










-
The aromatic substance in green tea is mainly composed of alcohols, aldehydes, ketones, esters, and olefins. The 'clean aroma' of green tea, is mainly comprosed of hexenal, hexenol, pentenol, hexenyl ester and 2-n-pentyl furan[1]. The main components of the 'fruit sweet fragrance' are linalool, linalool oxide, geraniol, nerol, farnesene, β-ionone, benzaldehyde, phenylacetaldehyde, (E)-2-octylenal, 3,5-octadien-2-one, α-cedarene, indole and jasmonolide, etc.[2]. In addition, some famous green teas have a 'chestnut fragrance', which is mainly due to terpenes and esters[3]. It has been reported that most of the aromatic substances in green tea have high aroma activity values, and a minor change in their content will lead to significant differences in people's perception of tea aroma[4]. The processing of tea has been proven to have an important impact on the aroma of green teas by regulating the synthesis or degradation of aromatic substances[5]. In the spring season, the post-harvest management of fresh leaves has become a serious problem for many tea manufacturers. Due to limited productivity, a large quantity of picked leaves cannot be processed to green teas in time, resulting in excessive water loss of picked leaves[6]. To tea manufacturers, how to preserve or even enhance the sensory quality of green tea products under the premise of prolonging the spreading period of picked leaves is an important issue which needs to be resolved. Enlightened by the preservation measures of fruits and vegetables after harvest[7], we developed three different low-temperature spreading methods based on conventional natural spreading, and performed a detailed analysis of the changes in the main chemical components of spreading leaves. In our previous work, we reported their effects on the metabolism of non-volatile components and the taste quality of green tea[8]. Here, we aim to reveal the influence mechanism of different spreading methods on the metabolism of aromatic substances and the aroma quality of green tea using headspace solid-phase microextraction (HS-SPME) combined with GC-MS and transcriptome analysis. This study will provide a genetic metabolic basis for applying low-temperature spreading technology in actual tea production.
-
This experiment was conducted in September. Natural spreading as CK treatment was conducted inside the tea factory with an indoor natural humidity of approximately 65 ± 2%. The air humidity would decrease slightly as the ambient temperature slightly increased. Therefore, the humidity parameter was set over 70% to minimize the humidity difference between CK and low-temperature treatments. Compared with environment humidity, temperature is the main factor affecting water-loss rate of postharvest plant crops[9].
The variation of water content in postharvest tea leaves under different spreading treatments has been published in our previous work[10]: with spreading time increasing, the moisture content in fresh tea leaves decreased gradually. While, the water-loss rate varied significantly among the leaves with different treatments. In CK, after spreading for 7 h, the moisture content decreased from 78.6 ± 0.7% to 72.1 ± 0.4%, with an average water-loss rate of 0.9%/h. In LTD and LTY, the spreading duration for the moisture content dropping to 72.2 ± 0.4% and 71.8 ± 0.3% was 14 h and 18 h, with an average water-loss rate of 0.46%/h and 0.38%/h, respectively. In LTCD, the water content decreased to 74.2 ± 0.2% after 48 h, with an average water-loss rate of 0.092%/h. This result indicated that moderately low-temperature (15 °C) conditions could significantly delay the dehydration of spreading leaves, and yellow-light irradiation or CO2 addition could further extend the spreading period.
The aroma quality in green tea products under different spreading treatments
-
The aroma quality scores of green tea products (Supplemental Table S1) were in the order of LTY (89.6) > LTCD (86.6) > CK (85.4) > LTD (84.9). The green teas with LTY treatment had the highest score due to a high fruity fragrance. The LTCD treatment had a lasting fragrance and scored slightly higher than CK and LTD. The results showed that low temperature spreading technology (especially yellow-light treatment) could improve the aroma quality in green tea and significantly extend the spreading period of fresh tea leaves after harvest.
The volatiles contents in green tea products under different spreading treatments
-
Supplemental Table S2 shows the relative content of aromatic compounds in green tea products. The relative amount of terpenes in green tea products was in the order of LTY (4.04 µg·g−1DW) > LTCD (3.00 µg·g−1DW) > LTD (2.50 µg·g−1DW) > CK (2.12 µg·g−1DW) (Fig. 1a) with a significant difference (p < 0.01) between low-temperature treatments (LTD, LTY, LTCD) and natural spreading treatment (CK). A variety of terpene alcohols were identified in green tea, including linalool, dehydrolinalool and four kinds of linalool oxides, geraniol, isogeraniol, tetrahydrogeraniol, neroliol, nerol, terpineol, farnesol, and verbena alcohol. The variation trend of the relative amount of terpene alcohols was consistent with the total amount of terpenes, with the highest amount in LTY, followed by LTCD and LTD (Fig. 1b). Some terpene olefins were also detected, such as butene oxide, β-ocimene, perillene, and β-farnesene, and their content was higher in LTY and LTCD than other treatments. The content of juniperene, cedarene, and butene had a slight difference among the four treatments. The amino acid-derived volatiles detected in dried teas were benzaldehyde, phenylacetaldehyde, phenethyl alcohol, methyl salicylate and indole, and their content under different treatments were in the following order: LTY > LTD > LTCD > CK (Fig. 1c). The fatty acid derived volatiles detected in green tea were octenal, hexenal, hexenol, and jasmone, which were significantly (p < 0.01) higher in LTD and CK than in LTY and LTCD (Fig. 1d). Terpenoids and amino acid-derived volatiles are conducive to the good aroma of green tea[11,12], monoterpenes and sesquiterpenes with rich sweet, floral and woody fragrance were reported to contribute most to the aroma of green tea[13]. However, some low-carbon fatty acid-derived volatiles have a negative effect on the aroma of green tea due to their crude green aroma[14]. The relative total amount of terpenes in dried tea samples was highest in LTY, followed by LTCD. The relative total amount of amino acid derivatives was also highest in LTY, followed by LTD, LTCD and CK (Supplemental Table S2). This result basically supported the sensory evaluation score of green tea products.
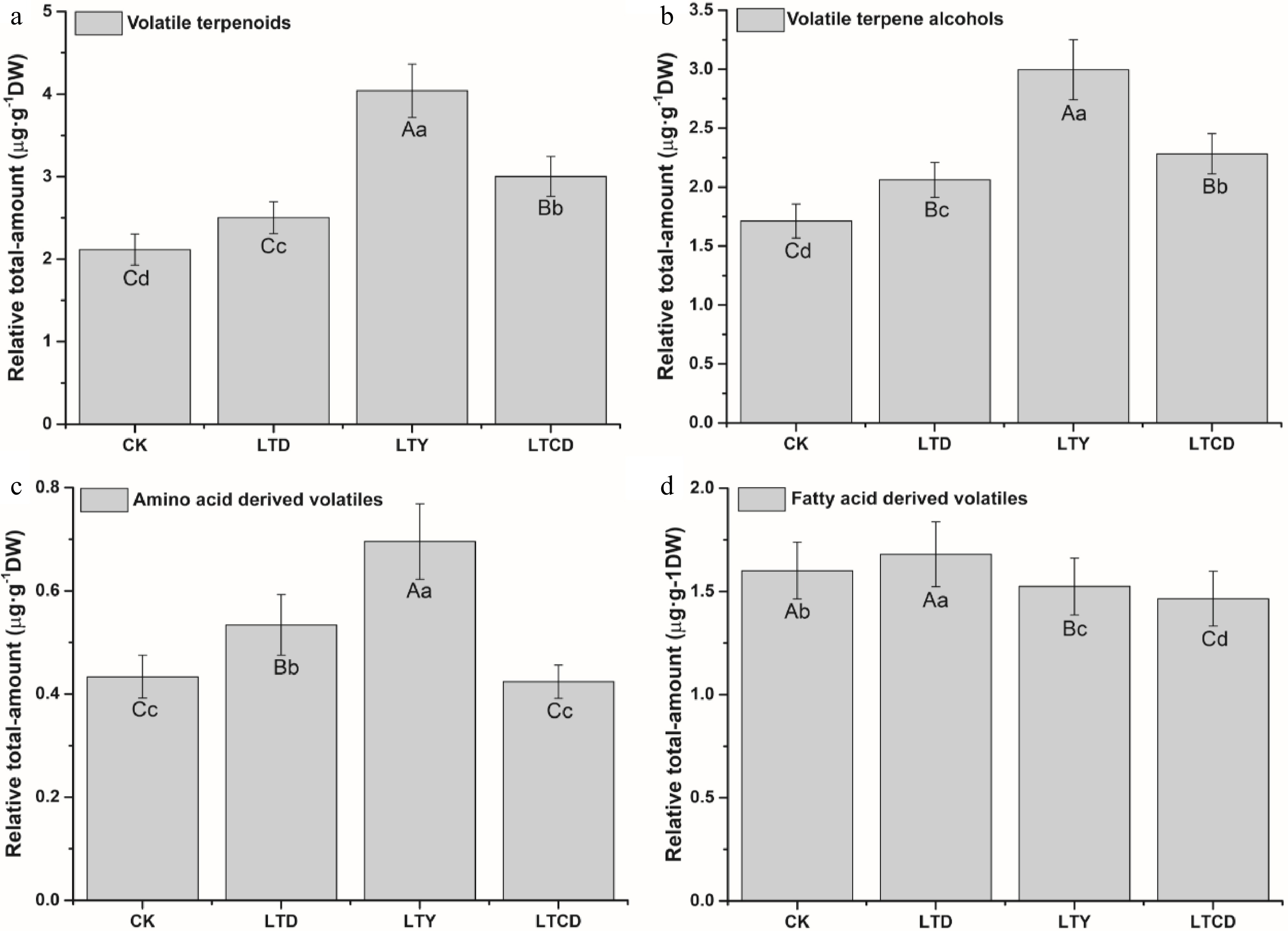
Figure 1. The relative total-content of volatile compounds in green tea products. (a) Volatile terpenoids; (b) volatile terpene alcohols; (c) amino acid-derived volatiles; (d) fatty acid-derived volatiles. Different upper/lowercase letters represent the significant difference at P < 0.01/0.05 level (ANOVA, Duncan test).
Differential volatile metabolites among four spreading leaf samples
-
Three main types of endogenous aroma volatiles in spreading tea leaves were detected, including terpenoids, fatty acid-derived volatiles and amino acid-derived volatiles, and a total of 45 compounds were identified (Supplemental Table S3). As shown in Fig. 2a, the relative content of terpenoids was significantly higher (p < 0.01) in samples with low temperature spreading than those with natural spreading, with the order of LTY > LTCD > LTD > CK > 0 h. Terpene alcohols were the major components of terpenoids, and the total-content variation (Fig. 2b) was basically consistent with that of terpenoids. Linalool, dehydrolinalool, linalool oxide, geraniol, nerolidol, nerol, terpineol, farnesol, cedarol and menthol were detected in tea leaves, and their content showed a significant increase (p < 0.05) after spreading, with the largest increase being observed in LTY.
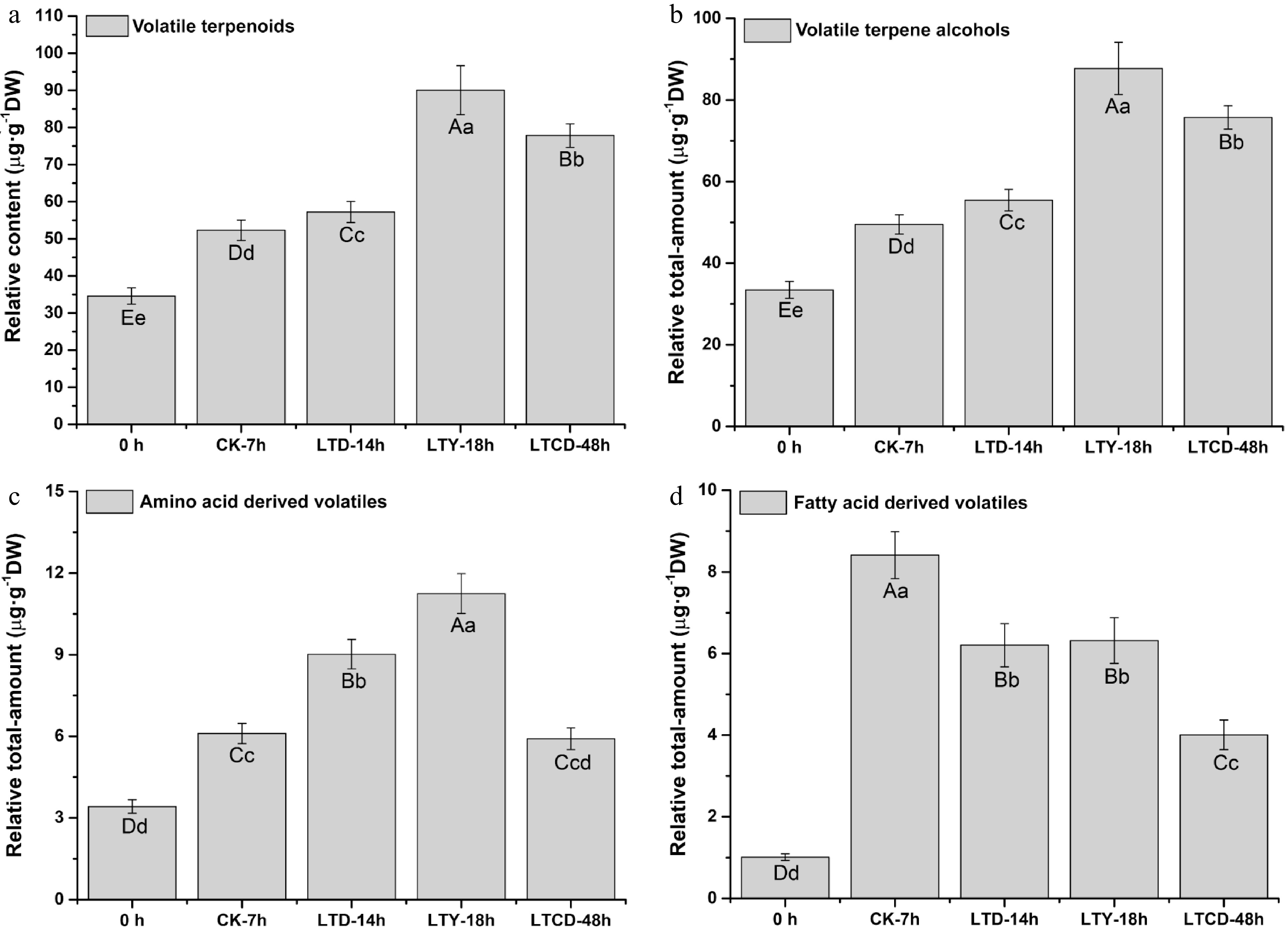
Figure 2. The relative total-content of volatile compounds in different spreading samples. (a) Volatile terpenoids; (b) volatile terpene alcohols; (c) amino acid-derived volatiles; (d) fatty acid-derived volatiles. Different upper/lowercase letters represent the significant difference at P < 0.01/0.05 level (ANOVA, Duncan test).
The amino acid-derived volatiles detected in spreading leaves were phenethyl alcohol, benzaldehyde, methyl salicylate, and indole. Benzaldehyde, phenethyl alcohol, and methyl salicylate belong to phenylalanine. The relative total-content of amino acid derivatives in spreading tea leaves were in the order of LTY > LTD > CK > LTCD > 0 h (Fig. 2c). Compared with benzaldehyde, phenethyl alcohol had a relatively higher content, and its content significantly increased after spreading at low temperature with 0 h (1.88 µg·g−1DW), LTD (2.59 µg·g−1DW), LTCD (3.20 µg·g−1DW), LTY (3.64 µg·g−1DW). Meanwhile, the largest increase in the content of methyl salicylate was also observed in the LTY treatment.
A number of fatty acid derivatives were also detected in the spreading leaves, including leaf alcohol, 2-hexenol, 2-ethylhexenol, heptanol, 3,7-dimethyloctenol, octenal, hexenyl butyrate, hexenyl hexanoate, hexenyl benzoate, and jasmone. The relative total amount of fatty acid derivatives could be lowered significantly by low temperature treatment (p < 0.05), and the order of their content was CK (8.41 µg·g−1DW) > LTY (6.32 µg·g−1DW) ≈ LTD (6.21 µg·g−1DW) > LTCD (4.01 µg·g−1DW) > 0 h (1.01 µg·g−1DW) (Fig. 2d). The relative content of jasmone was significantly higher (p < 0.05) in LTY than other treatments (Supplemental Table S3).
Gene expression of volatiles synthesis under different spreading treatments
-
Transcriptome analysis results showed that the differential metabolism of aromatic substances caused by different spreading treatments mainly involved the synthesis of terpenoids (monoterpenes and sesquiterpenes), the catabolism of α-linolenic acid and the metabolism of phenylalanine.
Gene expression of terpenoid synthesis under different spreading treatments
-
The mevalonate (MVA) pathway mainly occurs in the cytoplasm and is the synthetic pathway of farnesyl pyrophosphate, which is the precursor of sesquiterpenes, while the precursor of monoterpenes, geranyl pyrophosphate is synthesized in the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway, which mainly occurs in the plastid.
For the MVA pathway, the transcriptome analysis results (Fig. 3) and qRT-PCR results (Supplemental Fig. S1) revealed a total of six differentially expressed genes (DEGs) in the spreading leaves, with a significant (P < 0.05) up-regulation for ACAT2 (acetyl-CoA acyltransferase), HMG1 (3-hydroxy-3-methylglutaryl-CoA reductase), MVD (mevalonate pyrophosphate decarboxylase) and GPS1 (geranyl pyrophosphate synthase), and a significant (p < 0.05) down-regulation for HMGS (3-hydroxy-3-methylglutaryl-CoA synthase) and MK (mevalonate kinase). During spreading, the expression of ACAT2 decreased slightly in the early spreading stage and increased significantly in the later stage, while the expression of MVD continued to increase. At the end of spreading, the gene expression RQ value in LTY treatment was significantly higher (p < 0.05) than those in other treatments. The expression levels of HMG1 and GPS1 increased sharply in the early spreading stage, and showed a greater increase in the three low-temperature spreading treatments, especially for LTY.
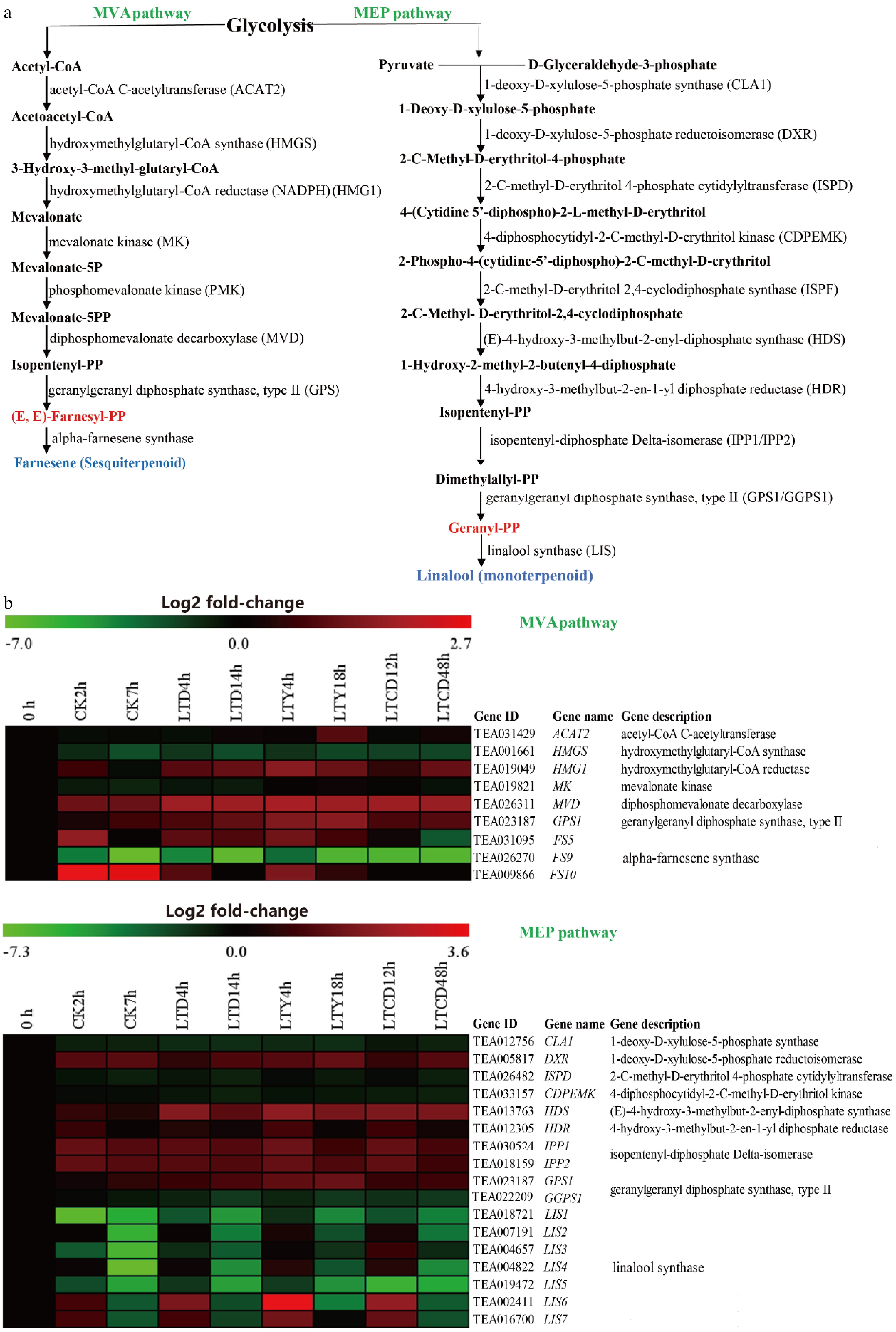
Figure 3. The expression of DEGs involved in terpenoid synthesis under different spreading treatments. (a) KEGG pathways, (b) heat maps of DEGs expression.
For the MEP pathway, a total of ten differentially expressed genes were identified, with a significant (p < 0.05) up-regulation for DXR (l-Deoxy-D-xylulose-5-phosphate reductoisomerase), HDS (4-hydroxy-2-methyl-2-E-butenyl-4-pyrophosphate synthase), HDR (hydroxy-methyl-butenyl pyrophosphate reductase), IPP1/IPP2 (isoprene pyrophosphate isomerase) and GPS1 (geranyl pyrophosphate synthase), while a significant (p < 0.05) down-regulation for CLA1 (5-phosphate deoxy xylulose synthase), ISPD (methylerythritol-cytosine dinucleotide synthase), and CDPEMK (4-(5-cytidine pyrophosphate)-2-C-methyl-D-erythritol kinase). The expressions of DXR, HDS, HDR, and IPP1 were up-regulated in all treatments, but the increase was larger in the three low-temperature treatments than in CK, with the most significant increase (p < 0.05) in LTY. The expression of GGPS1 showed a downward trend in the spreading process, while the expression of GPS1 showed a sharp increase in the early spreading stage, with the most significant (p < 0.05) increase in LTY (Supplemental Fig. S1). The above results indicated that the synthesis reaction of terpenoid precursors continues in fresh tea leaves after spreading. Compared to natural spreading (CK), the low-temperature and yellow light irradiation treatments could promote the synthesis of terpenoid precursors.
Farnesyl pyrophosphate can be synthesized to sesquiterpenes (farnesene, farnesol, nerolidol, etc.). A total of three differentially expressed genes (FS5, FS9, FS10) of the farnesene synthase (FS) gene family were identified in the different spreading treatments (Supplemental Fig. S1). The expression of FS9 was steadily down-regulated during spreading, in contrast to a significant increase (p < 0.05) in the expression of FS5 and FS10 in the early spreading stage of CK, LTD and LTY treatments, but a decrease in the LTCD treatment. The expression pattern of FS10 is consistent with that of FS5, with the highest up-regulation for LTY, followed by LTD and CK.
Geranyl pyrophosphate was the substrate for synthesizing monoterpenes (geraniol, linalool, nerol, etc.), and a total of six differentially expressed genes (LIS1, LIS2, LIS3, LIS4, LIS5, LIS6) in the linalool synthase (LIS) gene family were identified in the four different spreading treatments (Supplemental Fig. S1). The expression of LIS1, LIS5 and LIS6 was steadily down-regulated during spreading, while the expression of LIS2 and LIS4 were down-regulated in CK and LTD treatments, and up-regulated in the early spreading stage of LTY and LTCD treatments. The expression trend of LIS2 was consistent with that of LIS4, with the highest up-regulation in LTY treatment. Meanwhile, the expression of LIS3 was down-regulated in CK, LTD and LTCD, but significantly (p < 0.05) up-regulated in the early stage of LTY treatment, indicating that the synthesis reaction of some monoterpenoids and sesquiterpene volatiles was still proceeding, and terpenoid synthesis even showed an upward trend in the early spreading stage. Compared to the other treatments, LTY treatment showed a most obvious enhancement in the terpenoid synthesis reaction.
The activities of terpenoid precursor synthase, linalool synthase, and farnesene synthase were further analyzed during the spreading process (Supplemental Fig. S2). The activity of certain synthetase, farnesene synthase in the MVA pathway and linalool synthetase in the MEP pathway similarly had a significant increase (p < 0.05) during spreading. Compared with CK and LTD, LTY and LTCD could more obviously enhance the synthase activity of the MVA pathway.
Gene expression of α-linolenic acid metabolism under different spreading treatments
-
Phosphatidylcholine was catalyzed by phospholipase A2 (PLA2) and hydrolase family (LCAT2/ LCAT3/ DAD1) to produce α-linolenic acid. Transcriptome analysis (Fig. 4) and qRT-PCR (Supplemental Fig. S3) showed that a similar downward trend was observed in the expression of PLA2 and DAD1 during spreading, while the expression of LCAT2 and LCAT3 sharply up-regulated in the early spreading stage. α-Linolenic acid was catalyzed and degraded by lipoxygenase (LOX2/LOX6) to produce 13(S)-HpOTrE. A similar upward trend was observed in the expression of LOX2 and LOX6 in the early spreading stage of CK, LTD and LTY treatments. The above results indicated that the degradation reaction of α-linolenic acid was still proceeding during spreading and had a clear upward trend in the early spreading stage. Compared with CK, LTD and LTY showed no significant impact on the initial degradation of α-linolenic acid, while the CO2-controlled atmosphere (LTCD) produced a certain inhibitory effect, suggesting that higher temperatures may have a significant impact on the degradation reaction of α-linolenic acid while low-temperature plus CO2 is not conducive to the degradation of α-linolenic acid.

Figure 4. The expression of DEGs involved in α-linolenic metabolism under different spreading treatments. (a) KEGG pathways; (b) heat map of DEGs expression profiling from the RNA-Seq data.
13(S)-HpOTrE, a direct synthesis precursor of hexenal, hexenol and hexenyl ester, is catalyzed by lipohydroperoxide lyase (HPL1), alcohol dehydrogenase (ADH1) and acetyltransferase (CHAT). Except LTCD, the other three treatments increased the expression of HPL1 and ADH1, with the largest increase in CK, while the expression of CHAT (acetyltransferase) showed no significant change during spreading (Supplemental Fig. S3). The above gene expression pattern was consistent with the results of the aroma volatiles, suggesting that higher environment temperature (CK) facilitates the synthesis of fatty acid-derived volatiles, while the CO2 treatment (LTCD) showed a significant inhibitory effect on it.
Methyl jasmonate is synthesized through another downward metabolic pathway of 13(S)-HpOTrE, which required the catalysis of seven enzymes. The expression of AOS and AOC4 showed a steady in CK, LTD and LTY treatments, with the highest up-regulation in LTY, but down-regulation in LTCD) (Supplemental Fig. S3). In the OPR gene family, the expression of OPR2 was down-regulated during spreading, and the expression of OPR1 and OPR3 both increased sharply in the early spreading stage, followed by a slight decrease. In the ACX gene family, except for ACX4, the expression of ACX2 and ACX3 continued to increase during spreading, with the largest increase in LTY. Meanwhile, the expression of MFP2 also increased during spreading, with a higher up-regulation in LTY and LTD. The expression of PKT3 was up-regulated in the early spreading stage, also with the largest increase in LTY. The above results indicated that the synthesis reaction of methyl jasmonate was significantly enhanced during spreading, and most synthetic genes showed the highest expression in LTY treatment, suggesting that yellow light irradiation is a key contributor to the synthesis of methyl jasmonate.
The activity changes of key enzymes in the α-linolenic acid metabolic pathway during spreading were further analyzed (Supplemental Fig. S4). Compared to CK, LTD and LTY showed no significant effect on the initial degradation of α-linolenic acid, while the CO2 controlled atmosphere (LTCD) had a significant inhibitory effect. The results also indicate that the low temperature environment does not facilitate the production of hexenal and hexenol and the CO2 environment may significantly inhibit it; but for methyl jasmonate, low temperature and yellow light environment can significantly enhance the synthesis in spreading leaves.
Gene expression of phenylalanine metabolism under different spreading treatments
-
Transcriptome results (Fig. 5) and qRT-PCR results (Supplemental Fig. S5a) showed that the genes involved in the synthesis of phenylethanol and phenylacetaldehyde showed an increased, but not significant, expression. There are three enzymes involved in phenylethanol synthesis, including aromatic-L-amino acid decarboxylase (AAS), primary-amine oxidase (AO2), and phenylacetaldehyde reductase (PAR). During spreading, the expression of AAS kept decreasing, and the decrease was more obvious in the three low temperature treatments than in CK, with the greatest decrease in LTCD. Meanwhile, the expression of AO2 increased significantly (p < 0.05), with the most obvious increase for LTY treatment, and the expression of PAR was significantly higher (p < 0.05) in the three low temperature treatments than in CK, also with the largest increase in LTY, followed by LTCD.
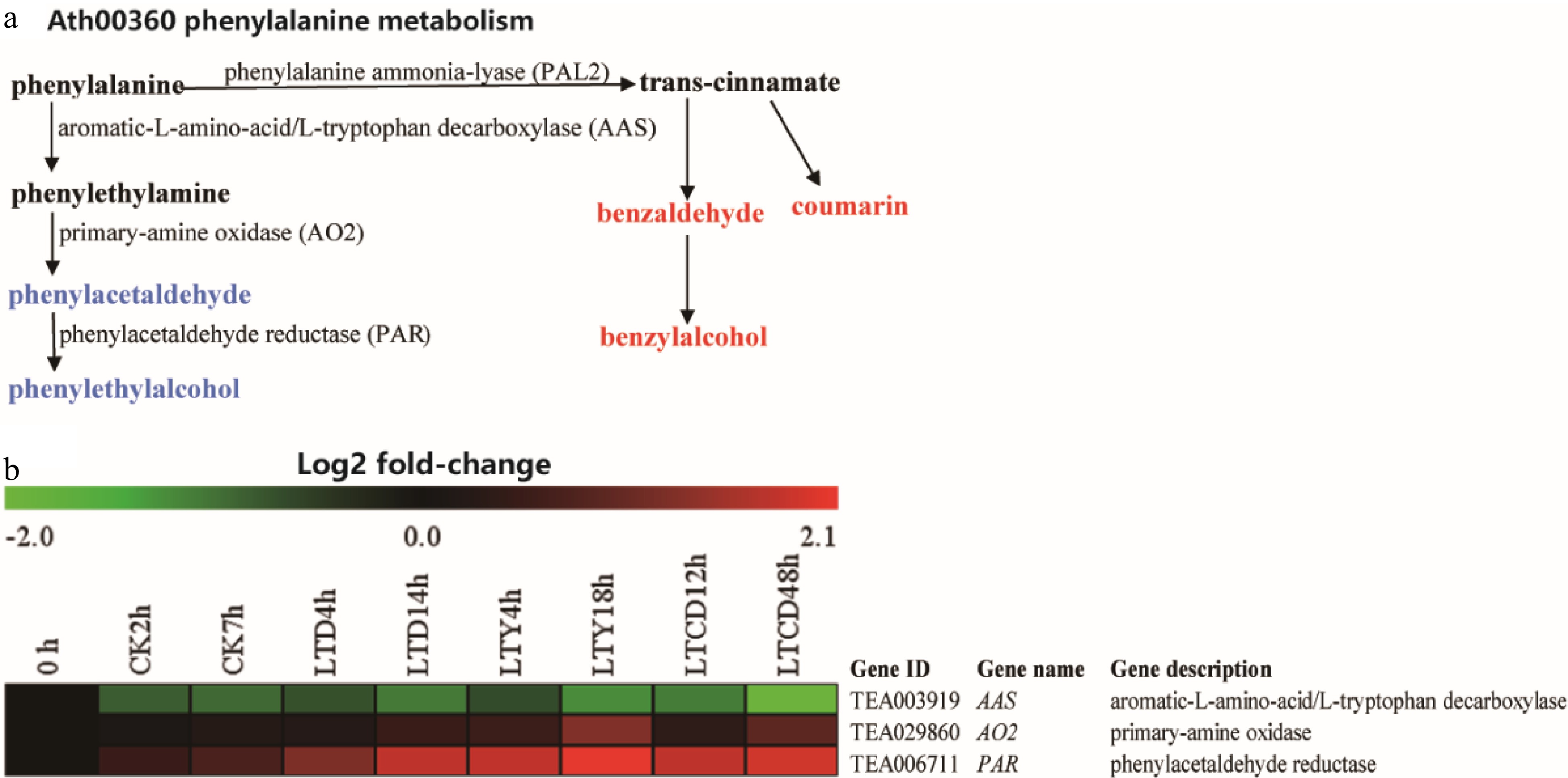
Figure 5. The expression of DEGS involved in phenylethylalcohol biosynthesis under different spreading treatments. (a) KEGG pathways; (b) heat map of DEGs expression profiling from the RNA-Seq data.
Changes in the activity of phenylacetaldehyde and phenylethanol synthase under different withering treatments were further analyzed (Supplemental Fig. S5b). The trend of the phenylacetaldehyde/phenylethanol synthase activity was roughly consistent with the expression pattern of the corresponding genes, indicating that the synthesis reaction of phenylethanol and phenylacetaldehyde is still proceeding and tends to increase during spreading, which is most obvious in LTY. As a result, it leads to a higher content of phenylethyl alcohol in LTY.
-
The experiment was designed to verify if a low-temperature spreading environment could preserve or even enhance the sensory quality of green tea products under the premise of prolonging the spreading period of picked tea leaves. To ensure that the physiological conditions of spreading leaves processed into tea products were consistent, we tried to control the moisture content of spreading leaves under the four treatments within the range of insignificant difference. For example, at the end of the treatments, the water-content in spreading leaves was 72.1 ± 0.4% (CK-7 h), 72.2 ± 0.4% (LTD-14 h), 71.8 ± 0.3% (LTY-18 h), and 74.2 ± 0.2% (LTCD-48 h), respectively. There were no significant differences among the three leaf samples of CK-7 h, LTD-14 h and LTY-18 h. Only the sample of LTCD-48 h was different (p < 0.05) from the other three samples. And the four leaf samples of CK-7 h, LTD-14 h, LTY-18 h and LTCD-48 h were processed into green tea products under the same processing method. Therefore, it was reasonable to analysis the variation of volatile and transcriptome during CK (CK-2 h, CK-7 h), LTD (LTD-4 h, LTD-14 h), LTY (LTY-4 h, LTY-18 h), and LTCD (LTCD-12 h, LTCD-36 h) treatments. Furthermore, it was similarly reasonable to compare the sensory quality and volatiles contents in the four tea products, because they were based on the relatively same conditions (the relatively same level of moisture-degree in withered leaves and the same tea processing methods) despite the different treatment duration.
The content of most aroma volatiles in fresh tea leaves increased significantly after spreading, indicating the necessity of spreading for enhancing green tea aroma. There are two possible explanations for the increase in the content of aroma volatiles in the spreading process: (i) aroma volatiles are synthesized de novo in the fresh tea leaves after harvest, and (ii) aroma volatiles result from hydrolysis by glycosidically bound volatiles. Studies have shown that the latter does not contribute much to the aroma enhancement of withered leaves[15]. Compared to natural spreading (CK), the three low-temperature treatments showed a larger increase in the content of aroma volatiles, which was consistent with previous studies[15,16], confirming the content increase of some terpene compounds like indole, jasmone, jasmine lactone, vanillin and other aroma volatiles under low-temperature spreading environment.
Under low-temperature plus yellow-light irradiation (LTY), the content of aromatic substances (mainly terpene alcohols) in fresh tea leaves increased significantly (P < 0.01). Fu et al.[17]found that, compared with dark treatment, withering with blue and red light was significantly higher (p < 0.05) in the content of indole after treatment for 2 h, and the contents of nerol, pentenol, and phenylacetaldehyde were relatively higher after the treatment for 4 h. Few studies have been performed on the application of modified CO2 preservation technology in tea relative to the related research on fruits. Lalel et al.[18] showed that the total amount of aroma volatiles was significantly higher (p < 0.05) in low-concentration CO2 (2%−3%) atmosphere preservation treatment for 20−30 days than the single low-temperature treatment.
Compared with CK, low temperature may enhance the expression of related genes in the terpenoid synthesis pathway to promote the synthesis and release of terpenoids. Katsuno et al.[16] found that the release of linalool in low temperature spreading leaves was significantly higher (peak area ratio > 2) than that in natural spreading leaves, and the expression of terpenoid synthase gene (TPS) was also significantly enhanced in low temperature spreading environments. The low-temperature induced up-regulation of terpenoid synthesis genes is also found in the preservation of vegetables[19] and fruits[20].
The buds, leaves, flowers and fruits of in vitro plants can still perceive external light signals after harvest[21] and light can stimulate the expression of key genes for terpenoid synthesis, thereby enhancing the synthesis and release of terpenoids. Fu et al.[17] found that the release of volatile terpenoids (linalool, geraniol) was significantly improved (p < 0.05) after a period of irradiation by LED blue light (470 nm) and red light (660 nm), and the fresh tea leaves also showed a significant increase in the expression level of the terpenoid synthase gene (TPS).
In this study, the transcriptome and qRT-PCR results revealed a few differentially expressed genes in the terpenoid synthesis pathway between LTCD and LTD treatments, but the expressions of ACAT2, MK and CLA1 were significantly up-regulated (p < 0.05) at the end of LTCD treatment, suggesting that, compared to LTD, the supplementation of CO2 can more significantly promote the synthesis of terpenoid precursors[22].
It was documented that the degradation and metabolism of fatty acids in fresh tea leaves during spreading are the main contributors to the sharp increase in the content of FADVs[12,23]. Compared with CK, the contents of hexanol and hexenyl esters were lower in the low-temperature spreading samples (LTD, LTY, and LTCD), which is consistent with previous observations in postharvest cantaloupe[24], strawberry[25], tomatoes[26]and other fruits and vegetables. Therefore, higher spreading temperature is the main inducer of the production and release of fatty acid derivatives in fresh tea leaves. The effect of light on the degradation of fatty acids varies in different postharvest plants. In the present study, the release of FADVs is not significantly different between LTY and LTD, possibly due to no significant difference in the expression of most genes in different treatments. The release of FADVs from post-harvest fresh tea leaves can be obviously inhibited by the CO2 atmosphere (LTCD). CO2 not only inhibits the aerobic metabolism of postharvest plants, leading to the deficiency of ATP and NADPH energy required for fatty acid degradation reaction[27], but is also involved in the inhibition of ethylene release in postharvest horticultural crops[28].
The release of phenylpropane volatiles was higher under low temperature spreading than under natural spreading (CK). One possible explanation is that the postharvest plants maintain the osmotic balance of the cells by producing and accumulating higher levels of phenylalanine in order to resist the stress of water loss in the low temperature environment[29], and then that higher levels of phenylalanine can also provide more synthetic precursors for phenylpropane volatiles[30], thus improving the production and release of phenylethanol.
The synthesis of phenylpropane volatiles and the expression of related genes can be enhanced by light. Cheng et al.[23] reported that the release of benzaldehyde/benzyl alcohol and phenylacetaldehyde/phenylethanol in isolated petunia flowers increased under LED red light, and the expression levels of PhPAL2, PhPAR, and other genes were significantly higher in light (p < 0.05) than in dark. The release of phenylethanol from fresh tea leaves showed a greater increase after CO2 treatment than a single low-temperature treatment. Similar observations were reported in the processing of post-harvest mango[18] and tomato[26].
-
In this research, the metabolism of key aroma substances in tea leaves was influenced obviously by different spreading treatments. Compared with natural spreading, the low-temperature spreading environment (especially low-temperature plus yellow-light treatment) could well retain or even enhance the aroma quality of green tea products. This research facilitates the understanding of volatile metabolism in postharvest tea leaves and provides a genetic metabolic basis for the application of low-temperature spreading in actual green tea processing.
-
Anhydrous ether (chromatographic grade) was purchased from Thermo Fisher Scientific Corporation (Waltham, Massachusetts, USA); ethyl decanoate, guaiacol (chromatographic grade) and anhydrous sodium sulfate (analytical grade) were purchased from Maclean Biochemical Technology Co., Ltd. (Shanghai, China); N-alkanes (C3-C25) was purchased from O2si Corporation (South Carolina, USA); the rapid RNA extraction kit, the Enzyme-Linked Immunosorbent Assay (ELISA) Kit, the TRUE Script Reverse Transcription (RT) Kit with gDNA Eraser and the 2×Sybr Green qPCR Mix were purchased from Beijing Adler Biotechnology Co.
Sample preparation
-
Tea 'Echa No. 10' [C. sinensis (L.) O. Kuntze] leaves comprising one bud with two leaves were harvested in September from the tea plantation of Huazhong Agriculture University (Wuhan, China). Picked tea leaves were spreading in four different treatments, namely CK: the indoor natural spreading [25 °C, 65% RH (relative humidity), 200 lx natural light]; LTD: low temperature (15 °C, 70% RH, darkness); LTY: low temperature plus yellow-lighting (15 °C, 70% RH, 250 lx / 9 W LED lights); LTCD: low temperature plus CO2 (15°C, 70% RH, 3% CO2 and darkness). At the end of each treatment, spreading leaves were subjected to fixation at 260 °C in a continuous roller fixation machine (6CST-50, Zhejiang Green Peak Machinery Co., Ltd., Quzhou, China). Subsequently, the de-enzymed leaves were placed on the tray of a far-infrared drying oven (101-3Y, Shanghai Zhetai Machinery Co., Ltd., Shanghai, China) and heated at 105 °C for 30 min, followed by cooling outside the oven, then heated again at 90 °C for 60 min, cooling again, and finally heated at 90 °C for 30 min. Treatment had been completed by three repetitions.
Analysis of volatile components in spreading leaves and green tea products
-
Volatile collection was conducted as described in the study by Xu et al.[31] with some minor modifications. Briefly, each leaf sample (1.0 g) was firstly placed into a 20 mL sealed glass vial, followed by adding 5 mL of boiling deionized water which contained 4.516 µg of guaiacol as an internal standard (used guaiacol as the IS[32], putting the vial immediately into a water bath heater at 60 °C, and allowing the volatiles to be adsorbed for 60 min by a CAR/PDMS (85 µm; 1 cm) coating fiber (Supelco, Inc., Bellefonte, PA, USA)).
GC-MS analysis was performed under the following conditions: instrument, a gas-mass spectrometer (TRACE DSQ II, Thermos); the chromatographic column, DB-5MS (30 mm × 0.25 mm × 0.22 μm, Agilent, USA); the carrier gas, helium (99.999%); flow rate, 1.0 mL/min; the inlet temperature, 250 °C; the transfer line temperature, 250 °C; the column temperature, 40 °C; sample injection without splitting. Heating program: initial temperature 40 °C, hold for 2 min, increase to 85 °C at 5 °C/min, hold for 2 min, then increase to 110 °C at 2 °C/min, 130 °C at 7 °C/min, 230 °C at 5 °C/min, and hold for 8 min. Ionization method: electron energy 70 eV, ion source temperature 230 °C, and scanning range 60−550 m/z.
Volatile compounds were identified by comparing with the standard substance (N-alkanes C3-C25), the published retaining index[32] as well as the National Institute of Standards and Technology (NIST) mass spectral database. The relative content of volatile compounds was calculated as the formula:
Relative content (mg/g DW) = peak area (compound) / peak area (internal standard) * 4.516 / sample dry weight
Identification of DEGs involved in volatile metabolism
-
The spreading tea leaf samples (0 h, CK-2 h, CK-7 h, LTD-4 h, LTD-14 h, LTY-4 h, LTY-18 h and LTCD-12 h, LTCD-48 h) were performed for the transcriptome analysis based on the reference genome TPIA (Tea Plant Information Archive)[33], and detailed sequencing results have been published[10]. DEGs, related to the metabolism of volatiles (e.g., sesquiterpenes, monoterpenes, jasmonic acid, methyl jasmonate, benzyl alcohol, phenethyl alcohol, benzaldehyde, phenylacetaldehyde, etc.) were identified using KEGG annotation and significantly enriched via KEGG pathways, and filtrated by |log2 (fold change)| ≥ 1 and an adjusted p < 0.05 in pairwise comparison groups.
Validation of the DEGs by quantitative real-time PCR (qRT-PCR)
-
qRT-PCR analysis was performed to verify RNA-seq results. The relative expression levels of the selected transcripts, normalized against Csβ-ACTIN expression levels, were calculated using the 2−ΔΔCᴛ method. Each reaction was performed in three biological replicates. The primer information was listed in the Supplemental Table S4.
Determination of enzymes activity related to volatile metabolism
-
Enzyme activity was determined as described in the study by Luo et al.[34]. Total proteins were first extracted from tea leaf samples using a Plant Total Protein Extraction Kit, and their concentrations were detected using a BCA Protein Concentration Determination Kit. Finally, the extracted protein samples were used for analyzing the activities (units/g of protein) of key enzymes involved in terpene biosynthesis, α-linolenic acid catabolism, phenylethyl alcohol biosynthesis by using the ELISA Kit according to the manufacturer's instructions.
Organoleptic evaluation
-
The organoleptic evaluation of four green tea products (CK, LTD, LTY, and LTCD) was performed by five panelists using a 100-score grading system (GB/T 23776-2018), with 30% assigned to aroma, 30% to taste, 10% to the shape of dry tea leaves, 10% to the color of dry tea leaves, 10% to liquor color, and 10% to infused leaf.
Statistical analysis
-
Statistical analysis was performed using Excel and SPSS19.0 software. All the results were shown as mean ± standard error. Multiple comparisons among groups of data were performed by one-way analysis of variance (ANOVA, Duncan test) and Tukey's HSD test at the level of p < 0.05 and 0.01. Experimental data were visualized using Origin 9.1 and Adobe Illustrator CS6 software.
- This work was supported by the National Key R & D Program of China (Project No. 2018YFD1000600) and the Fundamental Research Funds for the Central Universities, Huazhong Agricultural University (Project No. 2662017PY053).
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 Sensory quality scores of the green tea products.
- Supplemental Table S2 The relative contents (µg/g DW) of aroma components in green tea products.
- Supplemental Table S3 The relative contents (µg/g DW) of aroma components in different spreading samples.
- Supplemental Table S4 PCR primers used for gene expression analysis.
- Supplemental Fig. S1 The expression levels of DEGs involved in terpenoid biosynthesis under different spreading treatments.
- Supplemental Fig. S2 The enzyme activity of key enzymes involved in terpenoid biosynthesis under different spreading treatments.
- Supplemental Fig. S3 The expression levels of DEGs involved in the catabolic metabolism of α-linolenic acid under different spreading treatments.
- Supplemental Fig. S4 The enzyme activity of key enzymes involved in the catabolic metabolism of α-linolenic acid under different spreading treatments.
- Supplemental Fig. S5 The expression levels of DEGs (a) and the enzyme activity of key enzymes (b) involved in biosynthesis of phenylethyl alcohol under different spreading treatments.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yu X, He C, Li Y, Zhou J, Chen Y, et al. 2021. Effects of different spreading treatments on the formation of aroma quality in green tea. Beverage Plant Research 1: 14 doi: 10.48130/BPR-2021-0014

Effects of different spreading treatments on the formation of aroma quality in green tea
- Received: 03 November 2021
- Accepted: 26 November 2021
- Published online: 13 December 2021
Abstract: The current research was carried out to verify if low-temperature spreading treatments (CK, natural spreading; LTD, low temperature + dark; LTY, low temperature + yellow light; LTCD, low temperature + CO2) had a positive effect on the aroma quality of green tea products under the premise of prolonging the spreading period of picked tea leaves. Compared to CK, LTY significantly improved the aroma quality in green tea products. The spreading leaves with LTY treatment showed the highest relative content of terpene volatiles and amino acid-derived volatiles. Furthermore, terpene precursor synthesis genes, such as linalool synthase genes (CsLIS2, CsLIS3, CsLIS4) and farnesene synthase genes (CsFS5, CsFS10) were significantly up-regulated in spreading leaves after LTY treatment. This research furthers our understanding of volatile metabolism in postharvest tea leaves and provides a genetic metabolic basis for the application of low-temperature spreading in green tea processing.
-
Key words:
- Low-temperature spreading /
- Aroma quality of green tea /
- SPME-GC-MS /
- Volatile metabolism