









-
Tea (Camellia sinensis) is one of the mainstream beverage crops in the world[1]. In the last decade, Chinese tea production has increased year on year, accounting for approximately 41.78% of the total world production (Food and Agriculture Organization of the United Nations statistics;
www.fao.org/faostat ). Tea flowers are the reproductive organs that profoundly impact the quality and yield of the tea seeds, the raw material for tea seed oil production[2]. In addition, tea seeds are also essential hybrid breeding materials[3]. Tea seed oil is rich in linoleic acid, tea polyphenols, and vitamin E. It is not only a healthy cooking oil but also further processed into biodiesel[4]. However, the prevalence of many pathogens has directly led to significant losses of tea products, including Alternaria spp.[5,6]. Alternaria alternata is a fungus epidemic worldwide and pathogenic to the leaves, flowers and fruits of many plants[7]. A. alternata also produces Alternaria toxin during plant infestation, which poses a risk to both plants and humans[8]. A. alternata has been reported to cause disease in many crops, such as kiwifruit[9], strawberries[10], olives [11] and rice[12]. In October 2021, flower blight was found on 18% of the flowers on 1/5 of the tea plants in a tea plantation located in Hefei, Anhui Province, China. In this study, morphological and molecular characterizations, as well as pathogenicity tests were conducted to identify the causative agent of the flower blight observed on the flowers of tea plants. -
The flower blight samples of tea plants were collected from a tea plantation in Hefei, Anhui Province, China (East longitude 117.27, North latitude 31.86). To isolate the pathogen, the diseased flowers were treated with 70% alcohol for 30 s and 1% NaOCl for 3 min, then washed three times with sterile water. The sample flowers were then blotted on sterile filter paper to dry the surface and subsequently transfer to potato dextrose agar (PDA) for incubation. After 4 d, single colonies were collected and separately transferred to fresh PDA. The isolated strains were grown on PDA at a temperature of 25 °C in the dark. On the 7th day after inoculation, the main characteristics of the colonies (texture, color, pigment release) were evaluated using microscopy (microscope: Zeiss Axio Vert. A1). After 15 d of incubation, colonies were rinsed with sterile distilled water to obtain their conidia and chlamydospore.
Molecular identification of the pathogen and phylogenetic construction
-
Seven-day-old mycelial cultures were collected for DNA extraction using the CTAB method[13]. Amplifications of internal transcribed spacer (ITS)[14], glyceraldehyde-3-phosphate dehydrogenase (GAPDH)[15], beta-tubulin (TUB)[16] by gene-specific primer pairs (Table 1) were conducted[7]. These primers were synthesized by Generalbiol (Chuzhou, China). Polymerase chain reaction (PCR) amplification reaction system was 20 μl, including 1 μl of DNA template (100 ng/μl), 0.5 μl of each primer (10 μM), 10 μl of 2×FastTaq Premix Buffer (TOLOBIO), and 8 μl of ddH2O. The conditions of the thermal cycler for PCR were: initial denaturation at 95 °C for 5 min, followed by 30 cycles in a thermal cycler (S1000 Thermal Cycler, Bio-Rad, USA) with denaturation at 95 °C for 30 s, annealing temperature at 55 °C (for ITS)/58 °C (for GAPDH)/51 °C (for TUB) for 30 s, extension at 72 °C for 30 s, and final extension at 72 °C for 10 min. PCR products were analyzed by 1.2% (w/v) agarose gels (containing 0.06‰ Gel Red nucleic acid dye) and run on a horizontal electrophoresis system (Bio-Rad, USA) at 120 V for 20 min. The forward and reverse strands of the PCR products were sequenced by Sanger sequencing, and the sequencing service provider was Generalbiol (Chuzhou, China). The obtained gene sequences were eventually submitted to the NCBI GeneBank and the accession numbers of these sequences are listed in Table 1.
Table 1. The barcode region/gene sequenced by the fungus, acronym, the primers used and the Genbank accession number of the isolate.
The sequences derived from ITS, GAPDH, TUB were concatenated into combinatorial sequences using Sequence Matrix (Table 2). Multiple sequences were compared using Clustal W of MEGA7.0. The phylogenetic tree was constructed using the maximum likelihood method (1,000 replicates) based on the Tamura-Nei model.
Table 2. Alternaria species from different crops used in the phylogenetic analysis.
Species Isolate
identificationHost/isolation-source Country GenBank accession number ITS GAPDH TUB A. alternata GB-GJ-2-3 Tribolium castaneum Korea MG554320 MH423920 MH423925 A. alternata CS36-1 C. sinensis China KY814631 KY814636 KY814626 A. alternata CS36-3 C. sinensis China KY814633 KY814638 KY814628 A. alternata CS36-5 C. sinensis China KY814635 KY814640 KY814630 A. mirabibensis CPC 38838 Plant litter Namibia MW175361 MW173104 MW173140 A. iranica EGS51-075 Allium Korea/China JF331513 JF331456 JF331440 A. vanuatuensis CNU093020 Allium Korea/China JF331501 JF331488 JF500412 A. prasonis EGS52-006 Allium Korea/China JF331514 JF331457 JF331589 A. radicina BMP0055 Daucus carota USA EU136660 EU142008 EU139382 A. panax CNU085010 araliaceous Korea JF417569 JF417650 JF417623 A. panax CNU3531 Araliaceae spp. Korea JF417574 JF417655 JF417628 A. kareliniae CTU C036 Karelinia caspia China KY945068 KY945065 KY945077 Pathogenicity test
-
The flower buds and leaves from a second-year tea plant (commercial variety 'Huangjinya') grown in a greenhouse were selected for pathogenicity testing. The fungus, which had been cultured on PDA for 15 d, was stirred by adding 10 ml of ddH2O. The liquid was then filtered through a single layer of sterile gauze to remove the mycelium. The filtered liquid was added to Tween 20 and adjusted to a concentration of 0.05%. The conidia were dropped on a hemocytometer plate and observed under a microscope to calculate the conidia concentration. The final concentration was adjusted to 106 conidia/ml[17]. Two groups of plant materials, including undamaged flower buds and leaves punched with a sterile needle (0.8 to 1 mm),were separately sprayed with conidia or ddH2O. These two experiments were repeated three times using three independent healthy tea plants. All treated plants were incubated in a light incubator (16 h light/8 h dark) at 25 °C. From the tissues of infected flowers, the fungi were re-isolated and re-identified based on morphological and molecular barcoding regions to fulfill the Koch hypothesis.
-
In October of 2021, in comparison to the healthy flowers (Fig. 1a), typical flower blight symptoms were observed on some flowers of tea plants in a tea plantation (Fig. 1b). Onset of the flower blight appeared on September or October of each year. The disease began with punctated brown disease spots and then rapidly expanded to the whole petals, and consequently a large number of flowers died. As the temperature decreased, the symptoms were relieved.
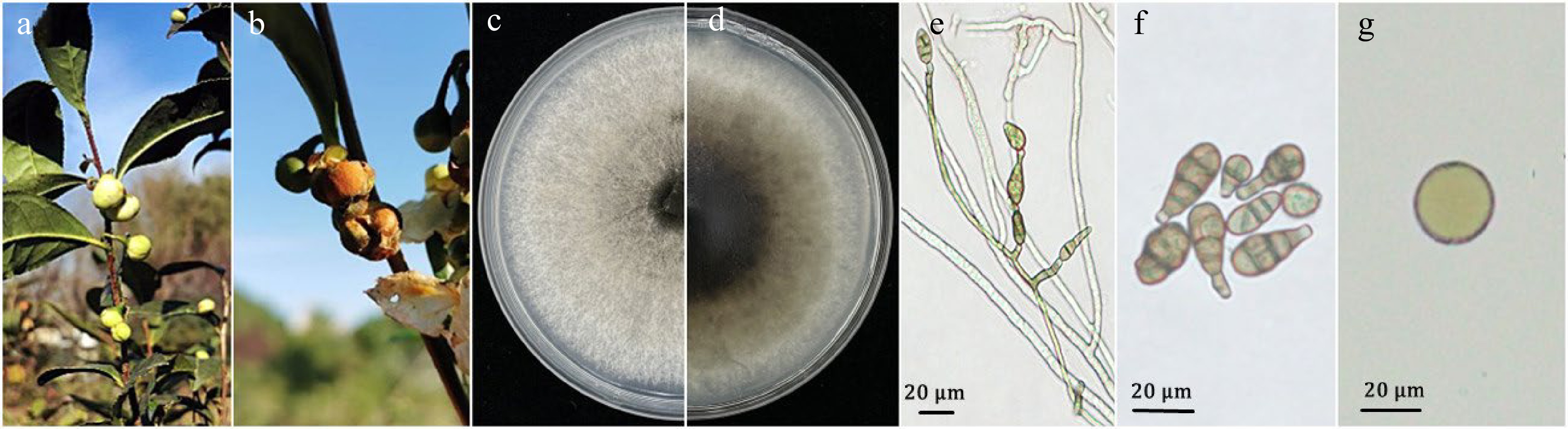
Figure 1.
Alternaria alternata strains AAU-CS-F4, AAU-CS-F9 and AAU-CS-F10 inciting flower blight on Camellia sinensis. (a) Healthy flower bud; (b) natural flower blight appearing on tea plants taken in the field; (c, d) colony on PDA for 7 d (front and reverse); (e) short chains of conidia of dwarf dendrites; (f) conidia; (g) chlamydospore. Scale bars = 20 μm.
Diseased flowers were collected from tea plantations to isolate pathogens. The junctions across diseased and healthy tissues were collected and cultured on PDA. Eventually eight independent single-spore isolates were obtained from diseased tea plant flowers. These isolates were further incubated on PDA under dark conditions for 7 d at 25 °C and abundant mycelium were grown with a central gray-black to gray-brown color accompanied by white margins (Fig. 1c & d). After 15 d of incubation, the colonies on PDA produced a large number of conidia and a few chlamydospore. Conidiophores on mycelium showed solitary conidia or short chains, occasionally branched (Fig. 1e). Microscopy observations showed that the conidia were triangular in shape, brown in color, with beak-like projections at the tips, and 1-6 layers of transverse septa and 0-4 layers of longitudinal septa on the spores. The size of the conidia was 9.2 to 35.3 µm in length (
$ \bar{x} $ $\bar{x} $ $ \bar{x} $ In order to further determine the species identity of these single-spore isolates, their ITS, GAPDH and TUB gene regions were selected for molecular characterization based on the taxonomic study of the genus Alternaria[7,16]. The sequences of the ITS, GAPDH and TUB genes derived from the eight single-spore isolates were amplified and analyzed by multiple sequence alignment, and SNPs were found in the sequences of the ITS, GAPDH and TUB genes. The eight strains were accordingly classified into three distinct groups based on differential SNPs categorization, among which AAU-CS-F4, AAU-CS-F9 and AAU-CS-F10 were the representative strains of each of the three groups, i.e., Group I (AAU-CS-F4, AAU-CS-F6 and AAU-CS-F7), Group II (AAU-CS-F3, AAU-CS-F5, AAU-CS-F8 and AAU-CS-F9) and Group III (AAU-CS-F10). AAU-CS-F10 and AAU-CS-F4, AAU-CS-F9 have one-base SNP in the ITS gene region, and AAU-CS-F4 and AAU-CS-F9, AAU-CS-F10 have one and two base SNPs in the TUB and GAPDH gene regions, respectively. On the basis of NCBI-BLASTn analysis, the ITS sequences of AAU-CS-F4, AAU-CS-F9 and AAU-CS-F10 showed 100%, 100% and 99.81% identity with A. alternata accessions KP124302, KP124298 and KP124332, respectively. Moreover, the GAPDH sequences showed 99.65%, 100% and 100% identity with A. alternata records KP124159, KP124155 and KP124187, respectively. Consistently, the TUB sequences showed 99.82%, 100% and 100% similarity with the accessions KY814630, KY814628 and JQ811937 of A. alternata, respectively. In the phylogenetic tree constructed from the combined sequences of ITS, GAPDH, TUB genes, AAU-CS-F4, AAU-CS-F9 and AAU-CS-F10 were closely clustered into the same branch with A. alternata and this branch was in the ML tree with 98% bootstrap support (Fig. 2).

Figure 2.
Phylogenetic analysis of AAU-CS-F4, AAU-CS-F9 and AAU-CS-F10. Phylogenetic tree was inferred from the combined ITS, GAPDH and TUB sequences of the genus Alternaria. The phylogenetic tree was constructed using the maximum likelihood method (1,000 replicates) by MEGA 7.0. The isolates in this study are marked with a red star. Scale bars represent 0.01 nucleotide substitutions per locus.
Flower buds and leaves from healthy tea plants were selected for pathogenicity testing. Healthy flower buds from tea plants were sprayed with 106 conidia/ml spore suspension and ddH2O, respectively. The treated tea plants were incubated in a light incubator (16 h light/8 h dark) at 25 °C. After 3 d, no lesion symptoms were found in the control group (Fig. 3a & b). Meanwhile, lesions appeared on the surface of the flower buds inoculated with the spore suspension, showing a brown to dark brown color, and the lesion symptoms were similar to those found in the garden (Fig. 3c & d). The disease-causing fungus was re-isolated from the lesioned tissues and A. alternata was again identified as the pathogen, and Koch's theorem was therefore fulfilled in this study. In order to explore whether AAU-CSs also have strong pathogenicity on tea leaves, Healthy leaves were inoculated with ddH2O (Fig. 4a) and spore suspension (Fig. 4c) , respectively. As a result, no pathological phenotypes from ddH2O treatment (Fig. 4b), the spore suspension treatment resulted in weak pathological symptom at the pinholes of spore-inoculated leaves after 10 d (Fig. 4d).

Figure 3.
Symptoms observed on the flowers in C. sinensis plant grown at 3 d post-inoculation. (a, b) Three day sterile water group and (c, d) spore inoculation group.

Figure 4.
A. alternata strains inciting inconspicuous leaf disease on C. sinensis at 10 d post-inoculation. Symptoms observed on a leaf in C. sinensis grown as the sterile water group and spore inoculation group after 0 d (a, c) and 10 d (b, d).
Tea plants are extensively cultivated in the mountains of southern China. However, the infection by pathogenic fungi often leads to a considerable reduction in the quality and yield of tea tree products. A. alternata is a worldwide phytopathogenic fungus that causes substantial crop yield reductions[19]. A. alternata has been identified as the causal pathogen of both flower blight in Geraldton wax[20], Zinnia acerosa[21], orchid[22] and Olea europaea[23] and leaf spot disease in C. sinensis[5] and C. japonica[24]. However, the diseases of tea plant flowers have rarely been reported, and the pathogen causing flower blight is still unknown. To our knowledge, this is the first report of A. alternata causing flower blight of tea plant in China. In our study, A. alternata was identified as the causal pathogen of flower blight observed in C. sinensis, directly leading to flower withering and reduction of seed production. It is worth mentioning that A. alternata isolates from flowers showed much weaker virulence to leaves (Fig. 4d) than that isolated from leaves in the pathogenicity of inoculated leaves[25]. Due to some SNPs found in ITS-GAPDH-TUB sequences among these isolates, we speculated that A. alternata may have evolved a large number of different physiological races in different geographical areas and produced differential pathogenicity in different organs/tissues.
-
This is the first report of A. alternata, being isolated from flowers of tea plants suffering from flower blight in Hefei, China. This result will provide a foundation effort aimed at presenting tea plant diseases caused by A. alternata in the future.
Xie X. and Wang L. acknowledge State Key Laboratory of Tea Plant Biology and Utilization, Anhui Agricultural University for providing tea plants for the pathogenicity experiments. This study was supported by grants from the National Natural Science Foundation of China (31900257, 31972474).
-
The authors declare that they have no conflict of interest.
- Copyright: 2022 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xie X, Yu Q, Li X, Liu Y, Wang L. 2022. First report of Alternaria alternata causing flower blight on Camellia sinensis in Hefei, China. Beverage Plant Research 2:16 doi: 10.48130/BPR-2022-0016 

First report of Alternaria alternata causing flower blight on Camellia sinensis in Hefei, China
- Received: 25 July 2022
- Accepted: 24 August 2022
- Published online: 13 September 2022
Abstract: Flowers are an essential organ for sexual reproduction of higher plants. Severe lesions along with flower blight on tea (Camellia sinensis) plants were observed in the experimental tea plantation located in Hefei (China). The pathogens isolated from diseased flowers matched the morphological peculiarity of Alternaria alternata. The species characteristics of A. alternata were further confirmed by both pathogenicity tests and multi-gene phylogenetic analyses by using internal transcribed spacer (ITS), glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and beta-tubulin (TUB). The combined phylogeny analysis using sequences derived from the ITS, GAPDH and TUB showed that the isolated pathogens belong to the genus Alternaria. Pathogenicity tests conducted on healthy tea flowers and leaves manifested typical symptoms of flower blight while weaker symptoms of leaf spot, demonstrating the A. alternata isolates were the causal agents of flower blight disease on tea plants. This fungus is first reported as a pathogen causing flower blight on C. sinensis in this study.
-
Key words:
- Alternaria alternata /
- Camellia sinensis /
- flower blight /
- fungi /
- pathogen detection