









-
Transcription factors (TFs) are essential in domesticating plants and are targets for molecular breeding. They play a key role in binding to cis-acting regulatory elements[1]. More than 1000 TF genes have been predicted in angiosperm genomes, and these TF genes can be classified into 58 families based on their DNA-binding domains. The WRKY gene family is the seventh largest TF family in flowering plants[2], where 74 WRKY genes have been identified in dicotyledonous Arabidopsis[3], while 109 WRKY genes have been identified in monocotyledonous rice[4].
The WRKY gene family shares a highly conserved WRKY domain at the N-terminus and a metal chelating zinc finger-like motif at the C-terminus[5]. Some variants of the WRKY domain, such as WRKYGEK, WRKYGKK, WSKYEQK, WRKYSEK, WRRYGQK, WSKYGQK, WVKYGQK, WKKYGQK, WRICGQK, WRMCGQK, WIKYGQK, and WKRYGQK, have been found in various plants[6,7]. Two main types of zinc finger-like motifs C2H2 (C-X4-5-C-X22-23-H-X-H) and C2HC (C-X7-C-X23-H-X1-C) have been identified[5]. All WRKY domains and zinc finger-like motifs are for structural stabilization[5]. According to the phylogenetic analysis, WRKY proteins are classified into three major groups—I, II, and III—based on the number of WRKY domains and pattern features of the zinc finger-like motif[8]. Only group I members have two WRKY domains, whereas those with one WRKY domain belong to group II or III[8]. Group II has five subgroups—IIa, IIb, IIc, IId, and IIe[8]. Those with C2HC (C-X7-C-X23-H-X1-C) zinc finger-like motifs belong to group III, whereas the members of groups I and II have C2H2 (C-X4-5-C-X22-23-H-X-H) zinc finger-like motifs[5,8].
WRKY proteins participate in various plant life activities, including defense against stresses, growth and development, biosynthesis, and regulation of hormone signaling[9,10]. As key regulators in plant immune response to various biotic stresses, WRKYs have been extensively investigated in rice, Arabidopsis, soybean, wheat, pepper, and wild tomato[11−23]. In rice, OsWRKY62.1 and OsWRKY67 have been established as important regulators against rice pathogens[12,22]. WRKY8, WRKY29, WRKY38, WRKY48, WRKY62, and WRKY33 are involved in response to pathogens in Arabidopsis[11,13,18−20]. Moreover, GmWRKY136, GmWRKY53, GmWRKY86, TaWRKY49, TaWRKY62, CaWRKY27, CaWRKY40, CaWRKY40b, CaWRKY6, and SpWRKY1 are involved in defense responses induced by biotic stress[14−17,21,23].
In addition to the response to these biotic stresses, WRKY genes can function effectively in most abiotic stress responses or tolerances in various plants. For example, WRKY25, WRKY39, and WRKY54 in Arabidopsis, PoWRKY13 in tomato, and CaWRKY40 in pepper respond to heat stress[24−28]. In contrast, AtWRKY34, VvWRKY24, SlWRKY33, and VbWRKY32 are associated with cold tolerance[29−32]. In addition, GhWRKY39-1, GbWRKY1, GmWRKY13 and GmWRKY17 are identified as key regulators in response to salt stress[9,33−35].
Among the various abiotic stresses that plants are exposed to during growth and development, drought is one of the most detrimental environmental factors limiting growth, development, distribution range, and especially plant yield[36]. WRKY genes can regulate the abscisic acid signaling pathway in response to drought[37]. WRKY can also resist drought stress by enhancing dehydration tolerance, reducing stomatal density, modulating ethylene response factors, and effectively eliminating reactive oxygen species by activating cellular antioxidant systems[2]. Thirty-seven WRKY genes that respond to drought stress have been identified in many plant species, including model and non-model plants. In model plants, WRKY1, WRKY1, WRKY18, WRKY40, WRKY46, WRKY54, WRKY57, WRKY60, WRKY63, and WRKY70 in Arabidopsis, and OsWRKY11, OsWRKY45, OsWRKY30, OsWRKY80, and OsWRKY47 in rice, play positive or negative roles in responding to drought stress. Numerous WRKY genes regulating drought resistance have also been identified in non-model plants, such as PbrWRKY53 in Pyrus betulaefolia[38], FvWRKY42 in the diploid woodland strawberry (Fragaria vesca)[39], and FcWRKY70 in Fortunella crassifolia to name a few[40].
In addition to stress response, WRKY genes play important roles in a range of processes such as root development, senescence regulation, seed germination, and plant nutrient utilization[41−47]. For example, WRKY42, WRKY45, WRKY75, and WRKY6 in Arabidopsis, and OsWRKY74 and OsWRKY80 in rice are involved in plant nutrient utilization[42]. AtWRKY6, AtWRKY53, and OsWRKY45 can regulate leaf senescence[46]. Moreover, AtWRKY12, AtWRKY13, AtWRKY71, and OsWRKY11 are associated with floral development in angiosperms[41,44,47].
Based on these important roles, the identification of WRKY gene family members in plants is the basis for further functional studies. However, identification is difficult because of the large number of WRKY gene family members in most plants. This study examines two pertinent questions to address this difficulty: (1) Is there a sequence pattern for quick identification? and (2) Can this sequence pattern be used for the identification of stress-responsive WRKY genes in Dendrobium catenatum?
D. catenatum, a perennial herb of Dendrobium in Orchidaceae, has important medicinal properties and high ornamental value. In China, D. catenatum is found in many provinces with wide differences in both temperature and humidity, including Anhui, Zhejiang, Guangxi, Hunan, Yunnan, and Guizhou. The additional stress-related genes in D. catenatum may have contributed to its adaptation to a wide range of environments[48]. Its epiphytic lifestyle and wide distribution range are closely related to its stress tolerance.
Because there may be a common amino acid pattern of WRKY genes that respond to stress, a confirmatory study was performed using D. catenatum as an example. First, all sequences of reported functional WRKY genes were summarized, and the sequence features of genes belonging to groups II-c, II-d, and II-e that responded to stress were completely analyzed, and the common patterns in each of these three subgroups were determined, which might be markers for identifying stress-resistant WRKY genes. Second, D. catenatum was used to verify the accuracy of these patterns. The WRKY gene family members were identified in D. catenatum and their sequence characteristics were analyzed. The candidate stress-responsive WRKY genes were identified in D. catenatum based on these sequence patterns. The consistency of sequence characteristics between candidate WRKY members in D. catenatum and identified sequence patterns were compared. Finally, the DcaWRKY gene expression pattern was analyzed in different tissues under drought stress, and the expression patterns of seven randomly selected genes were confirmed.
-
All reported functional WRKY sequences were obtained, and their basic information and sequence features were analyzed. The number of amino acids, molecular weight, theoretical isoelectric point (pI), instability index, aliphatic index, and grand average of hydropathicity (GRAVY) of each WRKY protein sequence were calculated by ExPASy protparam (
https://web.expasy.org/protparam.html ). All amino acid sequences of the WRKY gene family from reported plants were aligned by Multiple Alignment using Fast Fourier Transform (MAFFT) with the E-INS-I strategy[49]. The results of multiple alignments were visualized by GeneDoc software (www.softpedia.com/get/Sc-ience-CAD/GeneDoc.shtml ).Phylogenetic analysis of the WRKY family members from Arabidopsis, rice, and other reported species
-
All Arabidopsis and rice genomic data were downloaded from the Phytozome v12.1.6 database (phytozome.jgi.doe.gov) to create a protein dataset. A hidden Markov model (HMM) search was performed against the protein database using the WRKY domain file (PF03106) that was downloaded from the Pfam database to identify WRKY family members[50]. To verify the preliminary results, each predicted sequence was analyzed using Pfam, simple modular architecture research tool (SMART), and NCBI CD-Search (
www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi )[51]. Sequences with obvious errors were excluded.All WRKY sequences in Arabidopsis and rice, as well as reported WRKY gene sequences, were used to reconstruct a phylogenetic tree by the maximum likelihood (ML) method using PhyML 3.0 software[52]. Here, 1000 bootstrap replicates were used to test the reliability of the tree. Inspired by a Shimodaira–Hasegawa-like procedure, the approximate likelihood-ratio test branch support was estimated with the Whelan and Goldman model[52].
Identification of stress-related amino acid sequence patterns
-
To identify the stress response sequence patterns in groups II-c and II-e, sequences of stress-related genes clustered in the same branch in the subgroups were aligned by MAFFT with the E-INS-I strategy[49]. Stress-responsive amino acid sequence patterns in groups II-c and II-e were identified separately by the results of multiple comparisons by GeneDoc software.
Similarly, sequences of WRKY members of group II-d clustered in a branch related to abiotic stress were aligned to identify the amino acid sequence patterns in this subgroup.
Identification and characterization of WRKY members in D. catenatum
-
All WRKY protein sequences of D. catenatum in the two genome versions were downloaded for further analysis[48,53]. The file of the WRKY domain (PF03106) was used to search against the D. catenatum genomes by performing hmmsearch using HMMER software[49,50].
Various physicochemical parameters, including theoretical pI, the number of amino acids, aliphatic index, molecular weight, instability index, and GRAVY for DcaWRKY proteins, were computed by the ExPASy protparam online tool.
The WRKY family gene structure in D. catenatum was analyzed using the Batch SMART plug-in in TBtools[54]. The exons, introns, and untranslated regions of the DcaWRKY genes were analyzed and visualized by loading the GFF files of D. catenatum to the TBtools software, which was also used for analyzing and searching for conserved motifs.
Multiple sequence alignment and phylogenetic analysis of WRKY family members in D. catenatum
-
MAFFT was used to align and manually adjust the protein sequences of all WRKY family members in D. catenatum using the E-INS-I strategy[49]. By submitting the multiple sequence results to GeneDoc software, the multiple sequence alignments could be visualized and analyzed.
To study the phylogenetic relationships of DcaWRKY proteins and orthologs in Arabidopsis and rice, a phylogenetic tree was reconstructed using the ML method approach with 1000 bootstrap replicates and the Whelan and Goldman model by PhyML[52].
Expression analysis of all WRKY family members in D. catenatum under drought stress
-
Transcriptome data used in this study was from Wang et al.[55]. Briefly, healthy individuals (~12 cm tall) of D. catenatum were grown in a greenhouse (12/12 h light/dark, light intensity ~100 μmol·m−2·s−1; 28/22 °C day/night; 60%/70% relative humidity day/night). After first-day irrigation was performed, water was withheld for 10 d to simulate drought stress. The fourth and fifth leaves from the apex and young roots were collected from each plant at 9:00 h when the base material was in a drought and normal growth state. Salmon v0.9.1 was used to estimate gene abundance as reading counts based on alignment patterns[56].
The DcaWRKY gene expression patterns in each tissue were analyzed, and heatmaps were generated using TBtools software. The Fragments Per Kilobase of exon model per Million mapped fragments (FPKM) values for each gene in all root and leaf samples were log2 transformed[54,56]. Each sample had three biological replicates. Those with more than 2.0-fold change in expression under drought treatment compared with normal treatment were identified as differentially expressed genes.
Quantitative real-time polymerase chain reaction (qRT-PCR) analysis of selected DcaWRKY genes
-
Among all the differentially expressed DcaWRKY genes in both roots and leaves, four genes with decreased expression and three genes with increased expression when responding to drought stress were randomly selected. These seven genes — Dca002550, Dca002715, Dca005648, Dca006787, Dca007842, Dca010430, and Dca016437 — were used for further qRT-PCR analysis. Total RNA was extracted using the cetyltrimethylammonium bromide (CTAB) method. Extracted RNA was reverse-transcribed using the Fast Quant RT Kit with gDNase (Tiangen, Beijing, China).
The relative transcription levels of the selected WRKY genes during different treatments were analyzed by qRT-PCR. 18S rRNA was used as the internal control. The primer sequences for the selected genes were designed with the Primer3Plus interface (
www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi/ ). All qRT-PCR assays were performed on a Roche 480 real-time PCR system (LightCycler® 480; Roche, Basel, Switzerland) with three biological replicates. The reactions were conducted in a reaction volume of 10 μL with three technical replicates for each sample. The PCR program was as follows: 95 °C for 3 min and 45 cycles of 95 °C for 10 s, 65 °C for 20 s, 72 °C for 30 s, 95 °C for 5 s, and 60 °C for 1 min. The 18S gene was used as an internal control to calculate the relative fold expression levels according to the Ct method. Finally, the TBtools software was used to generate a heatmap of DcaWRKY genes in leaves. -
All reported functional WRKY genes are summarized in Table 1. The majority of genes were from Arabidopsis, rice, and soybean, with 27, 16, and nine, respectively. Seven WRKY genes were identified in wheat and six WRKY genes were identified in land cotton, whereas only one functional WRKY gene was reported in Pyrus, Chrysanthemum, Tamarix, and Macrotyloma. Numerous functional studies of WRKY genes have mostly focused on model plants and crops, whereas reports on ornamentals, especially flowers and medicinal plants, are few.
Table 1. Reported functional WRKY genes summarized in this study.
Group Gene ID Species Function References III GhWRKY33 Gossypium hirsutum Tolerance to drought [58] III PbrWRKY53 Pyrus betulaefolia Tolerance to drought [38] II C GmWRKY54 Glycine max Tolerance to drought [59] II A GmWRKY27 Glycine max Tolerance to drought [58] III AtWRKY63 Arabidopsis thaliana Tolerance to drought [60] I TaWRKY2 and TaWRKY19 Triticum aestivum Tolerance to drought [61] II C OsWRKY11 Oryza sativa Tolerance to drought [57] III OsWRKY45 Oryza sativa Tolerance to drought [62] II A WRKY18, WRKY40 and WRKY60 Arabidopsis thaliana Tolerance to drought [63,64] I WRKY1 Arabidopsis thaliana Tolerance to drought [65] III WRKY46, WRKY54, and WRKY70 Arabidopsis thaliana Tolerance to drought [42] II C AtWRKY57 Arabidopsis thaliana Tolerance to drought [37] I A OsWRKY30 Oryza sativa Tolerance to drought [66] II OsWRKY80 Oryza sativa Tolerance to drought [67] III OsWRKY47 Oryza sativa Tolerance to drought [68] I FvWRKY42 Fragaria vesca Tolerance to drought [39] II D ZmWRKY58 Zea mays Tolerance to drought [41] II E ZmWRKY106 Zea mays Tolerance to drought [69] II A ZmWRKY40 Zea mays Tolerance to drought [69] II E CmWRKY10 Chrysanthemum morifolium Tolerance to drought [70] III and I TaWRKY1 and TaWRKY33 Triticum aestivum Tolerance to drought [71] II D ThWRKY4 Tamarix hispida Tolerance to drought [72] I MuWRKY3 Macrotyloma uniflorum Tolerance to drought [73] II D GhWRKY17 Gossypium hirsutum Tolerance to drought [34] III GhWRKY41 Gossypium hirsutum Tolerance to drought [74] I TaWRKY44 Triticum aestivum Tolerance to drought [75] III FcWRKY70 Fortunella crassifolia Tolerance to drought [40] II C GmWRKY12 Glycine max Tolerance to drought [76] III ZmWRKY79 Zea mays Tolerance to drought [77] II D GhWRKY21 Gossypium hirsutum Tolerance to drought [78] III SlWRKY81 Solanum lycopersicum Tolerance to drought [79] I SPF1 Ipomoea batatas Root development [43] I and II A ABF1 and ABF2 Avena fatua Seed germination [45] II B WRKY42 and WRKY6 Arabidopsis thaliana Plant nutrient [42] II C WRKY45 and WRKY75 III and II E WRKY74 and WRKY80 Oryza sativa Plant nutrient [42] III AtWRKY53 Arabidopsis thaliana Leaf senescense [46] II C OsWRKY11 Oryza sativa Floral development [41] II C AtWRKY12 and AtWRKY13 Arabidopsis thaliana Floral development [44] II C AtWRKY71 Arabidopsis thaliana Floral development [47] I AtWRKY2 Arabidopsis thaliana Reproductive development [80] I OsWRKY70 Oryza sativa Defense response [81] II C FvWRKY48 Fragaria vesca Pectin degradation [82] II E OsWRKY13 Oryza sativa Regulated ABA signaling and tolerance to salt [83] II C VlWRKY3 Vitis vinifera Response to Golovinomyces cichoracearum and tolerant to salt [84] II C GhWRKY68 Gossypium hirsutum Reduced salt tolerance and drought resistance [85] I GhWRKY25 Gossypium hirsutum Tolerance to salt [86] I VvWRKY24 Vitis vinifera Tolerance to cold [29] I AtWRKY25 and AtWRKY33 Arabidopsis thaliana Tolerance to heat [25] I AtWRKY34 Arabidopsis thaliana Negative regulator in cold stress [31] III AtWRKY53 Arabidopsis thaliana Reduced drought resistance [87] III AtWRKY63 Arabidopsis thaliana Regulated ABA signaling [60] III AtWRKY54 Arabidopsis thaliana Response to heat stress [28] II C OsWRKY72 Oryza sativa Sensitive to salt, sucrose, and ABA [88] III OsWRKY74 Oryza sativa Tolerance to cold and Pi deprivation [89] II A OsWRKY76 Oryza sativa Tolerance to cold [90] III OsWRKY89 Oryza sativa Tolerance to UV [91] II A GmWRKY17 Glycine max Reduced salt tolerance [34] III BcWRKY46 Brassica campestris Tolerance to salt [92] III BhWRKY1 Boea hygrometrica Tolerance to salt [93] III and I VpWRKY1 and VpWRKY2 Vitis pseudoreticulata Tolerance to salt and cold [94] II A VpWRKY3 Vitis pseudoreticulata Tolerance to salt [95] III TcWRKY53 Thlaspi caerulescens Negative regulator in osmotic stress [96] I NaWRKY3 Nicotiana attenuata Sensitive to mechanical damage [97] I and II D JrWRKY2 and JrWRKY7 Juglans regia Tolerance to drought and cold [98] III SbWRKY30 Sorghum bicolor Tolerance to salt and drought [99] II C SbWRKY50 Sorghum bicolor Tolerance to salt [100] II A MdWRKY30 Malus domestica Tolerance to salt and osmotic stress [101] II C GbWRKY1 Gossypium barbadense Tolerance to salt [35] I VbWRKY32 Verbena bonariensis Tolerance to cold [28] II C OsWRKY67 Oryza sativa Negative regulator of innate defense response [22] II A OsWRKY62.1 Oryza sativa Positive regulator of PTI and ETI against pathogens [12] III AtWRKY38 and AtWRKY62 Arabidopsis thaliana Response to bacterial pathogen [11] II A,II C, and II B GmWRKY136, GmWRKY53, and GmWRKY86 Glycine max Tolerance to SCN [23] II C and III TaWRKY49 and TaWRKY62 Triticum aestivum Tolerance to stripe rust [102] II A CaWRKY40b and CaWRKY40 Capsicum annuum Negative regulation of plant immunity [17,24] II B CaWRKY6 Capsicum annuum Tolerance to R. solanacearum [15] I SpWRKY1 Solanum pimpinellifolium Tolerance to Phytophthora infestans [16] II D ZmWRKY17 Zea mays Negative regulator of salt stress [103] II D GhWRKY39-1 Gossypium hirsutum Tolerance to salt [33] II C AtWRKY8 Arabidopsis thaliana Defense response [13] II E CaWRKY27 Capsicum annuum Response to Ralstonia solanacearum infection [14] II C AtWRKY48 Arabidopsis thaliana Tolerance to P. syringae [20] II E AtWRKY29 Arabidopsis thaliana Resistance to P. syringe [18] II C PoWRKY13 Populus Response to heat stress [26] tomentosa III SlWRKY33 Solanum lycopersicum Tolerance to cold [30] II D GmWRKY13 Glycine max Response to salt and mannitol [9] To analyze the branch specificity of the WRKY gene family that responded to stress, a phylogenetic tree was reconstructed from AtWRKY genes, OsWRKY genes, and all reported functional WRKY genes (Fig. 1). All reported functional WRKY proteins were divided into three groups—I, II, and III, with group II being further divided into five subgroups—II-a, II-b, II-c, II-d, and II-e. Most WRKY proteins were widely distributed in all these groups rather than concentrated in the same branch. However, OsWRKY67, SbWRKY50, GmWRKY21, AtWRKY8, and AtWRKY48 play important roles under stress and were clustered in a branch of group II-c. Similarly, as essential stress-related genes, WRKY80 and WRKY13 in rice, WRKY29 in Arabidopsis, CaWRKY27 in pepper, and ZmWRKY106 in maize clustered in one branch of group II-e. In addition, reported genes responding to abiotic stress, including ZmWRKY58, GhWRKY21, GhWRKY17, ThWRKY4, GmWRKY13, GhWRKY39-1, and JrWRKY7, were all located in group II-d.
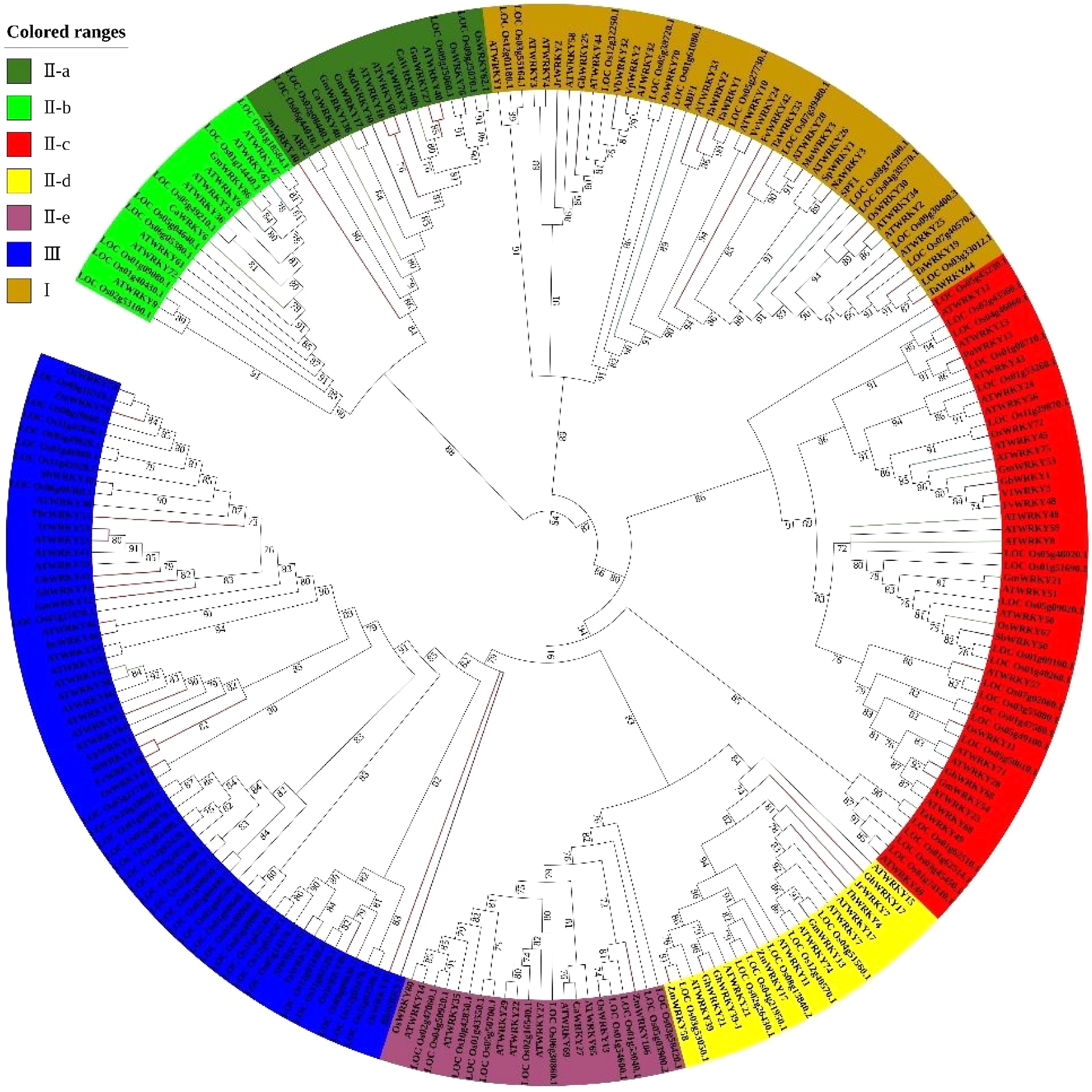
Figure 1.
Phylogenetic analysis of WRKY family proteins in Arabidopsis, rice, and other reported species.
Therefore, because of these distribution features of reported functional WRKY genes, the identification of amino acid sequence patterns in response to stress focused on groups II-c, II-d, and II-e.
Identification of the pattern of stress-related WRKY genes
-
To further characterize the stress-responsive WRKY proteins for identifying the amino acid patterns in different subgroups, the properties of all drought-resistant WRKY proteins (molecular weight, pI, instability index, etc.) were analyzed (Supplemental Table S1). The molecular weight of stress-related WRKY proteins ranged from 16801.82 to 74561.15, PI ranged from 4.86 to 9.96, and the instability index ranged from 37.86 to 88.65, showing a wide range of values.
From the alignment results, the amino acid patterns responding to stress were found in groups II-c, II-d, and II-e (Fig. 2). In contrast to groups II-d and II-e, where only stress-related WRKY genes were reported, group II-c contained WRKY genes that play important roles in plant growth and development besides stress-response. All stress-related genes clustered in the group II-e branch had the amino acid pattern 'PSD-S/A/L-WAWRKYGQKPIKGSPYPR-G/S-YYRCSSSKGC'. Similarly, the amino acid pattern 'VPA-I/V-S-X-K-M/L/V/I-ADIP-P/A/V-D-D/E-Y/F-S-WRKYGQKPIKGSP-H/Y-PRGYYKCS-S/T-V/M-RGCPARKVER' was found in the sequences of reported genes responding to abiotic stress clustered in the group II-d branch, which might be closely related to stress. In addition, for group II-c, an amino acid sequence pattern 'T-R/K-S/T-E/Q/D-V/I/L-E/D-I/V/H/N-L/M-D/E-D-G/E-F/Y-K/R-WRKYG-Q/K-K-A/T-VKN-S/N-P' in the stress-related genes clustered in one of the branches. From the alignment results, the sequences of WRKY genes involved in plant growth and development belonging to group II-c (FvWRKY48, ATWRKY75, ATWRKY45, ATWRKY12, ATWRKY13, and ATWRKY71) did not match this amino acid sequence pattern, further supporting the accuracy of this amino acid pattern in group II-c (Fig. 2a).

Figure 2.
Multiple sequence alignment of reported WRKY family proteins. (a) Multiple sequence alignment of reported WRKY proteins in group II-c. (b) Multiple sequence alignment of reported WRKY proteins in group II-d. (c) Multiple sequence alignment of reported WRKY proteins in group II-e.
Identification of all WRKY genes in D. catenatum
-
To identify DcaWRKY genes, all the potential WRKY genes were extracted from two genome versions of D. catenatum; an old genome version with 63 WRKY genes and an updated genome version with 60 WRKY genes[48,53]. After sequences were aligned by MAFFT, all 60 genes from the updated genome version were present in the old genome version, except for three genes (Dca028175, Dca028770, and Dca027312). After verifying the sequence characteristics, Dca028175 and Dca027312 had the 'WRKYGQK' domain and a zinc finger motif. Dca028770 had the variant WRKY domain 'WRKYGKK' but no zinc finger motif. For all three genes containing the WRKY domain, the WRKY gene family members could be used for further analysis. In total, 63 candidate WRKY gene family members were identified in D. catenatum. To validate them, the amino acid sequences of all members were searched against NCBI, Pfam, and SMART databases for the presence of the WRKY domain. The results showed that all sequences contained the WRKY domain (Table 2).
Table 2. Characteristics of WRKY genes in D. catenatum.
Gene ID WRKY domain No. of
domainsNo. of
exonsNo. of
intronsGroup Conserved heptapeptide Zinc finger Zinc finger type Dca000627 WRKYGQK − − 1 3 2 II E Dca000637 WRKYGQK C2H2 CX4CX23HXH 1 2 1 II C Dca000671 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 5 4 I Dca000699 WRKYGQK C2H2 CX4CX23HXH 1 2 1 II C Dca000873 WRKYGQK C2H2 CX5CX23HXH 1 5 4 II B Dca002197 WKKYGQK C2H2 CX4CX23HXH 1 2 1 I Dca002205 WRKDGTH/WRKYATN C2H2/C2H2 CX4CX23HXH/CX4CX23HXH 2 6 5 I Dca002550 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II E Dca002715 WRKYGQK C2H2 CX4CX23HXH 1 3 2 II C Dca003067 WRKYGQK C2HC CX7CX23HXC 1 3 2 III Dca003180 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II D Dca003732 WRKYGQK C2H2 CX4CX23HXH 1 2 1 II C Dca004998 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 4 3 I Dca005043 WRKYGQK C2HC CX7CX23HXC 1 3 2 III Dca005048 WRKYGEK C2HC CX7CX23HXC 1 2 1 III Dca005648 WRKYGQK C2H2 CX4CX23HXH 1 3 2 II C Dca005780 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 5 4 I Dca006278 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II D Dca006505 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 6 5 I Dca006646 WRKYGQK C2HC CX7CX23HXC 1 3 2 III Dca006787 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II E Dca007186 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 5 4 I Dca007842 WRKYGQK C2H2 CX5CX23HXH 1 5 4 II B Dca008357 WRKYGQK C2H2 CX5CX23HXH 1 4 3 II A Dca008967 WTKYGQK C2H2 CX4CX23HXH 1 3 2 I Dca008968 WNKYGQK C2H2 CX4CX23HXH 1 2 1 I Dca008985 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 4 3 I Dca009368 WRKYGQK C2H2 CX4CX23HXH 1 3 2 II C Dca010430 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 5 4 I Dca010993 WRKYGQK C2H2 CX4CX23HXH 1 2 1 II C Dca011499 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 3 2 II C Dca011569 WRKYGQK C2H2 CX5CX23HXH 1 5 4 II B Dca011912 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II A Dca011914 WRKYGQK C2H2 CX5CX23HXH 1 4 3 II A Dca012410 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II E Dca012846 WRKYGQK C2H2 CX4CX23HXH 1 3 2 II C Dca013146 WRKYGQK C2HC CX7CX23HXC 1 3 2 III Dca013149 WRKYGQK C2HC CX7CX23HXC 1 3 2 III Dca013150 WRKYGEK C2HC CX7CX23HXC 1 3 2 III Dca014563 WRKYGQK C2HC CX7CX23HXC 1 2 1 III Dca015482 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II D Dca015639 WRKYGKK C2H2 CX4CX23HXH 1 3 2 II C Dca015848 WRKYGQK C2H2 CX4CX23HXH 1 2 1 II C Dca015914 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 3 2 II C Dca016437 WRKYGQK C2H2 CX4CX23HXH 1 2 1 II C Dca016988 WRKYGQK C2H2 CX5CX23HXH 1 2 1 II E Dca017113 WRKYGQK C2H2 CX4CX23HXH 1 3 2 II C Dca018137 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 4 3 I Dca018897 WRKYGQK C2H2 CX5CX23HXH 1 4 3 II A Dca019319 WRKYGQK C2HC CX7CX23HXC 1 3 2 III Dca019656 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II E Dca019840 WRKYGKK C2H2 CX4CX23HXH 1 3 2 II C Dca020159 WRKYGQK C2H2 CX4CX23HXH 1 2 1 II C Dca020342 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 5 4 I Dca020473 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II E Dca021638 WRKYGEK/WRKYGEK —/C2HC —/CX7CX23HXC 2 5 4 III Dca023070 WRKYGQK C2H2 CX5CX23HXH 1 3 2 II D Dca024256 WRKYGQK C2H2 CX5CX23HXH 1 2 1 II E Dca024393 WRKYGQK/WRKYGQK C2H2/C2H2 CX4CX22HXH/CX4CX23HXH 2 4 3 I Dca026708 WRKYGQK C2H2 CX4CX23HXH 1 4 3 II A Dca027312 WRKYGQK C2HC CX7CX23HXC 1 3 2 III Dca028175 WRKYGQK C2H2 CX4CX23HXH 1 3 2 II C Dca028770 WRKYGKK - - 1 1 0 II C Phylogenetic
analysis and characterization of WRKY gene family members in D. catenatum -
WRKYs in D. catenatum, Arabidopsis, and rice were subjected to phylogenetic analysis to investigate the phylogenetic relationships and potential functions of these DcaWRKY genes (Fig. 3). The WRKY genes in D. catenatum were divided into the three groups I, II, and III based on amino acid sequence similarity, which is the same as the widely accepted classification system for this gene family in Arabidopsis[6]. Among all WRKY genes in D. catenatum, the number of groups I, II, and III was 14, 38, and 11, respectively. There were five subgroups in group II, which were IIa, IIb, IIc, IId, and IIe, and the number of WRKY genes in these groups was five, three, 18, four, and eight, respectively. The results indicated that more than half of WRKY genes were clustered in group II.
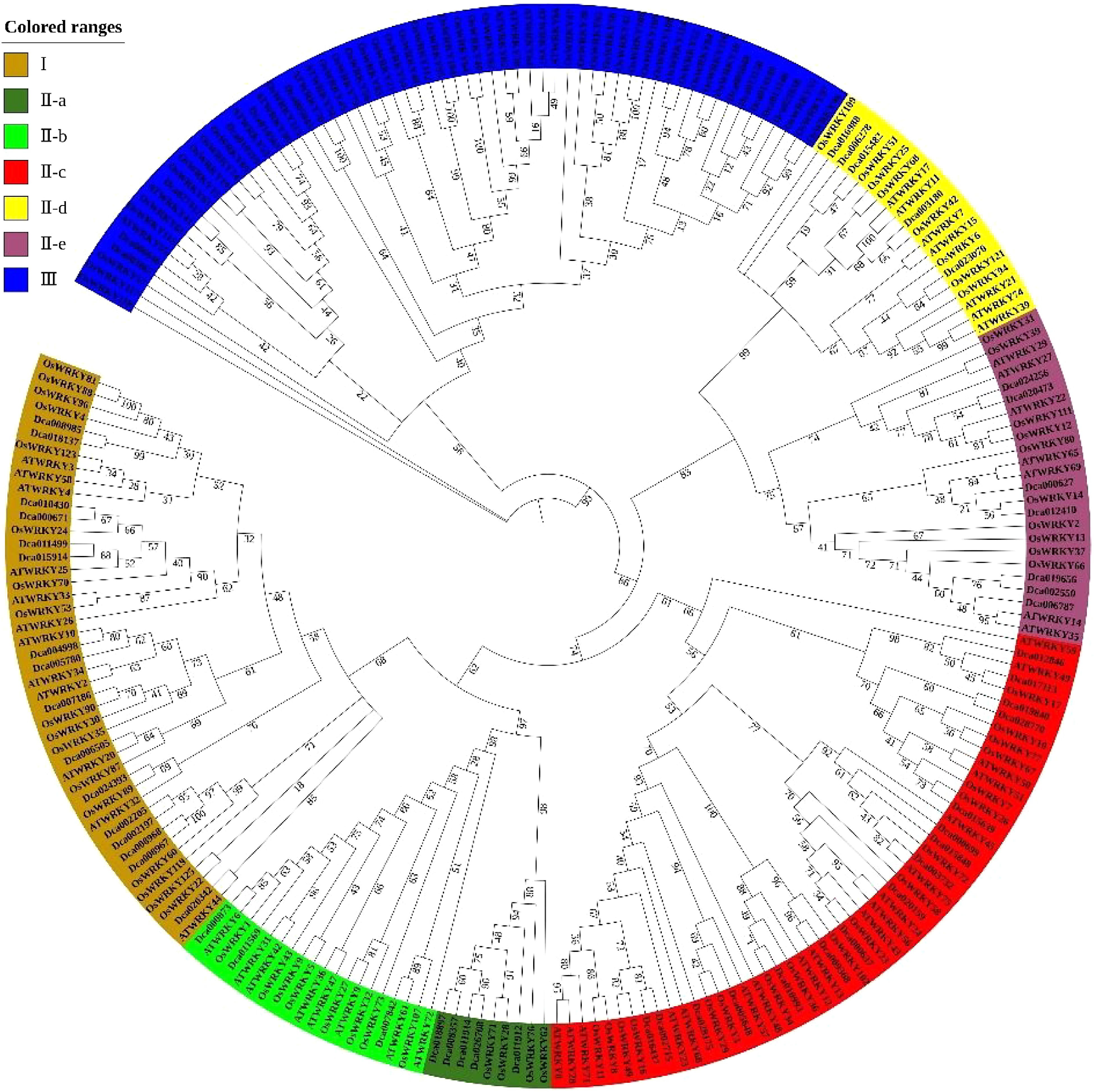
Figure 3.
Phylogenetic analysis of WRKY family proteins in D. catenatum.
All DcaWRKY proteins were analyzed by ExPASy protparam to calculate the number of amino acids, molecular weight, pI, instability index, aliphatic index, and GRAVY. The molecular weight of DcaWRKY proteins varied from 10,998.29 to 78,355.63, pI ranged from 4.57 to 10.63, and the instability index ranged from 34.57 to 76.36 (Supplemental Table S2). Analysis of DcaWRKY proteins indicated that WRKY family members in D. catenatum did not show specificity in physicochemical properties, which is the same as reported drought-resistance WRKY proteins.
To further understand the biological functions of DcWRKY genes, WRKY domain types, intron/exon distribution, and conserved motifs were analyzed according to their phylogenetic relationships. Of the 77 WRKY domains, 65 contained perfectly conserved WRKYGQK domains, whereas the other domains differed in one or more amino acids in the conserved WRKY signature. In addition, eight conserved motifs in DcaWRKY proteins were found by TBtools. To better understand the phylogenetic relationships and classification of WRKY members in D. catenatum, the intron/exon distribution was analyzed by TBtools. The number of introns in DcaWRKY genes varied from zero to five. Similarity in gene structure and sequence characteristics of WRKY members in the same branch were used to validate the reliability of the phylogenetic classification (Fig. 4).
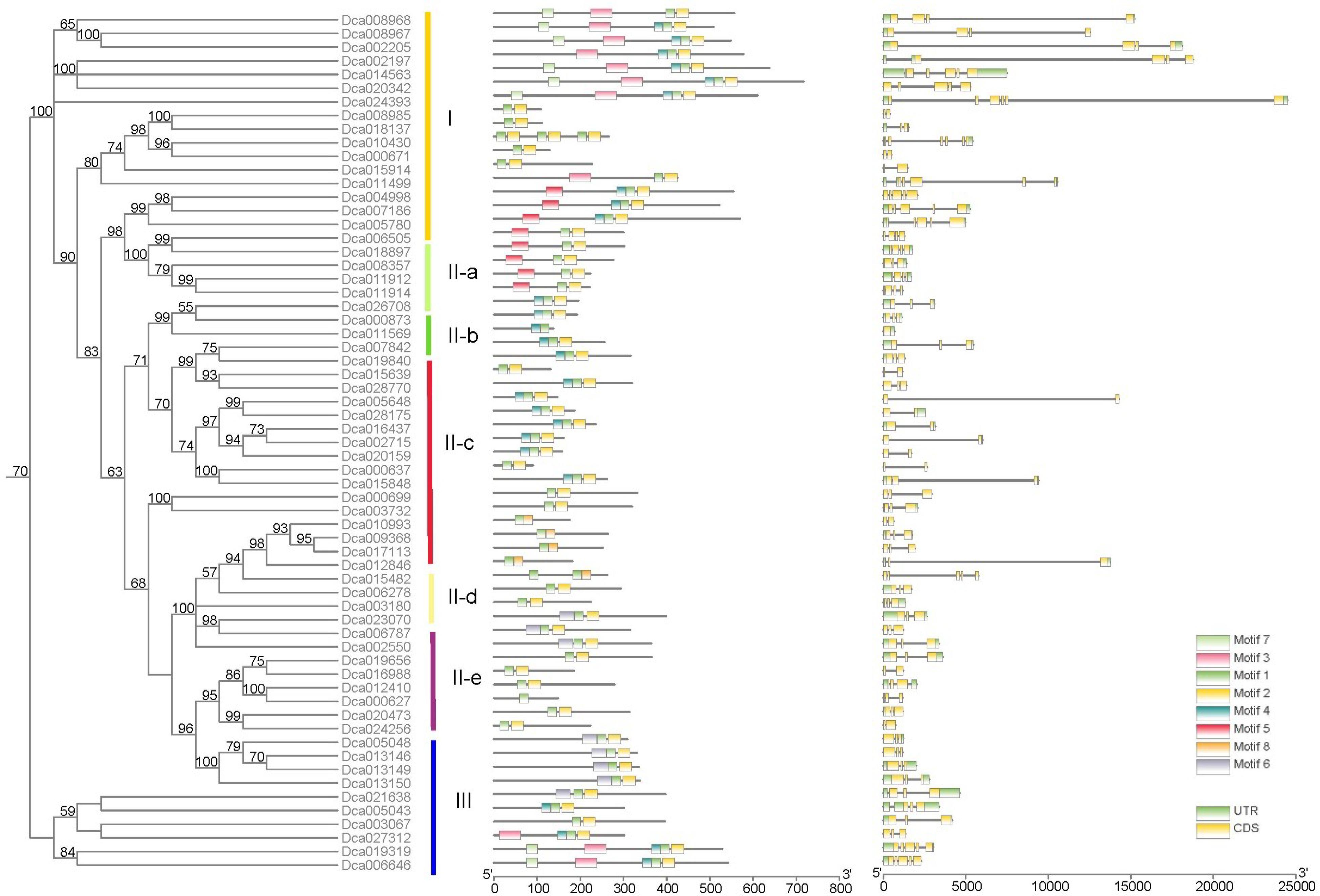
Figure 4.
Conserved motifs and gene structure of DcaWRKY genes according to phylogenetic relationships.
Group I contained two WRKY domains and two C2HC zinc finger motifs, but three DcaWRKY proteins (Dca002197, Dca008967, and Dca008968) contained only one WRKY domain and a C2H2 zinc finger motif. Besides, members of group I had five WRKY domain variant types—WKKYGQK in Dca002197, WTKYGQK in Dca008967, WNKYGQK in Dca008968, and both WRKDGTH and WRKYATN in Dca002205. Notably, the variant types WRKDGTH, WRKYATN, WTKYGQK, and WNKYGQK were first found in D. catenatum (Table 2). The number of introns in group I members widely varied from one to five. For example, Dca002205 and Dca006505 had five introns, whereas Dca002197 and Dca008968 had only one intron. Furthermore, all the DcaWRKY members of group I contained motifs 1 and 2. Interestingly, motif 7 was unique to group I (Fig. 4).
Group II proteins were close to group I proteins in the phylogenetic tree, but they contained only one WRKY domain, except for Dca011499 and Dca015914, which had two domains and two C2H2 zinc finger motifs. Besides, the only WRKY variant type WRKYGKK in group II was found in group II-c members Dca015639, Dca019840, and Dca028770 (Table 2). Similar to group I, the number of introns in group II varied from one to four except for Dca028770, which lacked introns. However, the number of introns was more focused in each subgroup of group II compared with group I. For example, groups II-a, II-b, and II-d contain three, four, and two introns, respectively. Among the three subgroups, the distribution and quantity of the motifs in all genes of the same subgroup were the same. Motifs 6, 1, and 2 were dispersed in group II-d. All members of group II-b had motifs 5, 1, 4, and 2, whereas DcaWRKY members of group II-a had motifs 5 and 2. Besides, motif 5 was uniquely dispersed in group II (Fig. 4).
All members of group III had one WRKYGQK domain, except for Dca021638, which had two WRKYGEK domains. Unlike groups I and II, the zinc finger motif of most WRKY proteins in group III was replaced by a C2HC zinc finger motif. For Dca0021638, variants of WRKYGEK/WRKYGEK or loss of a C2HC zinc finger motif might contribute to its classification in group III rather than group I (Table 2). Besides, all group III members had two exons except for Dca005048 (one), Dca014563 (one), and Dca021638 (four). The motif analysis results indicated that all members of group III contained motifs 1, 2, 6, and 8, and motif 8 was only dispersed in group III (Fig. 4).
Identification of the potential stress-responsive members of groups II-c, II-d, and II-e in D. catenatum
-
The identified stress-related amino acid pattern 'PSD-S/A/L-WAWRKYGQKPIKGSPYPR-G/S-YYRCSSSKGC' was used to identify WRKY genes of group II-e in D. catenatum. After multiple sequence alignment, Dca006787, Dca002550, Dca012410, and Dca019656 shared amino acid patterns and were identified as potential stress response genes (Fig. 5). Likewise, Dca019840, Dca005648, and Dca028770 of group II-c were also identified using the sequence pattern 'T-R/K-S/T-E/Q/D-V/I/L-E/D-I/V/H/N-L/M-D/E-D-G/E-F/Y-K/R-WRKYG-Q/K-K-A/T-VKN-S/N-P' (Fig. 5). In addition, using the 'VPA-I/V-S-X-K-M/L/V/I-ADIP-P/A/V-D-D/E-Y/F-S-WRKYGQKPIKGSP-H/Y-PRGYYKCS-S/T-V/M-RGCPARKVER' amino acid pattern, one potential gene, Dca023070, of group II-d in D. catenatum was identified as responsive to abiotic stress (Fig. 5).

Figure 5.
Multiple sequence alignment of identified DcaWRKY proteins.
Expression pattern analysis of DcaWRKY genes
-
Different WRKY genes have different tissue-specific expression patterns. To identify the accuracy of these patterns, the expression of all WRKY genes in roots and leaves of D. catenatum was separately analyzed under drought stress (Fig. 6, Supplemental Fig. S1).
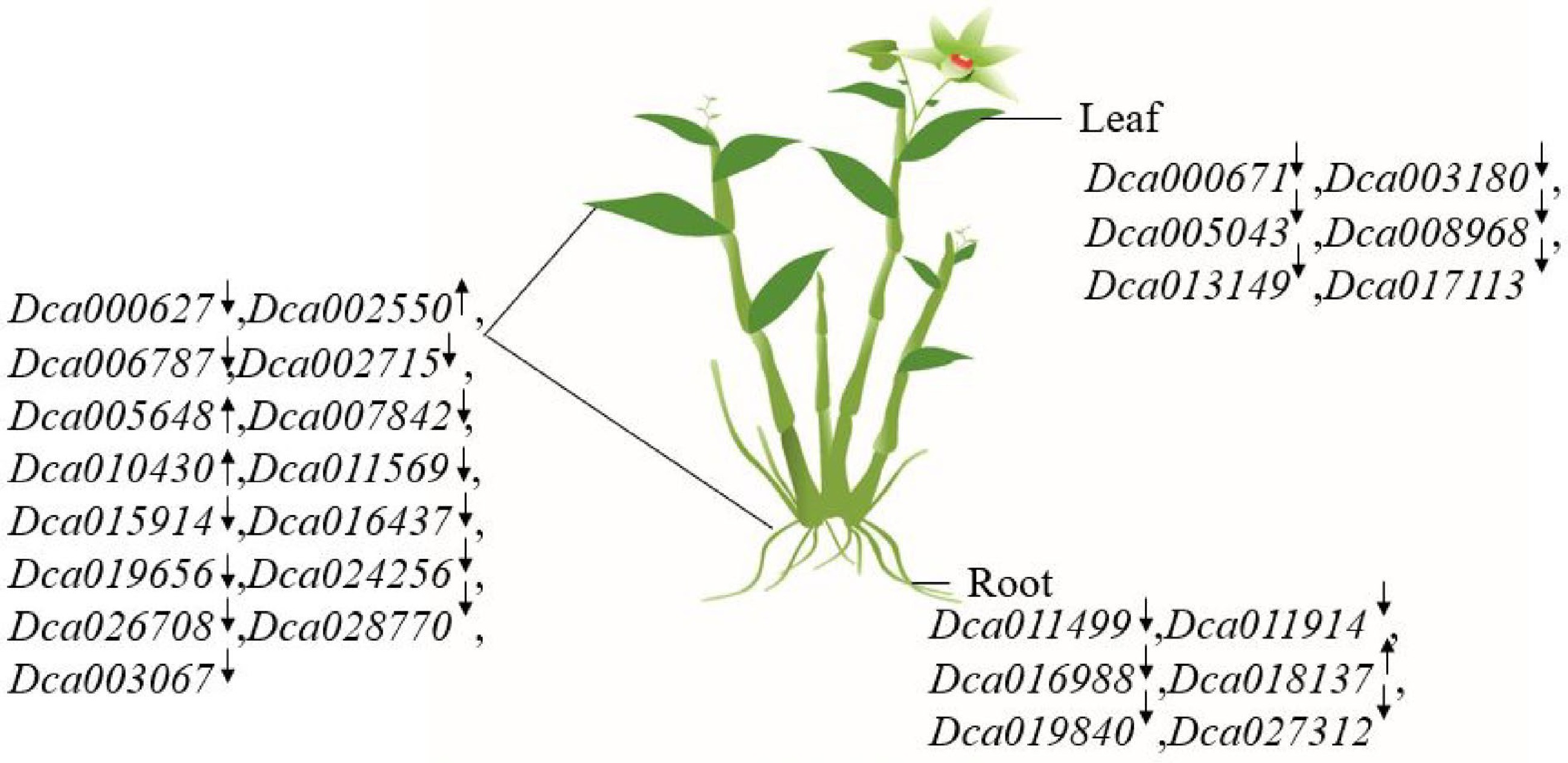
Figure 6.
The summarized figure of DcaWRKY genes expression in roots and leaves. '↑' indicates that gene expression increased under drought treatment. '↓' indicates decreased expression under drought treatment. Dca000671, Dca003180, Dca005043, Dca008968, Dca013149, and Dca017113 were differently expressed in leaves under drought treatment; similarly, Dca011499, Dca011914, Dca016988, Dca018137, Dca019840, and Dca027312 were differently expressed in roots. Dca000627, Dca002550, Dca006787, Dca002715, Dca005648, Dca007842, Dca010430, Dca011569, Dca015914, Dca016437, Dca019656, Dca024256, Dca026708, Dca028770, and Dca003067 were expressed both in leaves and roots under drought treatment.
A total of 29 differentially expressed WRKY genes in roots and leaves of D. catenatum were identified by the data. In roots, the expression of 22 DcaWRKY genes (Dca000627, Dca002550, Dca002715, Dca003067, Dca005648, Dca006787, Dca007842, Dca010430, Dca011499, Dca011569, Dca011914, Dca015914, Dca016437, Dca016988, Dca018137, Dca019840, Dca023070, Dca024256, Dca026708, Dca027312, Dca019656, and Dca028770) showed a change in drought-treated plants compared with untreated plants. Among these, most genes showed a decrease in expression, especially the Dca010430 gene, which induced a > 8.0-fold decrease in response to drought treatment. In contrast, Dca003067 and Dca018137 were upregulated and showed an opposite expression pattern (Fig. 7). In leaves, 22 WRKY members that were differentially expressed when responding to drought were considered drought-responsive genes; these included Dca000627, Dca000671, Dca002550, Dca002715, Dca003067, Dca003180, Dca005043, Dca005648, Dca006787, Dca007842, Dca008968, Dca010430, Dca011569, Dca012410, Dca013149, Dca015914, Dca016437, Dca017113, Dca024256, Dca019656, Dca026708, and Dca028770. All of these showed a decrease in their expression levels, except for Dca002550, Dca005648, and Dca010430, whose expression levels showed 2.0- to 11.0-fold increase in drought-treated plants compared with untreated plants (Fig. 8).
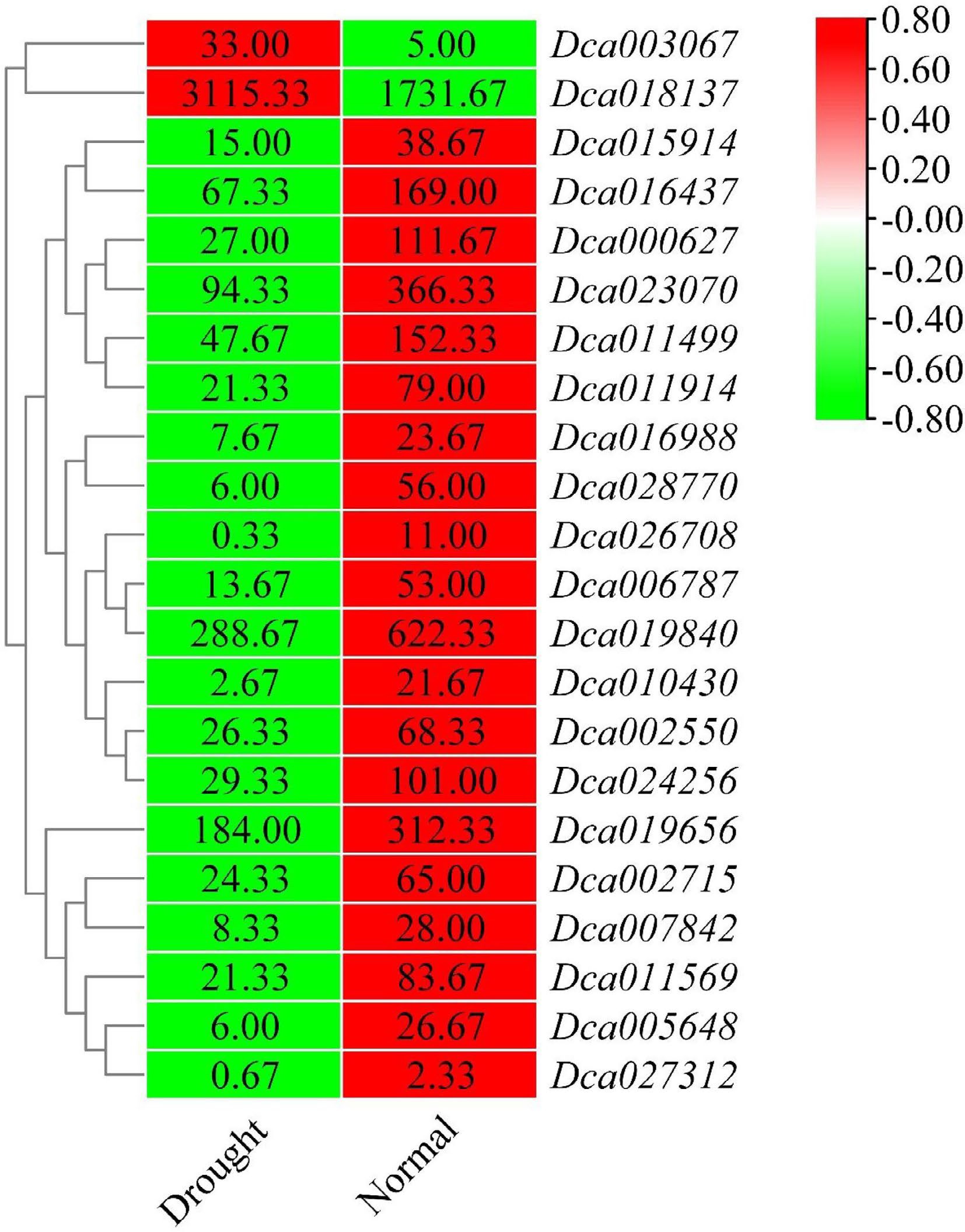
Figure 7.
Heatmap of differentially expressed DcaWRKY genes under drought stress in roots. The color scale shows increasing expression levels from green to red, which represents log2-transformed FPKM.
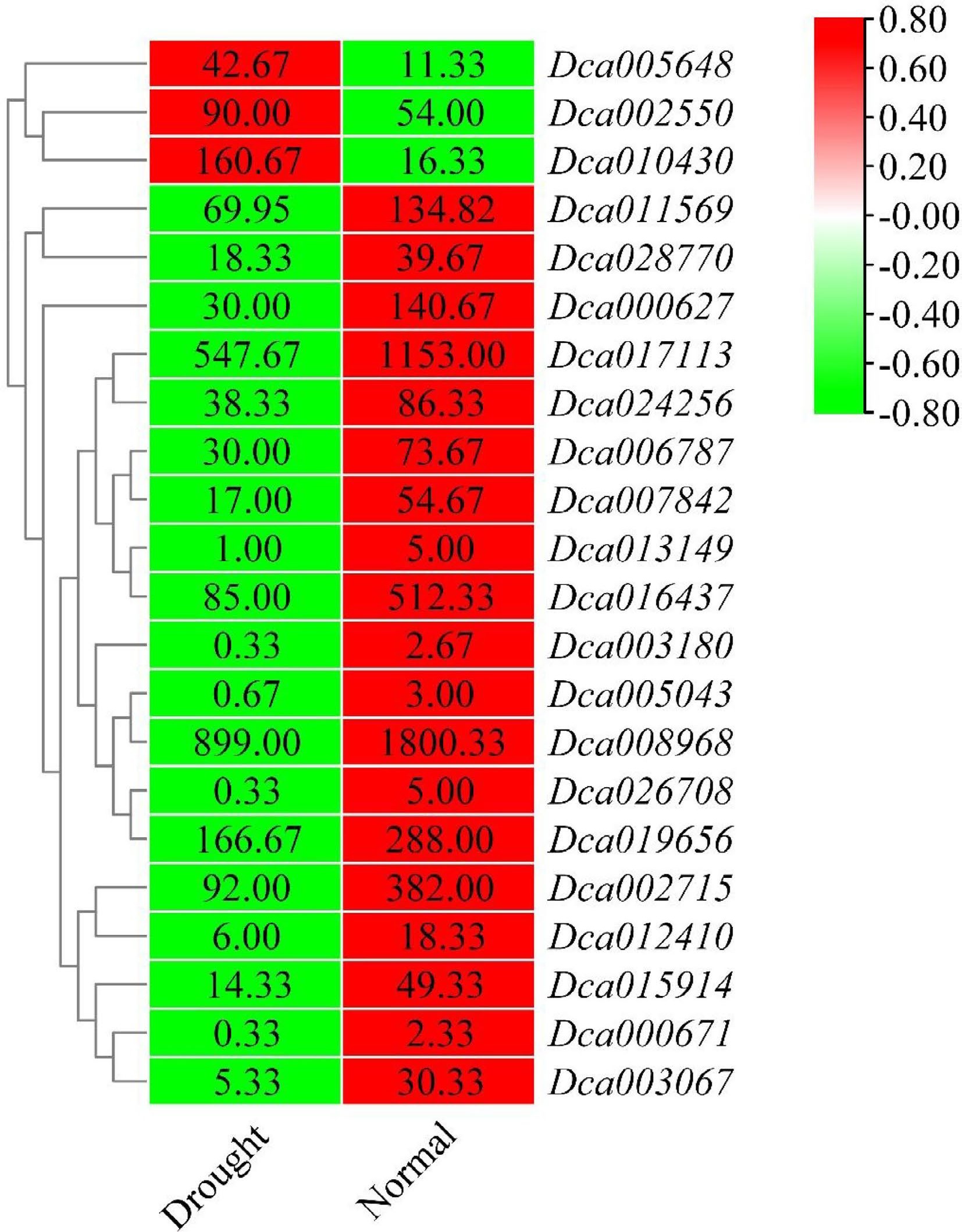
Figure 8.
Heatmap of differentially expressed DcaWRKY genes under drought stress in leaves. The color scale shows increasing expression levels from green to red, which represents log2-transformed FPKM.
A total of 15 genes in both roots and leaves showed a significant difference in expression levels when responding to drought stress, including Dca000627, Dca002550, Dca006787, Dca002715, Dca005648, Dca007842, Dca010430, Dca011569, Dca015914, Dca016434, Dca024256, Dca019656, Dca026708, Dca028770, and Dca003067. Interestingly, the identified drought-responsive gene Dca003067 showed the opposite expression patterns in the leaves and roots. Drought treatment led to a significant decrease in the expression level in leaves but an increased expression in roots.
In leaves, seven drought-responsive genes, including three genes whose expression increased and four genes whose expression decreased, were randomly selected for validation by qRT-PCR. These genes were Dca002550, Dca002715, Dca005648, Dca007842, Dca010430, Dca016437, and Dca006787 (Table 3, Fig. 9). All of them were differentially expressed in drought-treated plants compared with untreated plants. Thus, the expression patterns of these seven genes obtained by qRT-PCR were consistent with the transcriptome analysis results, confirming the reliability of the transcriptome data.
Table 3. qRT-PCR primers of DcaWRKYs.
Primer name Sequence (5'-3') Dca002550-F GTGTTCGAGCTCAACCATCA Dca002550-R TGATCGTGATCTCCCATGAA Dca005648-F GGCCGATTCACCGAATAATA Dca005648-R TTTCAACACGCTTCTTCACG Dca006787-F GCGATCTCTTTGCCTCAAAC Dca006787-R TTCCTTGCTGAGCATCCTTT Dca007842-F GCTCCTCTACCACCCATTCA Dca007842-R GTGAGGTCGAGGGTGATTGT Dca010430-F AGGAAGTCTGACGACGGCTA Dca010430-R CGAGTGGACTGAGGCTTAGG Dca016437-F ATCGTTGCACCACACAGAAG Dca016437-R AAGTCATGGTGGAAGCTTGG 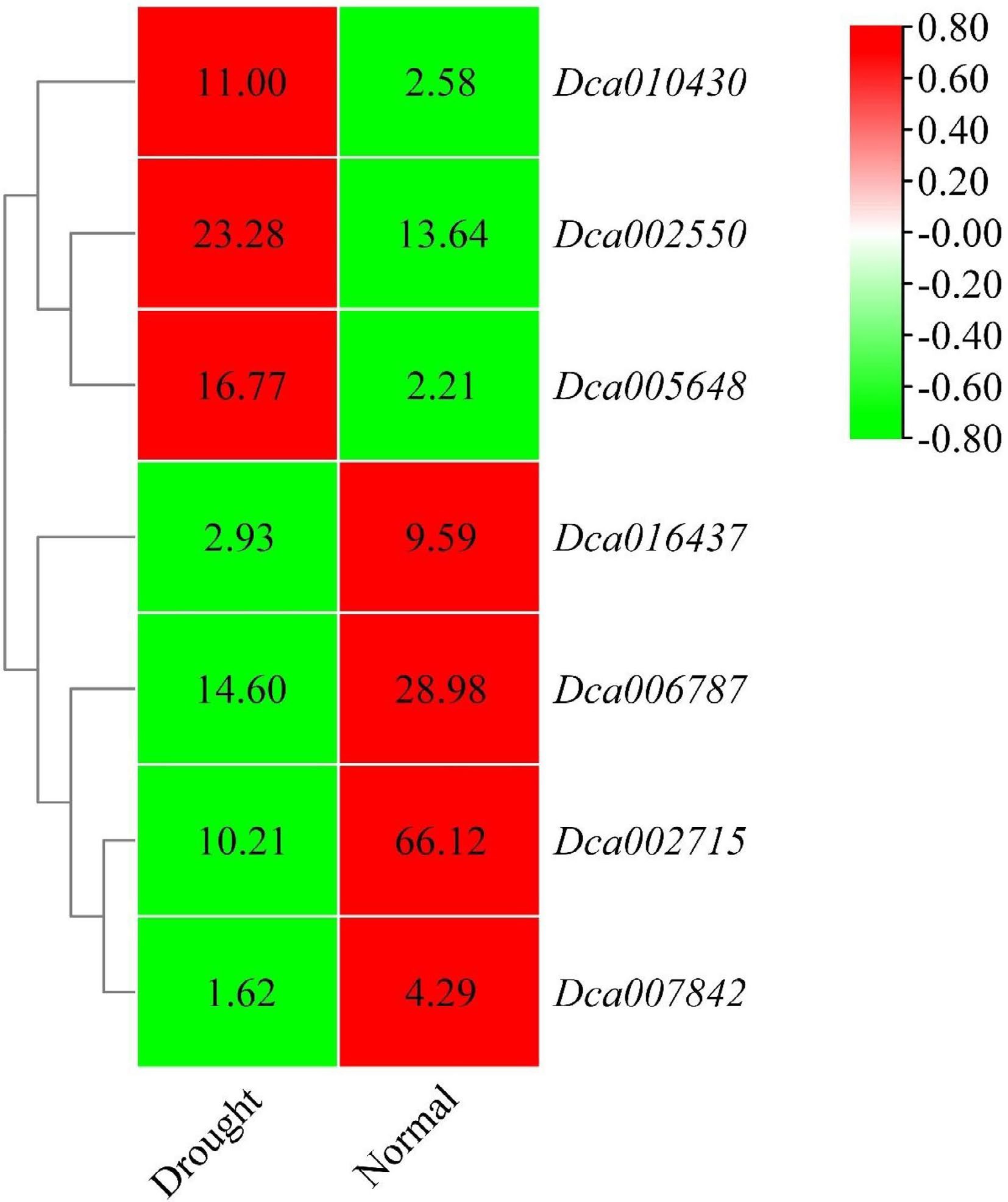
Figure 9.
Heatmap of
the selected DcaWRKY genes in leaves. The color scale shows increasing expression levels from green to red, which represents log2-transformed FPKM. -
After a comprehensive analysis of reported functional WRKY gene sequences, gene structure, and phylogenetic analysis, the amino acid patterns 'T-R/K-S/T-E/Q/D-V/I/L-E/D-I/V/H/N-L/M-D/E-D-G/E-F/Y-K/R-WRKYG-Q/K-K-A/T-VKN-S/N-P', 'VPA-I/V-S-X-K-M/L/V/I-ADIP-P/A/V-D-D/E-Y/F-S-WRKYGQKPIKGSP-H/Y-PRGYYKCS-S/T-V/M-RGCPARKVER', and 'PSD-S/A/L-WAWRKYGQKPIKGSPYPR-G/S-YYRCSSSKGC' were found in groups II-c, II-d, and II-e, respectively, which might be valid features of genes responding to stress in these three subgroups. Then, D. catenatum was used to validate these amino acid patterns. Four WRKY genes in group II-e (Dca006787, Dca002550, Dca012410, and Dca019656), three genes in group II-c (Dca019840, Dca005648, and Dca028770), and one gene in group II-d (Dca023070) were identified by homology searches. The expression of the identified DcaWRKY genes in roots and leaves was further analyzed under drought stress using qRT-PCR data. The expression levels of all potential genes identified as responsive to abiotic stress (stress under the drought treatment) were found to significantly change compared with untreated plants, suggesting that these identified amino acid patterns were valid and feasible for identifying abiotic stress in D. catenatum.
Although further studies are needed, the amino acid patterns identified in response to stress not only provide an ideal method for quickly identifying stress-related genes in D. catenatum but also offer a new perspective for the identification of functional genes in other species.
Diverse expression patterns of DcaWRKY genes in different tissues
-
In this study, DcaWRKY gene expression and the phylogeny of WRKY genes were analyzed. Various expression mechanisms of WRKY genes in D. catenatum were found under drought stress.
All differentially expressed WRKY genes in roots and leaves were comprehensively analyzed. The expression changes in WRKY genes in roots and leaves were analyzed first. Among all 29 differentially expressed WRKY genes, the expression levels of 14 DcaWRKY genes were changed only in roots or leaves when responding to drought stress. Among these 14 WRKY genes, seven were differentially expressed only in roots, including Dca011499, Dca011914, Dca016988, Dca018137, Dca019840, and Dca027312. Similarly, Dca000671, Dca003180, Dca005043, Dca008968, Dca013149, and Dca017113 were differentially expressed only in leaves. Some DcaWRKY genes might be differentially expressed only in specific tissues when responding to drought stress. Among the 14 WRKY genes, the expression levels of Dca002550, Dca005648, and Dca010430 were downregulated in leaves but were upregulated in roots in drought-treated plants compared with untreated plants. In contrast, under drought stress, Dca003067 expression was highly upregulated in leaves and downregulated in roots. Except for these three genes, all 11 genes among the 14 WRKY genes showed the same trend of expression in roots and leaves when responding to drought stress. Of the 11 DcaWRKY genes, only the expression of Dca003067, clustered in group III, was upregulated in drought-treated plants, whereas other genes were clustered in group II, and their expression levels were downregulated compared with the control. Genes with expression patterns consistent with phylogenetic branching correlations were clustered in group II; thus, we presume that the downregulation of expression levels is associated with the characteristics of group II. The molecular mechanisms of drought responses in different tissues of D. catenatum might be different.
WRKY phylogeny was systematically analyzed in Arabidopsis, rice, and D. catenatum. According to the phylogenetic tree, the members that belong to the same branch might possess a conserved function because of their common evolutionary processes. A close relationship was found between Dca016437 and Dca002715 and OsWRKY11, which was identified as an anti-drought gene in rice by the phylogenetic tree. These two genes might have the same response mechanism as OsWRKY11, whose ectopic expression resulted in constitutive expression of defense-associated genes to enhance tolerance to drought stress in rice[57].
This work was supported by the Young Talent Project of Hebei Agricultural University Foundation (YJ201848), and the Youth Fund of Hebei Province Natural Science Foundation (C2019204295). We would like to thank TopEdit (
www.topeditsci.com ) for linguistic assistance during the preparation of this manuscript.-
The authors declare that they have no conflict of interest.
- Supplemental Table. S1 The basic amino acids information of reported functionalWRKY genes.
- Supplemental Table S2 The basic amino acids information of DcaWRKY genes.
- Supplemental Fig. S1 Expression heatmap of DcaWRKY genes in roots and leaves.The color scale shows increasing expression levels from green to red, which represents log2-transformed FPKM.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhao Y, Zheng Y, Jiang L, Niu Y, Yang Y, et al. 2022. Identification of stress-related characteristics of the WRKY gene family: A case study of Dendrobium catenatum. Ornamental Plant Research 2:21 doi: 10.48130/OPR-2022-0021 

Identification of stress-related characteristics of the WRKY gene family: A case study of Dendrobium catenatum
- Received: 30 April 2022
- Accepted: 16 November 2022
- Published online: 23 December 2022
Abstract: As one of the largest families of transcription factors (TFs) in plants, the WRKY TF family plays a key role in regulating plant responses to various biotic and abiotic stresses. However, there is no confirmed method to quickly identify stress-responsive members from the WRKY gene family. In this study, all reported functional WRKY genes were first analyzed, and the amino acid patterns in response to stress were identified in group II-c (T-R/K-S/T-E/Q/D-V/I/L-E/D-I/V/H/N-L/M-D/E-D-G/E-F/Y-K/R-WRKYG-Q/K-K-A/T-VKN-S/N-P), group II-d (VPA-I/V-S-X-K-M/L/V/I-ADIP-P/A/V-D-D/E-Y/F-S-WRKYGQKPIKGSP-H/Y-PRGYYKCS-S/T-V/M-RGCPARKVER), and group II-e (PSD-S/A/L-WAWRKYGQKPIKGSPYPR-G/S-YYRCSSSKGC). WRKY genes in Dendrobium catenatum were used to validate the accuracy of these patterns. A total of 63 DcaWRKY genes were identified, their gene structures, conserved motifs, and gene expression patterns were analyzed, and a phylogenetic tree was constructed. Gene expression patterns were then analyzed under drought stress, and seven DcaWRKY genes (Dca002550, Dca002715, Dca005648, Dca007842, Dca010430, Dca016437, and Dca006787) were randomly selected to determine their expression levels and verify their expression patterns by quantitative real-time polymerase chain reaction analysis. The identified amino acid patterns were validated by drought-responsive WRKY genes in D. catenatum, confirming the accuracy of these amino acid patterns and providing valuable insights into further research of the WRKY family in D. catenatum.
-
Key words:
- WRKY genes /
- Abiotic stress /
- Biotic stress /
- Amino acid pattern /
- Dendrobium catenatum /
- Phylogenetic analysis /
- Gene expression