









-
Due to its vitamin, mineral and amino acid content, kiwifruit is an economically and nutritionally important fruit crop. Due to its good marketability and high yield compared to other cultivars, 'Hayward' is the most widely planted in the world[1]. As with other horticultural crops, fruit size is an important indicator of the commercial desirability of kiwifruit[2,3]. After fruit set, there is an active cell division phase followed by a cell expansion phase; both together promote fruit growth and determine the size of the fruit[4]. At this stage, plant growth regulators (PGRs), especially auxins (IAA), cytokinins (CKs) and gibberellins (GAs), are the main regulators of fruit cell division and expansion[5]. IAA and GAs regulate fruit growth by promoting fruit expansion[6,7], while CKs promote fruit cell division[8]. Some genes related to hormone synthesis or signaling pathways are also known to regulate fruit development, such as auxin response factor (ARFs) and transport inhibitor response 1 (TIR1)[9], GA biosynthesis enzymes such as GA 20-oxidase3 and GA 3-oxidase, and the negative regulator of GA signaling DELLA[10].
Carbohydrates are essential for fruit development. In addition to the regulatory roles of PGRs, carbohydrate availability also affects fruit growth. Higher plants are able to transfer organic carbon produced by photosynthesis from the source leaves to the sink tissue via the sieve element/chaperone cell complex[11]. The sink organs utilize photosynthetic products, and this capacity is known as sink strength[12]. The sink strength determines the distribution of carbohydrates in the whole plant[13]. In kiwifruit, photosynthetic products are transported as sucrose, and rapid elimination of sucrose is necessary to continue phloem unloading and to facilitate carbon transport from source to sink organs[14]. Sucrose synthase (SUSY) and invertases are two different types of enzymes that contribute to the cleavage of sucrose[15]. Starch metabolism also plays an important role in kiwifruit fruit development. Key enzymes for starch synthesis and degradation have been revealed, including ADP-glucose pyrophosphorylase (AGPase), starch branching enzyme (SBE), and β−amylase (β-AMY)[16].
Since PGRs coordinate multiple aspects of fruit development and ripening, they can be used during commercial production to increase fruit fresh weight and yield. The growth response to PGR application depends on the dosage and timing of application[17]. Forchlorfenuron (CPPU) is a widely used synthetic PGR that is believed to work synergistically with endogenous auxin to increase the size and cluster weight of a variety of fruit crops and induce parthenogenesis[18,19]. In horticultural production, it is applied to kiwifruit, grape[20], watermelon[21], apple[22] and pear[23] to promote fruit set and increase crop yield. In Hayward kiwi, CPPU treatment increases sugar accumulation and decreases acidity and hardness[24,25]. Patterson et al.[26] showed that CPPU treatment increases kiwifruit fresh weight by promoting cell expansion. Ainalidou et al.[27] showed that CPPU increases 'Hayward' kiwifruit size by enhancing cell expansion rather than cell division, while Wu et al.[25] demonstrated that CPPU promoted 'Hongyang' kiwifruit enlargement by promoting both cell division and expansion. Although there have been many studies on the effect of CPPU treatment on kiwifruit fruit growth, the molecular mechanism by which CPPU treatment regulates fruit enlargement (the period of rapid fruit development) is unclear.
Understanding the mechanisms of fruit size regulation has important implications for kiwifruit quality improvement and breeding. In the present study, we combined transcriptomic and physiological data to reveal the changes in the metabolism and signal transduction related to sugars and phytohormones within kiwifruit after CPPU treatment during the rapid fruit developmental stage.
-
'Hayward' kiwifruit was grown in Shangde Village, Yangling District, Shaanxi Province, China. Thirty vines with uniform growth were selected as experimental subjects. Forchlorfenuron (CPPU) treatment was performed as previously described[17]. The young fruit was immersed in CPPU solution (50 mg/L) for 2−3 seconds at 21 d after bloom (DAB). Fruits treated with water were used as control. Ten kiwifruit samples were randomly collected at 1 week (28 DAB), 2 weeks (35 DAB), 3 weeks (42 DAB), 4 weeks (49 DAB), 7 weeks (70 DAB), 11 weeks (98 DAB), and 19 weeks (154 DAB, fruit was considered mature for harvest on this day) after treatment, respectively. The fruits were quickly cut into small pieces after being harvested and frozen in liquid nitrogen. All samples were stored at –80 °C until further analysis. Another five fruits were harvested to determine the physiological indexes of fresh weight, cross-sectional diameter, soluble solids content and dry matter content.
Determination of soluble sugars and starch
-
Soluble sugars were extracted according to methods previously described[28]. Fruit samples (0.1 g) were mixed with 1.4 ml 75% methanol (pre-chilled at –20 °C) and 100 µl 400 ppm Ribitol and incubated at 70 °C on mental dry bath at 950 rpm for 30 min. The extract was centrifuged at 12000 rpm for 10 min. The supernatant was removed to a new centrifuge tube and then 750 µl chloroform (pre-chilled at –20 °C) and 1400 µl ddH2O were added, vortexed, and centrifuged at 2200 rpm for 15 min. An appropriate amount of supernatant was vacuum dried for 1 h. The dried product was then derivatized with methoxyamine hydrochloride and N-methyl-N-trimethylsilyl-trifluoroacetamide. After being derivatized, an Agilent 7890A GC/5975C MS system (Agilent Technology, Palo Alto, CA, USA) was used to analyze the metabolites.
The sugar extract was washed with 80% ethanol to remove soluble sugars and precipitated using 1 ml 0.2 mol/L NaOH to determine starch content. The precipitate was boiled in boiling water to form a gel, and then acetic acid was added to adjust the pH to 4.5. The starch was then cleaved with amyloglucosidase at 55 °C for 1 h, after which the enzyme was inactivated in boiled water for 1 min. After the solution was cooled and centrifuged, the supernatant was used to determine the starch content by spectrophotometric method in A540. The starch content was calculated by comparison to a glucose standard curve.
Determination of enzyme activity related to sugar metabolism
-
The enzyme was extracted using the method described by Moscatello et al.[14]. Sucrose phosphate synthase (SPS) activity was determined in a two-step assay. The enzyme solution (35 µl) was incubated for 30 min at 37 °C in a 70-µl volume of assay medium containing 50 mM HEPES-KOH (pH 7.5), 15 mM MgCl2, 1 mM EDTA, 4 mM fructose-6-phosphate, 14 mM glucose-6-phosphatase and 16 mM UDPG. In the control, the fructose-6-phosphate and glucose-6-phosphatase were replaced with 8.5 μl water. After incubation, 70 µL of 1 M NaOH was added to the reaction solution, and the reaction was finished by placing in a boiling water bath for 10 min. After removing and cooling, 0.75 ml of 30% HCl and 0.25 ml of 1% resorcinol were added to the solution, and the reaction was carried out at 80 °C for 8 min. Colorimetry was performed using a spectrophotometer at A480.
The sucrose synthase (SUSY) reaction system (500 µl) contained 80 mM MES, 100 mM sucrose, 5 mM UDP (the control is replaced by water), and 100 µl enzyme solution (the control is replaced by inactivated enzyme solution). After adding 500 µl 3,5-dinitrosalicylic acid (DNS), the mixture was incubated at 30 °C for 30 min. DNS (500 µl) was added after the mixture was cooled to room temperature, and the reaction was terminated in a boiling water bath for 5 min. To determine NINV activity, the assay (500 µl) contained 80 mM acetic acid-sodium acetate buffer (pH 4.5), 100 mM sucrose and 100 µl enzyme solution (the control is replaced by inactivated enzyme solution). The reaction was mixed with 500 µl of DNS and incubated for 30 min at 30 °C. After cooling, 500 µl of DNS was added followed by a boiling water bath for 5 min. Both SUSY and NINV were used as control with water and colorimetric using spectrophotometer under A540 conditions.
RNA extraction and library construction
-
Total RNA was extracted by CTAB method. The cDNA was generated by a reverse transcription kit (Takara Bio, USA), according to the manufacturer's instructions. RNA was extracted from kiwifruit samples and controls after one, two, and three weeks of CPPU treatment, respectively. After testing the quality of the extracted RNA, the library construction and sequencing process followed the standard protocols provided by Illumina. cDNA libraries were sequenced on an Illumina high-throughput sequencing platform, based on Sequencing By Synthesis (SBS) technology. Clean data (clean reads) were obtained by removing reads containing adapter, poly-N and low-quality reads from the raw data. The clean data (six treatments and three replicates) were aligned separately to the kiwifruit reference genome database (http://kiwifruitgenome.org/organism/3) using hisat2[29]. After converting the result files using Samtools[30] to BAM files, the statistics were compiled for reads with uniquely matched genes using FeatureCount[31] and then analyzed for differential expression using DEGseq2[32]. Gene function was annotated using GO and KEGG analysis. Genes with an adjusted P-value < 0.05 and a Fold Change ≥ 1.5 found by DEseq were assigned as differentially expressed genes (DEGs).
Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) analysis
-
To verify the accuracy of the RNA-sequencing results, we used the same RNA samples to detect the expression levels of nine randomly selected genes by qRT-PCR. The length of all qRT-PCR products was designed to be between 170 to 200 using primers listed in Supplemental Table S1. qRT-PCR was performed using a QuantStudio 5 real-time PCR system (manufacturer). The reaction volume for the qRT-PCR assay was 20 µl, including 10 µl of 2 × ChamQTM SYBR qPCR Master Mix (Dingling), 1 µl of each primer (10 μM), 2 µl of cDNA and 6 µl of water. The reaction conditions were as follows: hold at 94 °C for 2 min, followed by 40 cycles of 94 °C for 15 s and 60 °C for 30 s, and completed by the melting curve analysis procedure. The relative expression levels of genes were calculated according to the 2−ΔΔCᴛ method[33]. AcActin (GI:149938963) was used as control[25].
Statistical analysis
-
The physiological data were analyzed by one-way ANOVA. Significant differences for all data were assessed by SPSS version 16.0 (SPSS Inc., USA) according to Duncan's multiple range test (p < 0.05).
-
Consistent with a previous study[14], CPPU treatment markedly increased the weight and size of kiwifruit fruit, with the most growth occurring from 28 to 49 DAB(Fig. 1a, 1b). CPPU treatment markedly enhanced initial fruit growth, especially at 28 and 35 DAB. At harvest, the application of CPPU significantly increased the fresh weight of kiwifruit. At 154 DAB, the fresh weight of kiwifruits treated with 50 mg/L CPPU increased by 64% compared to untreated fruits. Compared with untreated fruits, the vertical (Fig. 1c) and horizontal (Fig. 1d) diameters of the fruits increased from 62.12 to 76.91 mm and from 52.65 to 60.23 mm after CPPU treatment at 154 DAB, respectively. Vertical diameter and horizontal diameter increased by 23.81% and 14.39% at harvest, respectively. The maximum daily fruit growth rate after CPPU treatment was observed at 28 DAB and 35 DAB (Fig. 1b). CPPU treatment increased the growth rate of 'Hayward' fruit from 28 DAB to 35 DAB, a period representing the cell division phase of fruit development. Our data shows that the greatest contribution of CPPU treatment to fruit development was between 28 and 42 DAB. Based on this result, RNA-seq was performed to analyze the changes in transcript abundance during the key period of kiwifruit fruit development with CPPU treatment.
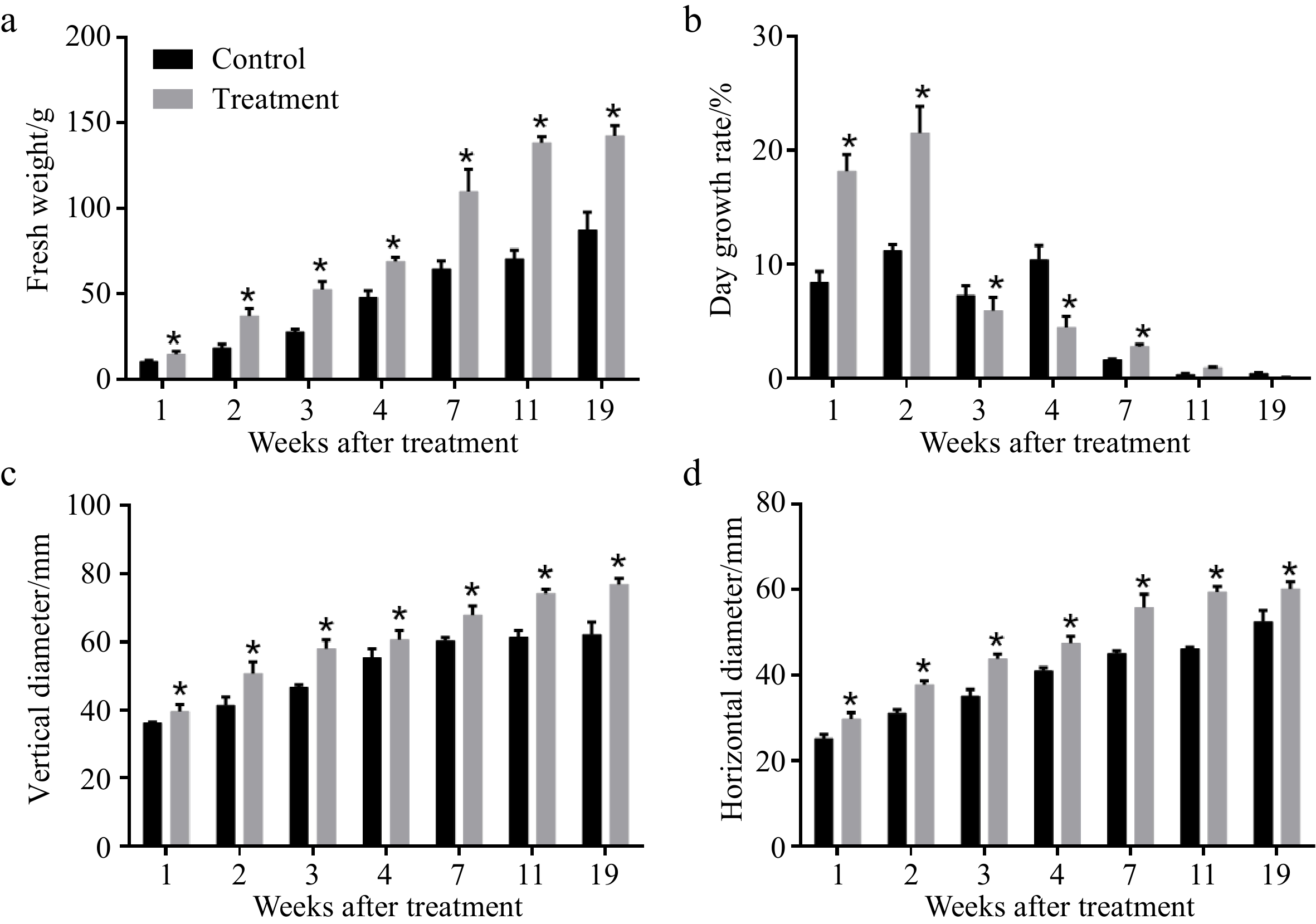
Figure 1. Effects of CPPU on (a) fresh weight, (b) day growth rate, (c) vertical diameter, and (d) horizontal diameter of 'Hayward' kiwifruit. * Indicates significant difference between the treatment and the control based on t-test (p < 0.05).
CPPU application affected the contents of dry matter, starch and soluble sugars in 'Hayward' kiwifruit
-
The dry matter, starch and soluble sugar content were determined to reveal the effect of CPPU treatment on the carbohydrate content of kiwifruit. The dry matter content was not significantly different between treated and untreated kiwifruit (Fig. 2a), which means that the application of CPPU had no obvious effect on the dry matter content of 'Hayward' fruit. Starch content increased markedly in CPPU-treated kiwifruit (Fig. 2b). Except at 28 DAB, the starch content of the treatment group was higher than the control group. At harvest, starch content in the treatment group had increased by 20% compared to the control group. CPPU treatment caused changes in soluble sugar within 'Hayward' fruits. The content of sucrose (Fig. 2c), glucose (Fig. 2d) and galactose (Fig. 2f) were significantly increased by exogenous CPPU during fruit development, especially in the initial growth stages. In addition, no significant differences in fructose (Fig. 2e) content between the treated and control groups were detected until 49 DAB. After 49 DAB, fructose content in the treatment group was much higher than that in the control group.
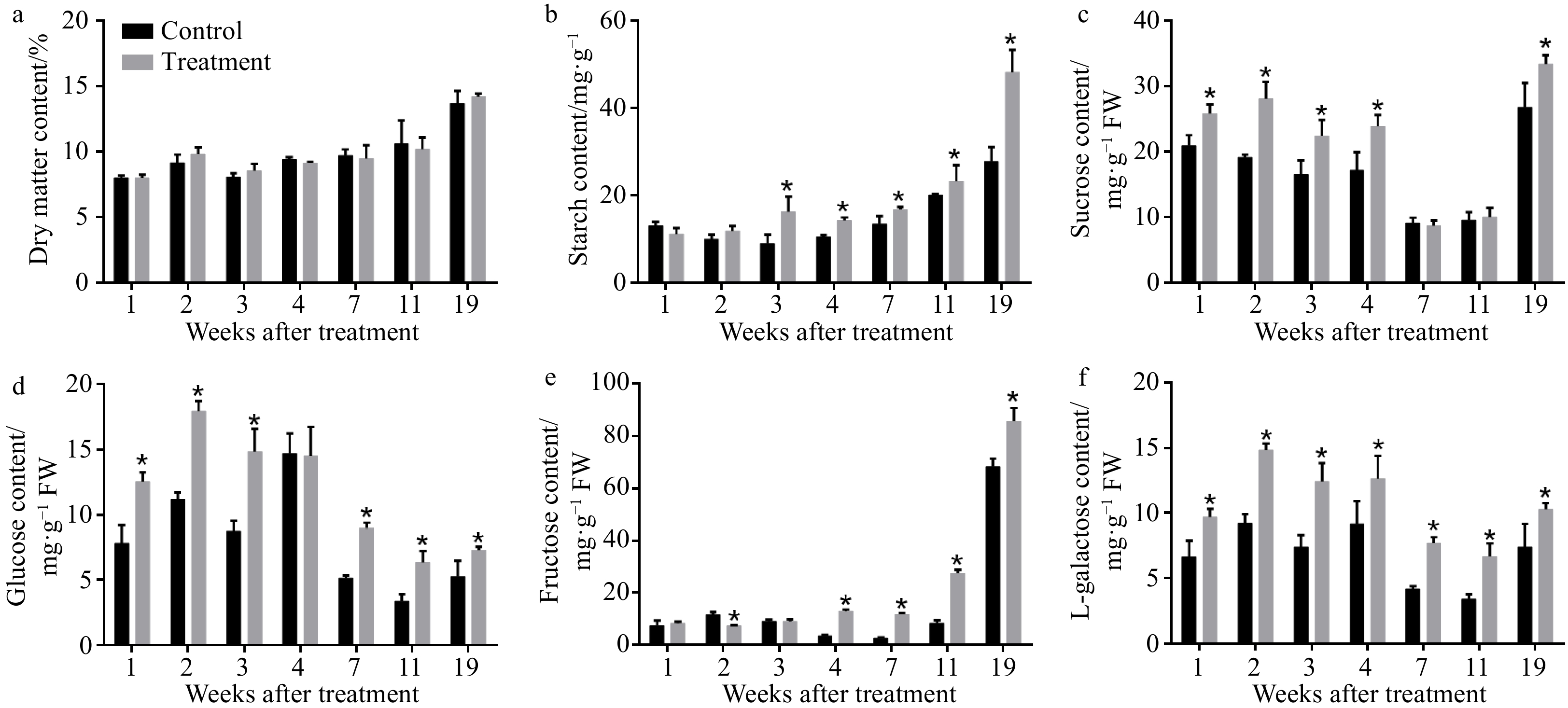
Figure 2. Effects of CPPU on (a) dry matter, (b) starch, (c) sucrose, (d) glucose, (e) fructose, and (f) L-galactose concentrations of 'Hayward' kiwifruit. * Indicates significant difference between the treatment and the control based on t-test (p < 0.05).
CPPU treatment affected enzyme activities related to sugar metabolism in 'Hayward' kiwifruit
-
The application of exogenous CPPU enhanced the activity of sucrose synthase (SUSY) (Fig. 3a), acid invertase (AINV) (Fig. 3b) and sucrose phosphate synthase (SPS) (Fig. 3c) in early stages of fruit development. The activity of AINV was much higher than that of SUSY and SPS at 28 to 42 DAB.
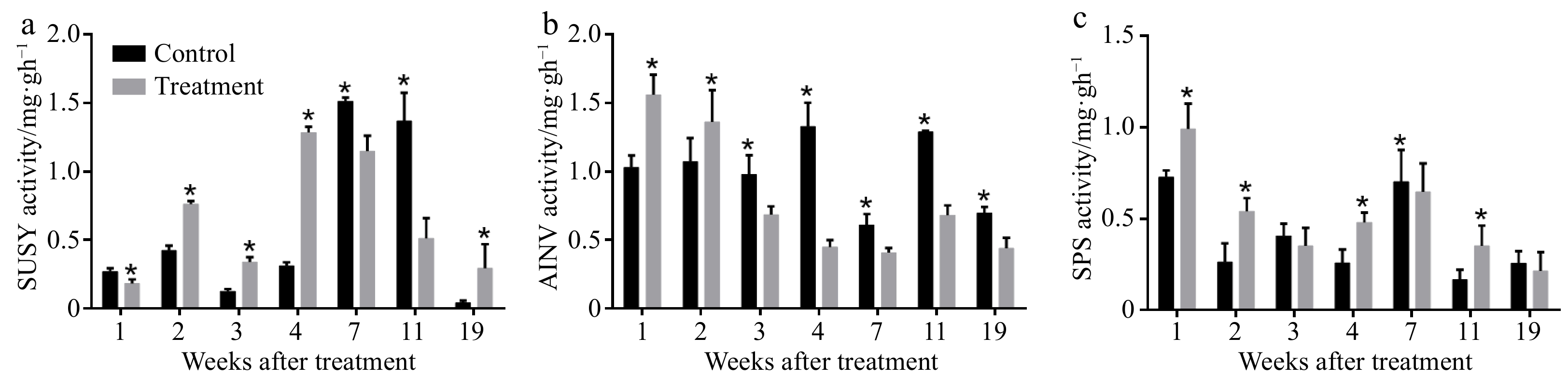
Figure 3. Effects of CPPU on (a) sucrose synthase (SUSY), (b) acid invertase (AINV), and (c) sucrose phosphate synthase (SPS) activity in 'Hayward' kiwifruit. * Indicates significant difference between the treatment and the control based on t-test (p < 0.05).
CPPU treatment affected endogenous hormone content of 'Hayward' kiwifruit
-
Hormones that regulate fruit growth by controlling cell division and cell expansion have been elucidated, including auxin, cytokinin and gibberellin[5]. To examine the effect of CPPU treatment on endogenous hormones in fruits, we measured the content of cytokinin (zeatin, ZR), gibberellin (gibberellic acid 3, GA3), abscisic acid (ABA) and auxin (IAA) in both treated- and untreated-fruits. From 28 to 49 DAB, the content of IAA and ZR in the treatment and control groups as well as the content of ABA in the control group showed an increasing trend. No significant difference was detected in IAA content between treatment and control groups from 28 to 42 DAB (Fig. 4a). At 49 DAB, the IAA content of the CPPU-treated fruits was reduced. In addition, CPPU treatment reduced the GA content in the fruit (Fig. 4b). From 28 to 35 DAB, exogenous CPPU treatment increased the endogenous ZR content (Fig. 4c). However, at 49 DAB, the endogenous ZR content of CPPU-treated fruits was lower than the control. CPPU treatment reduced the ABA content in fruits, especially at 42 to 49 DAB (Fig. 4d). Overall, at 28 to 42 DAB, exogenous CPPU treatment increased the content of endogenous IAA and cytokinin and decreased the content of GA and ABA compared to the untreated controls.
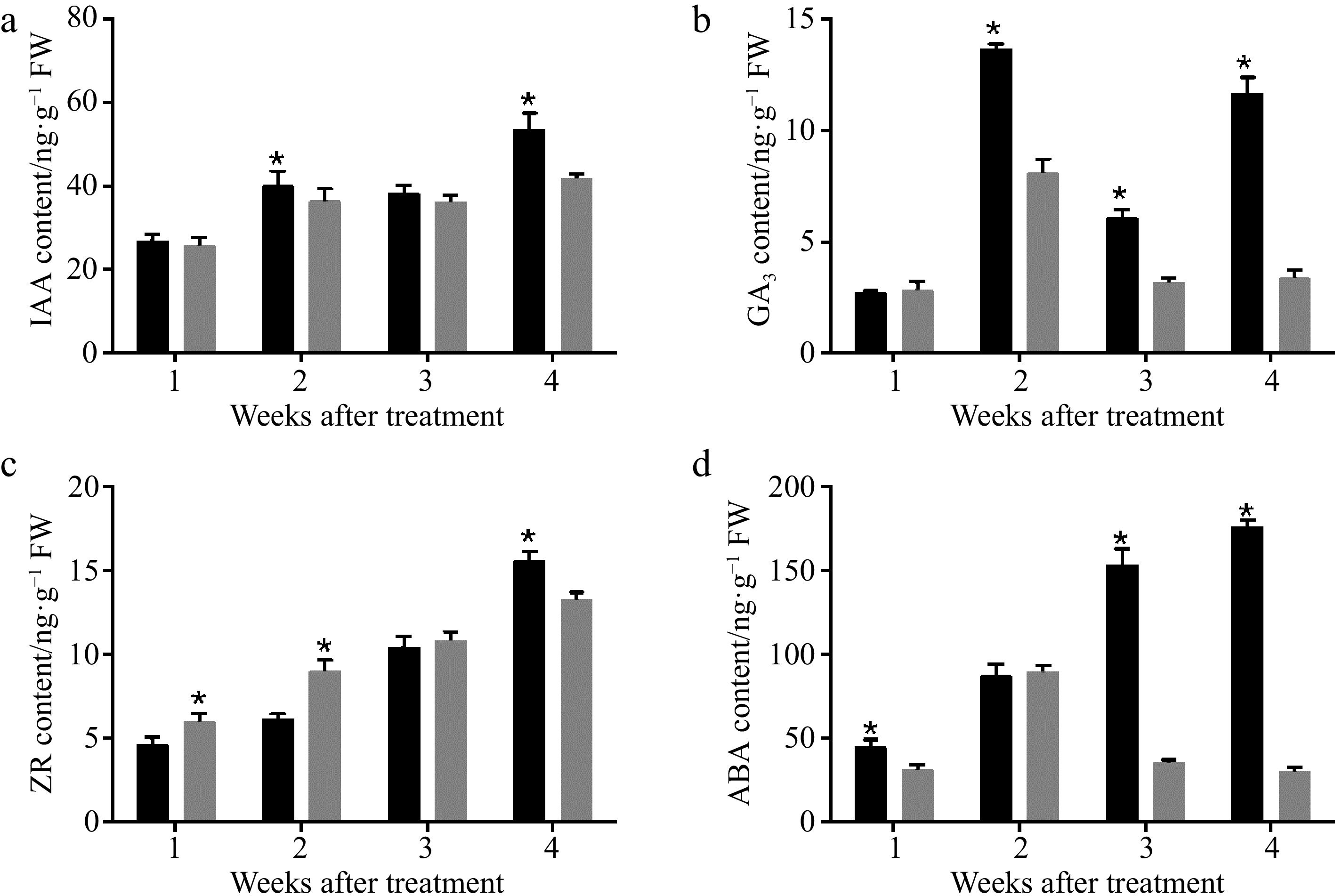
Figure 4. Effects of CPPU on the levels of (a) auxin (IAA), (b) gibberellin (GA3), (c) cytokinin (zeatin, ZR), and (d) abscisic acid (ABA) in 'Hayward' kiwifruit. * Indicates significant difference between the treatment and the control based on t-test (p < 0.05).
Identification and analysis of DEGs in early stages of kiwifruit development
-
RNA-seq was performed to investigate the genes affected by CPPU treatment during the fastest stages of 'Hayward' kiwifruit development. High-throughput sequencing was conducted in kiwifruit samples at 28, 35, and 42 DAB, which represent the period of the most growth of kiwifruits, with three biological replicates at each time point (Fig. 5). Genes with Fold Change > 1.5 and FDR < 0.05 were considered differentially expressed. All genes were compared between the CPPU-treated kiwifruit and the control, and 3,066 differently expressed genes (DEGs) were discovered, of which 1,135 were not assigned a potential function or identity. DEGs between B1/A1 (28 DAB CPPU-treatment/28 DAB control), B2/A2 (35 DAB CPPU-treatment/35 DAB control) and B3/A3 (42 DAB CPPU-treatment/42 DAB control) were obtained. The pair-wise comparison B1/A1 had the largest number of DEGs. To verify the RNA-seq results, nine related and/or interesting genes were selected for qRT-PCR analysis (Supplemental Fig. S1). Functional classification was assigned based on data from multiple databases.

Figure 5. Identification of Differentially Expressed Genes (DEGs) in kiwifruits in response to CPPU treatment. (a) Overall distribution of sample gene expression. (b) A volcano plot of DEGs. (c) Vinn diagrams of DEGs between B1/A1 (28 DAB CPPU-treatment/28 DAB control), B2/A2 (35 DAB CPPU-treatment/35 DAB control) and B3/A3 (42 DAB CPPU-treatment/42 DAB control).
Annotation and functional characterization
-
Twelve clusters with different expression patterns were identified by hierarchical clustering analysis of the screened DEGs (Fig. 6a), in which cluster 1 represented DEGS that were up-regulated over time with CPPU treatment, and cluster 4 represented the DEGs down-regulated at all three time points with CPPU treatment. The functions of DEGs in cluster 1 are mainly related to solute transport, RNA biosynthesis and enzyme functions. In cluster 4, the predicted functions of the DEGs were mainly related to phytohormone action, cell wall organization, solute transport and enzymes. Cluster 3 (K3) contained the most DEGs, which were down-regulated at 28 DAB with CPPU treatment but showed no significant changes at 35 and 42 DAB. The functions of DEGs in cluster 3 mainly included photosynthesis, protein modification, solute transport, RNA biosynthesis, cell cycle organization, cytoskeleton organization and enzyme.
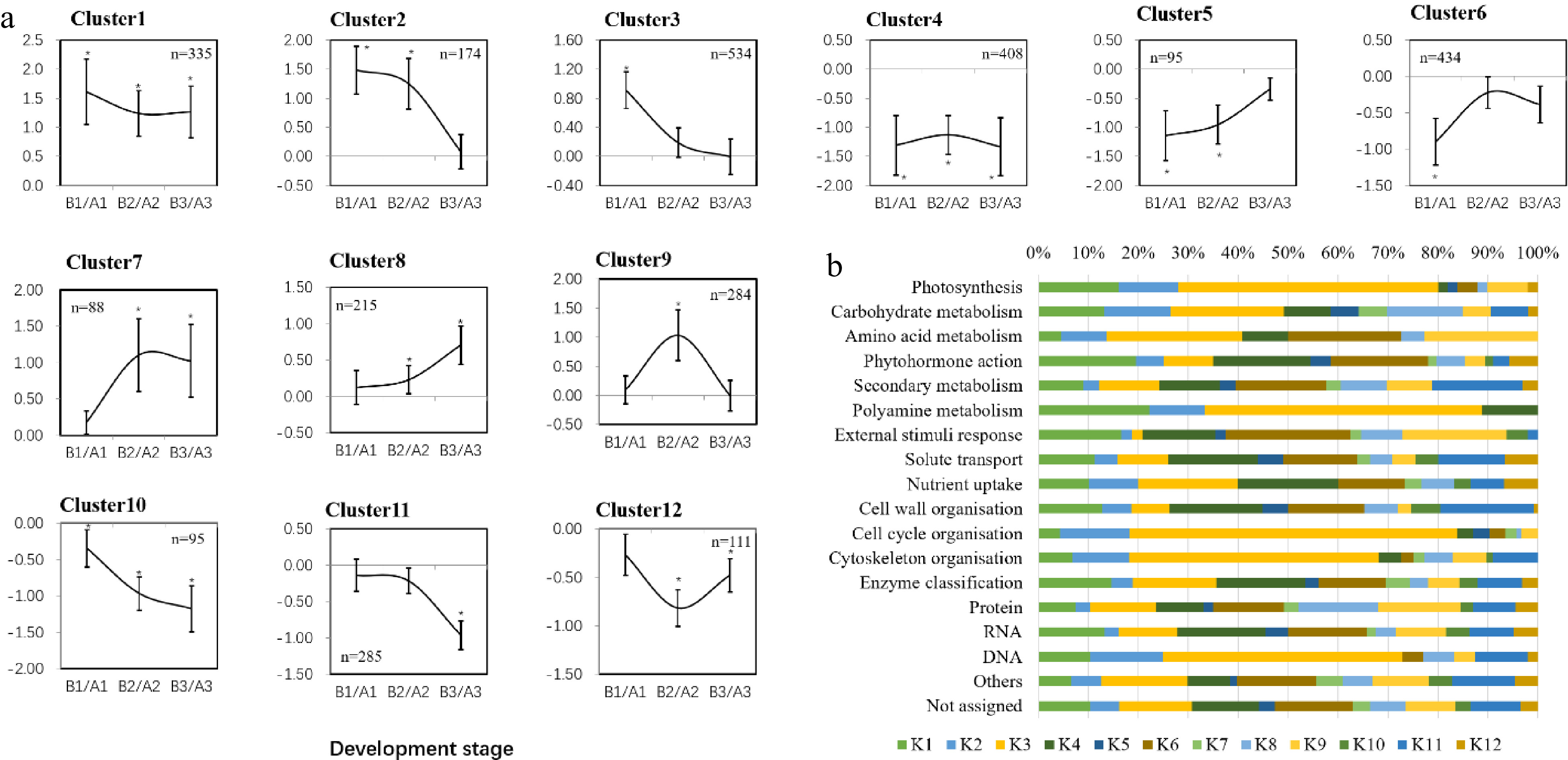
Figure 6. Different gene expression patterns and enriched functional categories. (a) Main expression profiles trends. Twelve main patterns were identified from the differentially expressed genes. The y-axis shows the log2 fold changes of CPPU treatment/control at the same sampled point. Error bars indicate standard deviation. (b) Functional category enrichment (modified MapMan bins) of upregulated and downregulated genes among the different expression patterns.
Expression of genes related to sugar metabolism and soluble sugar transport in CPPU-treated fruits
-
Transcriptome analysis showed that CPPU application led to changes in the abundance of genes involved in carbohydrate metabolism and sugar transport during the early stages of kiwifruit development (Fig. 7).

Figure 7. Major carbohydrate metabolism pathways and carbohydrate accumulation that showed changes in kiwifruit treated with CPPU. The dynamics of gene expression during the three periods (from 28 to 42 DAB) are represented by a group of three boxes, with colors representing up-regulation (red) or down-regulation (blue) with CPPU treatment. Fru, fructose; Glc, glucose; CWINV, cell wall invertase; SUSY, sucrose synthase; CIN, cytoplasmic invertase; VIN, vacuolar invertase; UDPG, UDP-D-glucose; UGP, UDPG-pyrophosphorylase; G1P, fructose-1-phosphate; PGM, phosphoglucomutase; FK, fructokinase; HK, hexokinase; F6P, fructose 6-phosphate; G6P, glucose 6-phosphate; PGI, phosphoglucose isomerase; PFK, phosphofructokinase; F1,6P, Fructose-1,6-diphosphate; AGPase, ADP-glucose pyrophosphorylase; GBSS, granule-bound starch synthase; SBE, starch branching enzyme; BMY, β-amylase; AMY, α-amylase; PHS, α-glucan phosphorylase; Mal, maltose; SUT, sucrose transporter; SWEET, sugar transporter; HT, hexose transporter; vGT, vacuolar glucose transporter; TST, tonoplast sugar transporter; EDR6, sugar transporter; PGPT, glucose 6-phosphate transmembrane transporter.
CPPU treatment affected the changes of starch metabolism genes. After CPPU treatment, a few genes related to starch synthesis, APL1 (ADP-glucose pyrophosphorylase) in cluster 2 and APL2 and APL4 in cluster 3, were up-regulated. A granule-bound starch synthase gene GBSS1 and two beta-amylase genes, BAM3 and BAM9, in cluster 4 were also up-regulated. Two alpha-amylase genes (psr99540 and pss31212) were down-regulated, while two alpha-glucan phosphorylase genes (pss30499 and psr86180) were up-regulated (Fig. 7).
CPPU treatment also affected the expression of genes related to sucrose degradation (Supplemental Table S2). The transcript levels of VIN1 (vacuolar invertase) and SUSY1 (sucrose synthase) increased. Besides, two alkaline sucrose-specific invertase genes, including CIN2, were down-regulated after CPPU treatment. Furthermore, two hexokinase genes (psr95417 and psr86807) were significantly up-regulated. At 28 DAB, the transcript level of FK3 was increased. In addition, we found that the expression of SUT1 was significantly up-regulated, and the expression of SUT2 was significantly down-regulated (Fig. 7). Changes in transcript abundance indicate that CPPU treatment increased the expression of genes related to starch synthesis and sucrose degradation.
CPPU application affected gene expression of cell cycle organization, cell wall organization and phytohormone action
-
Hormones such as auxin, gibberellin and cytokinin play important roles in regulating the growth and development of fruits, especially during early fruit growth[4,5]. Significant changes were seen in hormone content and gene expression related to hormone metabolism and signal transduction during the period of rapid fruit development response to CPPU treatment (Supplemental Table S3). Exogenous CPPU treatment reduced the cytokinin content within kiwifruit during early fruit development, corresponding to a decrease in the expression of genes associated with cytokinin synthesis and an increase in genes related to cytokinin degradation, such as LOG1 and CKX2. The content of gibberellin and abscisic acid as well as the expression of synthesis-related genes such as GA3OX1 and NCED3 showed decreasing trends. In addition, CPPU treatment affected the expression of genes related to hormone signaling pathways within the fruit. Genes in the cytokinin and abscisic acid signaling pathways, such as AHK4, ARR9 and PYL4-like, were increased, while the expression of genes related to gibberellin and auxin signaling decreased. The gibberellin signal transducer DELLA1 was down-regulated in expression at 28 DAB, The genes TIR1 and IAA26 in the auxin signaling pathway were inhibited.
Plant hormones have been shown to promote fruit enlargement by regulating cell division and elongation during fruit development. Analysis of the RNA-seq results showed that the expression of genes related to cell division and cell wall organization were up-regulated after CPPU treatment during the early stages of fruit development (Supplemental Table S4). Twelve cyclin genes, like CYCU2 and CYCA1, were up-regulated 1- to 2-fold at 28 DAB. In addition, the expression levels of CDKB1 and CDC2C, which play key roles in the control of the cell cycle, were all increased. Interestingly, the expression of expansin genes, such as EXPA8 and EXPA10, showed decreasing trends.
-
CPPU is commonly used to improve the yield of horticultural crops such as grapes, pears and kiwifruits[20−26]. Consistent with previous studies[27], the application of CPPU was found to increase the fresh weight and size of kiwifruit in our experiments. Substantial weight gains in the treatment group were observed at 28 and 35 DAB with CPPU application. Until harvest, the fresh weight of kiwifruit from treated fruit increased by 64% compared to the control group. We found that CPPU contributed the most to fruit weight growth at 28 to 45 DAB, i.e. 1 to 3 weeks after treatment, which is consistent with the study of Nardozza et al.[34]. Based on this, we performed transcriptome analysis during the fastest growing period of kiwifruit.
Fruit growth and development are inseparable from adequate energy supply and regulation by hormones[4]. The growth of fruit is mainly determined by the number and size of the cells in the tissue. However, cell division and expansion cannot be achieved without the supply of sugar. During the early stage of fruit development, most of the carbon transported from the source organ is metabolized through glycolysis and tricarboxylic acid cycle (TCA). As the development of the fruit slows down sugar metabolism, soluble sugar slowly accumulates, and the excess sugar is converted into starch for storage[35]. Sucrose metabolism plays a key role in fruit development and yield, mainly by producing a range of sugars used either as metabolites to promote growth and to synthesize essential compounds (including protein, cellulose and starch) or as signals to regulate microRNA expression, transcription factors and other genes as well as hormonal signaling[36−39]. In our study, CPPU treatment accelerated sugar metabolism and promoted starch accumulation in kiwifruit. Sucrose synthase and invertase are key enzymes in the cleavage of sucrose in cells, and sucrose synthase is considered to be the key enzyme affecting sink strength[36]. Transcriptomic data observed an upregulation of SUSY1 and NINV gene expression after CPPU treatment, which was consistent with the increase in their metabolites. Up-regulation of FK and HK gene expression was also observed. The increase in sucrose and glucose content, accompanied by no difference in fructose content, may indicate that more fructose was catabolized and converted to G-6-P to enter the tricarboxylic acid cycle, which provides the carbon skeleton and energy for fruit growth, leading to fruit enlargement. In addition, the activity of NINV was higher than that of SUSY, which may indicate more sucrose accumulation toward the vesicles. Moreover, the expression of enzymes related to starch synthesis, such as AGPase, GBSS, SBE, BMY, and AMY, were significantly up-regulated during fruit development, and a gradual increase in early starch content was observed, suggesting that CPPU promotes starch accumulation in early fruit development. We therefore hypothesized that CPPU treatment increased the distribution of carbohydrates to organs, increased fruit strength, and provided sufficient carbon skeleton and energy for cell division and expansion.
Hormones are generally considered to play roles in fruit development and ripening, and can interact with each other to regulate fruit growth[5], especially auxin, cytokinin and gibberellin. In early fruit development, the role of auxin is to promote cell division and cell expansion, and it is thought to act together with gibberellin to influence cell enlargement during fruit development[40]. During Arabidopsis fruit development, auxin activates gibberellin signaling and induces gibberellin synthesis[41]. In our experiments, we found that CPPU treatment affected not only the content of hormones within the fruit but also the expression of genes involved in hormone synthesis and signal transduction pathways. After CPPU treatment, the content of auxin and gibberellin decreased. In the meantime, genes in auxin and gibberellin signal transduction were down-regulated. The polar transport of auxin also plays an important role in regulating plant growth and development, and the efflux of auxin in plants is mainly controlled by PIN-FORMED (PIN) family proteins[42]. The expression levels of genes related to auxin metabolism, like the PIN1C transporter and GH3.3, an indole-3-acetic acid amide synthase gene, were decreased. In addition, the expression of TIR1 and Aux/IAA genes in the auxin signaling pathway was downregulated. The expression of gibberellin 3-oxidase (GA3ox), a key gene for GA3 synthesis, and the negative regulator DELLA, in the GA signal transduction pathway, were also down-regulated. Previous studies demonstrated that treatment with exogenous CPPU reduced the content of endogenous cytokinin during early fruit growth[43]. In our study, we observed a decrease in cytokinin (CTK) levels until 49 DAB, and CTK biosynthesis genes like LOG1 were down-regulated at 28 to 42 DAB. CTK signal transduction genes like AHK4, ARR9 and ARR12 and degradation genes like CKX2 and CKX3 were up-regulated. This result illustrates that endogenous cytokinin synthesis was reduced in fruit after CPPU treatment, but cytokinin content inside the fruit was increased and CPPU treatment enhanced cytokinin signaling. Lewis et al.[43] showed that the synthetic cytokinin CPPU promoted fruit development, but the endogenous cytokinin concentration was decreased. During kiwifruit fruit development, the cell division period is before 45 d, followed by the cell expansion period. The main roles of auxin and cytokinin in this period are to promote cell elongation[5]. But the decrease of auxin and gibberellin content, consistent with the down-regulation of the expression of auxin and gibberellin signaling genes, indicated that CPPU treatment may have inhibited cell elongation in early fruits. The increase in cytokinin content and up-regulation of gene expression for the cytokinin signaling pathway indicated that CPPU treatment promoted cell division in fruits during rapid development. This is consistent with an increase in cyclin gene expression and a decrease in cell expansion-related genes in the transcriptome data(Fig. 8).
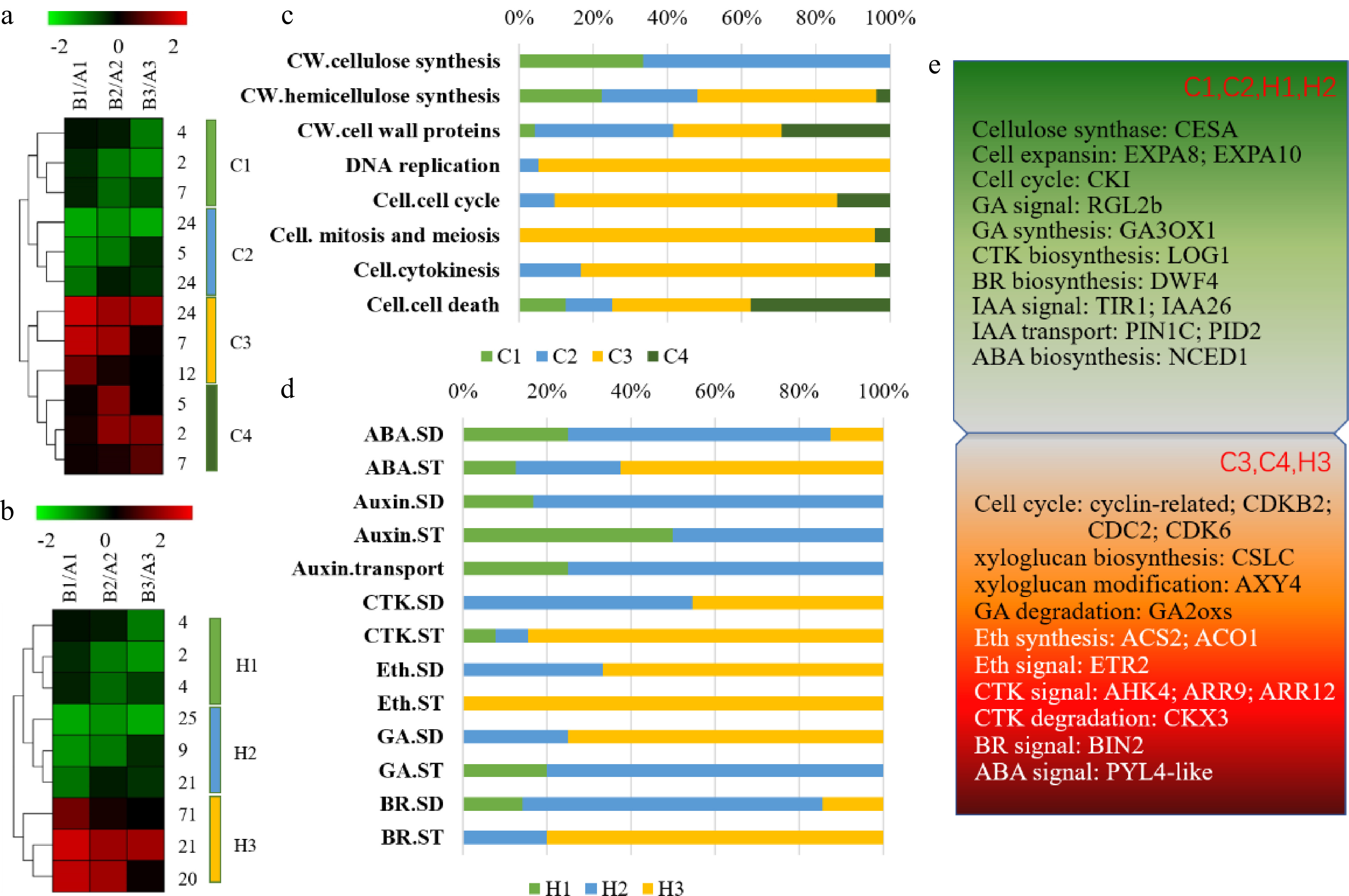
Figure 8. Expression profiles of genes associated with fruit development and growth, including genes involved in cell growth and cell wall synthesis, as well as hormone signaling. (a) Heat map of genes involved in cell and cell wall pathways. Genes that varied significantly along the three developmental stages (28, 35, 42 DAB) clustered into four patterns (C1−C4) based on 12 K-means clusters. (b) Heat map of the genes involved in hormone pathways. Expression patterns along the three developmental stages (28, 35, 42 DAB) clustered into three patterns (H1−H3) based on 9 K-means clusters. (c), (d) Distribution of genes among groups C1−C4 and H1−H3, respectively. (e) Trends in expression abundance of representative genes from 28 to 45 DAB. CW, Cell wall; SD: synthesis-degradation; ST, signal transduction.
-
CPPU treatment increased internal cytokinin content but decreased endogenous cytokinin synthesis and reduced auxin and gibberellin content in early fruit development. In addition, the up-regulation of gene expression related to cytokinin signaling and the down-regulation of gene expression related to auxin and gibberellin signaling indicated that CPPU treatment enhanced the effect of cytokinin in early fruit development. Consistent with this result, transcriptomic data showed upregulation of cell cycle-related genes at 28 to 45 DAB, particularly at 28 DAB, and downregulation of cell expansion-related genes. The CPPU treatment also accelerated the allocation of carbohydrates to the fruit. The increase in sucrose and glucose content, but not in fructose content, at the early stage indicated that more fructose was metabolized and then entered the tricarboxylic acid cycle and other pathways, thus generating more energy to support physiological activities such as cell division.
- This work was supported by the Program for the National Natural Science Foundation of China (No. 31901970). We thank the Horticulture Science Research Center at the College of Horticulture, NWAFU for their technical support in this work.
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 Oligonucleotide primers used in this study.
- Supplemental Table S2 Genes related to sucrose degradation.
- Supplemental Table S3 Genes related to hromone metabolism and signalling.
- Supplemental Table S4 Genes related to cell cycle.
- Supplemental Fig. S1 qRT-PCR test of differential genes. * Indicates significant difference between the treatment and the control based on t-test (p < 0.05).
- Copyright: © 2022 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen X, Zhu L, Song J, Zhang J, Li M. 2022. Transcriptomics reveals the molecular mechanism underlying kiwifruit expansion following forchlorfenuron (CPPU) treatment. Fruit Research 2: 4 doi: 10.48130/FruRes-2022-0004

Transcriptomics reveals the molecular mechanism underlying kiwifruit expansion following forchlorfenuron (CPPU) treatment
- Received: 19 December 2021
- Accepted: 22 February 2022
- Published online: 21 March 2022
Abstract: Kiwifruit is an economically and nutritionally valuable fruit. Forchlorfenuron (CPPU) is widely used to improve fruit set and yield of kiwifruit. However, the mechanism through which CPPU regulates fruit enlargement remains unclear. Physiological and RNA-seq data revealed that the greatest contribution of CPPU treatment to fruit growth occurred at 28 to 42 d after bloom (DAB). CPPU application accelerated sugar metabolism by increasing the content of sucrose through increasing the transcripts of genes such as SUSY, HK, and FK in sucrose metabolism. In the early stages of fruit development, up-regulation of the starch synthesis gene AGPase and the decomposition gene β-AMY indicated the occurrence of starch turnover. After CPPU treatment, genes related to the synthesis and signal transduction of auxin and gibberellins were down-regulated. During the early fruit developmental stage, the cytokinin content and the expression of signal transduction genes were increased. In addition, the expression of cyclin genes, related to cell division, was up-regulated within three weeks after CPPU treatment. This indicates that CPPU treatment promoted cell division during the fastest growth stage, which would lead to an increase in cell number and further increases in fruit size.
-
Key words:
- Kiwifruit /
- Forchlorfenuron /
- Fruit development /
- Sugar metabolism /
- Hormone.