









-
Macadamia, also known as Hawaii nut, belongs to the family Proteaceae, and M. integrifolia F. Mull., an evergreen tree without taproots and a shallow root system, is suitable for subtropical climate, and is classified as a subtropical fruit tree[1]. Macadamia nut contains more than 70% fat, 9% protein and eight essential amino acids; thus, its crispy kernels have abundant unsaturated fatty acids with high nutrition and health value[2−4]. Long-term consumption of macadamia nuts, which is known as the 'queen of dried fruits', contributes to the prevention and treatment of cardiovascular diseases[5,6]. In addition, the kernel, shell, and oil meal of macadamia are processed into various drinks, oil[7], activated carbon[8], adsorbent, feed, and other products. Furthermore, macadamia trees are beautifully shaped, with dense branches and foliage, pretty and fragrant flowers, and solid and compact wood, and are ideal insect-resistant landscaping plants.
Macadamia is endemic to the subtropical rainforests of eastern Australia and is one of the few crops to have been domesticated from dicotyledons[9]. The plant is concentrated in Australia, USA, Kenya, South Africa, Costa Rica, Guatemala, Brazil, and other tropical countries[10]. Macadamia is one of the few rapidly expanding crops in USA, Australia, South Africa, New Zealand, and China. Over 500 selected landraces and varieties are present. The macadamia germplasm in the world ranges from 34° N to 34° S and involves more than 20 countries, but most commercial production areas are located in 16°–24° N, and the main production countries are China, Australia, South Africa, USA, and Kenya. In 2020, Macadamia had a global cultivation area of 6.05 million acres, with the largest cultivation area of 4.7 million acres in China. Thus, China has the highest macadamia production in the world.
Macadamia is highly heterozygous and predominantly outcrossing. Its genome size is 896 Mb, and all cultivars are diploid[11−14]. Study of its origin and evolutionary history requires a high-quality macadamia genome, and two macadamia genome research studies have been reported. The release of the HAES 741 macadamia genome opened a new era of macadamia genomics and was an important milestone in macadamia research. In 2016, Nock et al. used Illumina short-read sequence data to construct a highly fragmented genome sketch of HAES 741 with a total length of 518 Mb, 3,522-bp contig N50, and 4,745-bp scaffold N50, which annotated 35,337 protein-coding genes[15]. In 2020, they reassembled the HAES 741 genome using a combined strategy of Illumina short read sequences and PacBio long read sequences, with a 745-Mb genome and 413-kb scaffold N50. After fixing scaffolds to 14 chromosomes using seven genetic linkage maps, 34,274 protein-coding genes were predicted[12]. The studies of Ming et al.[16] and Nock et al.[12] used the world-famous macadamia nut variety Kau (Macadamia integrifolia Maiden & Betche) as the material and used three-generation sequencing to complete the genome assembly . The genome size is 794 Mb, contig N50 281 kb and they annotated 37,728 genes. They found the gene families related to oil synthesis and fruit shell development in the macadamia genome have expanded. Then they selected 112 representative cultivated and wild macadamia materials from Hawaii and Australia to resequence, and the results revealed a phylogenetic relationship of macadamia populations. Through 2−3 generations of artificial selection, selective clearance areas are generated, which provides genomic basis and direct evidence for the 'one-step' theory of asexual crop domestication. These findings provide a theoretical basis for the rapid domestication of new species[16]. At present, two studies on the macadamia genome have been reported, and both are for the same species. It is also the only genome in macadamia, falling short of the genome resources required to study the plant.
GDSL esterase/lipase protein (GELP) is a hydrolase that hydrolyzes thioesters, aryl esters, phospholipids, amino acids, and other substrates. This type of lipase is widely present in prokaryotes and eukaryotes. At present, GELP is widely involved in physiological activities such as normal plant growth and development, organ morphogenesis, secondary metabolism and stress, and plays an important role in oil metabolism of oil crop seeds[17,18]. Although GELP family genes have been identified and studied in other plants, they have not been identified in macadamia.
GUIRE 1 (GR1), an excellent cultivar bred in China, has evident advantages such as early fruit setting, high and stable yield, high quality, and strong stress resistance. The high-quality GR1 assembled genome described in this study will help in seed selection and breeding of macadamia and accelerate research on the molecular mechanisms in macadamia, thereby providing a database for macadamia researchers.
-
The genome of GR1, a highly heterozygous cultivar, was assembled from the beginning using 281 Gb long-read and long-sequence data obtained by nanopore sequencing. The total genome size was about 924 Mb, including 1,757 contigs, and the contig N50 and GC content were 1.97 Mb and 39.28%, respectively. Busco's evaluation of genome assembly quality and integrity showed that 95.7% of plant single-copy homologous genes were complete, and 74.3% of complete single-copy and 21.4% of multi-copy genes were complete. The preliminarily assembled macadamia genome had high quality and coverage. Using the macadamia genome published by Nock et al.[12] as the reference genome, the initially assembled GR1 genome was constructed to the chromosomal level, resulting in a super high-density genome map with a 792-Mb genome size, 46.24-Mb scaffold N50, and 14 chromosomes (Fig. 1; Table 1). The size of the GR1 genome we finally assembled is consistent with that of Kau (Supplemental Table 1). BUSCO was used to assess the quality and integrity of genome assembly, and 89.7% of plant single-copy homologous genes were intact. Complete single- and multi-copy genes accounted for 73% and 16.7%, respectively.
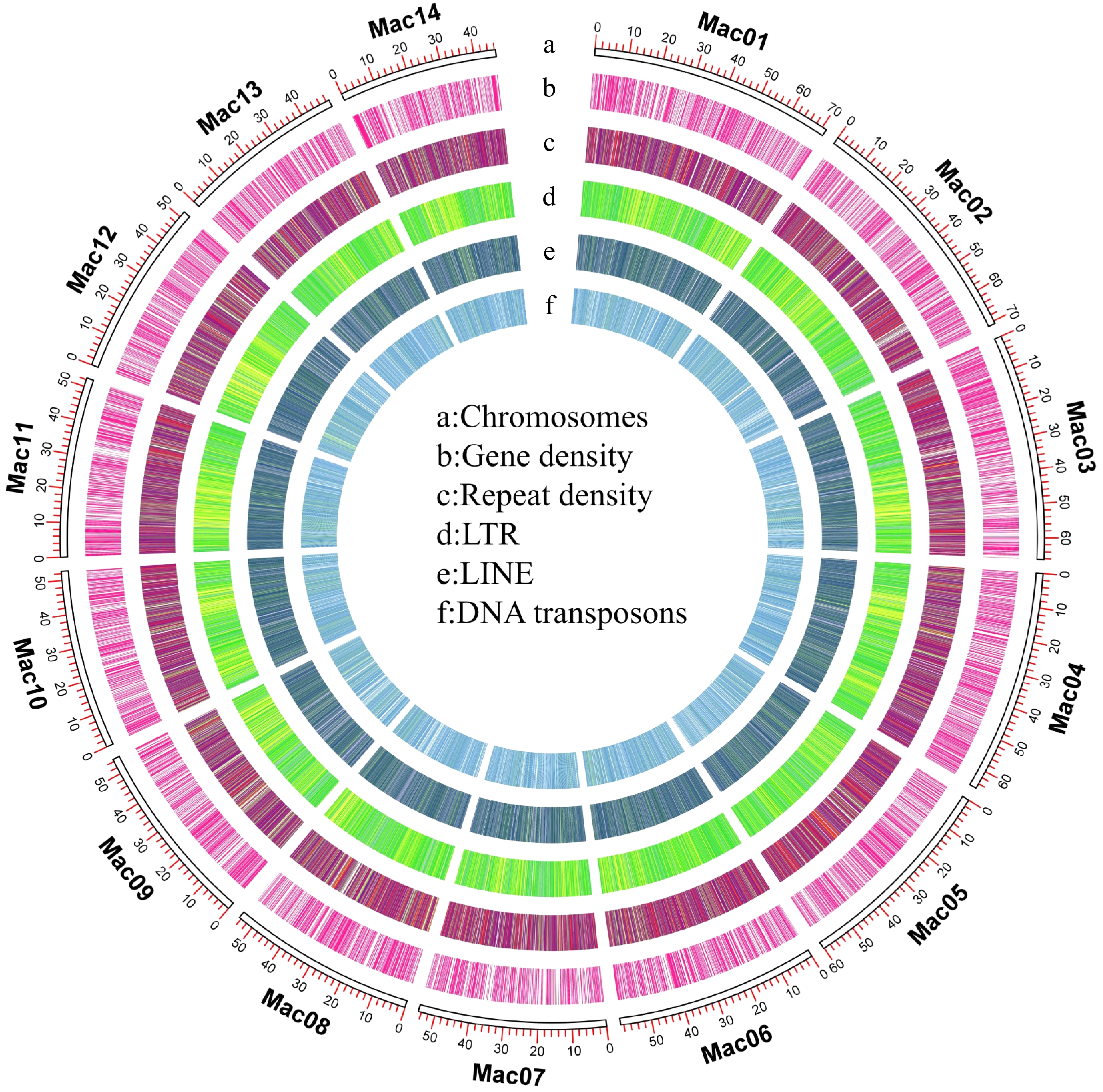
Figure 1.
Overview of the macadamia genome. (a) Chromosomes, (b) gene density, (c) repeat density, (d) long terminal repeats (LTRs), (e) long interspersed nuclear elements (LINEs), (f) DNA transposons.
Table 1. GR1 genome assembly statistics.
Statistic Contig length
(bp)Contig no. Scaffold length
(bp)Scaffold no. N50 1,971,883 124 55,868,876 7 N80 494,409 388 50,524,966 11 N90 166,444 729 47,725,559 13 Longest 30,625,757 − 77,511,179 − Total 924,331,529 1,757 792,388,653 14 Genome annotation
-
The repetitive sequences of the GR1 genome were obtained by homologous and de novo annotations to understand the genomic characteristics of GR1. About 56.77% of the genome was identified as duplicate regions. Among these regions, long terminal repeats (189 Mb, 23.96% of the genome) were identified as the main repeat sequences, followed by long interspersed nuclear elements (44 Mb, 5.63% of the genome) (Supplemental Table 2). A total of 2,543 tRNAs, 1,119 rRNAs, 248 microRNAs, and 1,562 snRNAs as non-coding genes were present in the GR1 genome. MAKER software combined with the AUGUSTUS pipeline ab initio gene prediction was used for protein-coding gene prediction, and proteins were compared to perform gene functional annotation for GR1. Finally, 37,657 protein-coding genes were obtained, which are consistent with the number of protein coding genes contained in 'Kau'. (Supplemental Table 3).
Expansion and contraction of gene families
-
Protein sequences of macadamia and 11 other species of the same family were compared to evaluate expansion and contraction in the macadamia gene family. These species were Telopea speciosissima (Proteaceae), dicotyledons: Arabidopsis thaliana, Coffea arabica, and Morus alba; monocotyledons: Setaria italica, Oryza sativa, Zea mays, Elaeis guineensis, Ananas comosus, and Nelumbo nucifera from the same family (Proteaceae) were used for gene family clustering, with Amborella trichopoda as the outgroup. OrthoVenn2 (
https://orthovenn2.bioinfotoolkits.net/home ) was used to identify gene families unique to all species. In macadamia, 37,657 genes were divided into 14,930 gene families, and 13,613 single-copy gene families were present.A phylogenetic tree was constructed using the single-copy genes of these single-copy gene families of the 12 species, with Amborella trichopoda as the outgroup. Evolutionarily, macadamia diverged from waratah, a member of the same family about 71.74 million years ago. Proteaceae diverged from Nelumbonaceae nearly 115.37 million years ago. Proteaceae diverged from Rubiaceae about 140 million years ago (Fig. 2a). Expansion or contraction of the gene family is an important feature in selective evolution. Compared with Nelumbo nucifera of the same family, macadamia evolution involved new genes and gene families, but their evolution occurred independently with a different degree of gene families being lost for each species. In macadamia, 1,498 gene families experienced expansion, and 5,327 gene families experienced contraction (Fig. 2a).
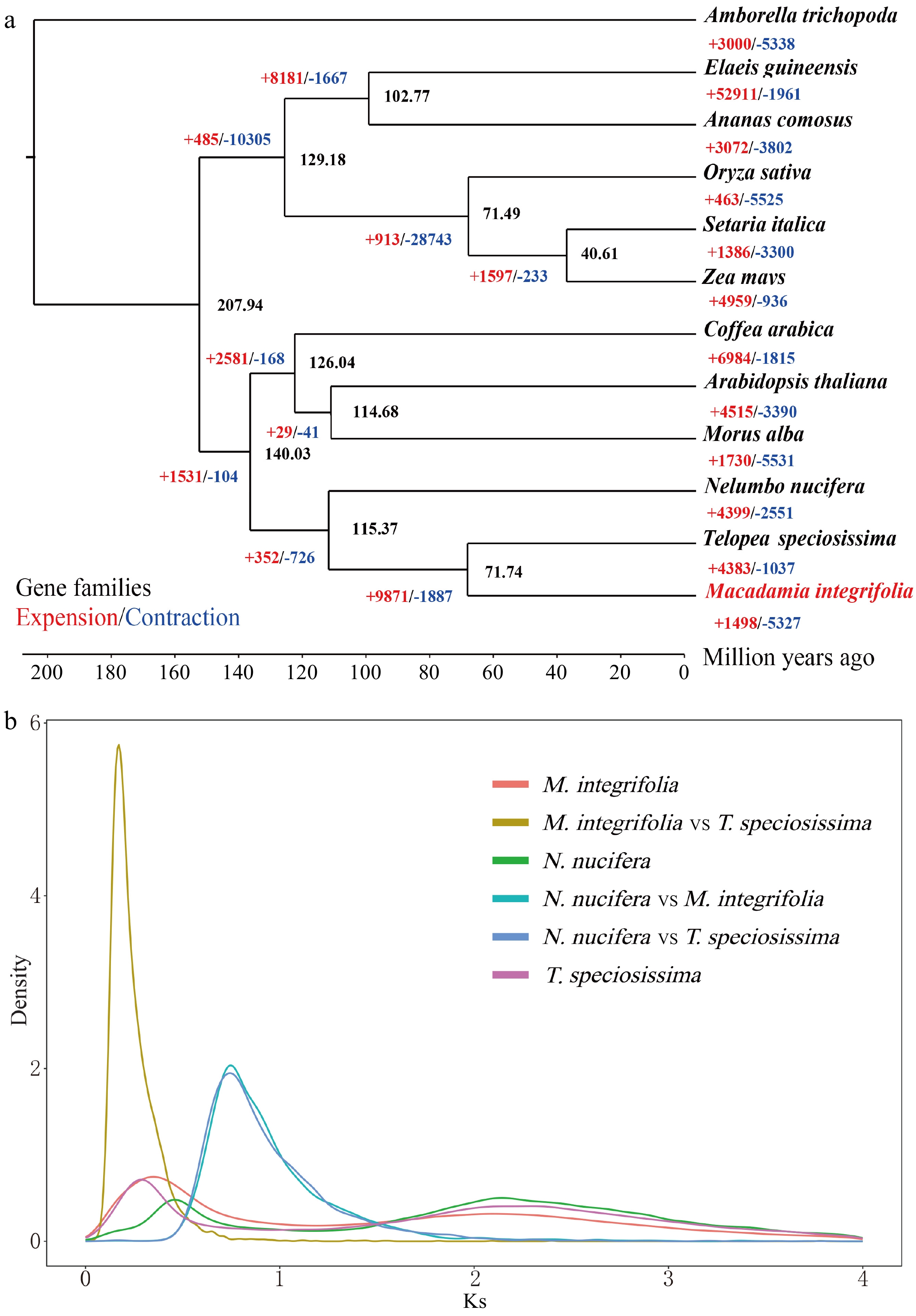
Figure 2.
Genome evolution analysis. (a) Phylogenetic tree of 12 species constructed using single-copy orthologs. (b) Frequency distribution of synonymous substitution rates (Ks) between homologous gene pairs in the syntenic blocks of M. integrifolia, M. integrifolia vs T. speciosissima, N. nucifera, N. nucifera vs M. integrifolia, N. nucifera vs T. speciosissima, and T. speciosissima.
Genome-wide replication and differentiation in macadamia
-
On the basis of the expansion and contraction of gene families, the synonymous substitution rates (Ks) of the homologous genes of macadamia, waratah, and A. thaliana were calculated, and Ks of the aligned homologous genes of different species was calculated to obtain the Ks curve of the genome and infer the WGD[19] event of macadamia. An evident peak between macadamia and waratah at Ks value of 0.34 indicated that a WGD event occurred in macadamia about 41 million years ago, and this WGD event might have been a doubling event shared by the Proteales (Fig. 2b).
Comparative genomic analysis
-
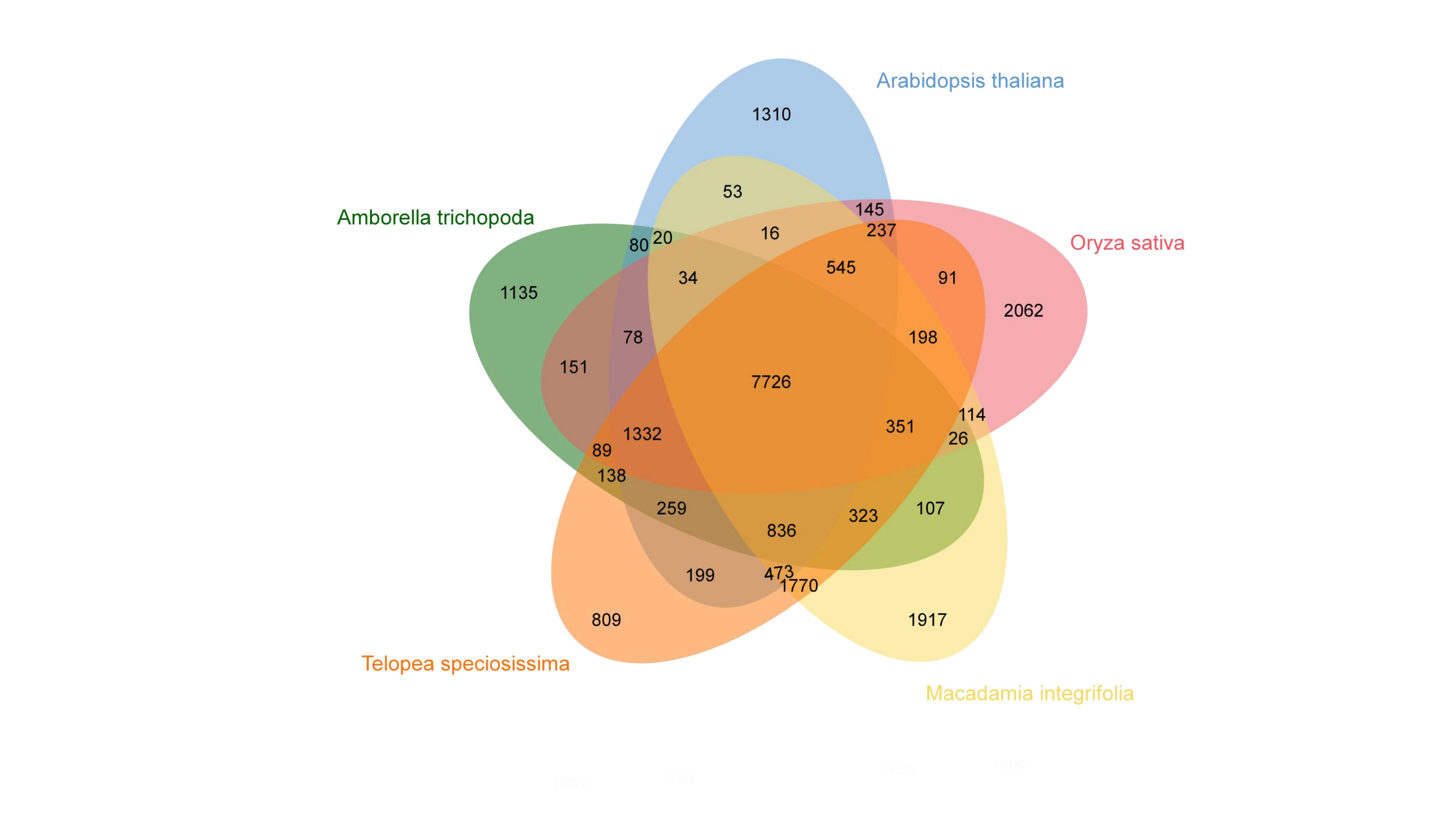
Comparative genomic analysis was performed using the genomes of macadamia, T. speciosissima, A. thaliana, A. trichopoda, and O. sativa. The results showed 7,726 gene families in the five species. The macadamia, T. speciosissima, A. thaliana, A. trichopoda, and O. sativa gene families were 1,917, 809, 1,310, 1,135 and 2,062, respectively (Supplemental Fig. 1). The Kyoto Encyclopedia of Genes and Genomes (KEGG)[20] analysis of the specific gene families in macadamia showed that these gene families were predominantly concentrated in metabolism-related biological functions such as metabolic pathways, biosynthesis of secondary metabolites, phenylpropanoid biosynthesis, and tryptophan metabolism (Fig. 3a). Further, the Gene Ontology (GO) enrichment analysis showed that the main functions of these gene families were signal transduction, protein phosphorylation, protein binding, and defense response (Fig. 3b).
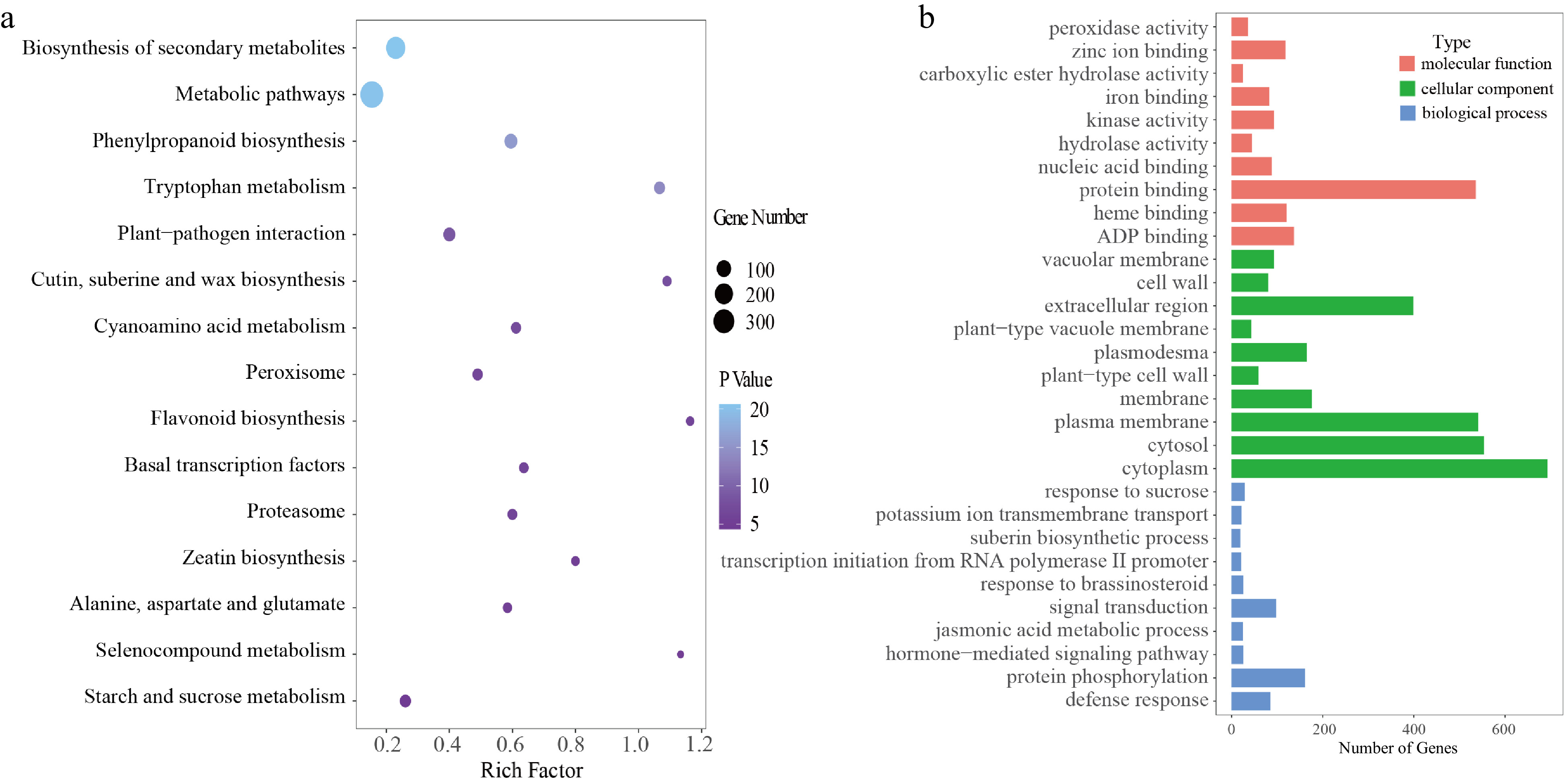
Figure 3.
Gene family analysis. (a) KEGG analysis of unigenes in the GR1 genome, (b) GO analysis of unigenes in the GR1 genome.
Genome-wide identification of GELP genes in M. integrifolia
-
We identified 120 GELP family members unevenly distributed on 14 chromosomes in macadamia, and most genes were located on chromosome 10 (Supplemental Fig. 2). In gene structure analysis, some differences were observed in the exon and intron structures of GELP family members. The number of exons of GELP family members ranged from 2 to 16, and the number of introns ranged from 1 to 15 (Fig. 4). Motif analysis revealed high motif conservation among GELP family members. Most GELP family genes contained motif1 and motif4, indicating that these two conserved motifs are particularly important in the GELP family. CAAT and TATA boxes were the most cis-acting elements in the promoters of all GELP family members, and they were present in all members of the GELP gene family. Among the GELP family members, 280 gene promoters contained cis-acting elements necessary for anaerobic induction, and 233 gene promoters contained abscisic acid cis-acting elements, 230 gene promoters contained methyl jasmonate cis-acting elements, 101 gene promoters contained salicylic acid cis-acting elements, 38 gene promoters contained auxin cis-acting elements, and 52 gene promoters contained gibberellin cis-acting elements. In addition, there were cis-acting elements for circadian control and defense and stress response (Supplemental Fig. 3).

Figure 4.
Analysis of conserved motifs and gene structure of the GELP family.
-
Macadamia has important economic value in the global food industry. A high-quality genome sequence is needed as a basis to promote research on the molecular mechanisms of macadamia and accelerate the breeding of superior varieties. GR1 is the first macadamia variety in China that is protected by intellectual property rights. This variety features early fruit setting, high yield, and strong stress resistance. The macadamia genome is relatively complex with high heterozygosity and outcrossing. To elucidate the genetic system and evolution of Proteaceae, we sequenced the genome of macadamia, providing new insights and genomic resources for breeding. We report a high-quality chromosome-scale genome assembly of passion fruit, with a contig N50 of 1.9 Mb and assembly to 14 pseudo-chromosomes. This reference genome is higher in continuity than the previously published macadamia genome, such as HAES 741, with a scaffold N50 ~413Kb. The high quality of our assembly can be attributed to the use of the unique combination of Nanopore sequencing[21,22] with chromosome-scale scaffolding via RagTag. The macadamia genome sequence provides an important resource for future molecular breeding and evolutionary studies.
This annotated chromosome-level reference genome of macadamia can provide important information on the gene content, duplication elements, gene location on 14 chromosomes and RNA types in macadamia. In addition, Macadamia is the most important genus of the family Proteaceae, and this high-quality assembled macadamia genome revealed the important role of macadamia in evolutionary history. The divergence between macadamia and its relative T. speciosissima occurred about 71.74 million years ago. The WGD event, a paleopolyploidy event, is common in plants, and it indicates the development of new gene functions or the formation of a new species. About 115.37 million years ago, Proteaceae and Nelumbonaceae differentiated to form a separate family, which provided a data base for future research on the evolutionary relationship of Proteales. This genomic information for macadamia will help clarify the evolutionary processes in Proteaceae species and contribute to improving the understanding of the physiological and morphological diversity of Proteaceae species.
Macadamia is rich in oil and unsaturated fatty acids, and the GELP gene family is involved in the regulation of plant growth and development, secondary metabolism and oil synthesis in fruits[23−26]. However, previous studies have found that GELP gene family members have been identified in Arabidopsis[27], rice, rape[24] and other plants, but studies of this gene family have not been reported in macadamia. To aid in future studies on oil synthesis in macadamia fruit as well as other molecular mechanisms, we sought to identify 120 members using our assembled macadamia gene genome. The number of GELP family members varies among species, which may be because of the different degree of evolution of the GELP family. Chromosome mapping analysis showed that GELP family genes were unevenly distributed on 14 chromosomes of macadamia. Gene structure analysis showed that most GELP genes contained 5 exons and 4 introns. GELP is structurally conservative. Promoter cis-acting element analysis showed that the promoters of GELP family members contained cis-acting elements related to hormone response and plant growth and metabolism. Therefore, the functions of GELP genes may be related to hormone response and growth and metabolism of plants.
-
Fresh leaves were collected from the Macadamia Germplasm Resource Nursery of Guangxi Institute of South Subtropical Agricultural Sciences, frozen in the field, and stored at −80 °C until DNA extraction. High-molecular weight genomic DNA was extracted from fresh-frozen macadamia leaves by the modified CTAB[28] method. Nanopore sequencing was adopted to obtain the long-read sequence data of 281 Gb.
Chromosome-level genome assembly
-
The chromosome-level genome assembled by Nock et al.[12] was downloaded from the NCBI (
www.ncbi.nlm.nih.gov/genome/?term=Macadamia+integrifolia ) database for use as the reference genome, and the genome of the Contig I version of GR1 was intended to be scaled to the chromosome level. The genomic size of cultivar 'HAES 741' assembled by Nock et al. was 744.64 Mb, with a scaffold N50 of 413.4 kb[12]. First, Nanopore sequence data were assembled de novo using NextDenovo. Redundans were used to remove the genome duplication[29]. Second, sequence errors from the preliminary assembly of the genome were removed using NextPolish[30]. Third, non-chromosomal sequences were removed from the downloaded reference genome sequences and only the chromosomal sequences were retained. Fourth, RagTag tools[31] were used for scaling the assembled GR1 genome to the chromosome level, and the GR1 genome sequence at the chromosome level was constructed. Finally, the integrity of the genome assembly based on a single-copy homologous plant-specific database was assessed using the benchmarking universal single-copy orthologs (BUSCO)[32] default setting.Gene prediction and functional annotation
-
Genome repeats were annotated from scratch by the RepeatMasker program (
www.repeatmasker.org ). Protein-coding genes were predicted by ab initio calculations, conserved protein homologs, and combinations of assembled transcripts based on repeated masked genomes. The homology prediction of protein sequences was performed using the macadamia genome HAES 741[12], waratah genome of family Proteaceae, and GR1 genome. In addition, transcriptome data of macadamia in different tissues at different periods was downloaded from the NCBI database. Transcriptome data were assembled by Trinity and used for preliminary annotation. The Augustus software (http://bioinf.uni-greifswald.de/augustus ) was used for gene prediction and annotation[33]. The MAKER[34] software was used to integrate these results in the final genetic model.Phylogenetic tree construction and evolution rate estimation
-
The OrthoMCL[35] program was used to identify macadamia, waratah, lotus flower, and the clusters of homologous genes of eight other species (A. thaliana, C. arabica, M. alba, S. italica, O. sativa, Z. mays, E. guineensis, and A. comosus) using the FastTree algorithm (v2.1.9)[36]. The maximum likelihood (ML) tree of single-copy homologous genes was constructed. This ML tree was converted to a super-time-scale phylogenetic tree by r8s[37] using the calibration time of the TimeTree website.
Syntenic analysis
-
The MCScanX toolkit[38] with default parameters was used to identify collinear blocks. Protein names were used as search queries for genomes of other plant species to find the best matching pair. Each aligned block represented homologous pairs derived from a common ancestor. The Nei–Gojobori method implemented in phylogenetic analysis by maximum likelihood (PAML)[39] was adopted to calculate the synonymous substitution rates (Ks) of homologs in the collinear region. The mean Ks value was considered to represent the collinear region.
Gene family analysis and comparison
-
The number of gene families in macadamia, T. speciosissima, A. thaliana, O. sativa, and A. trichopoda was calculated using the OrthoVenn2 website (
https://orthovenn2.bioinfotoolkits.net/home ). Besides macadamia-specific gene families, the protein sequences of specific gene families were screened and submitted to the KOBAS online website for GO and KEGG enrichment analyses.Pfam (
http://pfam.xfam.org/ ) was used to obtain the characteristic domain of the GELP gene family protein[40]. To build a more accurate hidden Markov model and predict all GELP family members in macadamia, the GELP family members in the GR1 genome were searched and screened by Hmmer 3.0, the screened sequences were compared with ClustalW, and the hidden Markov model of these verified sequences was constructed by hmmbuild. Finally, 120 GELP family members were screened. The gene structure of GELP family members was analyzed by TBtools[41] and the macadamia annotation file. The GELP family protein motifs were analyzed using MEME (http://meme-suite.org ) and visualized by TBtools. A 2000-bp sequence upstream of the GELP start codon (ATG) was obtained from the macadamia genome database as the promoter sequence. The cis-acting elements of promoters were predicted using the PlantCARE website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) and visualized by TBtools.Data availability
-
The whole genome sequence data reported in this paper have been deposited in the Genome Warehouse in China National Center for Bioinformation, under accession number PRJCA008938 that is publicly accessible at
https://ngdc.cncb.ac.cn/gwh . This research was supported by Guangxi Natural Science Foundation under Grant No.2019GXNSFBA18501.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (http://www.maxapress.com/article/doi/10.48130/TP-2022-0003)
-
Received 8 April 2022; Accepted 2 May 2022; Published online 22 May 2022
-
A chromosome-scale, high-quality macadamia GR1 reference genome was constructed through a combination of Nanopore sequencing, with 14 chromosomes
Protein sequences of macadamia and 11 other species of the same family were compared to evaluate expansion and contraction in the macadamia gene family
Proteaceae diverged from Nelumbonaceae nearly 115.37 million years ago and from Rubiaceae about 140 million years ago
Identified 120 GELP family members using our assembled macadamia genome.
-
# These authors contributed equally: Chengcai Xia, Sirong Jiang
- Supplemental Table S1 The comparison of GR1 and Kau genome assembly.
- Supplemental Table S2 Repeat sequences in the GR1 genome.
- Supplemental Table S3 Summary of the GR1 genome assembled chromosomes.
- Supplemental Fig. 1 Gene family analysis of five species.
- Supplemental Fig. 2 Chromosomal locations of DELP gene family.
- Supplemental Fig. 3 Analysis of cis-acting elements of the GELP family.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xia C, Jiang S, Tan Q, Wang W, Zhao L, et al. 2022. Chromosomal-level genome of macadamia (Macadamia integrifolia). Tropical Plants 1:3 doi: 10.48130/TP-2022-0003 

Chromosomal-level genome of macadamia (Macadamia integrifolia)
- Received: 08 April 2022
- Accepted: 02 May 2022
- Published online: 22 May 2022
Abstract: Macadamia from the family Proteaceae is a plant native to Australia and has long been favoured by people for its crispy and high nutritional and medicinal value. Here, the genome of GUIRE 1 (GR1), a highly heterozygous superior cultivar of macadamia nut, was sequenced and assembled using nanopore sequencing, and a 807-Mb genome (contig N50, 1.9 Mb; scaffold N50, 54.70 Mb) and 14 chromosomes were obtained. A total of 453 Mb (about 55.95%) repetitive sequences and 37,657 protein-coding genes were obtained by gene annotation and homologous protein comparison. Proteaceae diverged from Nelumbonaceae nearly 115.37 million years ago and from Rubiaceae about 140 million years ago. A genome-wide duplication (WGD) event occurred in macadamia 41 million years ago based on the WGD analysis. The functional enrichment analysis of M. integrifolia-specific gene families revealed their roles in signal transduction, protein phosphorylation, protein binding, and defense response. Here, a highly heterozygous genome of M. integrifolia was unlocked to provide a database for breeding and molecular mechanism research.
-
Key words:
- Macadamia genome /
- Proteaceae /
- Genome assembly /
- Genome annotation
{kind=link}
{kind=link}
{kind=link}