








-
Agricultural biostimulants are generally defined as formulations of compounds, substances or microorganisms that are applied to plants or soils to improve vigor, yields, quality and tolerance to stresses[1]. Exactly how these biostimulants function in the promotion of plant growth and health remains a mystery.
Endophytes are generally soil or seed-vectored microbes (often fungi and bacteria) that plants absorb into, or otherwise cultivate within their tissues, resulting in benefits to the host. These include increased nutrient absorption, improved development, increased abiotic and biotic stress tolerance, improved hardiness and sometimes increased resistance to pathogens or herbivores[2]. Another general feature of endophytes is that they inhabit plant tissues without causing outward symptoms or obvious expressions of their presence[2].
A Buddhist text that dates from the first millennium BCE describes the parable of the blind men and the elephant. In the parable, blind men were led to different sides of an elephant, then asked to describe the elephant by feeling it—but none could describe the entire elephant[3]. Investigators in two separate fields dealing with endophytic microbes and crop stimulation may be compared to the blind men of this Buddhist parable. Plant endophytism and crop biostimulation are two closely related and interacting phenomena that are not easily separated. In this article we will evaluate several biostimulants commonly used in agriculture and show how they function, either by becoming endophytic themselves or by stimulating absorption of soil microbes, thereby increasing endophytic activity.
-
It is difficult to determine how early in the domestication of crops humans began to employ microbial or other biostimulants. One likely historical use of microbial biostimulants is the 'corn medicine' used by Native American Indians. An example is the 'Iroquois Corn Medicine'[4]. It was the practice of the Iroquois to enhance corn growth by a method that may have used microbes from roots of non-cultivated grasses to provide microbes for corn that improves growth and nutrition of corn plants. It was recorded in the book 'Iroquois Foods and Food Preparation'[4] that the Iroquois would gather roots of common reed grass (Phragmites australis), bottle brush (Hystrix patula), Elymus canadiensis, and other common woodland grasses. Roots would be rinsed and placed in warm water, probably to aid in the removal or exiting of bacterial endophytes from roots (and root hairs) into the water. In some cases, roots were boiled—and this likely resulted in activation of Bacillus endospores in older parts of the roots. The Native Americans would then soak and partially germinate corn seeds in the liquid derived from wild grass roots. After inoculation with root bacteria, the seeds were planted in corn mounds. It is difficult to envision how the Native Americans developed their biostimulant methods, but it is likely that corn growth and health were considerably improved beyond the growth of corn without the corn medicine. It may also be considered that the method for inoculation with biostimulant microbes used by the Native Americans, and wherever farmers of different cultures used similar methods, was likely superior to that used in modern agriculture in terms of microbial diversity. In modern methods we tend to select a defined group of soil or plant microbes to use as biostimulants—while the Native Americans obtained a larger undefined community of biostimulant microbes from a selection of wild grasses, and this increased their chances of obtaining a selection (community) of microbes that were already adapted to grasses. In essence, the Native Americans transferred the entire root microbiome from wild grasses to cultivated corn to better cultivate their corn crops.
-
Plants in nature transfer microbes to their seeds that facilitate their growth, these microbes colonize seedling roots and stimulate seedling root growth and root hair elongation and protect seedlings from disease[5]. The microbial communities on seeds frequently require a microbial community maturation period as seeds are exposed to soil microbes and the elements (repeated periods of high moisture, lower moisture, fluctuations in temperature, etc.)[2]. Seed-vectored microbes may be lost from seeds due to how we manage seeds in domesticated plants without conditions in which the seed microbiome may mature[2]. Santhanam et al.[6] showed that the process of harvesting seeds from the field and storing them under dry refrigeration conditions was enough to result in the loss of protective microbes. These authors began annual cultivation of a wild tobacco species by harvesting seeds, storing them under cool dry conditions, then planting the following spring. After seven years of cultivation a wilt disease epidemic spread in the plants under cultivation. An examination of wild plants and cultivated plants showed that several microbes had been lost from the cultivated plants during the years of cultivation. The investigators then re-inoculated seeds with the microbes that had been lost and the wilt disease disappeared. The microbes were likely lost from seeds of the wild tobacco because the natural process of seed ripening involves a seed microbiome maturation period in which seeds mature on plant inflorescences or on the ground, where they are exposed to moisture and microbes. This maturation period permits the seed microbiome to increase in community diversity and density. Removal of seeds from plants and storage in dry protected conditions truncates the seed microbiome maturation phase and leaves seeds with an undeveloped microbiome and at a disadvantage without the microbes it needs for growth and survival.
This seed microbiome maturation phenomenon was shown to be important in the production of grass seeds[7]. Fescue and ryegrass seeds require a period where they are exposed to sufficient moisture to develop a diverse and dense seed microbiome on the surface of the seeds. Chen et al.[7] showed that seeds produced in very dry climates failed to develop a diverse microbe community on the seeds and as a consequence, had reduced germination rates or showed increased disease susceptibility along with reduced stress tolerance. Cotton seeds are generally treated with concentrated acids to degrade the cotton fibers so that seeds will flow better in planter machines[8]. However, as microbes are carried on the hairs of the seeds, removal of the hairs by the acid results in loss of the seed microbiome and leaves seeds at a disadvantage without their growth stimulatory and disease protective microbiome[8]. Similarly, some of our modern hybrid corn passes through an aseptic tissue culture phase that removes much of the microbiome and leads to plants that must recruit new microbes. However, because many of these microbes may be missing or reduced in soils managed with minimal organic material, or where soils are sterilized, reacquisition of beneficial microbes from unhealthy soils is difficult. Other seeds such as Bermuda grass (Cynodon dactylon) are covered with hulls that carry microbes[2]. These hulls are removed to increase seed germination rates, and this results in loss of the seed microbiome, as a result seeds must be coated with fungicides and other chemicals to facilitate their growth. It seems likely that many seeds in our cultivated crops have lost some microbes over the thousands of years that we humans have been cultivating them. This phenomenon may be described as a 'crop endophyte paradox' in that domestication, selection for enhanced productivity and continuous long-term cultivation of plants, without consideration of the presence of endophytes or their selection, frequently results in plants with fewer endophytes and less capability to survive naturaly without human intervention. Thus, many or all of our cultivated plant species could benefit from addition of biostimulant microbes to improve growth and reduce diseases.
-
Seed-vectored and soil microbes (mostly bacteria) function in the rhizophagy cycle[2, 9, 10]. Plants use soil microbes to transport soil nutrients (nitrogen, phosphorus, zinc, etc.) to themselves[2, 10]. Initially, microbes are attracted to the root tip by secretion of exudates (sugars, organic acids and other nutrients) by the root tip cells. The root cells around the root tip absorb the microbe cells into root cells, locating them in the periplasmic space (beneath the root cell wall and the root cell plasma membrane). The root cells then secrete superoxide onto bacteria, oxidizing away the bacterial cell walls and causing them to become naked protoplasts, and extracting nutrients from bacteria. Bacterial protoplasts are replicated within root cells by the action of root cell cyclosis where bacteria are circulated around the periphery of root cells, resulting in frequent replication of bacterial protoplasts, essentially cloning the bacterial protoplasts within root cells[10]. The presence of bacterial protoplasts within root cells triggers root hair elongation; without bacteria no root hairs are formed[2, 5, 9, 10]. As root hairs elongate, the bacteria are periodically ejected from small pores that form in the tips of the root hairs. Bacteria trigger hair elongation through production of ethylene and this triggers ejections of bacteria from the tips of hairs with each hair growth spurt[10]. After ejection from the hair tips, microbes grow in size, and reform cell walls and flagella, becoming competent again to acquire nutrients from the soil[9, 10]. The presence of rhizophagy microbes in plant roots results in increased exudate secretion from roots to cultivate microbes at the root tips; and further root hairs secrete exudates to rebuild bacteria after ejection from the root hairs. Thus, the rhizophagy cycle results in increased microbial activity both around and within roots. Because there are increased exudates in the rhizosphere, rhizophagy microbes and others in the soil may engage in 'nutrient mining', absorbing some nutrients and solubilizing other nutrients (e.g., potassium, calcium, magnesium, etc.) that the plant may absorb from the soil solution.
-
Biostimulant microbes may stimulate plant growth or health in multiple ways, including as epiphytes or endophytes, through functioning in the rhizophagy cycle or antagonists to pathogens. Commercial products may contain microbes that have biostimulant effects on plants, although there is some uncertainty as to how these biostimulant products actually function to promote plant growth or health[11]. Some of these commercial microbes were initially derived from soils, and others from fermented plant materials. Products may contain a selection of several endospore-forming bacteria belonging to genus Bacillus or related genera[2, 11]. Endospore formers are frequently used because they may be stable in formulations, where the dormant endospores are used in commercial products. However, other commercial products use freeze-dried mixes of bacteria like Pseudomonas spp. that do not form endospores[11]. In this case, the freeze-drying of the bacteria stabilizes them in commercial products. Other products can contain a large, sometimes incompletely defined, community of microbes from plant fermentation sources. These microbes may be freeze-dried or otherwise stabilized in the commercial product or less-optimally, they may continue to grow in the commercial product prior to use on plants. In general Bacillus spp. from any source or species may be endophytic (Fig. 1−3) in plants (entering root cells at root meristems) but the individual microbes may differ in growth promotional capabilities[1, 2, 5, 7, 9, 11]. The typical growth promotional effects on plants include increased root branching, increased root exudation, increased root hair growth, increased nutrient delivery (phosphate, nitrogen, potassium or micronutrients), increased plant growth in general and increased oxidative stress tolerance[2, 11].
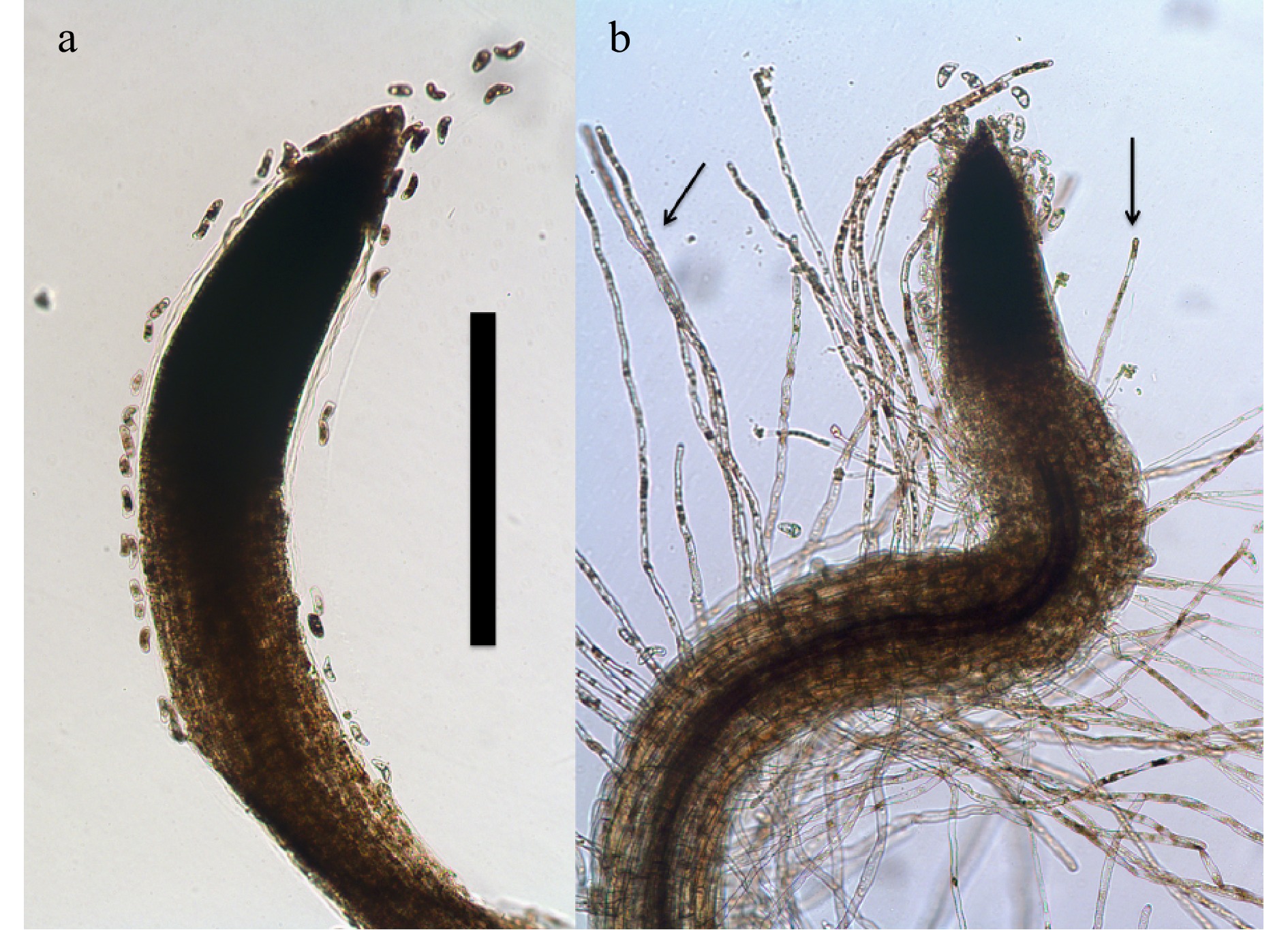
Figure 1. Root tips of 7-day-old Bermuda grass (Cynodon dactylon) seedlings growing on 0.7% agarose (stained with diaminobenzidine tetrahydrochloride; bar = 1 mm). (a) Root tip lacking root hairs due to absence of bacteria. (b) Root tip of seedling inoculated with a commercial product strain of Bacillus showing abundant presence of root hairs (arrows).

Figure 2. Root epidermal cells of wheat (Tritichum aestivum) seedling showing bacteria (arrows) within the root cell (stained with diaminobenzidine tetrahydrochloride; bar = 10 µm).
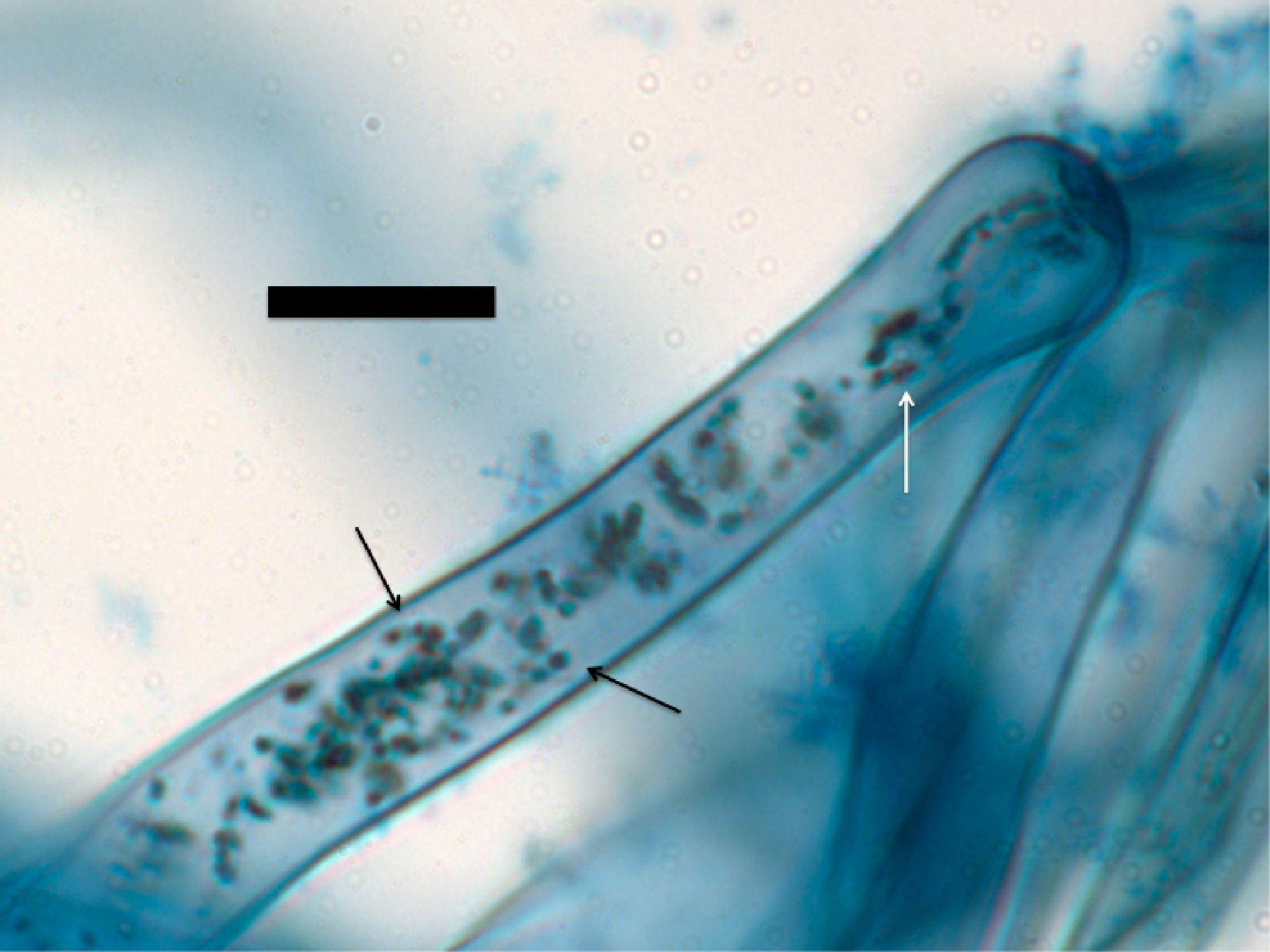
Figure 3. Root hair tip of Bermuda grass (Cynodon dactylon) seedling inoculated with commercial product strain of Bacillus sp. showing abundant presence of bacteria (arrows) within root hairs (stained with diaminobenzidine tetrahydrochloride; bar = 10 µm).
In some cases, soil isolated strains of Bacillus may be as good or better than some commercial product strains[11]. This suggests that some products with mixtures of microbes could be improved by specific formulation of mixtures of microbes each with a greater number of beneficial effects on seedlings. Some strains of bacteria may actually reduce plant growth through the phenomenon of 'endobiome interference' where microbes become endophytic and reduce plant growth rather than stimulate it[2].
-
Some plants host nitrogen-fixing microbes in their root nodules (e.g., rhizobia and actinorhizal symbioses) that are adapted to facilitate continuous nitrogenase activity and fixation of nitrogen from the atmosphere into forms that may be utilised. These well-evolved nitrogen-transfer symbioses are only present in certain families of plants (e.g., Fabaceae, Myricaceae, Rosaceae, Cucurbitaceae). However, diazotrophic endophytes are commonly distributed in many plant tissues in all families of plants[12]. Over several decades, there has been an effort to develop products from bacteria that may function in plants to provide fixed nitrogen to plants and reduce the requirement for nitrogen application to crops. The fixation by diazotrophic bacteria within, or in association with, plant tissues has come to be identified as 'associative nitrogen fixation'. Efforts to find and use diazotrophic endophytes in crop plants include those being made by Sharon Doty and collaborators at the University of Washington where bacteria isolated from poplar (Populus trichocarpa) and willow (Sitka sitchensis) are being developed into commercial products to stimulate crop growth[13]. A well-known diazotrophic endophyte is a strain of Glucoacetobacter diazotrophicus, initially isolated from sugarcane (Saccharum officinarum) where it was observed that sugarcane had been growing for decades without nitrogen applications[14]. There is considerable evidence that this diazotroph colonizes plant root tissues intracellularly, in a similar way to microbes that enter into the rhizophagy cycle[14].
-
Attempts are being made to achieve nitrogen fixation in cereals using endophytes that fix nitrogen. These efforts are exemplified by the work of Drs. Edward Cocking and David Dent[14]. In this work, the nitrogen-fixing bacterium Glucoacetobacter diazotrophicus, an endophyte of sugarcane, is being used in a range of crops, including cereals[14]. In sugarcane, as much as 40% of the nitrogen may come from endophytic bacteria like Glucoacetobacter diazotrophicus[15]. In sugarcane it has been found that nitrogen fixation occurs within or on the surface of roots[16]. It is thought that high oxygen levels suppress nitrogen fixation in leaves and stems. It has been suggested that formation of a pellicle (gelatinous mass of bacterial cells) by Glucobacter diazotrophicus in plant tissues excludes enough oxygen to permit nitrogen fixation in grasses[16]. Intracellular single cells or pellicles of this bacterium have been suggested to act as nitrogen fixing organelles called 'diazoplasts' may leak nitrogen into plant tissues[16−18].
Chang, Kingsley and White[10] outlined evidence for another mechanism, termed 'nutrient trap mechanism', for nitrogen fixation and transfer from intracellular diazotrophic bacteria. This mechanism is a nutrient trap, because once microbes are engaged in nutrient exchange with a plant cell, the microbes cannot escape from the process without being degraded or expelled by the plant cell. In this mechanism, intracellular bacteria secrete ethylene that triggers the host root cell (root hair) to grow and supply intracellular bacteria with exudates (carbohydrates that fuel nitrogenase activity); simultaneously, the root cell produces superoxide triggering bacteria to secrete antioxidant nitrogen (nitric oxide or ammonia) that combines with superoxide to produce nitrate that may be absorbed directly into root cells[10]. Nitric oxide secretion is thought to result in the chemical reaction NO (nitric oxide) + 2O (superoxide) → NO3 (nitrate); while secretion of ammonia would result in the chemical reaction 2NH3 (ammonia) + 9O (superoxide) → 2NO3 (nitrate) + 3H2O (water). It is notable that secretion of ammonia by bacterial cells would result in a nearly five-fold antioxidant capacity and twice the nitrate yield. Though not confirmed, secretion of ammonia is likely with diazotrophic bacteria where ammonia is the end product of nitrogen fixation[16]. Through exposure to superoxide produced by the plant cell, diazotrophic bacteria are forced to secrete antioxidant forms of nitrogen to prevent their own degradation. In the nutrient trap mechanism in root hairs, bacteria are subjected to alternating periods of high and low superoxide, with nitrogen fixation and bacterial replication in periods of low superoxide exposure and nitrogen extraction in periods of high superoxide exposure[10]. The function of nitrogen fixation in the nutrient trap mechanism in root hairs is closely tied to root cell growth where microbial cells are provided with nutrients to support replication and nitrogen fixation. Variations of the nutrient trap mechanism could also occur in leaves, where photosynthesis could provide photosynthate to fuel nitrogenase activity in cells, with bacteria being exposed to alternating periods of low and high superoxide levels. It is thus possible that many plants that are adapted to grow under low soil nitrogen conditions may use the nutrient trap mechanism to acquire nitrogen directly from diazotrophic bacteria growing within their cells[10]. Further research could evaluate this possibility.
-
One strategy for increasing plant supply of nitrogen from biological nitrogen fixation is to use endophytes that have been enhanced for nitrogen fixation by genetic modification[18]. This is a strategy that may be employed by commercial or university scientists [19]. Endophytic microbes (Methylobacterium spp.) that do not fix nitrogen could be employed as vehicles of nitrogen-fixation genes [19]. Most plants harbor Methylobacterium endophytes throughout all tissues of the plant[20]. Genes for nitrogen fixation from bacteria that normally fix nitrogen could be inserted into Methylobacterium endophytes. The nitrogen fixing Methylobacterium endophytes could then be used to inoculate plants, including cereals.
-
Some companies have developed, or are developing, multi-species microbe mixes intended to enhance plant growth, stress tolerance, disease resistance and nitrogen use efficiency [21]. Companies may screen for growth promotional microbes and obtain licenses of promising growth promotional microbes from university investigators—then test microbes to develop products for crops[21]. It is another unique approach to use fungal endophytes as carriers of endophytic bacteria. Fungi readily colonize seed surfaces and can hold or stabilize bacteria until seeds germinate and the microbes colonize seedlings[8]. Some products may constitute mixes of microbes that replace the lost seed microbiome and promote plant growth, disease resistance and nutrient use efficiency of seedlings[21].
-
Humic substances are naturally occurring molecules in soils that originate from microbial degradation of plants, primarily lignin from xylem[22]. Fossil deposits of humic substances are also available in large deposits in the earth in the form of the mineral Leonardite that is mined for various applications for agriculture and human health[23]. These substances are used extensively in agriculture as plant biostimulants[23]. They have been shown to do the following: 1) Improve plant mineral nutrition in terms of zinc, nitrogen, phosphorus, potassium, iron, magnesium, sulfur, manganese and copper; 2) Improve oxidative stress tolerance in plants, increasing resistance to abiotic stresses; 3) Improve root architecture, through increased root branching and root growth in general; 4) Increase plant exudation of nutrients from roots[24]; and 5) Increase microbial activity in the rhizosphere around roots[24]. There are two competing ideas as to how humic substances benefit plants. The first widespread hypothesis is the 'Improved Soil Properties Mechanism' (ISPM)[23]. According to this idea, humic substances have binding sites that function to increase soil cation exchange capacity, resulting in increased holding capacity for nutrients in soils and ultimately the conveyance of nutrients to plants. The ISPM focuses purely on the chemical properties of soil and humic substances[22, 23]. An alternate hypothesis may be referred to as the 'Signal Molecule Mechanism' (SMM)[25]. The SMM hypothesis emphasizes the direct 'hormone-like' effects that humic substances have on plant expression of genes and activities[25, 26]. In regard to the SMM hypothesis, it has been proposed that root cell plasma membrane receptors sense humic substances to activate ATPase and nitrate transporters, among other genes[26]. It is likely that both mechanisms contribute to the beneficial effects on plants.
Experiments conducted by James White and Ivy Chang focused on the effects of humic substances (Leonardite) on the rhizophagy cycle in plants. In those experiments it was found that humic substances in concentrations as low as 0.01% stimulated roots to increase internalization of bacteria into root cells at root tips. Exposure to humic substances resulted in increases in root gravitropic responses (i.e., more roots growing vertically, greater root hair elongation and root length growth (Table 1)).
Table 1. Humic acid effects on grass seedling (Poa annua) root gravitropic response, root hair and root length growth1.
Humic acid concentration
(wt./vol.)Gravitropic response2 Root hair length (µm)3
(n = 18)Root length
(n = 12)0% 41% (n = 177) 197 ± 10.4 9 ± 1.3 mm 0.01% 88.5% (n = 113) 537 ± 50 19 ± 2.1 mm 0.1% 89.5% (n = 143) N/A 39 ± 3.1 mm 1 Humic acid (mined and powdered Leonardite from EarthWorks Health, LLC (Norfolk, Nebraska, USA)) was incorporated into 0.7% agarose media. Seeds were then germinated and grown on the agarose for 7 days in lab ambient conditions, then seedling parameters were assessed.
2 Gravitropic response % = number of seedling roots that grow down into agarose ÷ total number of seedling roots × 100.
3 Data are presented as mean ± SD.Humic acids appear to serve as signal molecules for plants; they signal to plants, that microbes are active in degrading plant materials. Plants detect humic substances and increase the absorption of microbes into root cells at the root tip meristems. This absorption is evident because root hairs increase in length in response to humic substances. Previous research has shown that root hair length is directly proportional to the number of bacteria in root cells[2, 10]. Entry of microbes into cells at root tip meristems appears to be regulated by active secretion of reactive oxygen (hydrogen peroxide) from root tip cells. In media without humic substances, root tips secrete abundant reactive oxygen (Fig. 4a); when humic substances are incorporated into media around roots, reactive oxygen secretion from the root tip stops and roots cultivate microbes around the root (Fig. 4b), and microbes are internalized (absorbed) into root cells. Reactive oxygen secretion at root tips serves as a valve or switch for increasing rhizophagy cycle activity. Thus, humic substances appear to regulate entry of microbes into root cells and tissues.
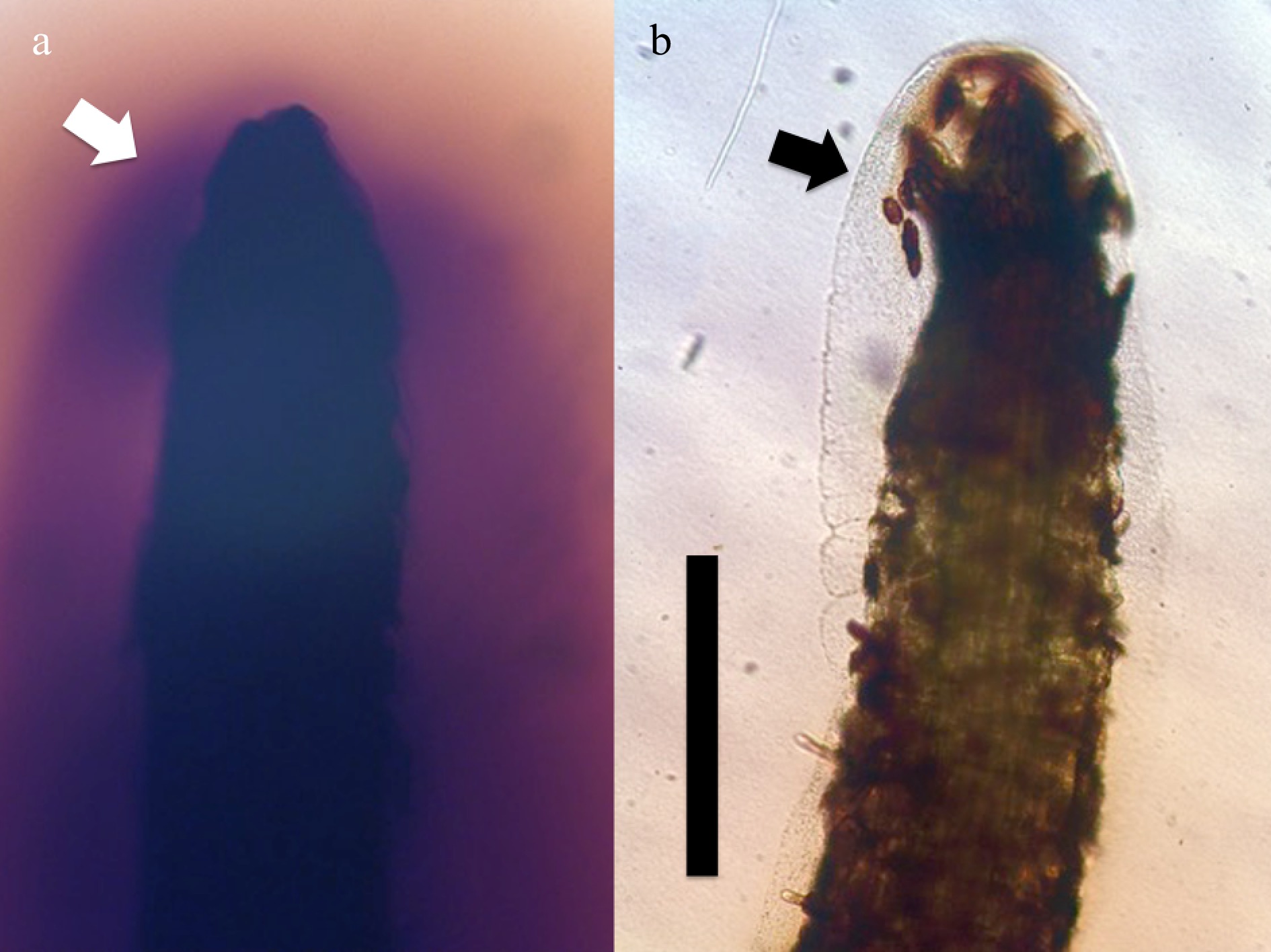
Figure 4. Root tips of grass Poa annua seedlings stained for ten hours at 20 oC with hydrogen peroxide stain diaminobenzidine tetrahydrochloride (bar = 1 mm). (a) Root tip (arrow) in agarose without humic substances, showing secretion of hydrogen peroxide (brown color) from root tip and absence of bacterial masses. (b) Root tip of Poa annua from the same experiment showing lack of reactive oxygen around root tip where humic substance (0.01% wt./vol.) is present in medium; bacteria (arrow) are also evident around the root tip.
-
In recent years, nutrient coatings have been used to stimulate seed germination and seedling growth[27]. Coatings range from plant extracts, algae, and proteins of various origins[27]. These nutrient coatings have been documented to increase nutrients available to plants and increase seedling resistance to stress and disease[27]. Soy flour coatings on broccoli (Brassica oleracea) seeds increased nitrogen content only slightly in seedlings, but markedly increased growth and chlorophyll content[28]. It was concluded by the authors that soy flour stimulated growth of plants through biostimulation rather than through nitrogen fertilization effects[28]. In studies using protein hydrolysates to coat seeds, it has been shown that coatings stimulate seed microbiomes and increase growth parameters as well as improve resistance to diseases and stresses[29]. All of the effects seen in these nutrient coatings on seeds are consistent with stimulation of the rhizophagy cycle in seedlings. The addition of nutrients to seed surfaces provides nutrients that increase growth of microbes that are carried on seed surfaces[2]. The growth of seed surface microbes increases microbes available for entrance into the rhizophagy cycle[9]. Many benefits exhibited in seedlings stem directly from rhizophagy cycle activity[9]. This includes increases in nutrients (nitrogen, phosphorus and zinc) and increased oxidative stress tolerance[2].
-
Soil contamination in agriculture is a frequent problem. Contaminants may include heavy metals, salts or other biological poisons[30]. There is an increasing recognition that endophytic biostimulant microbes may enhance plant growth to the extent that survival, growth and in many instances bio-extraction or degradation of the toxins is possible[30]. Arsenic (As) is a toxic metalloid according to the Environmental Protection Agencies of Europe and the United States[30]. Exposure to As can cause liver, endocrine, cardiovascular and kidney diseases[30]. It is also well established that As is a Class I carcinogen, and manifests in many types of cancers[31]. Arsenic is widely spread both by natural routes (geysers, volcanoes, natural soil composition) and anthropogenic routes (urban runoff, domestic and commercial activities, traffic and the mining industry, are a few examples.)[30]. Currently, mining is one of the most significant means for As contamination in soils due to the drifts generated once mines are abandoned[31]. Presence of As in terrestrial and aquatic ecosystems is causing biomagnification in upper trophic levels with high environmental and public health impacts[32]. Despite toxicity, some plants, bacteria and fungi are tolerant to this metalloid and other heavy metals[33], being able to accumulate them in tissues to high concentrations (hyperaccumulators), or they metabolize them to less toxic or volatile forms or prevent entry (excluders). Rice, one of the staples of much of the human population, is a hyperaccumulating plant and therefore a potentially dangerous food if As accumulates in the grain. The risk posed by rice depends both on the amount of rice consumed, and on the concentration of inorganic As in rice grains, moderated by the absorption rate in the intestine as well as in the dietary source[34, 35].
Some work acknowledges the role of endophytic bacteria in the metabolism and detoxification of As in plants, allowing their adaptation in stressful environments[36, 37]. Bacillus pumilus significantly reduces the amount of As in grains because of a high production of siderophores[38]. The same occurs with fungi and endophytic yeasts. Saccharomyces cerevisiae has been genetically engineered with the Waarsm gene, which encodes As methyltransferase. This gene comes from the soil fungus Westerdykella aurantiaca, which can remove As from contaminated soil. When this genetically modified strain is incorporated into the microbiome of rice plants, the concentration of As in grain is reduced and, in addition, this strain is a plant biostimulant bacterium[39]. The inoculation of a hyperaccumulator rhizobacterial strain, Ochrobactrum tritici As5 (a strain modified to increase its accumulative capacity), into the rhizosphere of rice plants reduced As presence inside the tissues of rice plants and reduced the inhibitory effect of the metalloid on the plant's growth parameters. Molina et al.[30] evaluated some other examples in which bacteria or fungi associated with plants altered the plant’s adaptive capacity against As stress.
Jasione montana L. is an As-tolerant plant that can grow in high concentrations of As[40, 41], although the mechanism of detoxification is uncertain. While some authors classify it as an As accumulator plant[42], others consider that it behaves as an As excluder plant[41]. Endophytic bacteria isolated from seeds, seedlings and mature plants were characterized according to Molina et al.[30]. Most plants were highly resistant to As, and bacteria isolated from them, functioned as plant biostimulant bacteria (Table 2). In order to evaluate possible application as microbe-based biostimulants against As stress, two bacteria, Pantoea eucalypti MC-12 from seeds and Pantoea conspicua MC-K1 from mature plants, and a mixture of all microbes were inoculated into Bermuda grass (Cynodon dactylon), as a laboratory model[30] (Fig. 5). This grass is very sensitive to 125 μM As concentration. The seeds (50 per plate, 5 plates per treatment) were placed on 0.7% agarose with 125 µM of As and incubated (or not) with single bacteria or with a bacterial mixture. When Pantoea eucalypti MC-12 was placed on seeds, the germination percentage (13.24% ± 1.7%) improved significantly with respect to the controls (non-inoculated seeds) (4.83% ± 1.72%)[30]. When seeds where inoculated with Pantoea conspicua (MC-K1) germination success was also superior to controls (12.74% ± 2.39%) (Fig. 5)[30]. This improvement was greater (19.43% ± 3.14%) when the seeds were inoculated with the bacterial mixture isolated from the seed and seedlings. These preliminary results show how seed-vectored endophytes and those from adult plants (vertical or horizontal transference) increase germination in situations of As stress. We propose that these endophytes, like others, may cause an adjustment in host plant tolerance to oxidative stresses, and this may increase tolerance to As, although more work is needed to confirm this mechanism for As stress tolerance. Further research could assess potential use of such microbes as biostimulant microorganisms under As stress conditions.
Table 2. Plant growth promotional features of Jasione Montana bacteria1.
Strain Species As MIC2 Phosphate solubilization3 Auxin
(µg/mL)% Inhibition Alternaria sp. (LB/PDA4) MC-12 Pantoea eucalypti 200 mM ++ 18.96 ± 8.79 0/0 MC-10 Bacillus siamensis 1 mM + 1.1 ± 0.12 100/82.9 MC-13 Pantoea sp. 200 mM + 2.3 ± 1.56 0/0 MC-14 Acinetobacter radioresistens 200 mM − 0.83 ± 0.03 25/0 MC-15 Unidentified 200 mM + 4.97 ± 0.36 20/43 MC-19 Unidentified 200 mM + 2.45 ± 0.29 0/0 MC-K1 Pantoea conspicua 450 mM + 1.96 ± 0.07 40/0 Bacterial mix N/A 200 mM ? 4.46 ± 0.01 20/65 1 Methodology and bacterial identifications in Molina et al.[30].
2 As MIC = As minimal inhibition concentration.
3 Bacteria were screened for phosphate solubilization by a plate assay method using Pikovskaya agar media: ++ Halo greater than a centimeter in diameter; + Halo less than a centimeter in diameter; − No halo[30].
4 Percent inhibition of radial growth = R1−R2/R1 × 100 where R1 is the farthest radial distance grown by the pathogen in the opposite direction of the antagonist and R2 is the distance grown on a line between the inoculation of the pathogen and the antagonist on media Luria Bertani agar (LB) and potato dextrose agar (PDA)[30].
Figure 5. Germination of Bermuda grass in 125 µM arsenate. (a) Control (no microbes). (b) Inoculated with Pantoea eucalypti MC-12. (c) Inoculated with Pantoea conspicua MC-K1. (d) Inoculated with the bacterial mixture isolated from the seeds and seedlings.
-
Diverse seed microbiomes containing a variety of microbes are important in growing and maintaining healthy, disease-free plants. However, the process of domestication and the application of modern agricultural practices have resulted in the elimination, or dramatic reduction, of the seed-vectored microbes of many crop plants. To combat this loss of microbe diversity, pre-modern cultures transferred microbes from wild plants to crop seeds. Today, some agricultural companies are developing, or have already developed, microbe-based biostimulants to enhance plant growth and development by taking naturally occurring or genetically modified microbes and introducing them to cultivated plants.
While all the details behind how microbes can affect plant growth and development have not been elucidated, many biostimulant microbes are known plant endophytes. Endophytic microbes participate in the rhizophagy cycle, bringing in soil nutrients that would have otherwise been unavailable to plants without endophytes. Not all endophytic microbes are beneficial, however certain species or genera may inhibit plant growth due to endobiome interference. More research could be done to determine how various endophytes interact with each other and their hosts so that more efficient and specialized biostimulant microbe combinations can be developed.
In addition to microbe-based biostimulants, humic-substance-based biostimulants are also being used to enhance plant growth and development. Humic substances appear to function as plant signaling molecules in the root and rhizosphere regions. Even low concentrations of humic acids increase the internalization of bacteria into root cells, thus acting as a regulator of the rhizophagy cycle and endophytism in plants. It may be possible that biostimulating effects of humic substances partially stem from their rhizophagy- and endophytism-enhancing properties.
The addition of proteins to seed surfaces provides organic nitrogen but also stimulates seed microbiomes during germination and increases the community of microbes that enter into the rhizophagy cycle and stimulate seedling growth and stress tolerance.
There is also evidence that microbes that increase plant tolerance to soil contaminants like As may also be endophytes, entering into tissues of plants and modifying plant response to contaminant stress.
We suggest that most of the beneficial effects of biostimulants stem from their participation in the rhizophagy cycle, either as endophytes being moved into the plant, chemical signalers that move the cycle along, or nutrients that foster the maturation of the microbiome. Further research could be done to illuminate the unknown aspects of the rhizophagy cycle and intracellular nitrogen fixation. In addition, it would be valuable to determine the mechanisms involved in how endophytic microbes affect their plant hosts, including effects on plant gene expression and secondary metabolite production in crops.
- Funding support was provided from USDA-NIFA Multistate Project W4147, the New Jersey Agricultural Experiment Station, and from a Cooperative Ecosystems Studies Unit CESU G16AC00433 grant between Rutgers University and the U.S. Geological Survey for control of invasive Phragmites australis, and Rey Juan Carlos University, grant DRÍADES-2020/00006/009. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
- The authors declare that they have no conflict of interest.
-
 This article has been contributed to by US Government employees and their work is in the public domain in the USA. It is an open access article under the terms of the Creative Commons Attribution License and not copyrightable (CC0), visithttps://creativecommons.org/share-your-work/public-domain/cc0.
This article has been contributed to by US Government employees and their work is in the public domain in the USA. It is an open access article under the terms of the Creative Commons Attribution License and not copyrightable (CC0), visithttps://creativecommons.org/share-your-work/public-domain/cc0.
-
About this article
Cite this article
White JF, Chang X, Kingsley KL, Zhang Q, Chiaranunt P, et al. 2021. Endophytic bacteria in grass crop growth promotion and biostimulation. Grass Research 1: 5 doi: 10.48130/GR-2021-0005

Endophytic bacteria in grass crop growth promotion and biostimulation
- Received: 15 March 2021
- Accepted: 02 June 2021
- Published online: 21 June 2021
Abstract: Plants naturally carry microbes on seeds and within seeds that may facilitate development and early survival of seedlings. Some crops have lost seed-vectored microbes in the process of domestication or during seed storage and seed treatment. Biostimulant microbes from wild plants were used by pre-modern cultures to re-acquire beneficial seed microbes. Today some companies have developed or are developing the use of microbes obtained from soils or plant sources to stimulate plant development and growth. Many of these biostimulant microbes are endophytic in plants. Biostimulant products also include humic substances, which appear to function as signal molecules in plants, triggering increased internalization of soil microbes into root cells and tissues. In addition, protein coatings on seeds fuel the growth of seed surface-vectored microbes, increasing microbial activity around and within roots. In this article, we provide evidence of the endophytic nature of many biostimulant microbes, and suggest that many of the beneficial effects of microbial biostimulants stem from their action as endophytes or as participants or stimulants of rhizophagy cycle activity.
-
Key words:
- Endophytes /
- Crop endophyte paradox /
- Nitrogen fixation /
- Rhizophagy cycle /
- Humic substances