ5, CsJAZ6 and CsJAZ7 induced significantly under waterlogging stress and waterlogging stress plus MeJA application. The mRNA level of CsJAZ1 rose to a maximum of 0.2 at 6 h under MeJA but fell again at 12, 24, 48, and 96 h in CK and also under waterlogging stress plus MeJA application. Concurrently, the expression level of CsJAZ1, CsJAZ2, and CsJAZ3 decreased at most of the timepoints in CK and waterlogging treated samples but contrastingly increased under MeJA treatment and peaked to a maximum of 0.17, 0.004 and 0.07 fold respectively. Therefore, we speculate that the JAZ genes, important regulators of the jasmonic acid signaling pathway, are involved in cucumber response to waterlogging stress.
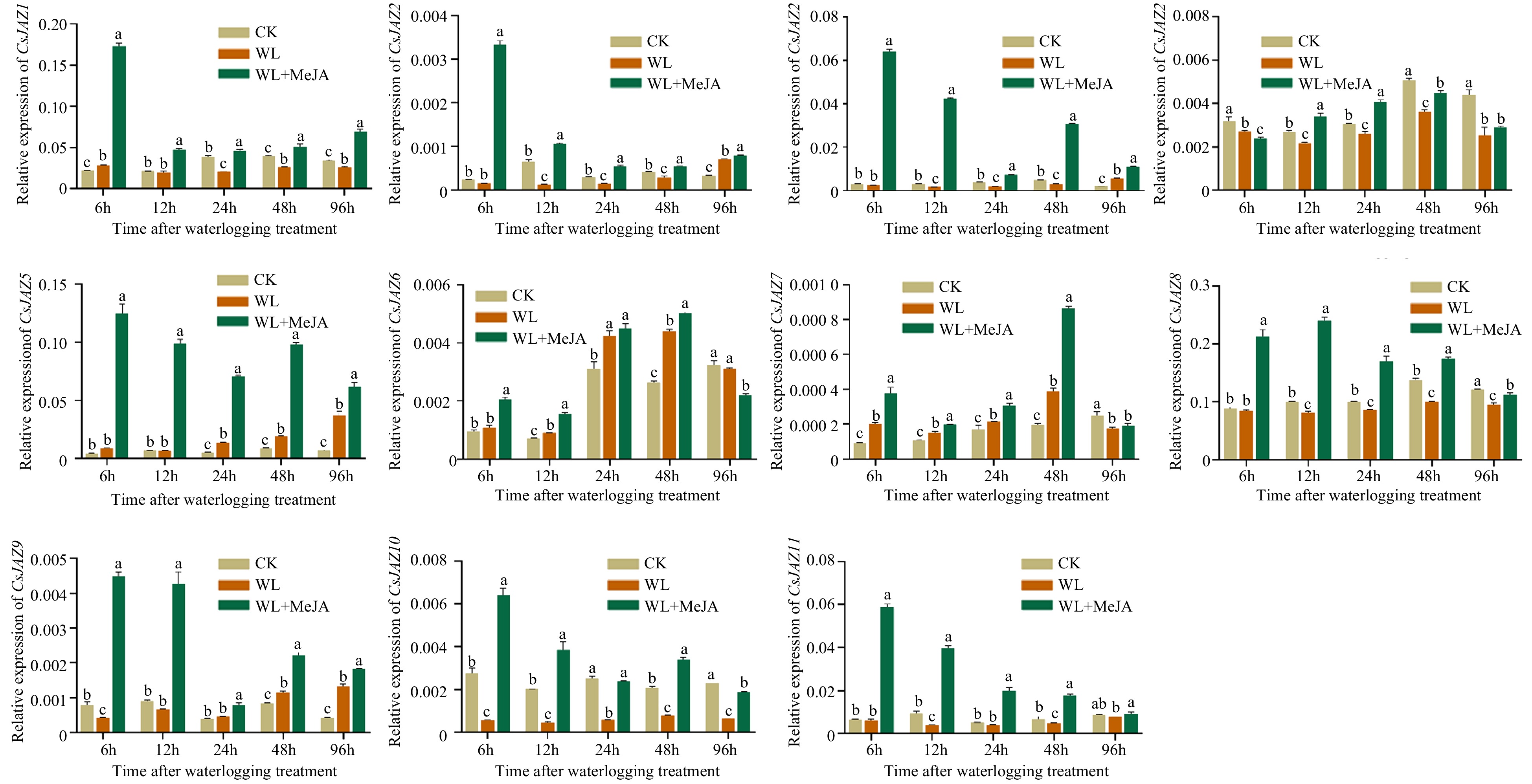
Figure 4.
Expression of CsJAZ genes in response to waterlogging treatment. The different lowercase letters above the bars indicate the significance (p < 0.05) of the relative expression level between two samples. The data for quantitative RT-PCR analysis were the mean and standard deviation of three bio-duplicates in the hypocotyl and three technical duplicates in each biological sample. The Y-axis is the relative level of expression.
Transcriptional expression patterns of CsJAZ in different tissues and post exogenous GA treatment
-
The RPKM values of CsJAZ in different tissues and post exogenous GA treatment were downloaded from the cucumber database (
http://cucurbitgenomics.org ) (Supplemental Table S6). Generally, all the CsJAZ genes were expressed in leaf, stem, ovary, tendril, fruit, root, male flower, and female flower (Fig. 5). Moreover, six genes (CsJAZ1, CsJAZ2, CsJAZ7, CsJAZ8, CsJAZ9, and CsJAZ11) were highly expressed in male flowers and three genes (CsJAZ3, CsJAZ5, and CsJAZ10) were highly expressed in fruit. CsJAZ1 was highly expressed in both leaf and stem. The CsJAZ4 and CsJAZ6 displayed dominant expression patterns especially in ovary tissue. Interestingly, further analysis revealed a correlation between tissue expression and evolution of the CsJAZ gene. For example, the CsJAZ5 and CsJAZ11 genes in group I, the CsJAZ2 and CsJAZ7genes in group III are closely related to the phylogeny of each homologous gene pair, but the tissue expression patterns are not consistent. These results indicate that CsJAZ genes have different expression patterns and tissue specificity. Moreover, the 11 CsJAZ genes were divided into three distinct groups. According to cluster analysis, eight organ tissues were also clustered into three distinct groups on the basis of their specific expression patterns.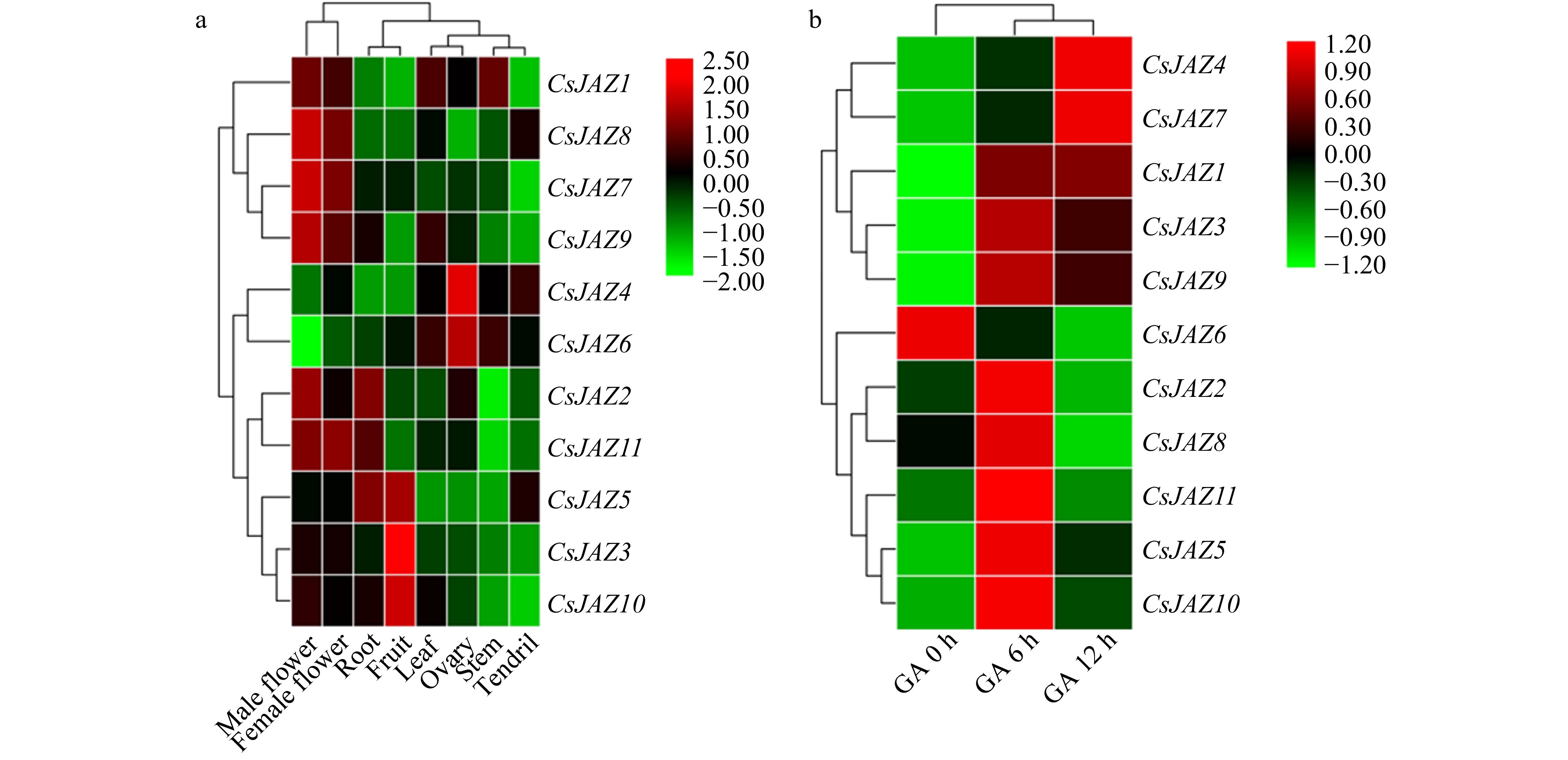
Figure 5.
Heatmap representation of CsJAZ genes in different tissue and post exogenous GA treatment of cucumber. The log2 transformation method was used to normalize and convert the RPKM values and displayed on the colored scale bar.
In response to GA treatment, all the CsJAZ genes were expressed following post exogenous GA application. In detail, the majority of CsJAZ genes (except CsJAZ6) were strongly upregulated after 6 h GA treatment. Among them, CsJAZ2, 5, 8, 10, 11 subsequently decreased at 12 h. On the other hand, CsJAZ1, 3, 9 were upregulated at 6 and 12 h after exogenous GA treatment. Noticeably, CsJAZ6 was significantly decreased by GA treatment from 6 h to 12 h. These results indicate that all CsJAZ genes respond to the plant hormone GA, and they may participate in complex hormone signaling networks.
Subcellular localization of CsJAZ proteins
-
In order to determine the subcellular localization of CsJAZ protein in cucumber cells, we cloned the CDS sequences of CsJAZ8 with specific primer, and finally generated a CsJAZ-pCAMBIA1301 construct. The construct encodes the CsJAZ protein fused with GFP. The CsJAZ-GFP fusion driven by the 35S promoter was introduced into the Tobacco epidermis. The GFP signal is detected in the nucleus of the transformed cell under a laser confocal microscope (Fig. 6), which is consistent with the prediction of Cell-PLoc 2.0 (
http://www.csbio.sjtu.edu.cn/bioinf/plant-multi ). Genes residing in the nucleus are generally considered as functional genes and could be involved in the regulation of various biological and cellular processes[33,34].
Figure 6.
Subcellular localization analysis of CsJAZ8-GFP fusion protein transiently expressed in Nicotiana tobaccum L. (a) Schematic representation of constructs used for subcellular localization of the CsJAZ8 protein. (b) The photographs were taken using green fluorescence (GFP), visible light, and merged light. Bar = 50 μm.
Homo- and heterodimeric interaction of CsJAZ proteins
-
In Arabidopsis, some JAZ proteins exhibit homologous and heterologous interactions mediated by the ZIM domain[14]. For this purpose, we used a yeast two-hybrid (Y2H) assay to determine whether these proteins interact to form homodimers or heterodimers. Due to the high degree of similarity in the nucleotide sequence of homologous genes, we selected 8 CsJAZ genes which were suppressed under waterlogging stress (CsJAZ1, CsJAZ2, CsJAZ3, CsJAZ4, CsJAZ8, CsJAZ9, CsJAZ10, and CsJAZ11), and cloned them. Through the toxicity self-activation test, we found that CsJAZ1 and CsJAZ3 have a self-activation phenomenon and cannot be mutually verified. Finally, 66 protein combinations were tested (Fig. 7). As shown in Fig. 7, the cucumber CsJAZ protein has homologous and heterologous interactions. The experimental results show that three of the six proteins (CsJAZ8, CsJAZ9, and CsJAZ10) exhibit homodimeric interactions, among which CsJAZ8 and CsJAZ9 exhibit strong homologous interactions, while CsJAZ10 homodimers interact with other members and displayed weak interaction capacity. As baits, CsJAZ4, CsJAZ8 and CsJAZ9 can interact strongly with other JAZ protein members. Meanwhile, the prey subordinate protein CsJAZ9 can interact with JAZ2, 4, 8, 9, 11, but not with CsJAZ10. On the other hand, as bait, CsJAZ9 can also interact with JAZ2, 4, 8, 9, 11, however, not interact with CsJAZ10. CsJAZ8 showed interaction with CsJAZ4, 8, and 9, both as prey and bait. Similarly, CsJAZ4 interacted with CsJAZ8 and 9 both as prey and bait. Therefore, we speculate that the cucumber JAZ gene may work in corresponding order to respond to waterlogging stress and perhaps other environmental stimuli through intra-family interaction.

Figure 7.
Yeast two-hybrid analysis of Homo- and heteromeric interactions in CsJAZ proteins. The full-length CsJAZ was fused with pGADT7 and pGBKT7 to generate prey and bait vectors, respectively, and then transformed into the yeast strain AH109. The interactions were determined by growth on SD-Trp/-Leu/-His/-Ade nutritional selection medium. The mating of pGBKT7-53/pGADT7-T7 was used as a positive control, and pGBKT7-Lam/pGADT7-T7 was used as a negative control.
-
The JAZ proteins belong to a plant-specific family known as TIFY[35]. To date, JAZ family proteins have been investigated in numerous plant species such as Arabidopsis thaliana[14,17,36], Oryza sativa[27], Glycine soja[29], Vitis vinifera[28], Nicotiana attenuata[37], Zea mays[38], Saccharum spp.[39], Melampsora larici-populina[30], Lycoris Aurea[40], Camellia Sinensis[31], Brassica juncea var. tumida[41]. However, there is currently no study available on the comprehensive understanding of JAZ proteins in cucumber. In this report, we retrieved the protein sequences of CsJAZ genes from the cucumber genome database. After that, we performed extensive bioinformatic analysis to unravel their evolutionary distribution and potential functions in the cucumber genome.
JAZ protein plays an important role in plant response to biotic and abiotic stresses as a key transcriptional inhibitor of plant jasmonic acid signaling pathway[42−44]. Analysis of CsJAZ promoters revealed a large number of hormones, growth and stress-related elements, but the types and numbers of elements contained in the different genes are not the same, indicating that the type and degree of response to hormone by cucumber CsJAZ gene is different, leading to functional differences between genes. In addition, some genes also contain multiple stress responsive cis-elements at the same time. It is speculated that CsJAZ may be involved in the regulation of different hormone signals to respond to environmental stress. In view of the presence of abiotic stresses response and hormone response cis-acting regulatory elements in the CsJAZ promoter region (Fig. 2), suggesting their potential involvement in regulating multiple abiotic stresses and could be vital in enhancing the resistance of cucumber against waterlogging stress.
JA deficient mutants opr3/dde1 and dde2-2 produced more ARs than the wild type[45,46]. Gutierrez et al.[47] showed that the accumulation of JA in Arabidopsis gh3 mutants was twice that of the wild type. This higher accumulation of JA resulted in the inhibition of AR. Another study highlighted the negative role of JA in the development of ARs. The arresting of the growth of ARs was observed in the Pepino (waterlogging sensitive line) and was found to be due to the increase in endogenous JA content. In contrast, the Zaoer N (resistant line) displayed reduced JA accumulation activity[48]. Similarly in the current study, the application of MeJA suppressed the ARs growth under waterlogging stress. The mechanism of how JA inhibits the growth of adventitious roots under waterlogging stress however needs further study.
We found that the JAZ gene, an important regulator of the jasmonic acid signaling pathway, was involved in cucumber response to waterlogging stress. Under waterlogging stress, the relative expression levels of all CsJAZ genes were up-regulated or down-regulated (Fig. 4). CsJAZ4 and CsJAZ8 genes were down-regulated at different time points after waterlogging stress. Therefore, we hypothesized that CsJAZ8 could be a potential candidate in regulating the mechanism of cucumber to waterlogging stress. In line with that, previous studies have shown that JAZ proteins are crucial in enhancing the defense mechanism of different plant species against various adverse conditions. For example, overexpression of GsJAZ2 enhances the resistance of soybean against alkali stress by regulating the intracellular pH homeostasis, maintaining the osmotic balance and expression of stress specific genes[49]. Overexpression of Apple MdJAZ2 also improved the tolerance of transgenic Arabidopsis to NaCl treatment[50]. There was no significant difference in the phenotype of cotton GhJAZ2 RNAi seedlings under salt stress, which may possess redundant functions with other family members in GhJAZ2 RNAi strains, but overexpression of GhJAZ2 increased the sensitivity of transgenic cotton to salt stress[51]. These studies showed that JAZ protein is widely involved in the study of salt and alkaline tolerance of plants. In addition, the JAZ protein plays a crucial role in plant resistance to biotic stress. Transgenic wheat lines that overexpressed truncated TaJAZ1 without Jas motif, accumulated more reactive oxygen species, which enhanced resistance to powdery mildew[52]. To date, no studies have explored the role of CsJAZ in cucumber waterlogging stress. Therefore, the specific role of the CsJAZ gene in cucumber needs further study.
In addition, exogenous MeJA treatment under waterlogging stress can significantly induce the expression of the JAZ genes. For example, the sugarcane ScJAZ1-ScJAZ7 gene increased sharply and peaked at 3 h after MeJA treatment before gradually down-regulating at 6 h to 12 h following treatment[39]. Similarly, most Camellia Sinensis JAZ genes were also significantly induced by MeJA treatment, and showed induced expression at 4 and 12 hours[31]. Delay in the response of LaJAZ5 genes to MeJA was observed, whereas LaJAZ6 was unaffected by MeJA[40]. In our study, the expression of CsJAZ genes was largely affected by both the MeJA treatment and waterlogging stress. We observed that six of the 11 CsJAZ genes (CsJAZ1, 2, 3, 5, 9, 10 and11) were up-regulated and reached their peak value at 6 h post MeJA treatment, showing consistency with previously reported studies[31] (Fig. 4).
Gene expression patterns are often related to gene function. Previous studies have shown that JAZ genes are differentially and constitutively expressed in plants. TaJAZ1, -4, -10, -11, and -14 respectively were expressed in the root, stem, leaf, stamen, and pistil tissues of wheat[53]. In sugarcane, seven ScJAZ genes were dominantly expressed in root, leaf, bud, stem pith, stem epidermis, and most of them were abundant in the bud and leaf tissues[39]. In H. brasiliensis, most HbJAZ genes such as HbJAZ1, HbJAZ2, HbJAZ7, HbJAZ8, HbJAZ9, HbJAZ10, and HbJAZ11 were expressed at higher levels in leaves than in bark[54]. Similarly in our study, the expression activities of 11 CsJAZ genes in various tissues of cucumber were observed (Fig. 5). These different expression patterns suggested that the 11 CsJAZ genes play a crucial role in the developmental processes of cucumber plants. For example, CsJAZ3, CsJAZ5, and CsJAZ10 displayed high expression levels in the fruit, and CsJAZ1, CsJAZ2, CsJAZ4, CsJAZ6, CsJAZ7, CsJAZ8, CsJAZ9, and CsJAZ11 had low levels in the same tissues. Only CsJAZ1 is highly expressed in leaves and stems, suggesting that CsJAZ1 may play an important role in sensing jasmonic acid signals in these structures. Collectively, the tissue-specific expression of CsJAZ genes indicates their possible involvement in cucumber growth and developmental activities.
A complex signal regulation network composed of a variety of plant hormone crosstalk is ubiquitous in plant growth and development[55,56]. The Jas domain at the C-terminal of the JAZ protein is very conserved, which interacts with ethylene (ETH) and other TFs, making the JA signaling pathway cross-linked with the ethylene signaling pathway in regulating plant development[57]. The weakly conserved NT domain at the N-terminal of the JAZ protein can interact with DELLA protein, and the main role of DELLAs is to inhibit gibberellin (GA) signal transmission. The interaction between the two results in the JA and GA signal pathway being linked in regulating the plant growth defense balance[58]. For instance, the majority of the TaJAZ genes in Triticum aestivum[53] and tea plant JAZ genes[31] respond to the plant hormone GA, the relative expression levels of these genes were upregulated after GA treatment. In our study, all the CsJAZ genes were changed significantly when post exogenous GA, and showed their responsive nature. Interestingly, not each of these responding genes contained the cis-acting elements involved in gibberellin-responsiveness like P-box or TATC-box. The promoter sequences of CsJAZ6 and 9 contained the GA responsive elements, indicating that CsJAZ6 and 9 were directly involved in GA signaling pathway of cucumber. CsJAZ1, 2, 3, 4, 5, 7, 8, 10, and 11 may respond to GA treatment by other unknown regulative pathways. These results indicated that CsJAZ genes may participate, directly or indirectly, in complex hormone signaling networks.
The subcellular localization experiment is generally performed to understand the behavioral nature of a certain gene. The JAZ protein of Arabidopsis[59] and Camellia Sinensis[31] plays the role of a nuclear protein, whereas the JAZ6 of sugarcane is a protein located in the cytoplasm and cell membrane[39]. OsJAZ1 was located in the cytoplasm, but it acted as a nuclear protein in the presence of the JA signal[60]. In addition, there may also be differences in subcellular localization of JAZ in the same species. For example, LaJAZ3, LaJAZ4 LaJAZ6, and LaJAZ7 of Lycoris aurea were located in the nucleus, LaJAZ2 and LaJAZ5 were located in the cytoplasm, while LaJAZ1 exists in the nucleus and cytoplasm[40]. Our study found that GFP-labeled CsJAZ8 is specifically localized to the nucleus (Fig. 6), which highlighted its involvement in the activation or suppression of transcription factors and determines the fate of the targeted proteins.
A TIFY/ZIM domain mediates homologous and heterologous interactions between JAZ proteins and also mediates interactions with NINJA, independent of plant hormones[14,19,61]. In previous studies, we found that all CsJAZ proteins contained a conservative TIFY motif in the ZIM domain. The yeast two-hybrid system proved that there were indeed homologous or heterologous dimer interactions between cucumber CsJAZ proteins (Fig. 7). Three of the six CsJAZ proteins (CsJAZ8, CsJAZ9, and CsJAZ10) exhibited homodimeric interactions. CsJAZ4 and CsJAZ8 can interact extensively with other JAZ protein members as baits. Homologous or heterologous interactions between JAZ proteins were also verified in Lycoris aurea[40] and Gossypium hirsutum[51]. Therefore, we hypothesized that cucumber JAZ protein may jointly respond to waterlogging stress through synergistic interaction among family members. However, the detailed network of cucumber JAZ protein regulating waterlogging stress interactions requires further verification to determine the specific role of the JAZ transcription factor in this regulation under waterlogging stress.
-
In conclusion, our study is the first to integrate genome-wide identification of the JAZ genes in cucumber. Eleven CsJAZ genes in the cucumber genome were identified and characterized. The following analysis, phylogenetic relationships, chromosomal location, gene duplication, selective evolutionary pressure, conserved domains, gene structure analysis, cis-acting regulatory elements of the JAZ family members, revealed the distinct roles of CsJAZ genes. Moreover, a quantitative PCR analysis of 11 CsJAZ genes revealed that these genes may be crucial in regulating the response of cucumber to waterlogging. Transcriptional expression patterns of CsJAZ in different tissues and under exogenous GA treatment were visualized. The subcellular localization experiment revealed that GFP-labeled CsJAZ8 is specifically localized to the nucleus. Further, cucumber CsJAZ proteins exhibit homologous and heterologous interactions. Our study explored research on the CsJAZ family in cucumber, and provides insightful knowledge regarding the role of CsJAZ proteins in waterlogging stress and could be useful in future cucumber breeding programs.
-
The whole-genome data of cucumber was downloaded from the Cucurbit Genomics Database (CuGenDB,
http://cucurbitgenomics.org ), the plant Jasmonate ZIM-domain protein sequence and cucumber protein data were also downloaded through NCBI (https://www.ncbi.nlm.nih.gov ) for blastall comparison. The protein sequence was obtained from Pfam (http://pfam.xfam.org )[62]. The NCBI Conserved Domain Database (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ) and the SMART program (http://smart.embl-heidelberg.de ) were used to further identify the JAZ protein structure and protein conserved domains and to remove redundancy. Finally, a total of 11 non-redundant CsJAZ genes were identified in the cucumber genome named CsJAZ1~CsJAZ11. ExPASy (https://web.expasy.org/protparam ) and PSORT tool (https://www.genscript.com/psort.html )[63,64] were used to calculate protein sequence length, molecular weight, and isoelectric point (pIs). The subcellular localization of all JAZ proteins of cucumber in different organisms were predicted using the online tool Cell-PLoc-2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2 )Phylogenetics, gene structure, motif composition analysis
-
In order to study the relationship between the JAZ gene phylogeny of cucumber, 12 Arabidopsis thaliana, 11 Vitis vinifera, 15 Oryza sativa, 13 Solanum lycopersicum, and 12 Populus trichocarpa JAZ protein sequences and 11 CsJAZ of cucumber were used. The amino acid sequences of conserved Jas domain were used for phylogenetic tree construction. Firstly, we performed ClustalW multiple sequence alignment of the JAZ protein sequences of six species, and set all parameters to default values[65], we then used the Neighbor-joining method[66], set the bootstrap replicates value to 1000 for phylogenetic tree construction using MEGA X software. According to the branches of the phylogenetic tree, it was divided into three subgroups, Group I, Group II, and Group III.
The cucumber whole-genome sequence was aligned with mRNA sequences to confirm the CsJAZ gene structures. The conserved motifs in the JAZ protein of cucumber are identified using the Motif Elicitation (MEME) online prediction tool (
http://meme-suite.org/index.html )[67], and the maximum number of motifs was set to 10, other parameters were set as default. In order to better understand the structure of the CsJAZ genes of cucumber, the Gene Structure Display Server program (GSDS) (http://gsds.gao-lab.org ) was used to map the whole genome sequence and CDS sequence of the CsJAZ genes and included the CsJAZ map of gene structure, and the exon and intron structure of CsJAZ genes.Chromosomal location, gene duplication analysis, and selective evolutionary pressure analysis
-
The distribution of the cucumber JAZ protein on the particular chromosome was visualized using MapChart software[68], according to the documents of genome annotation available on the website of the cucumber genome.
The Multiple Collinearity Scan toolkit (MCScanX) with default parameters was used to examine the gene duplication events of CsJAZ genes[69]. KaKs_Calculator2.0 software was used to perform non-synonymous (Ka)/synonymous (Ks) analysis on the members of the cucumber CsJAZ gene family (previously described by Wang et al., 2010[69]) to further analyze segmentally and tandemly duplicated CsJAZ genes. Ka/Ks ratio < 1 means purifying selection, ratio = 1 means neutral evolution, and ratio > 1 means positive selection[70].
Identification of cis-elements of cucumber JAZ genes
-
The 1500 bp upstream genomic DNA sequences of 11 CsJAZ genes were extracted and the cis-regulatory elements were analyzed using the Plant CARE website (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html )[71].Plant growth and waterlogging treatments
-
The waterlogging tolerant cucumber line Zaoer-N was used as the test material. On August 18, 2019, it was sown in a cave containing vermiculite : peat : perlite (1:3:1 v/v/v). The seedlings were grown in a glass greenhouse, 14 h at 28 °C/10 h at 18 °C (day/night) photocycle, and the relative humidity was 75%. Selecting Zaoer-N seedlings with the same two leaves and one heart (seedling age is about 15−20 d) for waterlogging treatment (waterlogging to the growth place of the first cotyledon), the seedlings were transferred into plastic containers filled with water[72]. After the preliminary exploration of different concentrations of MeJA treatment, a total of three treatments were set up, namely non-flooded treatment (CK), ordinary waterlogging treatment (WL), and waterlogging plus 500 μmol/L MeJA. The cucumber hypocotyls were collected at 6, 12, 24, 48, 96 h after treatment. Under all conditions including the control, three biological replicates were collected for each sample, which consisted of nine hypocotyls of the same line. All samples were immediately frozen in liquid nitrogen and kept at −80 °C to extract total RNA.
Total RNA Extraction, qRT–PCR analysis and transcriptomic analysis of CsJAZ in different tissues
-
The extract of cucumber hypocotyl total RNA was carried out using the TaKaRa MiniBEST Plant RNA Extraction Kit (TaKaRa, China), from 200 mg of hypocotyl samples, Nanodrop One (Thermo Fisher Scientific, USA) and 1% agarose gel electrophoresis was used to detect concentration and RNA completeness. 1 μg RNA was used in the first strand cDNA synthesis using a 20 μl reaction volume using the PrimeScript™ RT reagent Kit with gDNA Eraser (Perfect Real Time) (TaKaRa, China). Quantitative RT-PCR was performed using SYBR® Premix Ex Taq™ II (TaKaRa, China). The qRT-PCR reaction conditions were as follows: 50 °C for 2 min, 95 °C for 10 min, 40 cycles of 95 °C for 15 s, and 60 °C for 1 min. The primers used for the qRT-PCR assay of JAZ genes are listed in Supplemental Table S7. For each sample, the internal reference gene (ACTIN) was used to normalize the abundance of mRNA, and three replicates were performed. The relative expression level of genes was calculated using the 2−ΔΔCT method[73], and SPSS 16.0 software (IBM Corporation, USA) data was used for analysis. The different lowercase letters above bars indicated the significance (p < 0.05) of the relative expression level between two samples.
For measuring the relative amount of corresponding transcripts of CsJAZ genes in different plant tissues and post exogenous GA treatment, the transcriptomic data of CsJAZ genes were obtained from the cucumber genome database and visualized using TBtools[74].
Subcellular localization analysis
-
The coding region of the CsJAZ8 gene was amplified by PCR with specific primers CsJAZ8-GFP-F and CsJAZ8-GFP-R (Supplemental Table S7). The PCR product was ligated to the BamHI-KpnI sites of GFP fusion expression vector pCAMBIA1301-GFP to generate the transient expression vector pCAMBIA1301-GFP-CsJAZ8 by ClonExpress II One Step Cloning Kit (Vazyme Biotech, China). Next, the CsJAZ8-GFP fusion driven by 35S promoter was transformed into Agrobacterium tumefaciens GV3101 strain competent cells[75]. The cells were collected by centrifugation and suspended in a buffer solution containing 10 mM MES (pH 5.5), 10 mM MgCl2 and 200 μM acetylcholine, with a 600 nm optical density of 1.0. Using the tobacco transient transformation method, the cells were injected into one month old Ben's tobacco leaves with a needleless syringe. After 48−72 h, green fluorescence was detected in the tobacco epidermal cells under a laser confocal microscope (ZEISS LSM 980 with Airyscan2, Germany).
Yeast two-hybrid (Y2H) assay
-
Yeast two-hybrid system (Clontech, USA) based on Matchmaker gal4 was used. The pGBKT7 vector contains the GAL4 DNA binding domain, and the pGADT7 vector contains the GAL4 activation domain. The CDS sequence of the CsJAZ gene was amplified by PCR with gene-specific primers (Supplemental Table S7), and the full-length cDNA of CsJAZ was cloned into pGBKT7 (bait vector) and pGADT7 (prey vector). The inactive BD-CsJAZ2/CsJAZ4/CsJAZ8/CsJAZ9/CsJAZ10/CsJAZ11 and AD-CsJAZ1/CsJAZ2/CsJAZ3/CsJAZ4/CsJAZ5/CsJAZ6/CsJAZ7/CsJAZ8/CsJAZ9/CsJAZ10/CsJAZ11 plasmids were mixed and co-transformed into the yeast strain AH109. Yeast cells were cultured on a medium lacking leucine and tryptophan (SD/-Leu/-Trp) at 28 °C for 3−4 d. Then, yeast colonies were screened on a medium lacking leucine, tryptophan, histidine, and adenine (SD/-Leu/-Trp/-His/-Ade) at 28 °C for 3 d to determine possible interaction.
Statistical analysis
-
The statistical significance was performed using SPSS software (version 23.0, SPSS Inc., USA), using statistical analysis ANOVA and 95% and 99% confidence intervals (p ≤ 0.05 and p ≤ 0.01). The analyzed data were expressed as means ± standard deviation (SD) of three replicates for all measured parameters. GraphPad Prism 8.0 (GraphPad Software, Inc., LA Jolla, CA, USA) was used to produce the graphs in accordance with the data.
This research was supported by the National Natural Science Foundation of China (grant nos. 32030093 and 31801883) and Natural Science Foundation of Jiangsu Province (BK20180913).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Synteny analysis of JAZ genes in cucumber .
- Supplemental Table S2 Synonymous and nonsynonymous substitution rates for the duplication events in cucumber JAZ genes.
- Supplemental Table S3 GenBank accession numbers of reference species in evolutionary analysis.
- Supplemental Table S4 The description of cis-acting elements of the CsJAZ genes.
- Supplemental Table S5 The description of cis-acting elements of the CsJAZ genes associated with plant hormones and stress.
- Supplemental Table S6 RPKM values of CsJAZ genes in different tissues and post exogenous GA treatment.
- Supplemental Table S7 List of primers used in this study.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Pan J, Tu J, Sharif R, Qi X, Xu X, et al. 2021. Study of JASMONATE ZIM-Domain gene family to waterlogging stress in Cucumis sativus L.. Vegetable Research 1: 3 doi: 10.48130/VR-2021-0003 

Study of JASMONATE ZIM-Domain gene family to waterlogging stress in Cucumis sativus L.
- Received: 21 December 2020
- Accepted: 15 April 2021
- Published online: 29 April 2021
Abstract: Jasmonate ZIM Domain proteins (JAZs) provide a response to multiple abiotic stresses by critically regulating the JA signaling pathway. In this study, a total of 11 CsJAZ genes were identified in the cucumber genome database. In silico analysis revealed the presence of different cis-acting regulatory elements related to growth, stress and hormonal responsiveness in the upstream region of CsJAZ genes. The transcriptional expression pattern analysis showed that all CsJAZ genes were expressed in the tissues tested and all CsJAZ genes responded to plant hormone GA, suggesting they may participate in complex hormone signaling networks. An expression analysis based on qRT-PCR, indicated that cucumber CsJAZ8 was decreased significantly by waterlogging treatment but induced by exogenous methyl jasmonate application. Therefore, we speculate that CsJAZ8 could be a potential regulator of the response of cucumber to waterlogging stress. Additionally, transient expression assay showed that the CsJAZ8 protein is localized in the nucleus and could be involved in the regulation of various biological and cellular processes. Furthermore, Y2H assay revealed that cucumber CsJAZ proteins exhibit homologous and heterologous interactions. Subsequently, this research will contribute to the understanding of the CsJAZ gene family in Cucumis sativus. It also provides further insight into the role of CsJAZ proteins in waterlogging stress and may provide a new frontier in cucumber breeding for waterlogging tolerance.
-
Key words:
- Cucumis sativus L. /
- JAZ protein /
- Expression profiles /
- Y2H /
- Waterlogging stress