









-
Alternaria Nees is a ubiquitous dematiaceous hyphomycete genus, comprising over 790 species epithets, and approximately 368 species accepted within 29 sections[1−7]. Species of Alternaria occupy diverse ecological niches, from endophytes on various asymptomatic plant tissues to saprobes on a wide range of hosts and substrates (i.e., dead vegetation, paper, and food), as well as plant and animal (including human) pathogens worldwide[8−16]. The genus is cosmopolitan and widely distributed in Asia (e.g., India, Japan), Australia, Europe, and North America[17].
As invasive pathogens, Alternaria species are frequently isolated from different habitats such as the atmosphere, dust, indoor environments, soil, and damaged old buildings[11,12,18−22]. The most prevalent diseases of plants caused by Alternaria are leaf spots and defoliation with typical concentric zonatic symptoms featuring brown to black necrotic lesions surrounded by chlorotic areas on leaves[23], but can also infect flowers, fruits, roots, seedlings, and stems with different kinds of lesions[8,12,15]. These diseases reduce their market value and result in financial losses of important economic crops, such as cabbage, cucumber, fava bean, onion, potato, tomato and ornamental plants[8,11,12,15,22,24−26]. Most causal agents are restricted to Alternaria sects. Alternantherae D.P. Lawr. et al., Alternaria D.P. Lawr. et al., Brassicicola D.P. Lawr. et al., Crivellia (Shoemaker & Inderb.) Woudenb. & Crous, Gypsophilae D.P. Lawr. et al., Nimbya (E.G. Simmons) Woudenb. & Crous, Porri D.P. Lawr. et al., Radicina D.P. Lawr. et al., and Sonchi D.P. Lawr. et al. and occur on over 4,000 host and non-host specific plants[11,12,15,17,22,27,28]. Jayawardena et al.[25,26] showed that the majority of pathogenic Alternaria species infected a vast array of host species. Effective implementation of control strategies is generally hampered by misidentification. As an important plant pathogen, further details on phylogeny, diseases and symptoms, as well as morphological characters of Alternaria were also discussed by Jayawardena et al.[25,26].
Alternaria species have the ability to produce a wide spectrum of secondary metabolites. Potential phytotoxins produced by Alternaria are beneficial for biotechnological applications as biocontrol agents or mycoherbicides of innumerable plant species under diverse habitable regions[11,12,15]. Furthermore, Alternaria species also produced mycotoxins and are implicated in opportunistic animal and human diseases (e.g., alternariosis) that significantly affect the health of victims and can also contaminate food products. Alternaria alternata (Fr.) Keissl. and A. infectoria E.G. Simmons have frequently been reported as causative agents of phaeohyphomycosis in immuno-compromised patients and kidney transplant patients or airborne allergens[11,15,29−34].
Alternaria, currently belongs to Pleosporaceae of Pleosporales, Dothideomycetes[1,2], and was introduced by von Nees & Daniel[35], with A. tenuis Nees as the type species. von Keissler[36] considered A. tenuis to be conspecific with Torula alternata Fr.[37] and synonymized both A. tenuis and T. alternata with A. alternata which is currently designated as the generic type. Extensive morphology-based taxonomy of Alternaria was mainly dealt with by Simmons (1920–2013), who provided a monograph of Alternaria and recognized 275 species in the genus based on the patterns of sporulation and conidial morphology[38]. The latest taxonomic treatment of Alternaria was carried out by Lawrence et al.[13,15]. Lawrence et al.[13,15] described the asexual morph of Alternaria as alternarioid dematiaceous hyphomycetes with effuse, pigmented colonies, colorless hyphae, mononematous to caespitose, macronematous, simple or branched, pale brown to brown conidiophores, monotretic or polytretic, sympodial, conidiogenous cells, and dark pigmented, multi-celled, typically dictyosporous, or rarely phragmosporous conidia, some borne singly and most catenate chains. The sexual morph of Alternaria has only been reported for species in sects. Alternaria, Crivellia, Embellisioides Woudenb. & Crous, Eureka Woudenb. & Crous, Infectoriae Woudenb. & Crous, Nimbya and Panax D.P. Lawr. et al., and is characterized by small, dark brown, erumpent to superficial, globose to ovoid, glabrous, uni-loculate ascomata, with papillate ostioles, composed of thin-walled peridia, containing fissitunicate, cylindrical to cylindric-clavate asci, embedded in broad cellular pseudoparaphyses and muriform, ellipsoidal to fusoid, pigmented ascospores[11,15,24].
Over the course of taxonomic discussions of Alternaria, many genera have been considered to be the sexual morph of Alternaria, including Allewia E.G. Simmons, Crivellia Shoemaker & Inderb., Lewia M.E. Barr & E.G. Simmons, and Macrospora Fuckel[15,24]. Moreover, some sexual genera (viz. Clathrospora Rabenh., Comoclathris Clem., Leptosphaeria Ces. & De Not., and Pleospora Rabenh. ex Ces. & De Not.) have also been described with alternarioid asexual morphs, of which Pleospora were usually mentioned as the sexual morph of Alternaria[15]. However, Simmons[40] linked Pleospora with the asexual genus Stemphylium Wallr. Hitherto, Pleospora and Stemphylium were considered as congeneric, and Stemphylium was recommended to be used over Pleospora due to its wider use and earlier introduction[39]. Woudenberg et al.[11] demonstrated that Allewia, Brachycladium Corda, Chalastospora E.G. Simmons, Chmelia Svob.-Pol., Crivellia, Embellisia E.G. Simmons, Lewia, Nimbya E.G. Simmons, Sinomyces Yong Wang bis & X.G. Zhang, Teretispora E.G. Simmons, Ulocladium Preuss, Undifilum B.M. Pryor et al. and Ybotromyces Rulamort formed internal clades within Alternaria sensu stricto and thus these genera were synonymized and treated as sections of Alternaria. Macrospora was also considered as the sexual morph of sect. Nimbya and thus the genus was treated as a synonym of Alternaria[11,15]. The type species of Macrospora, M. scirpivora E.G. Simmons & D.A. Johnson, was synonymized under Alternaria as A. scirpivora (E.G. Simmons & D.A. Johnson), Woudenb. & Crous by Woudenberg et al.[11]. Based on the prior introduction of Alternaria, widespread use and number of the species, Rossman et al.[39] proposed to use Alternaria rather than Allewia, Crivellia and Lewia.
The DNA-based classification of the genus Alternaria has so far relied on over ten gene loci, including nuclear ribosomal DNA (LSU, SSU), the intervening ITS regions, mtSSU, protein-coding genes such as ACT, Alt-a1, CAL, GAPDH, RPB2, TEF1-α, THN, Tsr1, and the plasma membrane ATPase gene[7,11−15,25,26]. Multiple molecular methods have been investigated or proposed for distinguishing Alternaria species, including random amplified polymorphic DNA[41], amplified fragment length polymorphism[42], selective subtractive hybridization[43] and sequence characterized amplified genomic regions[44]. However, the standard gene regions and other protein-coding loci (e.g., ACT, CAM, RPB2, TEF1-α, Tsr1, TUB2 and chitin synthase) are not able to delineate species within all the sections of Alternaria, such as small spore species-groups like sect. Alternaria and sect. Infectoriae[12,45−48]. Hong et al.[49] illustrated that the Alt-a1 locus is reliable for Alternaria species identification. Lawrence et al.[14] used five protein-coding loci (viz. ACT, Alt-a1, CAM, GAPDH, and plasma membrane ATPase) for clarifying the phylogenetic hypothesis among Alternaria and revealed that the plasma membrane ATPase and CAM genes were the most suitable phylogenetic markers for molecular identification of Alternaria species. Woudenberg et al.[11] delineated phylogenetic lineages within Alternaria, and allied genera based on the multi-locus phylogeny of SSU, LSU, ITS, GAPDH, RPB2 and TEF1-α gene regions and introduced 16 new Alternaria sections. Subsequently, whole-genome sequencing has become an essential tool to delineate ambiguous species in Alternaria and other complex species[12]. Therefore, Woudenberg et al.[12] used multi-locus phylogeny based on ITS, GAPDH, RPB2, TEF1-α, Alt-a1, endoPG and OPA10-2 gene loci, coupled with whole-genome and transcriptome comparisons to discriminate species in sect. Alternaria. Lawrence et al.[15] provided a comprehensive taxonomic treatment of Alternaria with multi-locus phylogeny and accepted 27 sections in Alternaria, but later revised it to 28 accepted sections[7,15]. Recently, Gannibal et al.[6] and Ghafri et al.[7] introduced two new sections (i.e., sects. Helianthiinficiens and Omanenses) of Alternaria and thus, 29 sections were accepted[6,7,15].
Historical studies on Alternaria
-
The study of Alternaria and their allied genera has been debated for over 200 years. As summarized by Lawrence et al.[15], there are five chronological stages in the taxonomic studies of Alternaria. The first stage (1816–1850s) is when the genus Alternaria was first described in 1816, with A. tenuis as the type, but it was then confused with genera such as Macrosporium and Stemphylium. However, the first validly published species name was Torula alternata[37]. The second stage (1850s–1930s) involved publication of numerous alternarioid species, wherein Elliott[50] first attempted to revise the taxonomy and nomenclature of Alternaria and Macrosporium, but this resulted in an increasing number of nomenclatural problems within the alternarioid hyphomycetes. The third stage (1930s–1960s) includes various revisions of Alternaria made by Wiltshire[51], Neergaard[52] and Joly[53]. However, their work did not follow the rules of nomenclature, and despite wide adoption, these are not in practice to date. The fourth stage (1960s–2000s) is when Emory Guy Simmons (1920–2013) presented a complete reappraisal and revision of all names and taxa related to Alternaria, representing the most extensive compilations in the taxonomic history of the genus. The fifth stage (2000s–2015s) involved molecular phylogenetic methods to further investigate the taxonomy of Alternaria. Taxonomic studies integrating both morphological and molecular data were provided by Pryor & Gilbertson[54], Hong et al.[49], Lawrence et al.[13,14,55], Woudenberg et al.[11,12,22] and Grum-Grzhimaylo et al.[56]. In subsequent studies, the utility and reliability of different genes in deciphering phylogenetic relationships have been discussed by Woudenberg et al.[12] and Lawrence et al.[15].
The Alternaria sections
-
Alternaria sections are recognized based on molecular phylogenies, but these do not always correlate with species-groups that were earlier delineated based on morphological characteristics (Table 1)[11,13–15,22,56]. The species-groups A. alternata, A. alternantherae, A. brassicicola, A. infectoria, A. porri, A. radicina and A. sonchi were phylogenetically strongly supported by Chou & Wu[57], De Hoog & Horré[20], Hong et al.[49], Inderbitzin et al.[58], Lawrence et al.[14,55], Pryor & Bigelow[59], Pryor & Gilbertson[54], Pryor et al.[60], Runa et al.[21], Wang et al.[61], and Woudenburg et al.[11,12]. Lawrence et al.[14] introduced A. panax and A. gypsophilae as two species-groups and proposed eight species-groups to sections within Alternaria. The latest treatment of Alternaria were carried out by Lawrence et al.[15] who generalized the genus with 27 sections. Recently, Ghafri et al.[7] included sect. Omanenses Al Ghafri et al. to the genus. While Gannibal et al.[6] introduced a new section, sect. Helianthiinficientes, for A. helianthiinficiens which was previously demonstrated as a monotypic lineage in Woudenberg et al.[11] and Lawrence et al.[15].
Table 1. Synopsis of Alternaria sections based on the asexual morphs.
Alternaria sections Conidiogenesis structures Ecology and economy References Sect. Alternantherae Conidiophores Short to moderately long, with slightly enlarged conidiogenous tip. Species in this section are reported as plant pathogens that mainly cause leaf spots. [11,13,15,55] Conidia Large, ellipsoidal to ovoid, or subcylindrical, rarely narrow ellipsoidal, solitary or rarely paired, disto- and euseptate, transversely septate with no or 1–2 longitudinal or oblique septa, slightly constricted near some septa, with a long apical narrow beak, conidial beak unbranched, septate or aseptate, long filiform, sometimes swollen at the end, internal compartmentation occurs, with cell bright at end, with hexagonal, octagonal or rounded transverse sections lumina. Sect. Alternaria Conidiophores Short to long, straight or curved, simple or branched, with one or several apical conidiogenous loci. Species in this section are reported as plant pathogens on leaves, stems and fruits, and vegetables. Some species cause opportunistic infections of humans. Species in this section are also reported as resources of potential toxins and secondary metabolites. [11,12,15,100] Conidia Obclavate to long ellipsoid, small or moderate in size, septate, slightly constricted near some septa, with few longitudinal septa, in moderately long to long, simple or branched chains, form tapered beak or secondary conidiophore with one or a few conidiogenous loci. Sect. Brassicicola Conidiophores Short to moderately long, simple or branched, with one or several apical conidiogenous loci. Species in this section mainly cause black spot disease on a wide range of hosts, particularly on Brassica spp. such as cabbage, Chinese cabbage, cauliflower, oilseeds, broccoli and canola. Species in this section are also reported as sources of antibiotic masses. [11,14,101−103] Conidia Ellipsoid, ovoid or somewhat obclavate, small or moderate in size, septate, slightly or strongly constricted at most of the transverse septa, with or without longitudinal septa, in moderately long to long, simple or branched chains, with dark septa and cell walls. Apically or laterally form secondary conidiophores with one or a few conidiogenous loci. Sometimes produced chlamydospores. Sect. Chalastospora Conidiophores Short to long, simple or branched, with one or several conidiogenous loci. Species in this section are primarily reported as saprobes and causal agents of human diseases. [11,30,38] Conidia Pale to medium brown, narrowly ellipsoidal to ellipsoidal or ovoid, beakless, with no or multiple transverse eusepta and rarely longitudinal septa, solitary or in chains. Apically or laterally form secondary conidiophores with one or a few conidiogenous loci. Sect. Cheiranthus Conidiophores Short to moderately long, simple or branched, with one or several conidiogenous loci. Species in this section are primarily saprobes and pathogens on various plant hosts. [11,38,55] Conidia Ovoid, broadly ellipsoid with transverse and longitudinal septa, slightly or strongly constricted at the septa, in short to long, simple or branched chains. Sect. Crivellia Conidiophores Straight or curved, simple or branched, with geniculate, sympodial proliferations. Species in this section are mainly known as pathogens on opium poppy (Papaver somniferum L.), the sexual morph of which links with genus Crivella. [11,58] Conidia Cylindrical, straight to curved to inequilateral, with transverse septa, rarely constricted at septa, single or in short, simple or branched chains. Apically or laterally form secondary conidiophores. Sometimes produced microsclerotia or chlamydospores. Sect. Dianthicola Conidiophores Simple or branched, with or without apical geniculate proliferations. Species in this section mainly cause leaf spot and blight on economic vegetation hosts such as carnation (Dianthus sp.) and sesame (Sesamum indicum L.). [11,104,105] Conidia Narrowly ovoid or narrowly ellipsoid with transverse and few longitudinal septa, slightly constricted at the septa, with a long (filamentous) beak or apical secondary conidiophore, solitary or in short chains. Sect. Embellisia Conidiophores Simple, septate, straight or with geniculate sympodial proliferation. Species in this section are reported as pathogens on vegetable crops such as tomato and garlic. [11,106,107] Conidia Solitary, ovoid to subcylindrical, straight to inequilateral, with transverse septa; septa can be thick, dark and rigid in contrast to the external wall. Sometimes sporulated chlamydospores. Sect. Embellisioides Conidiophores Simple, septate conidiophores, straight or with multiple, geniculate, sympodial proliferations. Species in this section are mainly reported as saprobes in soil and pathogen on plant hosts. [9,108,109] Conidia Solitary or in short chains, obovoid to ellipsoid, with transverse and longitudinal septa, transverse septa can be thick, dark and rigid in contrast to the external wall. Apical or lateral, short secondary conidiophores may occur. Sometimes produced sexual morph and chlamydospores. Sect. Eureka Conidiophores Simple, septate conidiophores, straight or with geniculate, sympodial proliferations. Species in this section are reported as pathogens and endophytes that are active in the biotransformation of some secondary metabolites. [11,110,111] Conidia Solitary or in short chains, narrowly ellipsoidal to cylindrical, with transverse and longitudinal septa, slightly constricted at the septa, with a blunt rounded apex. Sometimes form apical or lateral, short secondary conidiophores and sporulated sexual morph and chlamydospores. Sect. Euphorbiicola Conidiophores Short to long, broad, apical and sometimes lateral, secondary conidiophores. Species in this section served as pathogens on economic plants such as Euphorbiicola sp. and Citrus sp. and also produced secondary metabolites. [11,112] Conidia Medium to large-sized, in short to moderately long chains, ovoid, obclavate, disto- and euseptate, with multiple transverse and some longitudinal septa, slightly constricted near some transverse septa, with no or a simple long beak in the terminal conidia. Sect. Gypsophilae Conidiophores Simple, or occasionally branched, with one or a few conidiogenous loci. Species in this section occur on the host family Caryophyllaceae. [11,14,38] Conidia Solitary or in short chains, ellipsoid to long ovoid, with multiple transverse and longitudinal septa, conspicuously constricted near some transverse septa. Apically form secondary conidiophores with one or two conidiogenous loci or laterally with a single conidiogenous locus. Sect. Helianthiinficientes Conidiophores Simple, or branched, with one or a few conidiogenous loci. Species in this section is well-known as a pathogen on sunflower and cosmos, and also associated with some other species in Asteraceae (i.e., Arctium sp. and Sonchus sp.). [6] Conidia Solitary or in short chains, large, narrowly or broadly ovoid, or ellipsoidal, with several transverse and longitudinal septa, constricted near septa, sometimes non-beaked. Apically form secondary conidiophores, or a few lateral secondary conidiophores, or short to very long filiform beak. Sect. Infectoriae Conidiophores Short to long, simple or branched, with one or several conidiogenous loci. Species in this section are known as saprobes as well as plant and human pathogens. [11,14,38,
70]Conidia Moderately long to long, branched chains, small or moderate sized, obclavate to long ellipsoidal, septate, slightly constricted near some septa, with few longitudinal septa. Apically or laterally formed long geniculate, multi-locus secondary conidiophores, with meristematic growth. Sect. Japonicae Conidiophores Short to long, simple or occasionally branched, with a single conidiogenous locus. Species in this section particularly occur on hosts in Brassicaceae. [11,14] Conidia Short to long ovoid with transverse and longitudinal septa, conspicuously constricted at most of the transverse septa, in short chains. Apically formed secondary conidiophores with single conidiogenous locus. Sect. Nimbya Conidiophores Simple, short to form moderately long, sometimes one to a few short to long, geniculate, sympodial metastasis. Species in this section are known as saprobes and plant pathogens. Species in this section produce phytotoxins [11,55,85,
113,114]Conidia Solitary or in short chains, narrowly elongate-obclavate, gradually tapering apically, with transverse disto- and eusepta, sometimes slightly constricted near eusepta. Sect. Omanenses Conidiophores Long, simple, with multiple geniculate, sympodial metastasis or short conidiogenous loci normally with a terminal cluster of three conidia. Species in this section consist of a core taxon A. omanensis which is saprobic on dead woods. [7] Conidia Solitary, obovoid and sphaeroid, non-beaked, with transverse and longitudinal septa. Sect. Panax Conidiophores Simple or branched, short to moderately long, with one or a few conidiogenous loci. Species in this section are known as pathogens causing blight on economic plants such as ginseng and American ginseng (Araliaceae). [11,14,115] Conidia Solitary, simple or branched, in short chains, obclavate to ovoid, with multiple transverse and longitudinal septa, conspicuously constricted near several transverse septa, apically formed secondary conidiophores with one or several conidiogenous loci, multiple lateral secondary conidiophores with a single conidiogenous locus. Sect. Phragmosporae Conidiophores Simple, short to moderately long, with one or multiple geniculate, sympodial metastasis. Species in this section are mainly known as saprobes from soil and marine environments. [11] Conidia Solitary or in simple short chains, broadly ovoid to long ovoid, ellipsoidal, curved, or limaciform, with multiple transverse and few to multiple longitudinal septa, some septa darkened, slightly to conspicuously constricted near several transverse septa, apically formed secondary conidiophores with one or several conidiogenous loci. Sect. Porri Conidiophores Short to long, simple, with one or several conidiogenous loci. Species in this section consist of some important phytopathogens and produce phytotoxins. [11,14,22,116,117] Conidia Solitary or in short to moderately long chains, with a simple or branched, long to filamentous beak, medium or large size, broadly ovoid, obclavate, ellipsoid, subcylindrical or obovoid, disto- and eusepta, with multiple transverse and longitudinal septa, slightly constricted near some transverse septa, apically or laterally formed secondary conidiophores. Sect. Pseudoalternaria Conidiophores Simple or branched, septate, smooth, medium brown, simple with a single apical pore, with short to long, simple to multi-geniculate secondary conidiophores with one to many conidiogenous loci. Species in this section are known as pathogens on plant hosts. [15] Conidia Mostly catenulate, ellipsoid to obclavate, medium brown to golden brown, with several transverse and longitudinal septa, smooth, secondary conidiophore may occur as a false beak. Sect. Pseudoulocladium Conidiophores Simple or branched, with short, geniculate, sympodial metastasis. Species in this section are reported as phytopathogens for human infection. [11] Conidia Obovoid, non-beaked with a narrow base, in simple or mostly branched chains, apically formed secondary conidiophores with multiple conidiogenous loci and laterally secondary conidiophores may occur with a single conidiogenous locus. Sect. Radicina Conidiophores Straight, simple or branched, short or long, with multiple, short geniculate, sympodial proliferations, with one to a few conidiogenous loci at the apex. Species in this section mainly occur on hosts in family Apiaceae. [11] Conidia Solitary or in short chains, moderate in size, broadly ovoid to narrowly ellipsoidal, beakless, with several transverse and longitudinal septa, apically formed solitary, short, secondary conidiophores. Sect. Soda Conidiophores Simple or occasionally branched, short to moderately long, with one conidiogenous locus. Species in this section are isolated from soda lake environments (Western Siberia, Russia). [56] Conidia Solitary or in short to long, simple or branched chains, moderate to very large in size, narrowly ellipsoid to elongate-ovoid or somewhat obclavate, septate, with transverse and longitudinal septa, conspicuously constricted at most of the transverse septa, produced microsclerotia or chlamydospores, apical or lateral short secondary conidiophores with a single conidiogenous locus may occur, and conidiogenous tip can be enlarged. Sect. Sonchi Conidiophores Simple or branched, with short, geniculate, with one or several conidiogenous loci. Species in this section mainly occur on a wide range of hosts within Asteraceae (Compositae). [14] Conidia Single or in short chains, medium to large size, subcylindrical, broadly ovoid, broadly ellipsoid or obclavate, with multiple transverse and few longitudinal septa, slightly constricted at the septa. Sect. Teretispora Conidiophores Simple, sometimes extending at the apex with one or two, geniculate, sympodial proliferations. Species in this section consist of a core species, Alternaria leucanthemi, which is a phytopathogen causing plant blight disease. [11,38] Conidia Single, long cylindrical, lacking a beak portion, with many transverse and a few longitudinal septa, constricted at most of the transverse septa, secondary conidiophores with single conidium from the base of primary conidium and rarely formed apically. Sect. Ulocladioides Conidiophores Short, geniculate, sympodial proliferations. Species in this section are mainly known as phytopathogens causing leaf spot disease and can be saprobes on a variety of host substrates as well as a causal agent of keratitis. [11,15] Conidia Obovoid, non-beaked with a narrow base, single or in chains, with apical secondary conidiophores. Sect. Ulocladium Conidiophores Simple, with one or two short, geniculate, sympodial proliferations. Species in this section are mainly isolated from plant litter and rarely from marine environments. Potential bioactivities were also reported. [11,118] Conidia Single, obovoid, non-beaked, with a narrow base. Sect. Undifilum Conidiophores Simple, septate, straight, or with geniculate sympodial proliferation. Species in this section mainly occur on hosts in family Fabaceae. [11] Conidia Ovate to obclavate to long ellipsoid, straight to inequilateral, single, transverse septa, septa can be thick, dark and rigid, and form unique germ tubes, which are wavy or undulate until branching. Presently, Alternaria contains 29 sections viz. sect. Alternantherae, sect. Alternaria, sect. Brassicicola, sect. Chalastospora, sect. Cheiranthus, sect. Crivellia, sect. Dianthicola, sect. Embellisia, sect. Embellisioides, sect. Euphorbiicola, sect. Eureka, sect. Gypsophilae, sect. Helianthiinficientes, sect. Infectoriae, sect. Japonicae, sect. Nimbya, sect. Omanenses, sect. Panax, sect. Phragmosporae, sect. Porri, sect. Pseudoalternaria, sect. Pseudoulocladium, sect. Radicina, sect. Soda, sect. Sonchi, sect. Teretispora, sect. Ulocladioides, sect. Ulocladium, and sect. Undifilum. Furthermore, seven species identified in Alternaria by multi-locus phylogenetic analyses and not accommodated among the 29 accepted sections of Alternaria are A. argyranthemi E.G. Simmons & C.F. Hill, A. brassicae (Berk.) Sacc., A. dennisii M.B. Ellis, A. peucedani S.H. Yu, A. soliardae E.G. Simmons, A. thalictrigena K. Schub. & Crous, and A. thlaspis (E.G. Simmons & J.C. David) D.P. Lawr., Rotondo & Gannibal[15].
Section Alternantherae was introduced by Lawrence et al.[14] for species group Alternaria alternantherae Holcomb & Antonop., comprising three species previously described as Nimbya species viz. A. celosiicola Jun. Nishikawa & C. Nakash., A. gomphrenae Togashi and A. perpunctulata (E.G. Simmons) D.P. Lawr., M.S. Park & B.M. Pryor, and the type species of the section, A. alternantherae[11,15]. Subsequently, the other three species were included in the sect. Alternantherae viz. A. crassoides (Davis) Gannibal, A. pimpriana V.G. Rao, and A. paragomphrenae Jun. Nishikawa & C. Nakash. that A. crassoides and A. pimpriana were previously accommodated in Nimbya[62−64]. Currently, seven species are accepted in this section.
Section Alternaria was introduced by Lawrence et al.[14] to accommodate Alternaria species, commonly referred to small-spored Alternaria groups. The main morphological feature that can be used to distinguish Alternaria sect. Alternaria from other sections is the short conidia produced in chains (frequently less than 60 µm in vitro)[11,14,65]. The sexual morph is known from A. alternata and described as typically small-sized, erumpent, globose to ovoid, smooth, dark brown, papillate ascomata, cylindrical to cylindric-clavate asci, and ellipsoid to fusoid, brown, eguttulate, smooth-walled ascospores, with 3–7 transverse septa, 1–2 longitudinal septa[15,24]. There were approximately 60 species accommodated in section Alternaria based on ITS sequence data[11]. However, Woudenberg et al.[12] accepted only 11 phylogenetic species and one species complex in this section. Gannibal[65] re-circumscribed and amended the section based on morphological assessments by Simmons[38]. Gannibal[65] included the other 37 morpho-species and accepted 59 species in this section. Subsequently, the other four species were included in this section by Gannibal & Lawrence[62] viz. A. calystegiae Nelen, A. diversispora (Thüm.) E.G. Simmons, A. guaranitica (Speg.) E.G. Simmons and A. macalpinei E.G. Simmons). Wanasinghe et al.[66] introduced A. doliconidium J.F. Li, Camporesi & K.D. Hyde on Rosa canina in Italy. Jayawardena et al.[67] also introduced A. italica J.F. LI, Camporesi & K.D. Hyde on Vitis vinifera in Italy. Nishikawa & Nakashima[63] also included A. iridicola (Ellis & Everh.) J.A. Elliott in this section. In 2022, Li et al.[68] introduced six saprobic species from Italy in this section (i.e., A. muriformispora J.F. Li et al., A. obpyriconidia J.F. Li et al., A. ovoidea J.F. Li et al., A. pseudoinfectoria J.F. Li et al., A. rostroconidia J.F. Li et al., and A. torilis J.F. Li et al.). In addition, Gou et al.[69] also introduced two Alternaria species as pathogens causing leaf spot or blight symptoms on Iris plants in China viz. A. setosae Y.N. Gou & J.X. Deng, and A. tectorum Y.N. Gou & J.X. Deng. Therefore, 73 species are now accommodated in this section.
Section Brassicicola was introduced by Lawrence et al.[14] for the species-group Alternaria brassicicola (Schwein.) Wiltshire. The section comprises five species viz. A. brassicicola, A. conoidea (E.G. Simmons) D.P. Lawr. et al., A. mimicula E.G. Simmons, A. septorioides (Westend.) E.G. Simmons, and A. solidaccana E.G. Simmons[11,15]. Multi-locus phylogenetic analyses demonstrated that sect. Brassicicola has close phylogenetic relationships with sects. Sonchi, Radicina, Gypsophilae, Porri, Alternaria, and Alternanatherae[11]. However, the conidial morphology of sect. Brassicicola is different from these sections in producing extremely small phragmosporous conidia with heavily melanized transverse septa[11,14,15].
Section Chalastospora was introduced by Woudenberg et al.[11] for a species group that was previously described as Chalastospora species. The section is typified by Alternaria cetera E.G. Simmons, and the other five species were also initially accommodated in this section, including A. abundans (E.G. Simmons) Woudenb. & Crous, A. armoraciae E.G. Simmons & C.F. Hill, A. breviramosa Woudenb. & Crous, A. malorum (Ruehle) U. Braun, Crous & Dugan, and A. obclavata (Crous & U. Braun) Woudenb. & Crous[11]. Interestingly, A. abundans and A. armoraciae can be distinguished from the other species in sect. Chalastospora by having mostly phragmoconidia that are short and not elongated as in other species of this section[15]. Marin-Felix et al.[70] included A. pobletensis Iturrieta-González, Dania García & Gené in this section and thus, seven species are listed in sect. Chalastospora.
Section Cheiranthus was introduced by Woudenberg et al.[11] to accommodate Alternaria cheiranthi (Lib.) P.C. Bolle, and A. indefessa (E.G. Simmons) Woudenberg & Crous (≡ Embellisia indefessa E.G. Simmons). Woudenberg et al.[11] treated a non-sporulating strain CBS 115.44 which was formally identified as A. resedae Neerg., in this section. However, A. resedae was treated as a synonym of A. septorioides E.G. Simmons in sect. Brassicicola. Thus, Woudenberg et al.[11] treated the strain CBS 115.44 as 'Alternaria sp.' Gannibal & Lawrence[62] assigned A. latifunda E.G. Simmons to this section based on morphology with conidia having many longitudinal septa. Hence, three species are accepted in this section[62]. Phylogenetic analyses demonstrated this section is sister to sects. Pseudoulocladium and Ulocladioides[11,14,15].
Section Crivellia was introduced by Woudenberg et al.[11] to accommodate the type species of Crivellia, C. papaveracea (De Not.) Shoemaker & Inderb. (asexual morph known as Brachycladium penicillatum Corda), and B. papaveris (Sawada) Shoemaker & Inderb. Both species are important pathogens of opium poppy[15]. Phylogenetic analyses based on ITS, GAPDH and TEF1-α sequences revealed that these two species clustered with the Alternaria-complex instead of Pleospora sensu stricto. Hence, Woudenberg et al.[11] transferred these two species to the new section of Alternaria as A. papavericola Woudenb. & Crous and A. penicillata (Corda) Woudenb. & Crous. However, Lawrence et al.[15] mentioned that the phylogenetic status of this section is uncertain. The sexual morph of A. penicillata was interdispersed with dark microsclerotia and macroconidiophores, forming medium-sized (320–400 × 220–300 μm), globose to depressed globose ascomata, with ellipsoidal ascospores (20–25 × 6–9 μm)[15].
Section Dianthicola was introduced by Woudenberg et al.[11] and is typified by Alternaria dianthicola Neerg. Three species were accommodated in this section, including A. dianthicola, A. elegans E.G. Simmons & J.C. David, and A. simsimi E.G. Simmons[11]. Xu et al.[71] introduced another pathogenic species, A. kareliniae B. Xu & Z.D. Jiang, causing leaf spot on Karelinia caspia (Pall.) Less. in China. However, the name was validly listed in Index Fungorum[72]. Thus, four phylogenetic species are known in this section. Phylogenetic analyses based on protein-coding genes showed that sect. Dianthicola has a close relationship with sect. Ulocladioides[11,15].
Section Embellisia was introduced by Woudenberg et al.[11] and is typified by Alternaria embellisia Woudenb. & Crous (≡ Helminthosporium allii Campan.). The section was established for the species previously described in Embellisia, including three species viz. E. allii E.G. Simmons, E. chlamydospora (Hoes, G.W. Bruehl & C.G. Shaw) E.G. Simmons, and E. tellustris E.G. Simmons. Embellisia was initially introduced to separate an atypical species of Helminthosporium Link[73] based on conidial and conidiophore morphology which is characterized by successive sympodial proliferations conidiophores and phragmoconidia, with distinctly dark, rigid and thickened transverse septa[15]. Phylogenetic analyses based on GAPDH, ITS and Alt-a1 genes demonstrated that the section has close relationships with sects. Phragmosporae, Soda, Chalastospora, Pseudoalternaria, and Infectoriae[11,15]. Woudenberg et al.[11] therefore, designated the new name for these three Embellisia species and transferred them to Alternaria sect. Embellisia, namely Alternaria chlamydosporigena Woudenb. & Crous, A. embellisia Woudenb. & Crous, and A. tellustris (E.G. Simmons) Woudenb. & Crous.
Section Embellisioides was introduced by Woudenberg et al.[11] to accommodate six species previously described as Embellisia species and named as Embellisia group III in Lawrence et al.[55]. The section consists of Alternaria botryospora Woudenb. & Crous, A. hyacinthi (de Hoog & P.J. Mull. bis) Woudenb. & Crous (type species), A. lolii (E.G. Simmons & C.F. Hill) Woudenb. & Crous, A. planifunda (E.G. Simmons) Woudenb. & Crous, A. proteae (E.G. Simmons) Woudenb. & Crous, and A. tumida (E.G. Simmons) Woudenb. & Crous[11,15]. These species were obtained from plants or the rhizosphere[15]. The sexual morph of species in this section was regarded as Allewia species and characterized by ovoid to spherical, dark, thin-walled, pseudothecial, papillate ascomata with markedly setose, subellipsoidal to subcylindrical asci and slightly inequilateral subellipsoidal immature ascospores. Mature ascospores are ellipsoid to subclavate, with multiple transverse septa and a discontinuous series of longitudinal septa[15]. Phylogenetic analyses supported the section as a sister group with sect. Eureka[11,15].
Section Euphorbiicola was introduced by Woudenberg et al.[22] and is typified by Alternaria euphorbiicola E.G. Simmons & Engelhard. Two species are currently accommodated in this section viz. A. euphorbiicola and A. limicola E.G. Simmons & M.E. Palm[22]. These two species were obtained from plant host families Euphorbiaceae and Rutaceae as saprobes and pathogens[17,22]. Woudenberg et al.[22] established sect. Euphorbiicola as a separate section with sect. Porri based on the formation of conidia in chains. Multi-locus phylogenetic analyses clearly separated the section from other species in sect. Porri[22].
Section Eureka was introduced by Woudenberg et al.[11] to accommodate four Alternaria species and the other two species previously described as Embellisia species which was mentioned as Embellisia group IV in Lawrence et al.[55]. Six species are currently known for this section, including Alternaria anigozanthi Priest, A. cumini E.G. Simmons, A. eureka E.G. Simmons (type species), A. geniostomatis E.G. Simmons & C.F. Hill, A. leptinellae (E.G. Simmons & C.F. Hill) Woudenb. & Crous, and A. triglochinicola Alcorn & S.M. Francis. These species were commonly isolated from plants and the rhizosphere[11,15]. The sexual morph is known for the type species of the section was regarded as Allewia species and characterized by spherical to ovoid, thin-walled, dark, papillate ascomata, with conspicuously setose, subcylindrical to subellipsoid asci, somewhat inequilateral, with subellipsoidal and slightly inequilateral juvenile ascospores. Ascospores are subclavate to ellipsoid, with transverse septa, discontinuous series of longitudinal septa when mature[15]. Multi-locus phylogenetic analyses based on the protein-coding genes demonstrated that the section has a close relationship with the morphologically similar sect. Embellisioides[11,15].
Section Gypsophilae was introduced by Lawrence et al.[14] to accommodate four Alternaria species, comprising A. gypsophilae Neerg. (type species), A. nobilis (Vize) E.G. Simmons, A. vaccariae (Săvul. & Sandu) E.G. Simmons & S.T. Koike and A. vaccariicola E.G. Simmons. Woudenberg et al.[11] recommended the other four species viz. A. axiaeriisporifera E.G. Simmons & C.F. Hill, A. ellipsoidea E.G. Simmons, A. juxtiseptata E.G. Simmons, and A. saponariae (Peck) Neerg. to this section based on multi-locus phylogeny. Based on morphological examination of Alternaria species producing conidia with many longitudinal septa, Gannibal & Lawrence[62] included A. longispora McAlpine in the sect. Gypsophilae. Consequently, Gannibal[74] introduced A. kamtschatica Gannibal from leaves of Dianthus barbatus in Russia. He et al.[3] introduced the other two new species in this section viz. A. barbata L. He & J.X. Deng and A. hispanica L. He & J.X. Deng from China. Currently, there are 12 species accommodated in this section that are restricted to the host family Caryophyllaceae[3,11,74]. The section has a close relationship with sects. Alternaria, Alternantherae, Euphorbiicola, and Porri[11,14,15].
Section Helianthiinficientes was introduced by Gannibal et al.[6] to accommodate Alternaria helianthiinficiens E.G. Simmons, Walcz & R.G. Roberts which was previously treated as a monotypic lineage in Woudenberg et al.[11] and Lawrence et al.[15]. Currently, only a single species is represented in this section[6]. The species was previously well-known as a causative pathogen on sunflower (Helianthus annuus L.) and cosmos (Cosmos bipinnatus Cav.) in Asia, Europe, and North America[6]. Gannibal et al.[6] reported the species on other hosts (i.e., Arctium sp. and Sonchus sp.) from Russia, suggesting that A. helianthiinficiens may also occur on other plant species in Asteraceae. Morphologically, A. helianthiinficiens resembles many species in sect. Porri in having large conidia[6]. However, multi-locus phylogenies analyzed by Woudenberg et al.[11] and Ghafri et al.[7] demonstrated that A. helianthiinficiens formed an independent lineage within Alternaria but could not be assigned to other known sections. Therefore, Gannibal et al.[6] established this new section.
Section Infectoriae was introduced by Woudenberg et al.[11] for Alternaria infectoria E.G. Simmons species-group, comprising approximately 45 accepted species in the sect. Infectoriae[15,24,70,75−80]. The human pathogenic genera Ybotromyces Rulamort (as Alternaria caespitosa (de Hoog & C. Rubio) Woudenb. & Crous) and Chmelia (as Alternaria slovaca (Svob.-Pol.) Woudenb. & Crous) were also embedded in sect. Infectoriae[11]. The section is typified by A. infectoria and taxa in this section are common saprobes and human pathogens as well as endophytes on apple leaves[15,77,80]. The sexual morph of sect. Infectoriae was linked to species in Lewia and is characterized by smooth-walled ascomata, subcylindrical or subellipsoidal asci and muriform ascospores with 5(−7) transverse septa and 1–2 longitudinal septa in central segments, with or without longitudinal or oblique septum in terminal cells[15]. The main refined morphological features of taxa in sect. Infectoriae are small conidia (usually less than 60 µm in vitro) and long secondary conidiophores[11,15,77]. Members of sect. Infectoriae are presumed to be homothallic mating-type genes that can produce protoascomata in axenic culture[15,81]. Phylogenetic analyses revealed the section as the sister group to sect. Pseudoalternaria and the most suitable genetic markers for 45 distinguishing species in the sect. Infectoriae are ATPase and cmdA genes[14,15,70].
Section Japonicae was introduced by Woudenberg et al.[11] with the type species of the section as Alternaria japonica Yoshii. The section was established to accommodate A. japonica together with A. nepalensis E.G. Simmons based on multi-locus phylogeny. Alternaria japonica was previously connected to the A. brassicicola species-group[14,54,59] but this connection was questioned by Hong et al.[49]. Bessadat et al.[82] included an additional species A. telliensis N. Bessadat, D. Ayad & P. Simoneau in this section; however, this species was invalidly introduced. Species in sect. Japonicae are frequently isolated from Brassicaceae hosts[11,15]. The phylogenetic status of the sect. Japonicae is uncertain within Alternaria[15].
Section Nimbya was introduced by Woudenberg et al.[11] and is typified by Alternaria scirpicola (Fuckel) Sivan. The section initially contained four species previously described as Nimbya species viz. A. caricis (E.G. Simmons) Woudenb. & Crous, A. scirpicola, A. scirpinfestans (E.G. Simmons & D.A. Johnson) Woudenb. & Crous, and A. scirpivora (E.G. Simmons & D.A. Johnson), Woudenb. & Crous. Gannibal[83] included the other two species, A. heteroschemos (Fautrey) Gannibal and A. juncicola (E.G. Simmons) Gannibal in this section. In addition, Ahmadpour[84] and Ahmadpour et al.[85] also introduced A. caricicola Ahmadp., A. cypericola Ahmadp., Poursafar & Ghosta, A. heyranica Ahmadp., Poursafar & Ghosta, and A. junci-acuti Ahmadp., Poursafar & Ghosta to this section. Hence, there are currently ten species accommodated in sect. Nimbya. It sounds that sect. Nimbya are restricted to Cyperaceae and Juncaceae host plant families[85]. The sexual morph of the section was referred to Macrospora Fuckel and is characterized by immersed to superficial, subglobose, ostiolate ascomata, with broadly cylindrical or clavate to obovoid asci and broadly ellipsoidal, brown to dark brown, multi-septate ascospores[15,24]. Section Nimbya is closely related to sects. Embellisia, Phragmosporae, Chalastospora and Infectoriae based on phylogenetic analyses of the combined GAPDH, RPB2 and TEF1-α sequence dataset[11].
Section Omanenses was introduced by Ghafri et al.[7], with the single species Alternaria omanensis as the type species of the section. Alternaria omanensis was isolated from dead wood in Oman as a saprobe and is known for both sexual and asexual morphs. The sexual morph of the section is characterized by superficial, subglobose to globose, or ovoid to cup-shaped (when dry), dark brown to black, carbonaceous ascomata, with a blunt ostiole, cylindrical to subcylindrical asci and pale brown to dark brown, muriform, subclavate to broadly obovoid or ellipsoid ascospores, with 3 transverse septa, 1–2 longitudinal septa in the central segments, without septa at the end cells, and constricted at the central septum (asexual morph see Table 1). Multi-locus phylogenetic analyses of a combined SSU, LSU, ITS, GAPDH, TEF1-α and RPB2 sequence dataset demonstrated that the section has a close relationship with sects. Embellisioides, Eureka and Ulocladium[7].
Section Panax was introduced by Lawrence et al.[14] and initially consisted of Alternaria calycipyricola R.G. Roberts, A. eryngii (Pers.) S. Hughes & E.G. Simmons and A. panax Whetzel (as A. panacis in Deng et al.[86] and Lawrence et al.[15]; type species). Woudenberg et al.[11] included A. avenicola E.G. Simmons and A. photistica E.G. Simmons to this section and the sexual morphs of these two species were known as Lewia avenicola Kosiak & Kwaśna[38] and L. photistica E.G. Simmons[40], respectively. Deng et al.[86] reported two pathogenic species in this section viz. A. araliae H.C. Greene and A. dendropanacis S.H. Yu & J.X. Deng that were associated with leaf spot and blight disease on Araliaceae in Korea, while Gannibal & Lawrence[62] included A. prasonis E.G. Simmons based on morphology. Recently, Hashemlou et al.[87] described A. hedjaroudei Y. Ghosta et al. on stems of Serratula coriacea Fisch. & C.A. Mey. from Iran in the sect. Panax. Thus, there are currently nine species accommodated in this section. PCR assays of mating-type genes indicated that members in sect. Panax are both homothallic and heterothallic species that are either capable of sporulating as sexual morphs in vitro or without an identified sexual morph. Phylogenetic analyses based on the GAPDH, RPB2 and TEF1-α sequences suggested that sect. Panax has a close relationship with A. thalictrigena and sect. Teretispora[11,15].
Section Phragmosporae was introduced by Woudenberg et al.[11] and is typified by Alternaria phragmospora Emden. The section contains six species viz. A. chlamydospora Mouch., A. didymospora (Munt.-Cvetk.) Woudenb. & Crous, A. limaciformis E.G. Simmons, A. molesta E.G. Simmons, A. mouchaccae E.G. Simmons, and A. phragmospora. These species are known from soil, seawater, seawater plants and animals, excluding A. didymospora which was found in equine nasal mucosa. There are no species associated with land plants in this section[15]. Phylogenetic results indicated the section is sister to sect. Embellisia, with A. didymospora and A. phragmospora were linked[11,15].
Section Porri was introduced by Lawrence et al.[14] and is typified by Alternaria porri (Ellis) Cif. Section Porri has been reported as the largest section of Alternaria with approximately 63 species revealed in the section based on multi-locus phylogeny[14,15,22]. A detailed study of this large-spored section was carried out by Woudenberg et al.[22]. The section displays a high level of genetic variation and contains many important plant pathogens, such as A. bataticola Ikata ex W. Yamam., A. porri, A. solani Sorauer and A. tomatophila E.G. Simmons, causing leaf and stem blight of sweet potato, purple blotch of onion and early blight of potato and tomato, respectively[22]. Gannibal[83] included A. rhapontici (Nelen) Gannibal in the section. Liu et al.[88] introduced A. physalidis H.F. Liu & J.X. Deng from Physalis alkekengi L. (Solanaceae) in China. Cai et al.[89] also introduced a pathogenic species A. yunnanensis Z.Y. Cai et al., which causes leaf spots on rubber trees in China. Poursafar et al.[90] introduced a pathogenic species A. guilanica Poursafar et al., on Solanum melongena L. with leaf spot and blight symptoms from Iran. Hence, 67 species are known in this section, making this section the second-largest section after sect. Alternaria. Multi-locus phylogeny demonstrated that the section is sister to sect. Euphorbiicola, and clustered with sects. Alternaria and Alternantherae[11,14,15,22].
Section Pseudoalternaria was introduced by Lawrence et al.[15] and is typified by Alternaria arrhenatheri D.P. Lawr., Rotondo & Gannibal. The section initially consisted of two species viz. A. arrhenatheri and A. rosae E.G. Simmons & C.F. Hill based on both phylogeny and morphology. Based on morphological examination, Gannibal & Lawrence[77] described a new taxon, A. parvicaespitosa Gannibal & D.P. Lawr. as a misidentified isolate previously identified as A. rosae by Zhu & Xiao[91]. Deng et al.[92] accommodated a new pathogenic species, A. brassicifolii S.H. Yu & J.X. Deng, causing necrotic leaf spots of Brassica rapa L. (Brassicaceae) in Korea in the section. However, Deng et al.[92] did not validly indicate the type specimens for their new species, and thus, the species is treated as invalid (nom. inval.) based on nomenclature article 40.1 (Shenzhen, China) that ‘Publication on or after 1 January 1958 of the name of a new taxon at the rank of genus or below is valid only when the type of the name is indicated’[93]. Subsequently, four other species were included in the section viz. A. altcampina Iturrieta-González, Dania García & Gené, A. ershadii A. Poursafar, Ghosta & M. Javan-Nikkhah, A. inflata Iturrieta-González, Dania García & Gené., and A. kordkuyana A. Poursafar et al.[70,94,95]. Currently, eight species are known in sect. Pseudoalternaria, all of which were confirmed using multi-locus phylogeny. Sect. Pseudoalternaria was shown to be closely related to sects. Infectoriae and Chalastospora[15,70].
Section Pseudoulocladium was introduced by Woudenberg et al.[11] to accommodate species previously described as Ulocladium species and is typified by Alternaria chartarum Preuss. Four species were initially included in the section viz. A. aspera Woudenb. & Crous, A. chartarum, A. concatenata Woudenb. & Crous, and A. septospora (Preuss) Woudenb. & Crous[11]. Based on morphology, Gannibal & Lawrence[96] included A. lanuginosa (Harz) Sacc. and A. sylvestris Gannibal & D.P. Lawr. Section Pseudoulocladium morphological resembles sects. Ulocladioides and Ulocladium but differs in simple or branched chains of conidia, whereas sect. Ulocladioides usually have densely geniculate conidiophores with clustered, short conidial chains, and secondary conidiophores are short with several conidiogenous loci. Section Ulocladium typically produces small, clustered, single conidia without chains[96]. Phylogenetic analyses of protein-coding genes revealed that the section has a sister relationship with sect. Dianthicola and clusters with sect. Ulocladioides[11].
Section Radicina was recognized by Pryor & Gilbertson[54] and formally established by Lawrence et al.[14]. The section was introduced to accommodate the radicina species-group and is typified by Alternaria radicina Meier, Drechsler & E.D. Eddy. Species in this section are pathogens occurring on Apiaceae[11,15]. Woudenberg et al.[11] and Lawrence et al.[15] listed five species in this section, including A. carotiincultae E.G. Simmons, A. petroselini (Neerg.) E.G. Simmons, A. radicina, A. selini E.G. Simmons and A. smyrnii (P. Crouan & H. Crouan) E.G. Simmons based on multi-locus phylogeny. Subsequently, Marin-Felix et al.[70] introduced A. chlamydosporifera Iturrieta-González, Dania García & Gené, isolated from rabbit dung in Spain, to the section. He et al.[4] introduced two new species in this section viz. A. divaricatae L. He & J.X. Deng and A. vulgaris L. He & J.X. Deng, both isolated from Umbelliferae (Apiaceae) in China. Hence, there are currently eight species accommodated in this section. Phylogenetic analyses demonstrated that the section has a close relationship with sect. Gypsophilae[15].
Section Soda was introduced by Grum-Grzhimaylo et al.[56] to contain three species isolated from soils at the different highly alkaline soda lakes in Russia, comprising Alternaria kulundae Bilanenko, Georgieva & Grum-Grzhim. (type species), A. petuchovskoi Bilanenko, Georgieva & Grum-Grzhim., and A. shukurtuzi Bilanenko, Georgieva & Grum-Grzhim. Species in this section showed a potential alkalitolerant to facultative alkaliphilic type of the adaptation. The sexual morph for the section is unknown. Multi-locus phylogeny of SSU, LSU, RPB2, ITS, and GAPDH showed that the section clustered with sects. Infectoriae, Chalastospora, and Embellisia[56].
Section Sonchi was described as the species-group by Hong et al.[49] and validly introduced by Lawrence et al.[14]. Only two species are accommodated in this section viz. Alternaria cinerariae Hori & Enjoji and the type species of the section A. sonchi Davis. Species in this section occur on a wide range of hosts in family Asteraceae[11,17]. The sexual morph of the section is unknown. Phylogenetic analyses based on the GAPDH, RPB2 and TEF1-α sequences showed that sect. Sonchi forms a sister clade with two monotypic lineages, A. brassicae (Berk.) Sacc. and A. helianthiinficiens E.G. Simmons, Walcz & R.G. Roberts[11]. Currently, A. helianthiinficiens was raised to the section rank of Alternaria by Gannibal et al.[6]. Ferreira & Barreto[97] designated the neotype of Acroconidiella tropaeoli (T.E.T. Bond) J.C. Lindq. & Alippi (≡ Heterosporium tropaeoli T.E.T. Bond) and proposed the new name for the species as Alternaria obtusa B.W. Ferreira & R.W. Barreto. The species is sister to sect. Sonchi[97].
Section Teretispora was introduced by Woudenberg et al.[11] to accommodate a single species, Alternaria leucanthemi Nelen, as the type of the section. The species was isolated from Leucanthemum maximum (Ramond) DC. (Asteraceae) and is characterized by simple primary conidiophores bearing 1–3 conidiogenous loci and generally solitary, cylindrical conidia, with 7–14(–17) transverse septa, and 3–7 longitudinal septa[15]. The sexual morph has not yet been described for the section. Phylogenetic analyses showed that sect. Teretispora is sister to A. thalictrigena, and clustered with sect. Panax. Thus, Woudenberg et al.[11] proposed to raise this species as a section, rather than a monotypic lineage.
Section Ulocladioides was introduced by Woudenberg et al.[11] and is typified by Alternaria cucurbitae Letendre & Roum. The section was introduced to accommodate ten species previously described as Ulocladium species based on phylogeny, place it distant from sect. Ulocladium. Section Ulocladioides is similar to the sect. Ulocladium, and is characterized by short, geniculate conidiophores, with sympodial proliferations and obovoid, non-beaked conidia, with a narrow base, single or in chains[11]. Gannibal & Lawrence[96] included the other ten species and thus, 20 species are currently known for this section. Phylogenetic analyses based on the GAPDH, RPB2 and TEF1-α sequences showed that sect. Ulocladioides has a close relationship with sects. Pseudoulocladium and Dianthicola[11].
Section Ulocladium was introduced by Woudenberg et al.[11] and is typified by Alternaria botrytis (Preuss) Woudenb. & Crous. The section is introduced to accommodate the epitype of the former Ulocladium as Alternaria botrytis (CBS 197.67) and additional three species viz. A. alternariae
(Cooke) Woudenb. & Crous, A. capsici-annui Săvul. & Sandu, and A. oudemansii (E.G. Simmons) Woudenb. Gannibal & Lawrence[96] included A. manihoticola (J.M. Yen) Gannibal & D.P. Law in the section based on morphological study and thus, five species are known for the section. Phylogenetic analyses based on the GAPDH, RPB2 and TEF1-α sequences showed that sect. Ulocladium is sister to the monotypic lineage A. argyranthemi[11]. Section Undifilum was introduced by Woudenberg et al.[11] and is typified by Alternaria bornmuelleri (Magnus) Woudenb. & Crous. The section consists of five species viz. A. bornmuelleri, A. cinerea (Baucom & Creamer) Woudenb., A. fulva (Baucom & Creamer) Woudenb. & Crous, A. gansuensis J. Li Liu & Y.Z. Li, and A. oxytropis (Q. Wang, Nagao & Kakish.) Woudenb. & Crous[11,98]. Section Undifilum resembles sect. Embellisia, but can be distinguished by conidial germination with the germ tube being wavy and unbranched[11,60]. Species in this section were isolated from Fabaceae as endophytes and produced a swaisonine toxic compound, causing a neurological disease of grazing animals[99]. Phylogenetic analyses of the GAPDH, RPB2 and TEF1-α sequences showed that the section forms an independent lineage closely related to a monotypic lineage Embellisia dennisii (M.B. Ellis) E.G. Simmons, (CBS 110533, CBS 476.90), which was resurrected as Alternaria dennisii M.B. Ellis in Woudenberg et al.[11].
Evolutionary and fossil studies of Alternaria
-
The study of fossil fungi has become an essential tool to understanding fungal evolution and diversification, as well as the correlation of fungi with other organisms coupled with historical functions in the ecosystem[119,120]. The detailed study of fungal fossils was limited in the early stages due to the technical factors used to study fossil fungi and visual matching for identifying similar extant species as well as poor preservation and unclear morphological characteristics[119−121]. Furthermore, the study of fossil fungi received little attention because of lack of interest, expertise and collaboration[119,120]. Samarakoon et al.[120] mentioned that although the study of fossil fungi is not an essential tool for fungal taxonomy, it is important for understanding the paleoecological conditions and calibrating divergent times of fungal evolution based on molecular clock studies. Hence, Samarakoon et al.[120] assimilated 16 selected fossil fungi in Ascomycota and provided detailed information based on descriptions, illustrations, minimum age estimations, and phylogenetic affinity, mostly regarding the epiphytic Dothideomycetes and Sordariomycetes.
The fossil record of Alternaria has also not been determined in the Kalgutkar and Jansonius database of fossil fungi[122]. However, there is a fossil record referred to Alternaria described as Polycellaesporonites alternariatus (Kalgutkar & Sigler) Kalgutkar & Janson (≡ Piriurella alternariata Kalgutkar & Sigler). Polycellaesporonites alternariatus was first described as Piriurella alternariata by Kalgutkar & Sigler[123] for the fungal fossil-produced dictyosporae spore group. The species was referred to Alternaria in forming muriform, ovoid to obclavate, rostrate, pale brown to brown conidia, arising singly or in clusters, with transverse septa more prominent and thicker than the longitudinal or oblique septa. The broader basal region distally tapered to a short or cylindrical beak with or without a dark thickened tip[123]. The species was found from Iceberg Bay formation at Kanguk Peninsula, Axel Heiberg Island and Northwest Territories, Canada with an estimated age during the late Palaeocene or early Eocene (40.4–58.7 MYA)[123]. However, the link between P. alternariatus and Alternaria has not yet been confirmed due to Alternaria being variable in shape, size and septation of conidium and was shown to be a species complex[11−14,22,55,123].
Evolutionary estimates based on molecular clocks have been increasingly common in fungal taxonomy in recent years[1,124−130], with some works including the Pleosporales[130−132]. Hyde et al.[128] proposed Kingdom Fungi as evolving during the Stenian to Calymmian era (1000–1600 MYA), the phyla evolved between Devonian to Cambrian (358–541 MYA), the classes evolved during Jurassic to Carboniferous (145–358 MYA) and the orders evolved during Cretaceous to Carboniferous (66–358 MYA). They also determined the higher ranks of fungi based on the divergence time estimations of Sordariomycetes, of which the familial rank would correspond to 50–150 MYA. Liu et al.[129] recommended that the orders of Dothideomycetes should have evolved between 100 and 220 MYA (crown age) and 130 and 310 MYA (stem age), and the families are ranked between 20–100 MYA (crown age). However, the divergence time estimations of Alternaria based on DNA sequence evidence remain unexplored.
In this research, we isolated Alternaria species from 65 specimens, collected from different plant hosts in Yunnan, China, Italy, Russia and Thailand from 2014 to 2019 and introduce 18 novel Alternaria species, all of which are represented by the hyphomycetous asexual morphs as saprobes on dead plant tissues. We also provide updated phylogenetic relationships for the sections in Alternaria based on phylogenetic analyses of a concatenated dataset from seven gene regions (ITS, LSU, SSU, TEF1-α, RPB2, GAPDH and Alt-a1 loci) and have estimated evolutionary divergence time for Alternaria.
-
Alternaria species, isolated from various hosts, were mainly collected in Italy, and partly from China (Yunnan), Russia and Thailand, during 2014–2019. Materials were brought to the laboratory in Zip-loc plastic bags and examined under a Motic SMZ 168 stereomicroscope. Morphological studies were conducted following the guidelines by Senanayake et al.[133]. Micromorphological characters of Alternaria species were examined under a Nikon ECLIPSE 80i compound microscope and images were captured using a Nikon ECLIPSE 80i compound microscope with a Canon EOS 550D digital camera. Measurements were made with the Tarosoft (R) Image Frame Work and images used for figures were processed with Adobe Photoshop CS3 Extended version 10.0 software. New species were justified based on Jeewon & Hyde[134] and registered in Faces of Fungi[135] and Index Fungorum[5].
Isolates were derived via single spore isolation following the method of Chomnunti et al.[136] and Senanayake et al.[133]. Germinating spores were transferred to potato dextrose agar (PDA; 39 g/L distilled water, DifcoTM potato dextrose, Montreal, Canada) or malt extract agar (MEA; 33.6 g/L sterile distilled water, DifcoTM malt extract, Montreal, Canada) media and incubated at 18–25°C. The cultural characteristics such as mycelium color, shape, texture and growth rate were determined after 1–8 weeks. The sporulation in vitro was induced on potato carrot agar (PCA; 20 g potato + 25 g carrot + 15 g agar/1 L) and observed after 8 weeks. The living cultures were preserved in PDA, the sterilized 10% glycerol, and double-distilled water (ddH2O) and deposited at the Mae Fah Luang University Culture Collection (MFLUCC), and duplicated at the Culture Collection of Kunming Institute of Botany (KUMCC/KUNCC) and China General Microbiological Culture Collection Center (CGMCC). The type and other collected specimens were deposited in the herbarium of Mae Fah Luang University (MFLU), Chiang Rai, Thailand and Herbarium of Cryptogams Kunming Institute of Botany, Academia Sinica (KUN-HKAS), China.
DNA extraction, PCR amplification and sequencing
-
Fungal isolates cultured on PDA or MEA at 25–28 °C for 25–30 d were used for genomic DNA extraction following the guidelines by Dissanayake et al.[137]. Fungal mycelium was scraped off and stored in a sterilized 1.5-ml microcentrifuge for further DNA extraction. Fungal genomic DNA was extracted using the Biospin Fungus Genomic DNA Extraction Kit (BSC14S1, BioFlux®, China), following the manufacturer's instructions.
DNA amplifications were conducted by polymerase chain reaction (PCR) with seven genes as listed in Table 2. Polymerase chain reaction (PCR) was performed in a ABI Veriti gradient PCR machine (Applied Biosystem, USA) with the total 25 μl reaction volume, containing 1 µl of DNA template, 1 µl of each forward and reverse primers, 12.5 µl of 2× Power Taq PCR Master Mix (mixture of EasyTaqTM DNA Polymerase, dNTPs, and optimized buffer, Beijing BioTeke Corporation, P.R. China) and 9.5 µl of sterilized double-distilled water (ddH2O). PCR thermal cycling conditions of each locus were set up following Woudenberg et al.[11] but adjusted as: for ITS, LSU, SSU, and TEF1-α was set up at initially, 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 50 s, elongation at 72 °C for 1 min; for RPB2 was set up at initially 95 °C for 2.30 min, followed by 35 cycles of denaturation at 95 °C for 30 s, annealing at 52 °C for 1 min, elongation at 72 °C for 1 min; for GAPDH was set up at initially 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 57 °C for 1 min, elongation at 72 °C for 90 sec; for Alt-a1 was set up at initially 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 57 °C for 30 s, elongation at 72 °C for 1 min; a final extension at 72 °C for 10 min, and finally hold at 4 °C. The PCR fragments were then checked on 1% agarose electrophoresis gels stained with ethidium bromide and visualized under the UV light using the Molecular Imager Gel Doc XR + Imaging system (BIO-RAD, USA). The amplified PCR fragments were sent to a commercial sequencing provider (TsingKe Biological Technology (Beijing) Co., Ltd, P.R. China) for purification and sequencing in both forward and reverse directions. Consensus sequences were incorporated with both forward and reverse sequences, computed by Bioedit v.7.1.3.0[138]. All acquired nucleotide sequences were deposited in GenBank (Supplemental Tables S2, S3).
Table 2. Gene loci and primers used in this study.
Gene loci Primers Sequence 5’–3’ References Internal transcribed spacer region (ITS, including the 5.8S gene) ITS5 GGA AGT AAA AGT CGT AAC AAG G [139] ITS4 TCC TCC GCT TAT TGA TAT GC 28S large subunit rDNA (LSU) LR0R GTA CCC GCT GAA CTT AAG C [140] LR5 ATC CTG AGG GAA ACT TC 18S small subunit rDNA (SSU) NS1 GTA GTC ATA TGC TTG TCT C [139] NS4 CTT CCG TCA ATT CCT TTA AG Alternaria major allergen (Alt-a1) Alt-for ATG CAG TTC ACC ACC ATC GC [49] Alt-rev ACG AGG GTG AYG TAG GCG TC Glyceraldehyde 3-phosphate Dehydrogenase (GAPDH) GDP-1 CAA CGG CTT CGG TCG CAT TG [141] GDP-2 GCC AAG CAG TTG GTT GTG C Plasma membrane ATPase (ATPase) ATPDF1 ATC GTC TCC ATG ACC GAG TTC G [14] ATPDR1 TCC GAT GGA GTT CAT GAT AGC C The second largest subunit of RNA polymerase II (RPB2) fRPB2-5f GAY GAY MGW GAT CAY TTY GG [142] fRPB2-7cR CCC ATR GCT TGY TTR CCC AT Translation elongation factor 1-α (TEF1-α) EF1-983F GCY CCY GGH CAY CGT GAY TTY AT [143] EF1-2218R ATG ACA CCR ACR GCR ACR GTY TG EF1-728F CATCGAGAAGTTCGAGAAGG [144] EF1-986R TACTTGAAGGAACCCTTACC Fossil calibration, divergence time and evolutionary rate estimations
-
Fossil calibrations used in the analyses followed the methodology described in Phukhamsakda et al.[131]. Two fossil and one secondary calibration were applied to estimate all other nodes in the tree. Fossil 1, Metacapnodium succinum (Metacapnodiaceae) was used to calibrate the minimum age of Capnodiales (normal distribution, mean = 100, SD = 150, providing 95% credibility interval of 346 MYA)[126,131,145−147] and fossil 2, Margaretbarromyces dictyosporus was used to calibrate the crown age of Aigialus (Aigialaceae) (gamma distribution, offset = 35, shape = 1.0, scale = 25, providing 95% credibility interval of 110 MYA)[131,148]. The split between Arthoniomycetes and Dothideomycetes was calibrated using the results from Phukhamsakda et al.[131] as the secondary calibration (normal distribution, mean = 300, SD = 50, providing 95% credibility interval of 382 MYA).
Evolutionary estimation based on molecular clock analysis was performed by BEAST 1.8.4[149]. Aligned sequence data were partitioned separately for each ITS, LSU, SSU, TEF1-α and RPB2 dataset, and were loaded to prepare an XML file constructed with BEAUti v1.8.4. Clock and substitution models were set to be independently estimated for each gene partition, while the tree prior parameters were set to be linked across partitions. A lognormal relaxed clock (uncorrelated) model was applied with a lognormal distribution of rates for each gene estimated. The best fit of substitution models was selected based on jModeltest2 v.0.1.1[150] for each gene partition, resulting as ITS = GTR+I+G, LSU = GTR+I+G, SSU = TIM2+I+G, TEF1-α = SYM+I+G, RPB2 = TIM2+I+G; Yule processed tree prior with a randomly generated starting tree. The analysis was performed for 100 million generations in BEAST v1.8.4, sampling parameters every 10,000 generations. The effective sample sizes (ESS) were checked by Tracer v1.6[151] and accepted when ESS values were higher than 200. The first 10% trees were discarded as a burn-in phase. The remaining trees were combined in LogCombiner 1.8.0. A maximum clade creditability (MCC) tree was generated by summarized data and estimated in TreeAnnotator v1.8.0. The tree was visualized with FigTree v1.4.[152].
Phylogenetic analyses
-
The quality of the generated ITS, LSU, SSU, TEF1-α, RPB2, GAPDH, Alt-a1 and ATPase sequences was checked using Bioedit v. 7.1.3.0[138] and subjected to the nucleotide BLAST search engine via the NCBI (
www.ncbi.nlm.nih.gov ) for checking potential contaminants or erroneous sequences as well as delineating the closely related taxa. All reference sequences were downloaded from GenBank (Supplemental Tables S1, S2, S3) based on recent publications[4,12,47,63,70,78,79,88,89].The multiple sequence alignments were automatically generated by MAFFT v. 7.452[153] (
https://mafft.cbrc.jp/alignment/software/ ), and manual improvements were made where necessary using BioEdit v. 7.2[138]. Individual gene alignments were separately analyzed by maximum likelihood (ML) in order to check the congruence of tree topology and thus the combined multi-locus phylogenetic trees were inferred based on Bayesian inference (BI) and maximum likelihood (ML) analyses. Five different datasets were generated to estimate phylogenic relationships of Alternaria sections (analysis 1), intraspecific variation of A. alternata (analysis 2), sect. Infectoriae (analysis 3), sect. Porri (analysis 4), and sect. Radicina (analysis 5).Maximum likelihood (ML) analyses were performed by RAxML[154] implemented in raxmlGUI 1.3[155] with 1000 bootstrap replicates and GAMMAI model of nucleotide substitution. MrModeltest v. 2.3[156] was used to determine the best-fit model of nucleotide substitution for each locus and incorporated into the analyses (Table 3). Bayesian inference (BI) analyses were performed by MrBayes v.3.1.2[157]. Markov Chain Monte Carlo (MCMC) of six simultaneous Markov chains were run with 1–5 million generations to determine posterior probabilities (PP)[158,159] and started from a random tree topology. Trees were frequently sampled at 100th generation and the temperature value of heated chain was set to 0.15. The extra runs were required when the average standard deviation of split frequencies was not lower than 0.01 after one million generations. The first 25% trees represented the burn-in phase of the analyses, were discarded and the remaining trees were used for calculating posterior probabilities (PP) in the majority rule consensus tree. The phylogram were visualized in FigTree v. 1.4.0[152] and edited in Microsoft Office PowerPoint 2016 (Microsoft Inc., USA). The final alignments and trees were submitted in TreeBASE (
www.treebase.org ) following the submission ID: 258523–258527.Table 3. The best nucleotide substitution model for each locus based on the Akaike Information Criterion (AIC) generated by MrModeltest v. 2.3.[156].
Phylogenetic analyses Nucleotide substitution models ITS LSU SSU GAPDH RPB2 TEF1-α Alt-a1 ATPase A1: Alternaria sections GTR+I+G GTR+G TrN+I+G SYM+I+G GTR+I+G TIM1+I+G GTR+I+G n/a A2: A. alternata GTR+I+G GTR+I+G GTR+I+G GTR+I+G TIM2 +G GTR+I+G GTR+I+G n/a A3: sect. Infectoriae GTR+I+G n/a n/a GTR+I+G n/a n/a n/a SYM+G A4: sect. Porri SYM+I+G n/a n/a TIM2+I+G GTR+I+G GTR+G GTR+I+G n/a A5: sect. Radicina GTR+I+G n/a n/a GTR+I+G TIM2+G GTR+I+G n/a n/a -
Representative strains of taxa in Pleosporales were analyzed based on a combined ITS, LSU, SSU, TEF1-α and RPB2 DNA sequence dataset comprised 227 strains of ingroup taxa. Four species in Arthoniomycetes (Arthonia dispersa UPSC2583, Dendrographa leucophaea f-minor, Roccella fuciformis Tehler 8171 and Schismatomma decolorans Ertz 5003) were selected as the outgroup taxa. The best scoring RAxML tree is presented in Fig. 1 with final ML optimization likelihood value of −61934.763756 (ln). RAxML analysis yielded 1,831 distinct alignment patterns and 19.69% of undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.252569, C = 0.228251, G = 0.283442, T = 0.235738, with substitution rates AC = 1.554349, AG = 4.323993, AT = 1.241832, CG = 1.108361, CT = 8.993327, GT = 1.000000. The gamma distribution shape parameter alpha = 0.307857 and the Tree-Length = 19.058776. Bayesian posterior probabilities (BYPP) from MCMC were evaluated with final average standard deviation of split frequencies = 0.009034. The final alignment and tree were submitted in TreeBASE as submission ID: 258527. The phylogenetic results of Pleosporales (Fig. 1) showed an overall similar tree topology with maximum clade credibility (MCC) tree (Fig. 2). Alternaria sections formed well-resolved and stable clades (up to 80% ML, 0.95 PP; Fig. 1) within Pleosporaceae; while the phylogenetic status of sects. Embellisioides and Eureka (Fig. 1: phylogenetic tree of Pleosporales) are not well-resolved, concurring with phylogenetic results of Alternaria sections (Analysis 1; Fig. 3: phylogenetic tree of Alternaria sections).
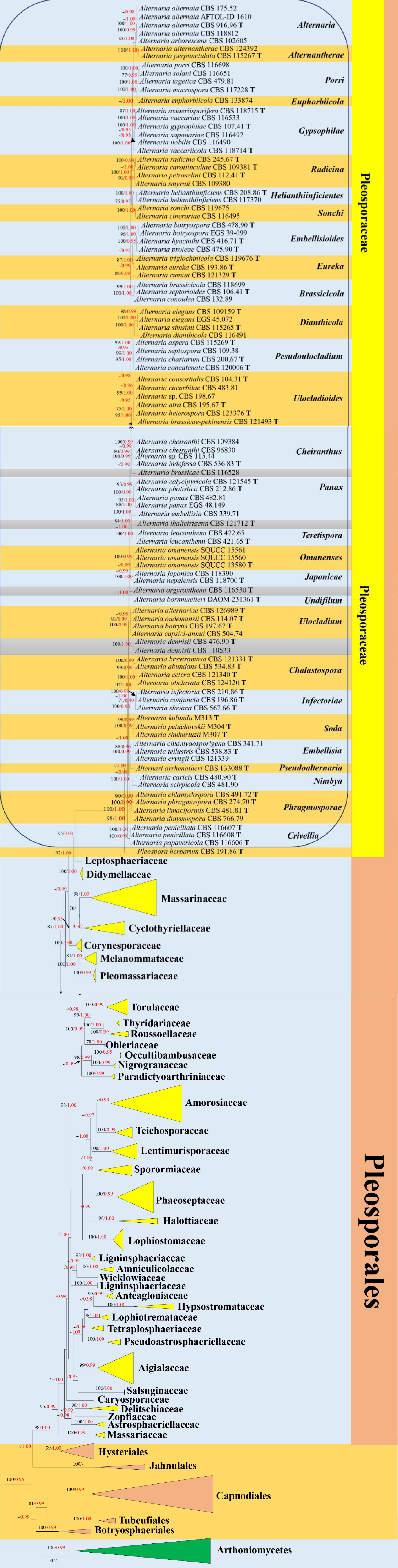
Figure 1.
Phylogenetic construction of Pleosporales using RAxML-based maximum likelihood analysis of a combined ITS, LSU, SSU, TEF1-α and RPB2 DNA sequence dataset. Bootstrap support values for maximum likelihood (ML, black) equal to or greater than 70% and Bayesian posterior probabilities (PP, red) equal to or greater than 0.95 PP are shown above the nodes. The tree is rooted to Arthoniomycetes (Arthonia dispersa UPSC2583, Dendrographa leucophaea f-minor, Roccella fuciformis Tehler 8171 and Schismatomma decolorans Ertz 5003). The type strains are indicated by boldface 'T'.
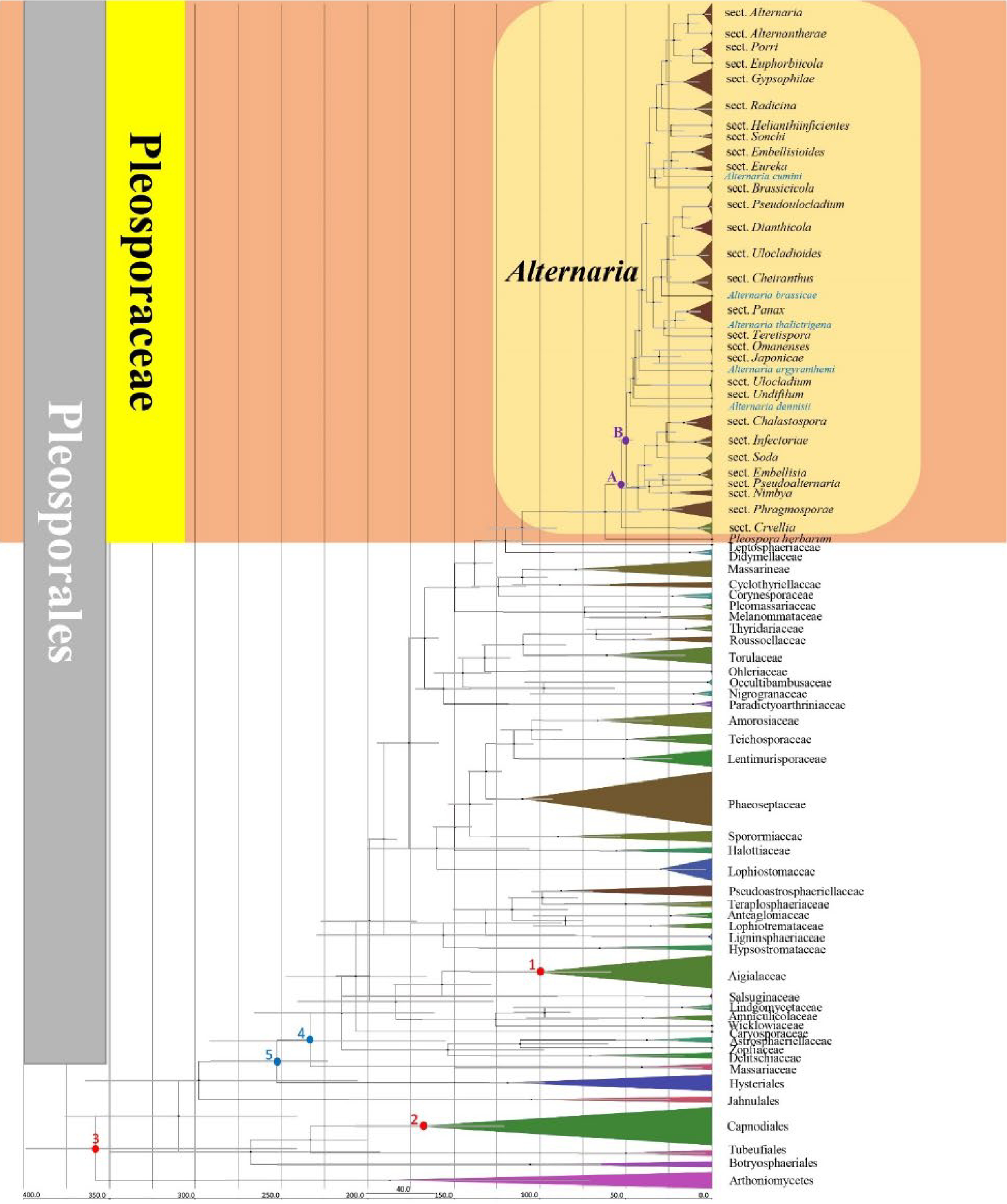
Figure 2.
Maximum clade credibility (MCC) tree with divergence times estimates obtained from BEAST. Numbers in red indicate the fossil (1, 2) and secondary (3) points. Numbers in blue indicate divergence time estimate of Pleosporales (stem age: 5, crown age: 4). Letters in purple indicate divergence time estimate of Alternaria (stem age: A, crown age: B). Single lineages of Alternaria species are highlighted in blue.

Figure 3.
Phylogenetic construction of genus Alternaria using RAxML-based maximum likelihood analysis of a combined ITS, LSU, SSU, TEF1-α, RPB2, GAPDH and Alt-a1 DNA sequence dataset. Bootstrap support values for maximum likelihood (ML, black) equal to or greater than 70% and Bayesian posterior probabilities (PP, red) equal to or greater than 0.95 PP are shown above the nodes. The tree is rooted to Stemphylium vesicarium (CBS 191.86) and Pleospora tarda (CBS 714.68). Newly generated strains are in blue. The type strains obtained from ex-type cultures are indicated by 'T' and the type strains obtained from specimens are indicated by 'H'.
According to divergence time estimates (Fig. 2), the stem and crown ages of Dothideomycetes are 358 (266–492) Mya and 310 (230–392) Mya in the Permian Period, respectively (Fig. 2). Pleosporales diverged with other orders roughly 253 (184–326) Mya in the Triassic Period. The crown age of Pleosporales is around 233 (168–301) Mya in the Late Triassic. The crown and stem ages of Dothideomycetes and Pleosporales in the MCC tree (Fig. 2) are well-supported, falling in the recommended divergence times for class and order status by Liu et al.[129] and Hongsanan et al.[1]. In Pleosporales, Pleosporinae diverged approximately 120 (84–159) Mya in Cretaceous. The stem age of Alternaria is at 62 (42–85) Mya and the crown age of Alternaria is at 53 (36–72) Mya in the age of late Paleocene to early Eocene. The species occurred in the sections that diverged earlier than other sections in Alternaria with beakless, rare multi and longitudinal septate conidia, less forming secondary conidiophores such as species in sects. Crivellia, Phragmospora, Ulocladium, and Undifilum, while later diverged sections mostly comprise species with beaks or multi-septate conidia forming secondary conidiophores with conidiogenous loci[11,12,14,15,22]. Divergence times of other sections in the analysis are shown in Table 4.
Table 4. Divergence times of Alternaria sections indicated in MCC tree. The age value with '*' indicates recent results lacking key coding gene strains.
Order Family Genus Sections Divergence times (crown age) Divergence times (stem age) Pleosporales 233 (168–301) Mya 252 (184–326) Mya Pleosporaceae 110 (79–148) Mya 120 (84–159) Mya Alternaria 53 (36–71) Mya 62 (42–85) Mya Alternaria sect. Alternantherae 0.4 (0–1.5) Mya 14 (6.7–21) Mya Alternaria sect. Alternaria 5 (1.7–10) Mya 14 (6.7–21) Mya Alternaria sect. Brassicicola 2.3 (0.5–5.5) Mya 33 (22–45) Mya Alternaria sect. Chalastospora 16 (8.8–26) Mya 26 (16–38) Mya Alternaria sect. Cheiranthus 11 (4.23–20) Mya 26 (16–38) Mya Alternaria sect. Crivellia 7.6 (1.5–19) Mya 53 (36–71) Mya Alternaria sect. Dianthicola 11 (5.4–18) Mya 17 (10–27) Mya Alternaria sect. Embellisia 7.4 (2.5–15) Mya 28 (14–43) Mya Alternaria sect. Embellisioides 11 (5–19) Mya 24 (14–36) Mya Alternaria sect. Eureka 14 (5.6–24) Mya 28 (18–44) Mya Alternaria sect. Euphorbiicola - 11 (5.6–17) Mya Alternaria sect. Gypsophilae 16 (7.6–26) Mya 27 (18–37) Mya Alternaria sect. Helianthiinficientes 0.11 (0–0.3) Mya 24 (13–36) Mya Alternaria sect. Infectoriae 9.5 (4–17) Mya 26 (16–38) Mya Alternaria sect. Japonicae – 31 (14–47) Mya Alternaria sect. Nimbya 24 (11–39) Mya 36 (28–51) Mya Alternaria sect. Omanenses – 30 (14–47) Mya Alternaria sect. Panax 14 (6.8–23) Mya 22 (12–33) Mya Alternaria sect. Phragmosporae 28 (13–44) Mya 42 (28–58) Mya Alternaria sect. Porri 6.7 (3–11) Mya 11 (5.6–17) Mya Alternaria sect. Pseudoalternata – 28 (14–43) Mya* Alternaria sect. Pseudoulocladium 2.1 (0.4–5.8) Mya 17 (9.5–27) Mya Alternaria sect. Radicina 9.3 (3.6–18) Mya 29 (19–40) Mya Alternaria sect. Soda 3 (0.5–8.4) Mya 32 (26–54) Mya Alternaria sect. Sonchi 6.8 (2–14) Mya 24 (13–36) Mya Alternaria sect. Teretispora 0.2 (0–1.03) Mya 27 (17–40) Mya Alternaria sect. Ulocladioides 8.1 (3–17) Mya 22 (13–32) Mya Alternaria sect. Ulocladium 0.9 (0.1–2.5) Mya 44 (32–60) Mya Alternaria sect. Undifilum – 45 (32–62) Mya Phylogenetic analyses of Alternaria sections
-
In this study, five phylogenetic trees were inferred to define the phylogenetic placements of the novel Alternaria species and relationships of taxa in Alternaria sections. The multi-locus phylogenetic tree (Fig. 3) demonstrated that 18 novel species were delineated in sects. Alternaria, Infectoriae, Porri and Radicina. Fourteen new species and two new records on host and geography are introduced in sect. Alternaria and two novel species are introduced to sect. Infectoriae, and the other two new species are introduced in sects. Porri and Radicina, respectively.
Analyses 1 revealed phylogenetic relationships of the representative Alternaria taxa in 29 sections and the novel species introduced in this study. The combined ITS, LSU, SSU, TEF1-α, RPB2, GAPDH and Alt-a1 sequence dataset comprises 189 taxa with Stemphylium vesicarium (CBS 191.86) and Pleospora tarda (CBS 714.68) as the outgroup taxa. Bayesian inference (BI) and maximum likelihood (ML) analyses of the combined dataset resulted in phylogenetic reconstructions with largely similar topologies. The best scoring RAxML tree is shown in Fig. 3, with the final ML optimization likelihood value of –42167.053232 (ln). The dataset consists of 4,385 total characters, including gaps (ITS: 1–679 bp, LSU: 680–1,532 bp, SSU: 1,533–2,459 bp, TEF1-α: 2,460–2,740 bp, RPB2: 2,741–3,314 bp, GAPDH: 3,315–3,908 bp, Alt-a1: 3,909–4,385 bp). RAxML analysis yielded 1,623 distinct alignment patterns and 15.42% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.248242, C = 0.254002, G = 0.257113, T = 0.240642, with substitution rates AC = 1.284821, AG = 3.337861, AT = 1.081773, CG = 0.849184, CT = 5.615040, GT = 1.000000. The gamma distribution shape parameter alpha = 0.184966 and the Tree-Length = 2.767524. Bayesian posterior probabilities (PP) from MCMC were evaluated with the final average standard deviation of split frequencies = 0.008657.
Present multi-locus analyses (Fig. 3) demonstrated that most Alternaria sections formed well-resolved clades with high support values (up to 70% ML, 0.95 PP), excluding sects. Cheiranthus, Omanenses, and Undifilum. Section Cheiranthus clustered with sects. Dianthicola and Pseudoulocladium with significant support in BI analysis (0.96 PP) but low support in ML analysis. Three representative species in sect. Cheiranthus (A. cheiranthi, A. indefessa and Alternaria sp.) grouped together with significant support in ML analysis (74% ML) but low support in BI analysis; while the clades of sect. Eureka and sect. Embellisioides were not well separated, and grouped together with high support values (85% ML, 0.99 PP). Alternaria cumini CBS 121329 formed an independent lineage basal to sect. Eureka and sect. Embellisioides with high support (93% ML, 0.99 PP). Three representative strains of A. omanensis formed a robust clade (100% ML, 1.00 PP), basal to sect. Eureka and sect. Embellisioides with significant support in BI analysis (0.95 PP), but low support in ML analysis. The putative strain of A. bornmuelleri (DAOM 231361), represented sect. Undifilum and formed an independent lineage (0.95 PP) with A. dennisii (CBS 476.90, CBS 110533).
Fourteen new species are introduced in sect. Alternaria, including A. arctoseptata, A. baoshanensis, A. breviconidiophora, A. ellipsoidialis, A. eupatoriicola, A. falcata, A. lathyri, A. macilenta, A. macroconidia, A. minimispora, A. oblongoellipsoidea, A. orobanches, A. phragmiticola and A. salicicola. These novel species formed independent well-supported subclades (up to 80% ML and 0.95 PP; Fig. 3) within sect. Alternaria. The new collection, A. doliconidium (MFLUCC 14-0020), clustered with the type strains (HKAS 100840, MFLUCC 17-0263) of A. doliconidium with high support values (99% ML, 100 PP; Fig. 3). However, the species did not form a well-resolved clade and clustered with other strains of A. alternata and A. italica.
Analyses 2 represented the intraspecific variation of Alternaria alternata corresponding with their hosts. Phylogenetic construction of A. alternata based on a combined ITS, LSU, SSU, TEF1-α, RPB2, GAPDH and Alt-a1 DNA sequence dataset comprised 110 strains with A. eichhorniae Nag Raj & Ponnappa (CBS 489.92, CBS 119778) as the outgroup. The best scoring RAxML tree is shown in Fig. 4, with the final ML optimization likelihood value of –6848.962092 (ln). The dataset consists of 4,377 total characters including gaps (ITS: 1–514 bp, LSU: 515–1,368 bp, SSU: 1,369–2,295 bp, TEF1-α: 2,296–2,540 bp, RPB2: 2,541–3,311 bp, GAPDH: 3,312–3,897 bp, Alt-a1: 3,898–4,377 bp). RAxML analysis yielded 74 distinct alignment patterns and 1.54% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.246749, C = 0.253377, G = 0.260395, T = 0.239479, with substitution rates AC = 5.780289, AG = 13.046393, AT = 1.475727, CG = 0.816872, CT = 36.600039, GT = 1.000000. The gamma distribution shape parameter alpha = 0.020000 and the Tree-Length = 0.029480. Bayesian posterior probabilities (BYPP) from MCMC were evaluated with final average standard deviation of split frequencies = 0.008559. The final alignment and tree were submitted in TreeBASE as submission ID: 258523. Alternaria alternata strains represented in this study were obtained from diverse plant hosts and humans. Forty-five new collections of A. alternata were included in the present analyses and are reported for different hosts and geography from China, Italy and Thailand. Multi-locus phylogenetic analyses (Fig. 4) showed a high intraspecific genetic variation of A. alternata. This phylogenetic result concurs with Woudenberg et al.[12]. However, the strains of A. alternata can be distinguished into five main subclades with significant support (up to 80% ML, 0.95 PP) in this study. Hence, existing strains of A. alternata may represent five species rather than a single species, and further work is needed to clarify the phylogeny.
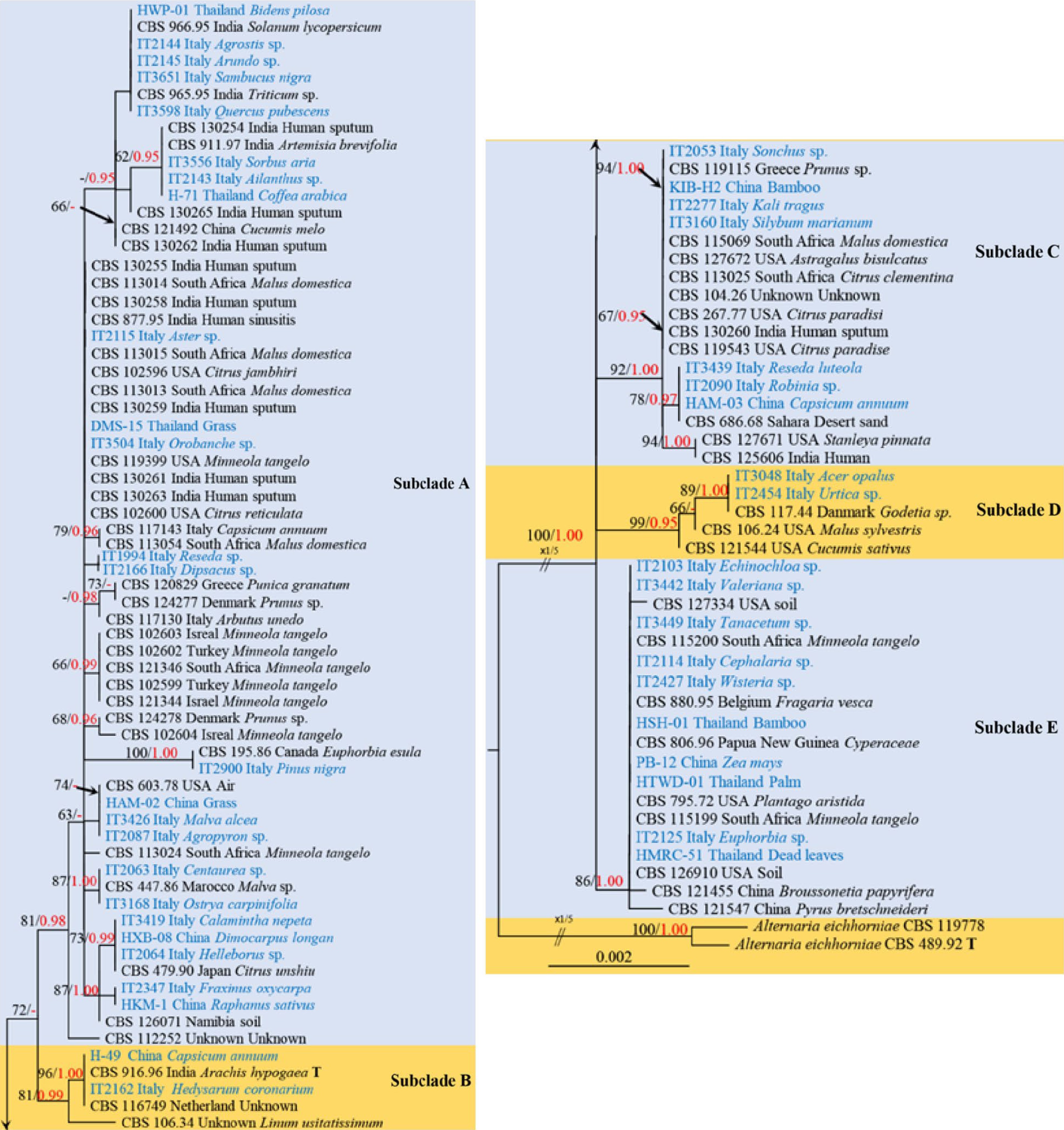
Figure 4.
Phylogenetic construction of Alternaria alternata using RAxML-based analysis of a combined ITS, LSU, SSU, TEF1-α, RPB2, GAPDH and Alt-a1 DNA sequence dataset. Bootstrap support values for maximum likelihood (ML, black) equal to or greater than 60% and Bayesian posterior probabilities (PP, red) equal to or greater than 0.95 are shown above the nodes. The tree is rooted to Alternaria eichhorniae (CBS 489.92) and Alternaria eichhorniae (CBS 119778). Newly generated strains are in blue, and the type strains are indicated by 'T'.
Analyses 3 represented phylogenetic relationships of two novel taxa, Alternaria arundinis and A. nodulariconidiophora, with other representative species in sect. Infectoriae and sect. Pseudoalternaria. Phylogenetic construction of sect. Infectoriae is based on a combined ITS, GAPDH and ATPase DNA sequence dataset comprising 68 strains of ingroup taxa and two taxa in sect. Chalastospora (A. malorum CBS 135.31 and A. abundans CBS 534.83) were selected as the outgroup taxa. The best scoring RAxML tree is shown in Fig. 5 with the final ML optimization likelihood value of –8,122.101740 (ln). The dataset consists of 2,280 total characters, including gaps (ITS: 1–561 bp, GAPDH: 562–1,081 bp, ATPase: 1,082–2,280 bp). RAxML analysis yielded 428 distinct alignment patterns and 7.89% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.221370, C = 0.311012, G = 0.250900, T = 0.216718, with substitution rates AC = 1.538856, AG = 2.663454, AT = 1.056408, CG = 1.258484, CT = 8.888843, GT = 1.000000. The gamma distribution shape parameter alpha = 0.122301 and the Tree-Length = 0.445658. Bayesian posterior probabilities (PP) from MCMC were evaluated with final average standard deviation of split frequencies = 0.008633. The final alignment and tree were submitted in TreeBASE as submission ID: 258524. Three strains of A. arundinis (MFLU 21-0313A, MFLU 21-0313B, MFLUCC 21-0128) formed a monophyletic subclade (92% ML, 0.99 PP), sister to A. incomplexa E.G. Simmons (CBS 121330) with significant support (89% ML, 1.00 PP), while A. nodulariconidiophora (MFLU 21-0315, MFLUCC 21-0131) clustered with Alternaria sp. (JS8-5, FA3-2) and A. humuli E.G. Simmons (CBS 119404) with significant support (87% ML, 1.00 PP). Many Alternaria spp. isolated from black head mold-affected wheat and barley in Iran were included in the present analyses and remained phylogenetically unresolved, concurring with Poursafar et al.[47]. Unfortunately, phylogenetic affinities of most species in this section are characterized by internally low support values, also in agreement with Poursafar et al.[47] and Marin-Felix et al.[70]. More informative phylogenetic markers such as ATPase and cmdA genes were suggested to use at species level identification due to species in sect. Infectoriae showing high genetic variation[14,15,70].

Figure 5.
Phylogenetic construction of Alternaria sect. Infectoriae using RAxML-based analysis of a combined ITS, GAPDH and ATPase DNA sequence dataset. Bootstrap support values for maximum likelihood (ML, black) equal to or greater than 70% and Bayesian posterior probabilities (PP, red) equal to or greater than 0.95 PP are shown above the nodes. The tree is rooted with taxa in sect. Chalastospora (Alternaria malorum CBS 135.31 and A. abundans CBS 534.83). Newly generated strains are in blue. The type strains obtained from ex-type cultures are indicated by 'T' and the type strains obtained from specimens are indicated by 'H'.
Analyses 4 represented phylogenetic relationships of the novel species, Alternaria brevirostra with other representative species in sect. Porri. Phylogenetic construction of Alternaria sect. Porri based on a combined ITS, GAPDH, TEF1-α, RPB2 and Alt-a1 DNA sequence dataset comprised 114 strains of 65 ingroup species. Five strains of three species in sect. Euphorbiicola (A. limicola CBS 483.90, CBS 117360 and A. euphorbiicola CBS 119460, CBS 198.86) and sect. Gypsophilae (A. gypsophilae CBS 107.41) were selected as the outgroup taxa. The best scoring RAxML tree is shown in Fig. 6 with the final ML optimization likelihood value of –11,626.467690 (ln). The dataset consists of 2,715 total characters, including gaps (ITS: 1–539 bp, GAPDH: 540–1,121 bp, TEF1-α: 1,122–1,463 bp, RPB2: 1,464–2,239 bp, Alt-a1: 2,240–2,715). RAxML analysis yielded 597 distinct alignment patterns and 3.43% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.231369, C = 0.294521, G = 0.244128, T = 0.229982, with substitution rates AC = 0.982217, AG = 4.158740, AT = 0.987304, CG = 0.590117, CT = 8.931720, GT = 1.000000. The gamma distribution shape parameter alpha = 0.207829 and the Tree-Length = 0.669039. Bayesian posterior probabilities (PP) from MCMC were evaluated with the final average standard deviation of split frequencies = 0.008326. The final alignment and tree were submitted in TreeBASE as submission ID: 258525. Two strains of the novel species, A. brevirostra (MFLUCC 21-0129, MFLUCC 21-0130), formed a sister clade with A. rostellata (CBS 117366) and clustered with A. nitrimali (CBS 109163), A. pipionipisi (CBS 116115), A. crassa (CBS 110.38) and A. macrospora (CBS 117128). However, phylogenetic relationships of the species in this subclade were not well-resolved. Concurring with Woudenberg et al.[11,22], sect. Porri displayed a high degree of genetic variation, and the phylogenetic status of many species remain unresolved in this study.
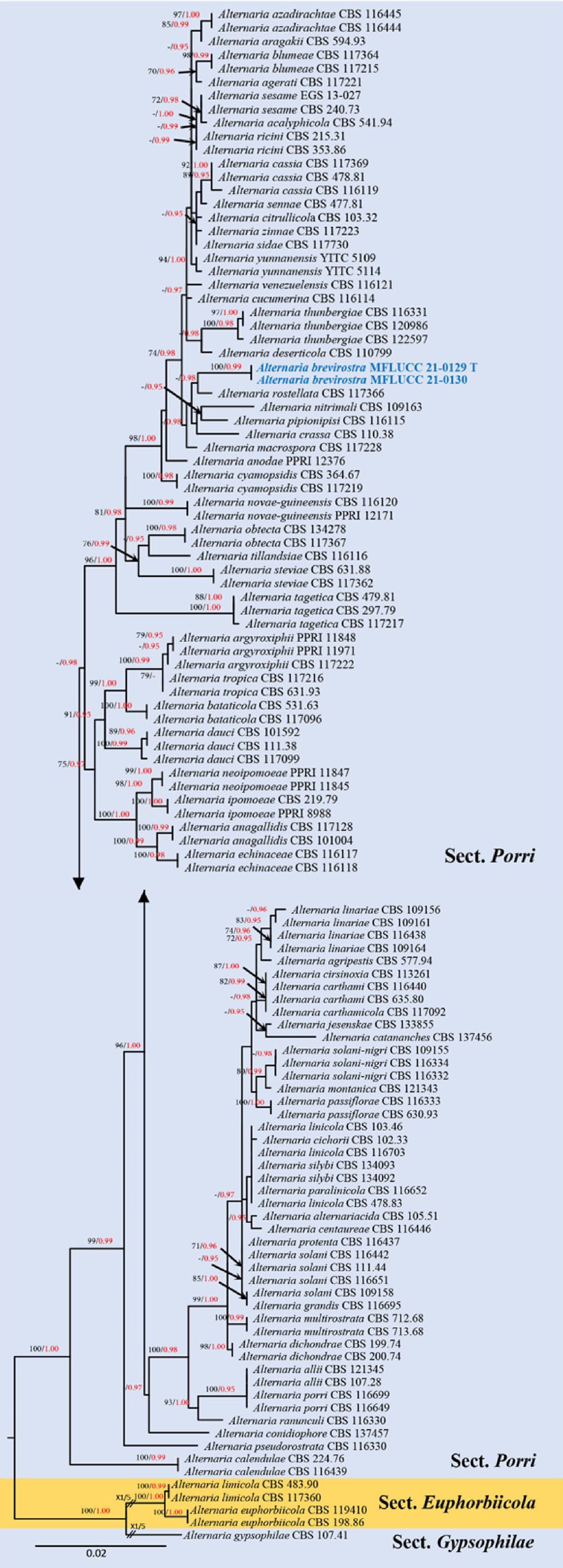
Figure 6.
Phylogenetic construction of Alternaria sect. Porri using RAxML-based analysis of a combined ITS, GAPDH, TEF1-α, RPB2 and Alt-a1 DNA sequence dataset. Bootstrap support values for maximum likelihood (ML, black) equal to or greater than 70% and Bayesian posterior probabilities (PP, red) equal to or greater than 0.95 PP are shown at the nodes. The tree is rooted to sect. Gypsophilae (Alternaria gypsophilae CBS 107.41). Newly generated strains are in blue and the type strains are indicated by 'T'.
Analyses 5 represented phylogenetic relationships of the new taxon, Alternaria phytolaccae, with other species in sect. Radicina and the closely related sect. Gypsophilae. Phylogenetic construction of sect. Radicina based on a combined ITS, TEF1-α, RPB2, and GAPDH DNA sequence dataset comprised 18 strains of ingroup taxa and A. helianthiinficiens (CBS 208.86, CBS 117370) was selected as the outgroup. The best scoring RAxML tree is shown in Fig. 7 with the final ML optimization likelihood value of –5,160.537105 (ln). The dataset consists of 2,209 total characters, including gaps (ITS: 1–521 bp, TEF1-α: 522–767 bp, RPB2: 768–1,636 bp, GAPDH: 1,637–2,209 bp). RAxML analysis yielded 220 distinct alignment patterns and 8.90% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.246820, C = 0.276063, G = 0.242024, T = 0.235092, with substitution rates AC = 1.221528, AG = 4.038587, AT = 0.589501, CG = 0.595656, CT = 9.444647, GT = 1.000000. The gamma distribution shape parameter alpha = 0.191394 and the Tree-Length = 0.206922. Bayesian posterior probabilities (PP) from MCMC were evaluated with final average standard deviation of split frequencies = 0.008477. The final alignment and tree were submitted in TreeBASE as submission ID: 258526. Two strains of the novel species, A. phytolaccae (MFLU 21-0314, MFLUCC 21-0135), formed a strong support clade (98% ML, 1.00 PP) and clustered with A. selini E.G. Simmons (EGS 25-198), A. petroselini (Neerg.) E.G. Simmons (CBS 112.41) and A. vulgaris L. He & J.X. Deng (YZU161234, YZU161235), with significant support (60% ML, 0.97 PP).
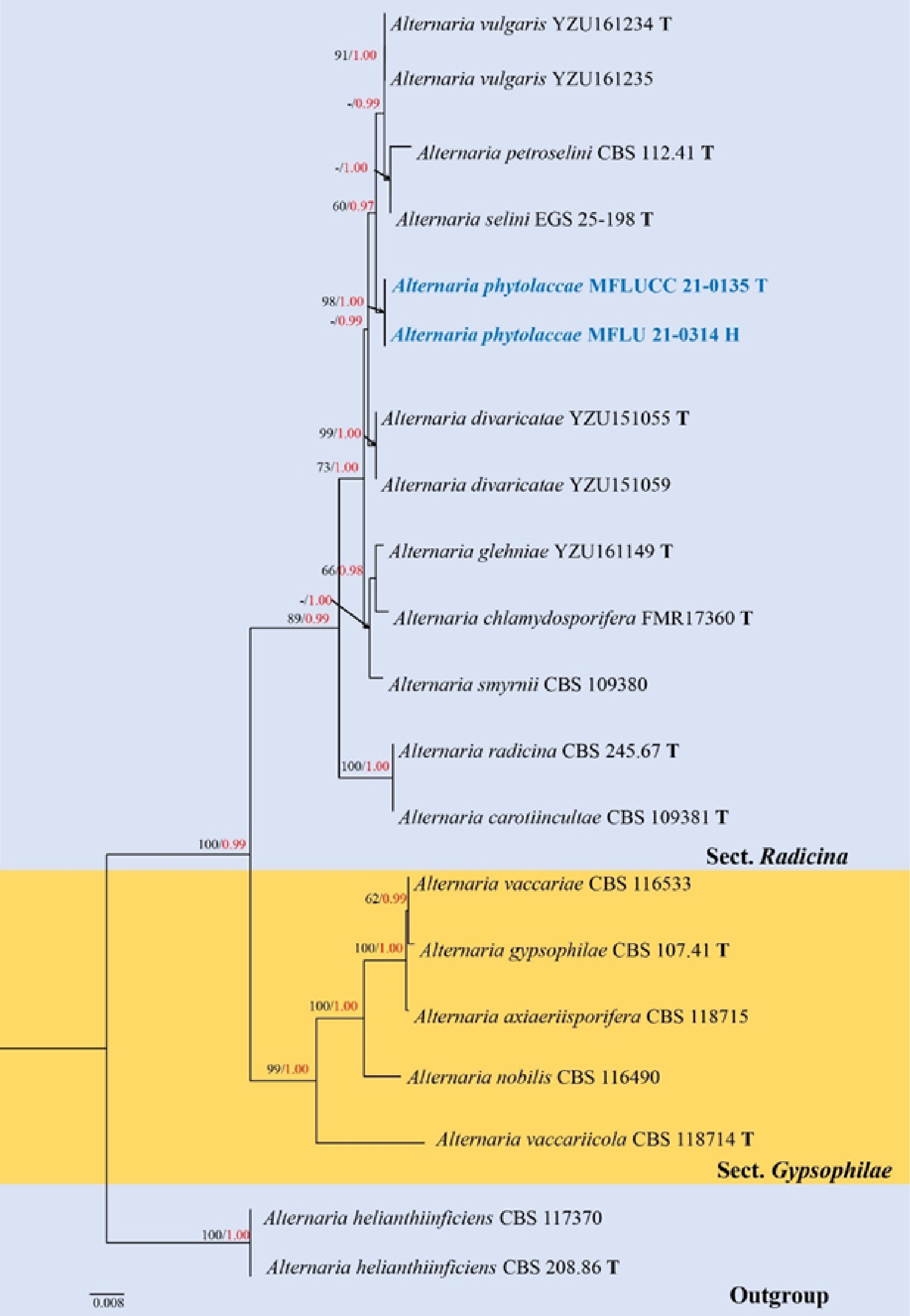
Figure 7.
Phylogenetic construction of Alternaria sect. Radicina using RAxML-based analysis of a combined ITS, TEF1-α, GAPDH and Alt-a1 DNA sequence dataset. Bootstrap support values for maximum likelihood (ML, black) equal to or greater than 60% and Bayesian posterior probabilities (PP, red) equal to or greater than 0.95 PP are shown above the nodes. The tree is rooted to A. helianthiinficiens (CBS 208.86 and CBS 117370). Newly generated strains are in blue. The type strains obtained from ex-type culture are indicated by 'T' and the type strains obtained from holotype specimen are indicated by 'H'.
Taxonomy
-
The current morphology-based taxonomy of Alternaria followed the treatment of Emory G. Simmons (1920–2013), who provided a monograph of Alternaria based on the patterns of sporulation and conidial morphology[38]. In addition, a comprehensive treatment with multi-locus phylogeny-based taxonomy was carried on by Woudenberg et al.[11,12,22]. In the present study, 18 saprobic species have been introduced into sects. Alternaria, infectoriae, Porri and Radicina based on morphological characteristics on host substrates, coupled with multi-locus phylogenetic evidence. In addition, the sporulation of novel species was also induced on OA, PCA and PDA following Emory G. Simmons’s criterion, of which A. arctoseptata, A. baoshanensis, A. breviconidiophora, A. ellipsoidialis, A. eupatoriicola, A. falcata, A. lathyri, A. macilenta, A. minimispora, A. oblongoellipsoidea, A. phragmiticola and A. salicicola in sect. Alternaria, A. arundinis and A. nodulariconidiophora in sect. infectoriae and A. phytolaccae in sect. Radicina, were sporulated on PCA (Fig. 8). Besides, A. alternata, A. doliconidium and A. macroconidia were sporulated on OA. While A. orobanches (sect. Alternaria) and A. brevirostra (sect. Porri) did not sporulate on any agar media.

Figure 8.
Sporulation in Alternaria spp. (a) A. eupatoriicola (MFLU 21-0319). (b) A. lathyri (MFLU 21-0297). (c) A. oblongoellipsoidea (MFLU 21-0310). (d) A. macilenta (MFLU 21-0305). (e) A. baoshanensis (MFLU 21-0296). (f) A. falcata (MFLU 21-0306). (g) A. ellipsoidialis (MFLU 21-0307). (h) A. arctoseptata. (i) A. salicicola (MFLU 21-0320). (j) A. arundinis (MFLU 21-0313). (k) A. phytolaccae (MFLU 21-0314). (l) A. breviconidiophora (MFLU 21-0317). (m) A. phragmiticola (MFLU 21-0316). (n) A. minimispora (MFLU 21-0295). Scale bars: (a)–(n) = 10 µm.
Section Alternaria D.P. Lawr., Gannibal, Peever & B.M. Pryor
Type species – Alternaria alternata (Fr.) Keissl.
Notes – Simmons[160] described the species-groups of Alternaria alternata, A. tenuissima, A. cheiranthi and A. brassicicola based on the morphological characteristics of sporulation. Lawrence et al.[14] revealed eight distinct asexual lineages of Alternaria based on a molecular phylogenetic approach using ten protein-coding loci incorporated extensive taxon sampling (176 species) and proposed eight novel sections for Alternaria, in which sect. Alternaria introduced by Woudenberg et al.[11] assigned an orthographic variant ‘Alternata’ for sect. Alternaria that is contradictory to ICBN Arts. 22.1 and 22.2. Thus, Lawrence et al.[15] resurrected the section named Alternaria. Most species in this section are small-spored, with concatenated conidia that can be found as saprobes and as pre- or post-harvest diseases in over 100 host plants as well as human pathogens[12,15]. Some important plant pathogens in this section such as A. arborescens can cause stem canker on tomato, and A. longipes caused brown spot disease on tobacco[12]. Major updated taxonomic treatment of sect. Alternaria was circumscribed by Woudenberg et al.[12]. The generic type of Alternaria, A. alternata is also accommodated in this section. Alternaria alternata displays high genetic variation, and thus, Woudenberg et al.[12] synonymized 35 morpho-species under A. alternata, of which three formae speciales and two pathotypes of A. alternata were recognized according to the detection of host-specific toxins. Woudenberg et al.[12] mentioned that the genome assembly showed high similarity between the isolates within sect. Alternaria (96.7%–98.2% genome identity) compared with isolates from other sections (85.1%–89.3% genome identity), while the synonymized morpho-species under A. alternata showed 1.4%–1.5% SNPs in their whole-genome reads. As Alternaria isolates were highly polymorphic, the low informative genes of the ITS, LSU and SSU were the least successful in separating the species in sect. Alternaria, while GAPDH was commonly used to distinguish all species in the section, except for distinguishing the A. arborescens species complex (AASC) from A. alternata. The other genes viz. Alt-a1, endoPG, KOG1058, OPA10-2 and RPB2 could separate all species in the section from A. alternata but could not separate different pairs of other species from one another[12].
Alternaria alternata (Fr.) Keissl., Beih. bot. Zbl., Abt. 2 29: 434 (1912)
Index Fungorum number: IF 119834, Facesoffungi number: FoF 03825; Fig. 9

Figure 9.
Alternaria alternata (MFLU 21-0302). (a), (b) Colonies on dead branch. (c)–(i) Conidiophores. (j)–(u) Conidia. Scale bars: (a) = 0.1 cm, (b) = 300 µm, (c)–(u) = 20 µm.
Basionym: Torula alternata Fr., Syst. Mycol. (Lundae) 3: 500. 1832. (nom. sanct.)
Synonyms: See Woudenberg et al.[12] and Index Fungorum[5]
Type details – India, on Arachis hypogaea (Fabaceae), 1 December 1980, E.G. Simmons, IMI 254138 [epitype; designated by de Hoog & Horré[20]], ex-epitype living culture, CBS 916.96 = ATCC 66981 = EGS 34.01; Germany, on fragments of a pithy stem, C.G.D. Nees von Esenbeck, No. 910, 262-129, a [neotype; designated by Simmons][161].
Saprobic on dead branches of Reseda sp. Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, composed of septate, branched, smooth, thin-walled, pale white to grey hyphae. Conidiophores (160.5–)179.5–184(–188) × (9–)12–14(–15.5) µm (
$ \bar{x} $ $ \bar{x} $ $ \bar{x} $ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, cottony, pale white to grey, reaching 5 cm in 7 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, subhyaline hyphae. Conidia sporulated on OA media within 15 d, phragmosporous to muriform, obclavate to obpyriform, light brown to brown, with branched or unbranched acicular or doliiform, aseptate, apical beak, formed branched, apically or laterally secondary conidiophores with 1–2 conidiogenous loci, olivaceous-brown to brown, 1–4 transversely euseptate, 1–3 longitudinal or oblique or Y-shaped septa in transverse divisions, borne in chains with at least 3 conidia, smooth to minutely verruculose.
Material examined – Italy, Province of Arezzo [AR], near Passo la Calla, on dead aerial stem of Reseda sp. (Resedaceae), 13 July 2014, E. Camporesi, IT1994 (MFLU 21-0302), living culture = MFLUCC 21-0797.
Notes – In this study, we obtained 45 new collections of Alternaria alternata from China, Italy and Thailand. Multi-locus phylogeny of ITS, LSU, SSU, TEF1-α, RPB2, GAPDH and Alt-a1 loci confirmed species identification of these 45 strains as A. alternata. These 45 strains formed various internal subclades within A. alternata and can be separated into five main subclades with 80% ML, 0.95 PP support (Fig. 4). These 45 collections showed a diverse range of host occurrences in families Adoxaceae, Arecaceae, Asteraceae, Betulaceae Brassicaceae, Caprifoliaceae, Fagaceae, Lamiaceae, Malavaceae, Orobanchaceae, Pinaceae, Poaceae, Rubiaceae, Resedaceae, Rosaceae, Sapindaceae, Solanaceae, Urticaceae and some unidentified plant litter (Table 5).
Table 5. Additional collections of Alternaria alternata collected from Yunnan, China, Italy and Thailand in this study.
Culture collection Original
codeHerbarium no. Origin Host and habitat Collection date Collector KUNCC 22-10823 IT2053 HKAS 124866
MFLU 15-2585Italy, Province of Forlì-Cesena, Premilcuore Dead stem of Sonchus sp. (Asteraceae) 18 August 2014 E. Camporesi KUNCC 22-10824 IT2063 HKAS 124867 Italy, Province of Forlì-Cesena, Verghereto, Montecoronaro Dead hanging stem of Centaurea sp. (Asteraceae) 20 August 2014 E. Camporesi KUNCC 22-10825 IT2064 HKAS 124868 Italy, Province of Forlì-Cesena, Fiumicello di Premilcuore Dead hanging stem of Helleborus sp. (Ranunculaceae). 28 August 2014 E. Camporesi KUNCC 22-10826 IT2087 HKAS 124869 Italy, Province of Forlì-Cesena, Quattro di Forlì Dead hanging stem of Agropyron sp. (Asteraceae) 1 September 2014 E. Camporesi KUNCC 22-10827 IT2090 HKAS 124870 Italy, Province of Forlì-Cesena, Predappio, Rocca delle Caminate Dead leaf petiole of Robinia sp. (Fabaceae) 4 September 2014 E. Camporesi KUNCC 22-10828 IT2103 HKAS 124871 Italy, Province of Forlì-Cesena, Meldola Dead hanging stem of Echinochloa
sp. (Poaceae)8 September 2014 E. Camporesi KUNCC 22-10829 IT2114 HKAS 124872 Italy, Province of Forlì-Cesena, Tessello Dead hanging stem of Cephalaria sp. (Dipsacaseae) 16 September 2014 E. Camporesi KUNCC 22-10830 IT2115 HKAS 124873 Italy, Province of Forlì-Cesena, Civitella di Romagna Dead hanging stem of Aster sp. (Asteraceae) 19 September 2014 E. Camporesi KUNCC 22-10831 IT2125 HKAS 124874 Italy, Province of Arezzo, Stia, Montemezzano Dead hanging stem of Euphorbia sp. (Euphorbiaceae) 22 September 2014 E. Camporesi KUNCC 22-10832 IT2143 HKAS 124875 Italy, Province of Forlì-Cesena, Galeata, San Zeno Dead hanging leaf petiole of
Ailanthus sp. (Simaroubaceae)30 September 2014 E. Camporesi KUNCC 22-10833 IT2144 HKAS 124876 Italy, Province of Forlì-Cesena,
Cabelli di Santa SofiaDead hanging stem of Agrostis sp. (Poaceae) 2 October 2014 E. Camporesi KUNCC 22-10834 IT2145 HKAS 124877 Italy, Province of Forlì-Cesena,
Monte MirabelloDead hanging leaf of Arundo sp. (Poaceae) 3 October 2014 E. Camporesi KUNCC 22-10835 IT2162 HKAS 124878 Italy, Province of Forlì-Cesena,
Santa SofiaDead hanging stem of Hedysarum coronarium L. (Papilionaceae) 7 October 2014 E. Camporesi KUNCC 22-10836 IT2166 HKAS 124879 Italy, Province of Forlì-Cesena, Meldola, Piandispino Dead hanging stem of Dipsacus sp. (Caprifoliaceae) 7 October 2014 E. Camporesi KUNCC 22-10837 IT2277 HKAS 124880 Italy, Province of Ravenna, Lido di Dante Dead hanging stem of Kali tragus (L.) Scop. (Amaranthaceae) 2 December 2014 E. Camporesi KUNCC 22-10838 IT2347 HKAS 124881 Italy, Province of Forlì-Cesena,
Collina di ForlìSeveral samaras of Fraxinus oxycarpa Willd. (Oleaceae) 21 January 2015 E. Camporesi KUNCC 22-10839 IT2427 MFLU 15-1823 Italy, Province of Forlì-Cesena, Forlì, Via Nenni Dead hanging stem of Wisteria sp. (Caprifoliaceae) 30 March 2015 E. Camporesi KUNCC 22-10840 IT2454 HKAS 124883 Italy, Province of Forlì-Cesena, Predappio, Rocca delle Caminate Dead stem of Urtica sp. (Urticaceae) 21 April 2015 E. Camporesi KUNCC 22-10841 IT2900 MFLU 16-1116 Italy, Province of Forlì-Cesena, Santa Sofia, Camposonaldo Dead needles of Pinus nigra
J.F. Arnold (Pinaceae)23 March 2016 E. Camporesi KUNCC 22-10842 IT3048 MFLU 16-2277 Italy, Province of Forlì-Cesena, Fiumicello di Premilcuore Dead hanging stem of Acer opalus Mill. (Sapindaceae) 27 July 2016 E. Camporesi KUNCC 22-10843 IT3160 MFLU 16-2883 Italy, Province of Forlì-Cesena, Forlì, Ravaldino in Monte Dead stem of Silybum marianum (L.) Gaertn. (Asteraceae) 15 November 2016 E. Camporesi KUNCC 22-10844 IT3168 MFLU 16-2904 Italy, Province of Forlì-Cesena, Forlì, Parco Urbano Dead hanging fruits of Ostrya carpinifolia Scop. (Betulaceae) 19 November 2016 E. Camporesi KUNCC 22-10845 IT3419 HKAS 124888 Italy, Province of Arezzo, Poppi, Quota Dead hanging stem of Calamintha nepeta (Lamiaceae) 25 July 2017 E. Camporesi KUNCC 22-10846 IT3426 HKAS 124889 Italy, Province of Arezzo, near Croce di Pratomagno Dead hanging stem of Malva alcea L. (Malavaceae) 1 August 2017 E. Camporesi KUNCC 22-10847 IT3439 HKAS 124890 Italy, Province of Arezzo, Stia, Montemezzano Dead hanging stem of Reseda luteola L. (Resedaceae) 13 August 2017 E. Camporesi KUNCC 22-10848 IT3442 HKAS 124891 Italy, Province of Arezzo, Montemignaio Dead hanging stem of Valeriana sp. (Caprifoliaceae) 12 August 2017 E. Camporesi KUNCC 22-10849 IT3449 HKAS 124892 Italy, Province of Forlì-Cesena, Bagno di Romagna, Riofreddo Dead hanging stem of Tanacetum
sp. (Asteraceae)22 August 2017 E. Camporesi KUNCC 22-1050 IT3504 MFLU 17-1778 Italy, Province of Forlì-Cesena, Bagno di Romagna, Acquapartita Dead hanging stem of Orobanche
sp. (Orobanchaceae)25 September 2017 E. Camporesi KUNCC 22-10851 IT3556 HKAS 124894 Italy, Province of Forlì-Cesena, Santa Sofia Dead leaf of Sorbus aria Crantz (Rosaceae) 15 November 2017 E. Camporesi KUNCC 22-10852 IT3598 HKAS 124895 Italy, Province of Ravenna, Faenza, Santa Lucia Dead leaves of Quercus pubescens Willd. (Fagaceae) 13 December 2017 E. Camporesi KUNCC 22-10853 IT3651 HKAS 124896 Italy, Province of Forlì-Cesena, Meldola Dead hanging branch of Sambucus nigra L. (Adoxaceae) 31 December 2017 E. Camporesi KUNCC 22-10854 KIB-H2 HKAS 124897 China, Yunnan, Kunming Institute of Botany Dead fallen leave of bamboo (Poaceae) 26 December 2014 J.F. Li KUNCC 22-10855 HKM-1 HKAS 124898 China, Yunnan, Kunming, Xundian Dead stem of Raphanus sativus L. (Brassicaceae) 13 March 2015 J.F. Li KUNCC 22-10856 H-49 HKAS 124899 China, Yunnan, Baoshan Dead branch of Capsicum annuum L. (Solanaceae) 22 October 2015 J.F. Li KUNCC 22-10857 PB-12 HKAS 124900 China, Yunnan, Pingbian, Dawei Mountain Dead stem of Zea mays L. (Poaceae) 20 September 2017 J.F. Li KUNCC 22-10858 HXB-08 HKAS 124901 China, Yunnan, Xishuangbanna Dead fallen leaves of Dimocarpus longan Lour. (Sapindaceae) 8 June 2018, J.F. Li KUNCC 22-10859 HAM-02 HKAS 124902 China, Yunnan, Honghe, Amu Mountain Dead fallen leaves of grass (Poaceae) 15 June 2018 J.F. Li KUNCC 22-10860 HAM-03 HKAS 124903 China, Yunnan, Honghe, Amu Mountain Dead aerial stem of Capsicum
annuum (Solanaceae)15 June 2018 J.F. Li KUNCC 22-10861 HSH-01 HKAS 124904 Thailand, Chiang Rai, Muang, Singha Park Dead culms of bamboo (Poaceae) 23 February 2016 J.F. Li KUNCC 22-10862 HMRC-51 HKAS 124905 Thailand, Chiang Mai, Mae Taeng, Mushroom Research Center (M.R.C) Dead fallen leaves of unidentified plant 23 March 2016 J.F. Li KUNCC 22-10863 DMS-15 HKAS 124906 Thailand, Chiang Rai, Doi Mae Salong Dead leaves of grass (Poaceae) 24 May 2016 J.F. Li KUNCC 22-10864 HTWD-01 HKAS 124807 Thailand, Chiang Rai, Mae Fah Luang Dead leaves of palm (Arecaceae) 25 September 2016 J.F. Li KUNCC 22-10865 H-71 HKAS 124908 Thailand, Chiang Rai, Doi Chang Dead leaves of Coffea arabica L. (Rubiaceae) 25 July 2018 J.F. Li KUNCC 22-10866 HWP-01 HKAS 124909 Thailand, Chiang Rai, Wiang Pa Pao Dead stems of Bidens pilosa L. (Asteraceae) 16 October 2018 J.F. Li The major subclade A formed a significant subclade with 81% ML, 0.98 PP support containing 62 strains with several internal branches. Newly generated strains HWP-01, IT2144, IT2145, IT3598, and IT3651 formed a single lineage with strains CBS 965.95 and CBS 966.95; these isolates occurred on host families Asteraceae, Adoxaceae, Fagaceae, Poaceae and Solanaceae as pathogens and saprobes. The saprobic strains H-71, IT2143, and IT3556 formed a single lineage with strain CBS 130254 (isolated from human sputum in India), and CBS 911.97 (isolated from Artemisia brevifolia (Asteraceae) in India) and clustered with CBS 130262, CBS 130265 (isolated from human sputum in India) and CBS 121492 (isolated from Cucumis melo (Cucurbitaceae) in China). Six strains isolated from human sputum and sinusitis in India (CBS 877.95, CBS 130255, CBS 130258, CBS 130259, CBS 130261, CBS 130263) clustered with three strains isolated from Malus domestica (Rosaceae) in South Africa (CBS 113013, CBS 113014, CBS 113015), strains isolated from Citrus jambhiri (CBS 102596), C. reticulata (CBS 102600) and Minneola tangolo (CBS 119399) in the USA, three new strains isolated from Aster sp. (IT2115), Orobanche sp. (IT3504) in Italy and unidentified grass (DMS 15) in Thailand. At the same time, the strain CBS 117143 (isolated from Capsicum annuum (Solanaceae) in Italy) formed a significant subclade (79% ML, 0.96 PP) with CBS 113054 (isolated from M. domestica in South Africa), and clustered with the two new isolates IT 1994 and IT 2166.
The strain CBS 120829 (isolated from Punica granatum (Punicaceae) in Greece), formed a single lineage with CBS 124277 (isolated from Prunus sp. (Rosaceae) in Denmark) and clustered with CBS 117130 (isolated from Arbutus unedo (Ericaceae) in Italy) with significant support in BI analyses (0.98 PP). Five strains isolated from Minneola tangelo in Israel, South Africa and Turkey (CBS 102559, CBS 102602, CBS 102603, CBS 121344, and CBS 121346) also formed a single lineage (66% ML, 0.99 PP) within subclade A. The strain CBS 124278 (isolated from Prunus sp. in Denmark) formed a single lineage with CBS 102604 (isolated from Minneola tangelo in Israel) with significant support (68% ML, 0.96 PP), while at the same time, the new isolate IT2900 formed a single lineage with CBS 195.86 (isolated from Euphorbia esula (Euphorbiaceae) in Canada) with high support (100% ML, 1.00 PP). Three new isolates (HAM 02, IT 2087, IT 3426) formed a single lineage with CBS 603.78 (isolated from the air in USA) and clustered with CBS 113024 (isolated from Minneola tangelo in South Africa). Two new isolates (IT 2063, IT 3168) formed a significant branch with 87% ML, 1.00 PP support clustered with CBS 447.86 (isolated from Malva sp. (Malvaceae) in Morocco). Three new isolates (HXB 08, IT3419, IT2064) also formed a significant support lineage (73% ML, 0.99 PP) with CBS 479.90 (isolated from Citrus unshiu in Japan) and clustered with IT2347, HKM 1 and CBS 126071 (isolated from soil in Namibia) with significant support (87% ML, 1.00 PP). The strain CBS 112252 formed an independent single lineage basal to subclade A with significant support (81% ML, 1.00 PP).
Subclade B was represented by five strains, including the ex-type strain of Alternaria alternata (CBS 916.96). Two new isolates (H-49, IT2162) formed a single lineage with the type strain of A. alternata (CBS 916.96) and CBS 116749 with 96% ML, 1.00 PP support, clustering with CBS 106.34 (isolated from Linum usitatissimum, Linaceae) with 81% ML, 0.99 PP support.
Subclade C contained seven new isolates from China and Italy (IT2053, KIB-H2, IT 2277, IT 3160, IT 3439, IT 2090 and HAM-03), clustering with strain CBS 119115 (isolated from Prunus sp. in Greece), two strains from South Africa (CBS 115069, CBS113025), four strains from USA (CBS 127672, CBS 267.77, CBS 119543, CBS 127671), two strains from India (CBS 130260, CBS 125606), CBS 686.68 (from Sahara Desert sand) and CBS 104.26, with 96% ML, 1.00 PP support.
Subclade D formed a high support subclade (99% ML, 0.95 PP) containing five strains that include two new strains (IT 3048, IT2454) isolated from Acer opalus and Urtica sp. in Italy. The two new strains (IT 3048, IT2454) formed a single lineage with CBS 117.44 (isolated from Godetia sp. in Denmark) with 89% ML, 1.00 PP support, clustering with CBS 106.24 (isolated from Malus sylvestris in USA) and CBS 121544 (isolated from Cucumis sativus in USA).
Subclade E formed a basal lineage with 86% ML, 1.00 PP support, including ten new isolates (IT2103, IT 3442, IT 3449, IT 2114, IT 2427, HSH-01, PB-12, HTWD-01, IT 2125, HMRC-51), two isolates from soil (CBS 127334, CBS 126910) and from Plantago aristida (Plantaginaceae) (CBS 795.72) in USA, two strains isolated from Minneola tangelo in South Africa (CBS 115200, CBS 115199), an isolate from Fragaria vesca in Belgium (CBS 880.95), an isolate from Cyperaceae in Papua New Guinea (CBS 806.96) and two strains isolated from Broussonetia papyrifera (CBS 121455) and Pyrus bretschneideri (CBS 121547).
Alternaria arctoseptata J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558434; Facesoffungi number: FoF12652; Fig. 10

Figure 10.
Alternaria arctoseptata (MFLU 21-0308, holotype). (a) Colonies on dead stem of Lathyrus sp. (Fabaceae). (b)–(g) Conidiophores bearing conidiogenous cells. (h), (i) Immature conidia. (j), (o) Conidia formed secondary conidiophores. (k)–(n) Mature conidia. Scale bars: (a) = 200 µm, (b)–(o) = 20 µm.
Etymology: Referring to the constricted septate conidia.
Holotype: MFLU 21-0308
Saprobic on dead standing stem of Lathyrus sp. (Fabaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, composed of dark brown hyphae. Conidiophores 50–100 × 8–12 µm (
$ \bar{x} $ $ \bar{x} $ $ \bar{x} $ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, hairy, fluffy, brown to dark brown, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, white to brown hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, white to light brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, hyaline to subhyaline hyphae, 2–5 µm diam; conidia formed in vitro within 30 d, borne in chains with at least two conidia, yellow to light brown, subglobose to ellipsoidal, 55 × 20 µm (Fig. 8h).
Material examined – Italy, Province of Forlì-Cesena, Predappio, Fiumana, on dead standing stem of Lathyrus sp. (Fabaceae), 2 September 2014, E. Camporesi, IT2088 (MFLU 21-0308, holotype), ex-type living culture = MFLUCC 21-0139.
Notes – Alternaria arctoseptata was isolated from the same host genus Lathyrus sp. as A. lathyri. Alternaria arctoseptata can be distinguished from A. lathyri in having paler conidia, with short narrow apical beak or formed secondary conidiophores. Furthermore, the conidiophores of A. arctoseptata are shorter, pale brown to light brown conidiophores with apically swollen knots, arising from stomatic base, which are darker in A. lathyri. Phylogenetically two strains of A. arctoseptata (MFLUCC 21-0139, MFLU 21-0308) formed a high support clade (100% ML, 0.99 PP) that clustered with A. baoshanensis and A. ovoidea with 86% ML, 0.99 PP support (Fig. 3), and distant from A. lathyri. Alternaria arctoseptata differs from A. baoshanensis in having apical beak, and larger conidia (60 × 25 µm vs. 38 × 18 µm) that are rather constricted at some septa as well as having rather monotretic, larger conidiogenous cells (11.5 × 12 µm vs. 6.2 × 7.5 µm). Alternaria arctoseptata also differs from A. ovoidea in having paler brown conidia that are rather constricted at some septa[68]. A comparison of RPB2 nucleotide pairwise shows that A. arctoseptata differs from A. baoshanensis in 10/559 bp (1.8% difference, no gap) and differs from A. ovoidea in 12/565 bp (2.1% difference, no gap). A comparison of Alt-a1 nucleotide pairwise shows that A. arctoseptata differs from A. baoshanensis in 8/474 bp (1.7% difference, no gap) and differs from A. ovoidea in 18/520 bp (3.5% difference, no gap). Based on distinct morphological characteristics and phylogenetic support, A. arctoseptata is introduced as a new species in this study.
Alternaria baoshanensis J.F. Li, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558435; Facesoffungi number: FoF 12653; Fig. 11

Figure 11.
Alternaria baoshanensis (MFLU 21-0296, holotype). (a) Colonies on dead rattan of Curcubita moschata. (b)–(e) Conidiophores with a distinct conidiogenous locus. (f)–(l) Variation in shape of conidia. Scale bars: (a) = 200 µm, (b)–(e) = 30 µm, (f)–(l) = 20 µm.
Etymology: Named after the locality, Baoshan (Yunnan, China), where the species was collected.
Holotype: MFLU 21-0296
Saprobic on rattan of Curcubita moschata (Duch ex Lam.) Duch ex Poiret (Cucurbitaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, composed of septate, branched, smooth, thin-walled, composed of dark hyphae. Conidiophores 80–100 × 12–20 µm (
$ \bar{x} $ $\bar {x} $ $ \bar {x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, hairy fluffy, brown, reaching 5 mm in 10 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, brown hyphae; conidia not formed in vitro within 60 days. Colonies growing on PCA, white to light brown colored, cottony, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, subhyaline to light brown hyphae, 2–5 µm diam; conidia formed in vitro within 60 d, borne in chains with at least 2 conidia, light brown to brown, ovoid to ellipsoidal, 40 × 15 µm (Fig. 8e).
Material examined – China, Yunnan, Baoshan, Shuizhai County, on dead rattan of Curcubita moschata (Cucurbitaceae), 25 October 2015, J.F. Li, H-50 (MFLU 21-0296, holotype), ex-type living culture = MFLUCC 21-0124.
Notes – Alternaria baoshanensis can be distinguished from other related species (A. actroseptata and A. ovoidea) in having versicolorous conidiophores[68]. Multi-locus phylogeny demonstrated that two strains of A. baoshanensis (MFLUCC 21-0124, MFLU 21-0296) form a robust clade (100% ML, 1.00 PP), sister to A. ovoidea with 67% ML, 0.96 PP support (Fig. 3). Alternaria baoshanensis differs from A. ovoidea in having shorter conidiogenous cells, with apically or laterally 1–2 conidiogenous loci (5–7 × 6–10 µm vs. 9–13 × 8.5–15 µm), while A. ovoidea has conidiogenous loci cicatrized on conidial secession[68]. Furthermore, A. baoshanensis has slightly smaller conidia (25–60 × 12–22 µm vs. 48–65 × 15.5–30 µm)[68]. Conidia of A. baoshanensis are varied in shapes, usually subglobose to ellipsoidal, solitary or borne in chain (at least 1–3 conidia), light brown to yellowish brown, 3–6 transverse septa, with 1–2 longitudinal or oblique septa in some transverse divisions. Whereas, conidia of A. ovoidea are solitary, ovoid, orangish brown to copper brown, with 1–3 transverse septa, and 1 longitudinal septum in transverse divisions[68].
Alternaria breviconidiophora J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558436; Facesoffungi number: FoF 12654; Fig. 12

Figure 12.
Alternaria breviconidiophora (MFLU 21-0317, holotype). (a) Colonies on dead branch. (b) Conidiophores arising on stomatic base. (c)–(g) Conidiophores. (h)–(q) Conidia. Scale bars: (a) = 50 µm, (b)–(g) = 20 µm, (h)–(q) = 10 µm.
Etymology: Named after its short conidiophores and small conidial structures.
Holotype: MFLU 21-0317
Saprobic on dead hanging branches of Digitalis sp. (Scrophulariaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, composed of brown to dark brown hyphae. Conidiophores 25–88 × 6–10 µm (
$ \bar {x}$ $ \bar {x}$ $ \bar {x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, cottony, brown to dark brown, reaching 5 mm in 10 d at 25ºC, mycelium superficial, effuse, radially striate, light brown hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, light brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, subhyaline to light brown hyphae, 2–5 µm diam; conidia formed in vitro within 60 d, borne in chains with at least 2 conidia, light brown to brown, globose or subglobose, 12 × 10 µm (Fig. 8l).
Material examined – Italy, Province of Forlì-Cesena, Rocca San Casciano, on dead hanging branch of Digitalis sp. (Scrophulariaceae), 7 April 2017, E. Camporesi, IT3308 (MFLU 21-0317, holotype), ex-type living culture = MFLUCC 22-0075.
Notes – Multi-locus phylogeny showed that two strains of Alternaria breviconidiophora form a robust subclade (100% ML, 1.00 PP; Fig. 3) basal to A. lathyri, A. muriformispora, and A. pseudoinfectoria with 100% ML and 1.00 PP support (Fig. 3). However, A. breviconidiophora differs from these three species in having small (
$ \bar {x}$ Alternaria doliconidium J.F. Li, Camporesi & K.D. Hyde, in Wanasinghe et al., Fungal Diversity: 10.1007/s13225-018-0395-7, (2018)[147]
Index Fungorum number: IF554202; Facesoffungi number: FoF 04041; Fig. 13

Figure 13.
Alternaria doliconidium (MFLU 21-0294). (a) Dead stem of Reseda sp. (Resdaceae). (b) Colonies on dead stem. (c)–(j) Conidiophores on natural substrate. (k)–(m) Conidiogenesis sporulated in vitro. (n) Germinated conidium. (o)–(r) Conidia ((q), (r) on natural substrate)). Scale bars: (a) = 0.1 cm, (b) = 500 µm, (c)–(r) = 20 µm.
Type details – ITALY, Province of Forli-Cesena [FC], Raggio di Santa Sofia, on dead aerial spines of Rosa canina L. (Rosaceae), 16 October 2014, E. Camporesi, IT2165 (KUN-HKAS100840, holotype), ex-type living culture = KUMCC 17-0263.
Saprobic on dead stems of Reseda sp. Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, with dark brown hyphae. Conidiophores (140–)167–176(–189.5) × 8–10(–10.5) µm (
$ \bar {x}$ $ \bar {x}$ $ \bar {x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from lateral cells. Colonies growing on PDA, hairy fluffy or cottony, grey to brown, reaching 5 cm in 15 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, grey to light brown hyphae. Conidia sporulated on OA within 30 d, phragmosporous to muriform, oblong to subglobose, with short, branched or unbranched, aseptate apical beak, formed apically, or laterally secondary conidiophores with 1 conidiogenous locus, olivaceous-brown to golden brown, sectored, 2–3 transversely eusepta, with 0–1 longitudinal or oblique or Y-shaped septum in transverse divisions, borne in chains with at least 2–4 conidia, verruculose to verrucose .
Material examined – China, Yunnan Province, Kunming, Kunyang Town, on dead stem of Reseda sp. (Resedaceae), 18 September 2014, J.F. Li, H-11B (MFLU 21-0294, new geographical and host record), living culture MFLUCC 14-0020.
Notes – Wanasinghe et al.[66] described Alternaria doliconidium on Rosa canina L. (Rosaceae) from Italy. In this study, we collected A. doliconidium on Reseda sp. as the first record on this host from China. Alternaria doliconidium was isolated from the same host species as A. alternata (MFLU 21-0302, MFLUCC 21-0797) collected in Italy. A nucleotide pairwise comparison showed that our strain MFLUCC 14-0020 differs from A. alternata (MFLUCC 21-0797) in 10/520 bp (1.9% difference, no gap) of GAPDH and 8/500 bp (1.6% difference, no gap) of Alt-a1 gene locus. Multi-locus phylogeny (Fig. 3) showed that our strain (MFLUCC 14-0020) forms a high support clade (99% ML, 1.00 PP; Fig. 3) with the ex-type strains of A. doliconidium (KUN-HKAS 100840, MFLUCC 17-0263) and clustered with A. italica (KUMCC 17-0123, MFLUCC 14-0421) and A. alternata (CBS 102595, CBS 175.52, YL1, YL2, CBS 916.96) with significant support in ML analysis (71% ML; Fig. 3). A comparison of nucleotide pairwise similarities showed that A. doliconidium is distinct from A. italica, and A. alternata (CBS 102595, CBS 175.52, YL1, YL2, CBS 916.96) in Alt-a1, GAPDH, RBP2, and TEF1-α (Table 6). However, the phylogenetic relationship between A. doliconidium and A. italica with A. alternata is unresolved in this study, pending further study.
Table 6. Nucleotide base comparison of Alternaria doliconidium (MFLUCC 14-0020) with closely related taxa.
Species name Strain no. Nucleotide difference of gene sequences (no gaps) A. doliconidium MFLUCC 14-0020 ITS TEF1-α RBP2 GAPDH Alt-a1 A. alternata CBS 102595 8/520 bp (1.5%) 14/256 bp (4.7%) 30/875 bp (3.4%) 16/582 bp (2.7%) 12/476 bp (2.5%) A. alternata CBS 916.96 8/520 bp (1.5%) 14/256 bp (4.7%) 28/875 bp (3.2%) 18/582 bp (3.1%) 12/476 bp (2.5%) A. alternata CBS 175.52 10/520 bp (1.9%) 16/256 bp (6.3%) 30/875 bp (3.4%) 17/582 bp (2.9%) 12/476 bp (2.5%) A. alternata YL1 9/520 bp (1.7%) 14/256 bp (4.7%) 30/875 bp (3.4%) 16/582 bp (2.7%) 12/476 bp (2.5%) A. alternata YL2 9/520 bp (1.7%) 14/256 bp (4.7%) 30/875 bp (3.4%) 17/582 bp (2.9%) 12/476 bp (2.5%) A. doliconidium HKAS 100840 1/520 bp, (0.2%) 0% N/A N/A N/A A. doliconidium MFLUCC 17-0263 1/520 bp, (0.2%) 0% N/A N/A N/A A. italica MFLUCC 14-0231 6/520 bp, (1.2%) 11/256 bp (4.3%) 25/875 bp, (2.9%) N/A N/A A. italica KUMCC 17-0123 7/520 bp, (1.3%) N/A N/A N/A N/A Alternaria ellipsoidialis J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558437; Facesoffungi number: FoF12655; Fig. 14

Figure 14.
Alternaria ellipsoidialis (MFLU 21-0307, holotype). (a) Colonies on dead hulls of Brassica sp. (b)–(g) Conidiophores bearing conidiogenous cells with a few apical conidiogenous loci. (h)–(q) Conidia. Scale bars: (a) = 300 µm, (b)–(g) = 30 µm, (h)–(q) = 20 µm.
Etymology: Named after its ellipsoidal conidia.
Holotype: MFLU 21-0307
Saprobic on dead hanging hulls of Brassica sp. (Brassicaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, with dark brown hyphae. Conidiophores 65–188 × 5–8 µm (
$ \bar {x}$ $ \bar {x}$ $ \bar {x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, hairy fluffy, brown to dark brown, reaching 5 cm in 10 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, white to grey-white hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, white to light brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia formed in vitro within 60 d, borne in chains with at least 2 conidia, light brown to brown, ellipsoidal to ovoid, or obpyriform, 60 × 30 µm (Fig. 8g).
Material examined – Italy, Province of Arezzo, Stia, Papiano, on dead hanging hulls of Brassica sp. (Brassicaceae), 1 September 2014, E. Camporesi, IT2086 (MFLU 21-0307, holotype), ex-type living culture = MFLUCC 21-0132.
Notes – Alternaria ellipsoidialis resembles A. falcata due to its ellipsoidal conidia with short beak and curved conidiophores. However, A. ellipsoidialis differs from A. falcata in having solitary conidiophores with several geniculate conidiogenous loci proliferations at apex, which is rather polytretic than A. falcata. In multi-locus phylogenetic analyses (Fig. 3), A. ellipsoidialis formed an independent subclade closely related with A. eupatoriicola and distant from A. falcata. Alternaria ellipsoidialis can be distinguished from A. eupatoriicola in having larger (60 × 28 µm vs. 48 × 20 µm), pale to yellowish brown conidia, and the conidiophores are more twisted at the apex in A. ellipsoidialis. A RPB2 nucleotide base comparison showed that A. ellipsoidialis differs from A. eupatoriicola in 9/550 bp (1.6% difference, no gap). In GAPDH, A. ellipsoidialis differs from A. eupatoriicola in 13/520 bp (2.5% difference, no gap), and in Alt-a1 the species differs from A. eupatoriicola in 10/480 bp (2.1% difference, no gap). Therefore, the new species A. ellipsoidialis is established.
Alternaria eupatoriicola J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558438; Facesoffungi number: FoF 12656; Fig. 15

Figure 15.
Alternaria eupatoriicola (MFLU 21-0319, holotype). (a) Colonies on dead hanging stem of Eupatorium cannabinum. (b)–(f) Conidiophores bearing conidiogenous cells. (g)–(q) Conidia. Scale bars: (a) = 300 µm, (b)–(m) = 20 µm.
Etymology: Named after the host genus 'Eupatorium cannabinum'.
Holotype: MFLU 21-0319
Saprobic on standing dead stem of Eupatorium cannabinum L. (Asteraceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, thick-walled, with dark hyphae. Conidiophores 50–160 × 5–8 µm (
$ \bar {x}$ $ \bar {x}$ $ \bar {x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from both end cells. Colonies growing on PDA, hairy fluffy, brown to dark brown, reaching 5 cm in 20 d at 25 ºC, mycelium superficial, effuse, radially striated, with irregular edge, white to light dark brown hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, white to brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia formed in vitro within 30 d, borne in chains with at least 2 conidia, yellow to brown, ovoid to obpyriform, 50 × 20 µm (Fig. 8a).
Material examined – Italy, Province of Arezzo, Badia Prataglia, on dead standing stem of Eupatorium cannabinum (Asteraceae), 2 October 2017, E. Camporesi, IT3518 (MFLU 21-0319, holotype), ex-type living culture = MFLUCC 21-0122.
Notes – Based on multi-locus phylogenetic analyses, two strains of Alternaria eupatoriicola formed a robust clade (100% ML, 1.00PP; Fig. 3), distinct from other species within sect. Alternaria. The species is basal to A. pseudoinfectoria, A. muriformispora, A. lathyri, A. breviconidiophora, A. torilis, A. phragmiticola, A. oblongoellipsoidea, and A. macilenta with 84% ML and 1.00 PP support.
Alternaria falcata J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558439; Facesoffungi number: FoF 12657; Fig. 16

Figure 16.
Alternaria falcata (MFLU 21-0306, holotype). (a) Colonies on dead hanging stem of Atriplex sp. (b)–(g) Conidiophores bearing conidiogenous cells. (h)–(m) Conidia. Scale bars: (a) = 150 µm, (b)–(m) = 20 µm.
Etymology: Referring to the curve conidiophores.
Holotype: MFLU 21-0306
Saprobic on dead standing stem of Atriplex sp. (Chenopodiaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, composed of dark, septate, branched hyphae. Conidiophores 70–130 × 5–8 µm (
$ \bar {x}$ $ \bar {x}$ $ \bar {x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, hairy or cottony, brown to dark brown, reaching 5 mm in 20 d at 25 ºC, mycelium superficial, effuse, radially striated, with irregular edge, white to light grey colored hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, light brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia formed in vitro within 60 d, borne in chains with at least 2 conidia, ellipsoidal or obpyriform, 45 × 15 µm (Fig. 8f).
Material examined – Italy, Province of Forlì-Cesena, Fiumicello di Premilcuore, on dead standing stem of Atriplex sp. (Chenopodiaceae), 29 August 2014, E. Camporesi, IT2079 (MFLU 21-0306, holotype), ex-type living culture = MFLUCC 21-0123.
Notes – In multi-locus phylogenetic analyses, two strains of Alternaria falcata formed an independent subclade (81% ML, 0.97 PP; Fig. 3) basal to A. obpyriconidia (MFLUCC 21-0121, MFLU 21-0300), A. macroconidia (MFLUCC 21-0134, MFLU 21-0301), A. salicicola (MFLUCC 22-0072, MFLU 21-0320), A. arctoseptata (MFLUCC 21-0139, MFLU 21-0308), A. ovoidea (MFLUCC 14-0427, MFLU 21-0298) and A. baoshanensis (MFLUCC 21-0124, MFLU 21-0296), respectively. Alternaria falcata differs from A. arctoseptata in having smaller (40 × 15 µm vs. 60 × 25 µm), subglobose to ellipsoidal, olivaceous-brown to brown conidia, solitary conidiophores with a curved apex, while its conidia are rather ellipsoidal to obpyriform or obclavate, brown to dark brown, constricted at the septa and conidiophores aggregated on the stomatic base in A. arctoseptata. Alternaria falcata differs from A. baoshanensis in having larger (40 × 15 µm vs. 38 × 18 µm) and longer (96 × 7 µm vs. 48 × 14 µm) solitary concolorous conidiophores, while the species differs from A. ovoidea in having smaller (40 µm × 15 µm vs. 55 × 27 µm) and paler brown conidia, with a short beak, and shorter (96 × 7 µm vs. 280 × 8 µm), curved conidiophores[68]. A RPB2 nucleotide pairwise comparison showed that A. falcata differs from A. arctoseptata in 10/565 bp (1.8% difference, no gap), differs from A. baoshanensis in 8/559 bp (1.4% difference, no gap) and differs from A. ovoidea in 9/565 bp (1.6% difference, no gap). A GAPDH nucleotide pairwise comparison showed that A. falcata differs from A. arctoseptata in 13/590 bp (2.2% difference, no gap), differs from A. baoshanensis in 13/578 bp (2.2% difference, no gap) and differs from A. oviodea in 11/570 bp (1.9% difference, no gap). An Alt-a1 nucleotide pairwise comparison showed that A. falcata differs from A. arctoseptata in 11/520 bp (2.1% difference, no gap), differs from A. baoshanensis in 8/474 bp (1.7% difference, no gap) and differs from A. ovoidea in 14/520 bp (2.7% difference, no gap). Thus, the new species A. falcata is established based on morphology and phylogeny.
Alternaria lathyri J.F. Li, Camporesi, Phookamsak & Bhat, sp. nov.
Index Fungorum number: IF 558440; Facesoffungi number: FoF 12658; Fig. 17

Figure 17.
Alternaria lathyri (MFLU 21-0297, holotype). (a) Colonies on dead stem dead aerial stem of Lathyrus sp. (b) Conidiophores and conidia. (c)–(h) Conidiophores bearing conidiogenous cells. (i)–(n) Conidia. (o) Germinated conidium. Scale bars: (a) = 200 µm, (b)–(o) = 20 µm.
Etymology: Named after the host genus 'Lathyrus'.
Holotype: MFLU 21-0297
Saprobic on dead stem of Lathyrus sp. (Fabaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, with dark hyphae. Conidiophores 100–130 × 11–15 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 12 h and germ tubes produced from lateral cells. Colonies growing on PDA, cottony, brown to dark brown, reaching 5 cm in 10 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, brown to dark brown hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, white to brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia formed in vitro within 30 d, borne in chains with at least 2 conidia, light brown to brown, ovoid to obpyriform, 55 × 20 µm (Fig 8b).
Material examined – Italy, Province of Forlì-Cesena, Galeata, Strada San Zeno, on dead aerial stem of Lathyrus sp. (Fabaceae), 11 January 2014, E. Camporesi, IT 1640 (MFLU 21-0297, holotype), ex-type living culture = MFLUCC 21-0140.
Notes – Multi-locus phylogeny demonstrated Alternaria lathyri has a close relationship with A. muriformispora (MFLUCC 22-0073, MFLU 21-0309) and A. pseudoinfectoria (MFLUCC 21-0126, MFLU 21-0311) with 87% ML and 0.99 PP support (Fig. 3). Alternaria lathyri can be distinguished from A. muriformispora in having smaller (55 × 25 µm vs. 83 × 29 µm) darker, and beakless conidia, with less conidial septation. The species also differs from A. pseudoinfectoria in having larger (55 × 25 µm vs. 33 × 19 µm), ovoid to obpyriform, dark grey-brown and beakless conidia, while A. pseudoinfectoria has subglobose to obclavate, or obpyriform, light brown conidia that usually formed long secondary conidiophores[68]. A RPB2 nucleotide pairwise comparison showed that A. lathyri differs from A. muriformispora in 9/570 bp (1.6% differences, no gap), and differs from A. pseudoinfectoria in 12/550 bp (2.2% difference, no gap). In Alt-a1, A. lathyri differs from A. muriformispora in 8/474 bp (1.7% differences, no gap) and differs from A. pseudoinfectoria in 8/480 bp (1.6% difference, no gap). In this study, A. lathyri is described herein as a new species occurring on the same host genus with A. arctoseptata.
Alternaria macilenta J.F. Li, Camporesi, Phookamsak & Bhat, sp. nov.
Index Fungorum number: IF 558441; Facesoffungi number: FoF 12659; Fig. 18
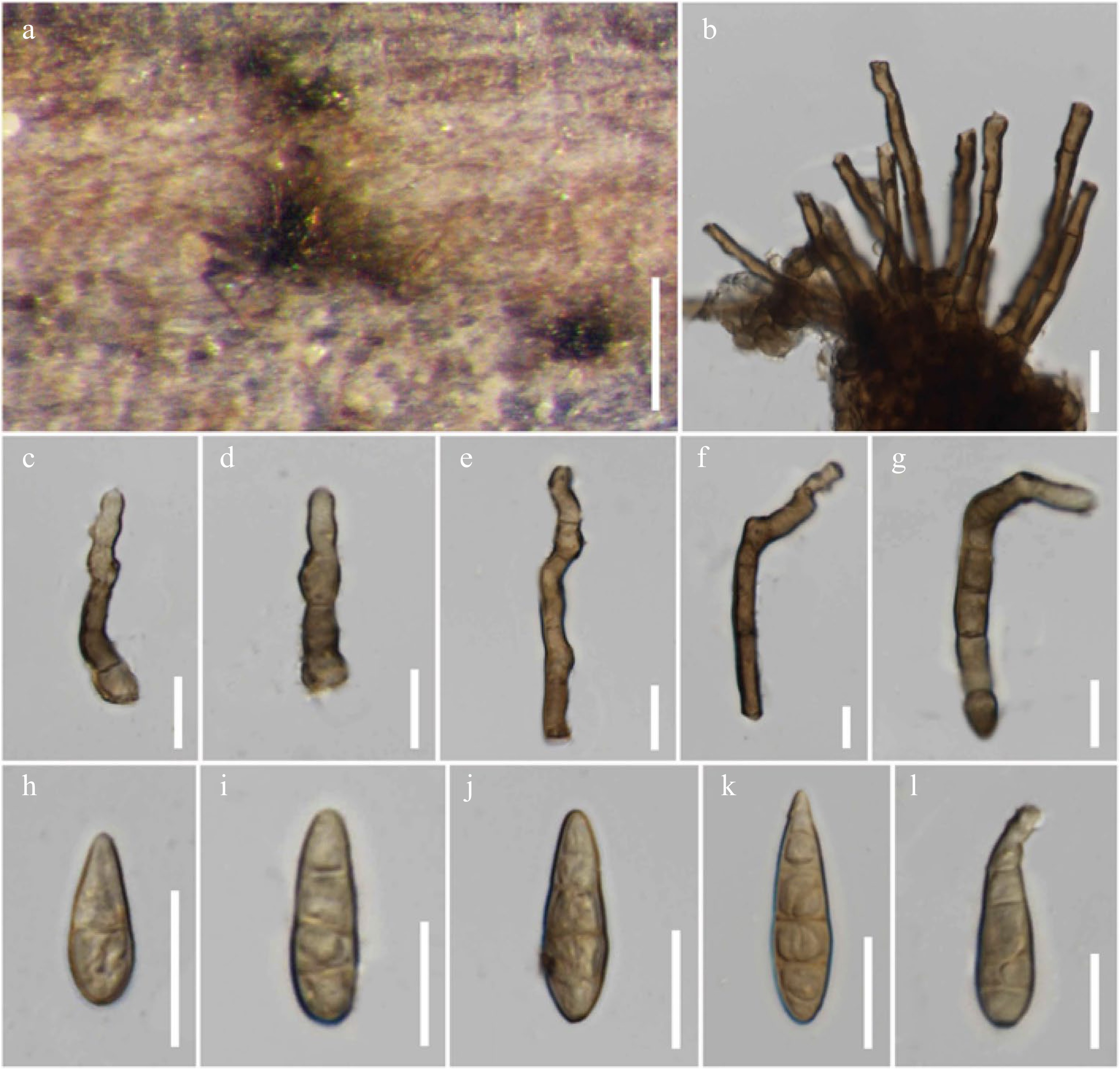
Figure 18.
Alternaria macilenta (MFLU 21-0305, holotype). (a) Colonies on dead stem of Scabiosa sp. (b)–(g) Conidiophores. (h)–(l) Conidia. Scale bars: (a) = 150 µm, (b)–(l) = 20 µm.
Etymology: Referring to its slender-shaped conidia.
Holotype: MFLU 21-0305
Saprobic on dead standing stem of Scabiosa sp. (Caprifoliaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, composed of dark hyphae. Conidiophores 70–100 × 10–12 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from both end cells. Colonies growing on PDA, cottony, brown to dark brown, reaching 5 cm in 10 d at 25 ºC, mycelium superficial, effuse, radially striated, with irregular edge, pale white to white hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, light brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia formed in vitro within 30 d, borne in chains with at least 2 conidia, light brown to brown, subcylindrical to obclavate, 35 × 10 µm (Fig. 8d).
Material examined – Italy, Province of Forlì-Cesena, Dovadola, on dead standing stem of Scabiosa sp. (Caprifoliaceae), 25 August 2015, E. Camporesi, IT 2076 (MFLU 21-0305, holotype), ex-type living culture = MFLUCC 21-0138.
Notes – Multi-locus phylogeny (Fig. 3) demonstrated that Alternaria macilenta clustered with A. oblongoellipsoidea and A. phragmiticola with 80% ML and 0.99 PP support. Alternaria macilenta can be easily distinguished from A. oblongoellipsoidea and A. phragmiticola in having subcylindrical to obclavate, beakless, pale brown to light greyish brown, 0–4-distoseptate conidia, while lacking longitudinal septum and conidiophores aggregated at the base. A RPB2 nucleotide pairwise comparison showed that A. macilenta differs from A. oblongoellipsoidea 10/560 bp (1.8% difference, no gap) and differs from A. phragmiticola in 9/560 bp (1.6% difference, no gap). In Alt-a1, A. macilenta differs from A. oblongoellipsoidea in 10/470 bp (2.1% difference, no gap) and differs from A. phragmiticola in 8/492 bp (1.6% difference, no gap).
Alternaria macroconidia J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558442; Facesoffungi number: FoF 12660; Fig. 19

Figure 19.
Alternaria macroconidia (MFLU 21-0301, holotype). (a) Specimen examined of Spartium junceum (Fabaceae). (b) Colonies on dead aerial branch of Spartium junceum. (c)–(j) Conidiophores bearing conidiogenous cells. (k)–(n) Immature conidia. (o)–(s) Mature conidia. (t) Germinated conidium. Scale bars: (a) = 0.5 cm, (b) = 1000 µm, (c)–(t) = 20 µm.
Etymology: Named after its large conidia (up to 60 µm).
Holotype: MFLU 21-0301
Saprobic on branches of Spartium junceum L. (Fabaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on host substrate, composed of septate, branched, smooth, thin-walled, white to light brown hyphae. Conidiophores 46–80 × 7–12 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 12 h and germ tubes produced from lateral cells. Colonies growing on PDA, cottony, white to light brown, reaching 5 cm in 7 d at 25 ºC, mycelium superficial, effuse, radially striate, with regular edge, white to light brown hyphae. Conidia sporulated on OA media within 15 d, muriform, ovoid to obclavate, with aseptate, short, paler brown, acicular apical rostrum, light brown to brown, 2–4 transverse eusepta, 0–1 longitudinal septum in transverse division, borne in chains, smooth, laterally formed branched or unbranched conidiophores with 1–2 conidiogenous loci. Colonies growing on PCA, pale white to light brown colored, cottony, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, subhyaline hyphae, 2–5 µm diam; conidia not formed in vitro within 60 d.
Material examined – Italy, Province of Forlì-Cesena, Santa Sofia, Collina di Pondo, on dead aerial branches of Spartium junceum (Fabaceae), 6 March 2014, E. Camporesi, IT1756 (MFLU 21-0301, holotype), ex-type living culture = MFLUCC 21-0134.
Notes – Alternaria macroconidia resembles A. phragmiticola in having obclavate conidia, with a short to long beak or rostrate apex when mature. However, A. macroconidia differs from A. phragmiticola in having slightly larger (75.6 × 27.7 µm vs. 70 × 25 µm), olivaceous brown to golden brown or brown conidia, rather monotretic conidiogenous cells and shorter (55 × 8.2 µm vs. 90 × 6.5 µm), aggregated conidiophores at the base. In multi-locus phylogenetic analyses, A. macroconidia formed a separate clade and is sister to A. salicicola (MFLUCC 22-0072, MFLU 21-0320) with 61% ML, 0.95 PP support (Fig. 3). Alternaria macroconidia differs A. salicicola (MFLUCC 22-0072, MFLU 21-0320) in having larger (75.6 × 27.7 µm vs. 45 × 32 µm), olivaceous brown to golden brown or brown conidia, with 3–5 transverse disto- or eusepta, and one longitudinal or oblique or Y-shaped septum in some transverse division. While A. salicicola (MFLUCC 22-0072, MFLU 21-0320) has subglobose to obclavate or obpyriform, light yellowish-brown to light brown conidia, with a longer beak, sectored, with several transverse and longitudinal distosepta. In GAPDH, A. macroconidia differs from A. salicicola in 23/569 bp (4% difference, no gap). In Alt-a1, A. macroconidia differs from A. salicicola in 12/520 bp (2.3% difference, no gap). Based on distinct morphological characteristics and phylogenetic support, A. macroconidia is introduced as a new species in this study.
Alternaria minimispora J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558443; Facesoffungi number: FoF 12661; Fig. 20

Figure 20.
Alternaria minimispora (MFLU 21-0295, holotype). (a) Colonies on rotten peel of Citrullus lanatus. (b)–(e), (h), (i) Conidiophores bearing conidiogenous cells. (f) Conidiogenous cells. (g) Secondary conidiophores arising from conidia. (j)–(q) Conidia. Scale bars: (a) = 300 µm, (b)–(e), (h), (i) = 30 µm, (f), (g), (j)–(q) = 10 µm.
Etymology: Named after its tiny conidia.
Holotype: MFLU 21-0295
Saprobic on rotten peel of Citrullus lanatus (Thumb.) Matsum et Nakai (Cucurbitaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, with dark hyphae. Conidiophores 65–175 × 8–13 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, cottony, white to grey, reaching 5 cm in 10 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, yellow white to grey hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, light brown to brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, subhyaline to light brown hyphae, 2–5 µm diam; conidia formed in vitro within 30 d, borne in chains with at least 2 conidia, light brown, ovoid to obpyriform or obturbinate, 20 × 10 µm (Fig. 8n).
Material examined – Thailand, Chiang Rai Province, Mae Fah Luang University, on rotten peel of Citrullus lanatus (Cucurbitaceae), 13 August 2016, J.F. Li, H-14 (MFLU 21-0295, holotype), ex-type living culture = MFLUCC 21-0127.
Notes – Alternaria minimispora resembles A. breviconidiophora due to small, subglobose to ovoid, dark brown conidia, with swollen knots conidiophores and paler brown conidiogenous cells. However, A. minimispora differs from A. breviconidiophora in having solitary, longer (129 × 10 µm vs. 45 × 9 µm) conidiophores, lacking a stomatic base and conidia that are rather rostrate than A. breviconidiophora. In multi-locus phylogenetic analyses, A. minimispora formed a sister clade with A. rostroconidia (64% ML, 0.99 PP; Fig. 3) and distant from A. breviconidiophora. Alternaria minimispora can be distinguished from A. rostroconidia in having smaller (20 × 9.5 µm vs. 66 × 22 µm), subglobose to ovoid, beakless conidia that conidia are rather short beak in A. rostroconidia[68]. A RPB2 nucleotide pairwise comparison showed that A. minimispora differs from A. breviconidiophora in 40/505 bp (7.9% difference, no gap) and differs from A. rostroconidia in 19/505 bp (3.8% difference, no gap). In GAPDH, A. minimispora differs from A. breviconidiophora in 12/550 bp (2.1% difference, no gap) and differs from A. rostroconidia in 10/545 bp (1.8% difference, no gap). In comparison, the Alt-a1 nucleotide shows that A. minimispora differs from A. breviconidiophora in 14/474 bp (3% difference, no gap) and differs from A. rostroconidia in 8/474 bp (1.7% difference, no gap).
Alternaria oblongoellipsoidea J.F. Li, Camporesi, Phookamsak & Bhat, sp. nov.
Index Fungorum number: IF 558444; Facesoffungi number: FoF 12662; Fig. 21

Figure 21.
Alternaria oblongoellipsoidea (MFLU 21-0310, holotype). (a) Colonies on dead stem of Cichorium intybus. (b)–(h) Conidiophores bearing conidiogenous cells. (i)–(l) Conidia. (m) Germinated conidium. Scale bars: (a) = 300 µm, (b)–(m) = 20 µm.
Etymology: Referring to its oblong to ellipsoidal conidia.
Holotype: MFLU 21-0310
Saprobic on dead stem of Cichorium intybus L. (Asteraceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, composed of septate, branched, smooth, thin-walled, brown to dark brown hyphae. Conidiophores 80–120 × 6.5–11 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, hairy or cottony, brown to dark brown, reaching 5 cm in 7 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, dark brown hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, light brown to brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia formed in vitro within 30 d, born in chains with at least 2 conidia, light brown, obclavate to obpyriform, 50 × 20 µm (Fig. 8c).
Material examined – Italy, Province of Forlì-Cesena, Meldola, on dead aerial stem of Cichorium intybus (Asteraceae), 8 September 2014, E. Camporesi, IT2102 (MFLU 21-0310, holotype), ex-type living culture = MFLUCC 22-0074.
Notes – In multi-locus phylogenetic analyses, Alternaria oblongoellipsoidea is sister to A. phragmiticola with 80% ML, 0.99 PP support and also clustered with A. macilenta. A RPB2 nucleotide pairwise comparison showed that A. oblongoellipsoidea differs from A. phragmiticola in 10/560 bp (1.8% difference, no gap) and also differs from A. phragmiticola in 8/492 bp (1.6% difference, no gap) based on an Alt-a1 nucleotide pairwise comparison. Alternaria oblongoellipsoidea can be distinguished from A. phragmiticola in having smaller (52 × 22 µm vs. 70 × 25 µm) oblong to ellipsoidal conidia, with 4–7 transverse eusepta, and 1–2 longitudinal or oblique or Y-shaped septa in transverse divisions. While A. phragmiticola has obclavate to obpyriform conidia, with longer rostrate beak and less conidial septation than A. oblongoellipsoidea.
Alternaria orobanches J.F. Li, Camporesi, Phookamsak & Jeewon, sp. nov.
Index Fungorum number: IF 558445; Facesoffungi number: FoF 12663; Fig. 22

Figure 22.
Alternaria orobanches (MFLU 21-0303, holotype). (a) Colonies on dead stem of Orobanche sp. (b)–(g) Conidiophores bearing conidiogenous cells. (h) Conidiophores bearing conidiogenous cells with attached conidia. (i)–(q) Conidia. (r) Germinated conidium. Scale bars: (a) = 100 µm, (b)–(r) = 20 µm.
Etymology: Named after its host occurrence on 'genus Orobanche'
Holotype: MFLU 21-0303
Saprobic on dead hanging stem of Orobanche sp. (Orobanchaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the host substrate, with dark hyphae. Conidiophores 50–90 × 15–18 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 12 h and germ tubes produced from lateral cells. Colonies growing on PDA, hairy fluffy, pale white to white, reaching 5 cm in 14 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, white hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, white to light brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia not formed in vitro within 60 d.
Material examined – Italy, Province of Forlì-Cesena, Predappio, Monte Mirabello, on dead hanging stem of Orobanche sp. (Orobanchaceae), 16 July 2014, E. Camporesi, IT1997 (MFLU 21-0303, holotype), ex-type living culture, MFLUCC 21-0137.
Notes – Alternaria orobanches resembles A. arctoseptata in having short, colorless conidiophores arising from a stomatic base. However, A. orobanches differs from A. arctoseptata due to its shorter, wider and less septate conidiophores (75 × 17 µm, 1–3-septate vs. 82 × 9 µm, 2–4-septate). Conidia of A. orobanches are obclavate to ovoid, pale yellowish-brown to pale brown while A. arctoseptata has subglobose to ovoid or pyriform, yellowish-brown to dark brown conidia. A GAPDH nucleotide pairwise comparison showed that A. orobanches differs from A. arctoseptata in 10/490 bp (2% difference, no gap) and also differs from A. arctoseptata in 13/480 bp (2.7% difference, no gap) based on Alt-a1 nucleotide pairwise comparison. Multi-locus phylogeny also supports the distinctiveness of these two species. Alternaria orobanches constitutes an independent lineage basal to other species in sect. Alternaria with high support (100% ML, 1.00 PP; Fig. 3).
Alternaria phragmiticola J.F. Li, Camporesi, Bhat & Jeewon, sp. nov.
Index Fungorum number: IF 558446; Facesoffungi number: FoF 12664; Fig. 23

Figure 23.
Alternaria phragmiticola (MFLU 21-0316, holotype). (a) Colonies on dead stem Phragmites sp. (b)–(g) Conidiophores bearing conidiogenous cells. (h)–(o) Conidia. Scale bars: (a) = 200 µm, (b)–(o) = 20 µm.
Etymology: Named after the host genus 'Phragmites', of which the species was collected.
Holotype: MFLU 21-0316
Saprobic on stem of Phragmites sp. (Poaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the substrate, with dark hyphae. Conidiophores 50–108 × 4–8 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from lateral cells. Colonies growing on PDA, hairy or cottony, brown to dark brown, reaching 5 mm in 20 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, yellow-white to grey hyphae; conidia not formed in vitro within 60 d. Colonies growing on PCA, light brown to brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, light brown hyphae, 2–5 µm diam; conidia formed in vitro within 60 d, borne in chains with at least 2 conidia, brown to dark brown, verruculose, obclavate to obpyriform, 70 × 25 µm (Fig. 8m).
Material examined – Italy, Province of Forlì-Cesena, Meldola, San Colombano, on dead aerial stem of Phragmites sp. (Poaceae), 28 September 2015, E. Camporesi, IT2630 (MFLU 21-0316, holotype), ex-type living culture, MFLUCC 21-0125.
Notes – Multi-locus phylogenetic analyses demonstrated that three strains of Alternaria phragmiticola formed a robust clade (100% ML, 1.00 PP; Fig. 3) and clustered with A. macilenta and A. oblongoellipsoidea. These three species formed a well-resolved clade together with significant support (84% ML, 1.00 PP; Fig. 3). Alternaria phragmiticola can be distinguished from A. macilenta and A. oblongoellipsoidea in having solitary, dark brown, conidiophores and obclavate to obpyriform, pale yellowish-brown to brown, conidia with a rostrate apex, while A. macilenta has brown, aggregated conidiophores, arising from a stomatic base and subcylindrical to obclavate, beakless, pale brown to light greyish brown, distoseptate conidia. Alternaria oblongoellipsoidea has more twisted conidiophores at the apex and oblong to ellipsoidal conidia, with more conidial eusepta.
Alternaria salicicola J.F. Li, Bulgakov & Phookamsak, sp. nov.
Index Fungorum number: IF 558447; Facesoffungi number: FoF 12665; Fig. 24

Figure 24.
Alternaria salicicola (MFLU 21-0320, holotype). (a) Colonies on dead twig of Salix alba. (b)–(e) Conidiophores bearing conidiogenous cells. (f)–(p) Conidia. (q) Germinated conidium. Scale bars: (a) = 150 µm, (b)–(q) = 20 µm.
Etymology: Named after the host genus Salix, of which the species was found.
Holotype: MFLU 21-0320
Saprobic on dead twig of Salix alba L. (Salicaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on host substrate, with dark hyphae. Conidiophores 110–150 × 10–15 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from lateral cells. Colonies growing on PDA, hairy or cottony, brown to dark brown, reaching 5 cm in 10 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, light brown to brown hyphae; conidia not sporulated in vitro within 60 d. Colonies growing on PCA, white to light brown colored, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, hyaline hyphae, 2–5 µm diam; conidia formed in vitro within 60 d, borne in chains with at least 2 conidia, brown to dark brown, with secondary conidiophores, acicular to doliiform, 45 × 30 µm (Fig. 8i).
Material examined – Russia, Rostov Region, Krasnosulinsky District, trees near Kudryuchya River, on dead twig of Salix alba (Salicaceae), 18 June 2015, T.S. Bulgakov, T-504 (MFLU 21-0320, holotype), ex-type living culture = MFLUCC 22-0072.
Notes – Alternaria salicicola is the only species collected from Russia in this study. In multi-locus phylogenetic analyses, two strains of A. salicicola formed a robust clade (100% ML, 1.00 PP; Fig. 3) sister to A. macroconidia.
Section Infectoriae Woudenb. & Crous, Study in Mycology 75: 194 (2013)
Type species – Alternaria infectoria E.G. Simmons.
Notes – Sect. Infectoriae is one of the largest and most complicated sections in Alternaria, containing approximately 50 species[11,70,77,79]. Species in this section often form white or nearly white, floccose colonies on nutrient-rich media and sporulated several conidial chains on low sugar media as extensive three-dimensional branching patterns, and the conidia usually produce long secondary conidiophores[47,162]. Although, members of sect. Infectoriae are commonly known as saprobes, many species such as A. infectoria and A. triticina have been reported as important pathogens on various plant hosts as well as human infections[14,15,47,79,81,163]. Additionally, species in this section produce secondary metabolites such as novae-zelandins and infectopyrone, rather than mycotoxins, which are unique to this section[47,163]. The latest updated account of species number in sect. Infectoriae was carried out by Marin-Felix et al.[70], who introduced six new species mostly collected from herbivore dung and plant litter and Iturrieta‐González et al.[79] who introduced three novel species causing human cutaneous infections. In this study, we introduce two other novel saprobic species collected from dead plants in Italy based on typical morphology and multi-locus phylogeny (Fig. 5).
Alternaria arundinis J.F. Li, Camporesi, Bhat & Jeewon, sp. nov.
Index Fungorum number: IF 558448; Facesoffungi number: FoF 12666; Fig. 25

Figure 25.
Alternaria arundinis (MFLU 21-0313, holotype). (a) Colonies on dead stem of Arundo donax. (b)–(f) Conidiophores bearing conidiogenous cells. (g)–(i) Conidia formed secondary conidiophores. (j)–(n) Conidia. Scale bars: (a) = 150 µm, (b)–(n) = 20 µm.
Etymology: Named after the host genus Arundo, of which the species was collected.
Holotype: MFLU 21-0313
Saprobic on dead leaf of Arundo donax L. (Poaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on the host substrate, with dark hyphae. Conidiophores 50–85 × 5–8 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, cottony, white to dark grey, reaching 5 cm in 10 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, white to grey hyphae; conidia not sporulated in vitro within 60 d. Colonies growing on PCA, light brown to brown, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, brown hyphae, 2–5 µm diam; conidia formed in vitro within 60 d, borne in chains with at least 2 conidia, brown to dark brown, with long secondary conidiophores, ellipsoidal to ovoid, or obclavate, 40 × 20 µm (Fig. 8j).
Material examined – Italy, Province of Forlì-Cesena, Predappio, Rocca delle Caminate, on dead aerial leaf of Arundo donax (Poaceae), 17 November 2014, E. Camporesi, IT2250 (MFLU 21-0313, holotype), ex-type living culture, MFLUCC 21-0128.
Notes – Alternaria arundinis is typical of sect. Infectoriae in forming secondary conidiophores, with white to dark grey colonies on PDA. Multi-locus phylogeny demonstrated that A. arundinis formed an independent lineage sister to A. incomplexa with significant support (89% ML, 1.00 PP; Fig. 5) and clustered with Alternaria sp. (H1-4) and A. anthropophila (CBS 541.94). Alternaria arundinis resembles A. incomplexa in having ellipsoidal to ovoid, or obclavate conidia, with 3(–5) transverse eusepta, and 1–2 longitudinal septa. However, A. arundinis can be distinguished from A. incomplexa in having shorter conidiophores (70 × 7 µm vs. 100 × 4 µm) and slightly larger conidia (38 × 20 µm vs. 18–25 × 5–8 µm), which formed secondary conidiophores with several conidiogenous loci, while the conidia can raise up to 30–40 × 8–13 µm, with 5–8 transverse septa and 0–4 longitudinal septa in A. incomplexa[164]. Alternaria arundinis differs from A. anthropophila in having shorter conidiophores (50–85 × 5–8 µm vs. 26–120 × 4–7 µm) with laterally and apically geniculate conidiogenous loci, while conidiophores are apically geniculate in A. anthropophila, the conidia of A. arundinis are shorter and wider (20–45 × 15–25 µm vs. 11–63 × 6–11 µm), and more longitudinal or oblique euseptate (1–2 vs. 0–1)[79]. An ITS nucleotide pairwise comparison showed that A. arundinis differs from A. incomplexa in 8/546 bp (1.5% difference, no gap) and differs from A. anthropophila in 10/546 bp (1.8% difference, no gap). In GAPDH, A. arundinis differs from A. incomplexa in 9/521 bp (1.7% difference, no gap) and differs from A. anthropophila in 14/521 bp (2.7% difference, no gap). In ATPase, A. arundinis differs from A. incomplexa in 16/1000 bp (1.6% difference, no gap) and differs from A. anthropophila in 21/1000 bp (2.1% difference, no gap).
Alternaria nodulariconidiophora J.F. Li, Camporesi, Bhat & Jeewon, sp. nov.
Index Fungorum number: IF 558449; Facesoffungi number: FoF 12667; Figs 26 & 27

Figure 26.
Alternaria nodulariconidiophora (MFLU 21-0315, holotype). (a) Colonies on dead stem of Heracleum sphondylium. (b)–(e) Geniculate conidiophores bearing conidiogenous cells. (f)–(m) Conidia. (n) Germinated conidium. Scale bars: (a) = 100 µm, (b)–(n) = 20 µm.
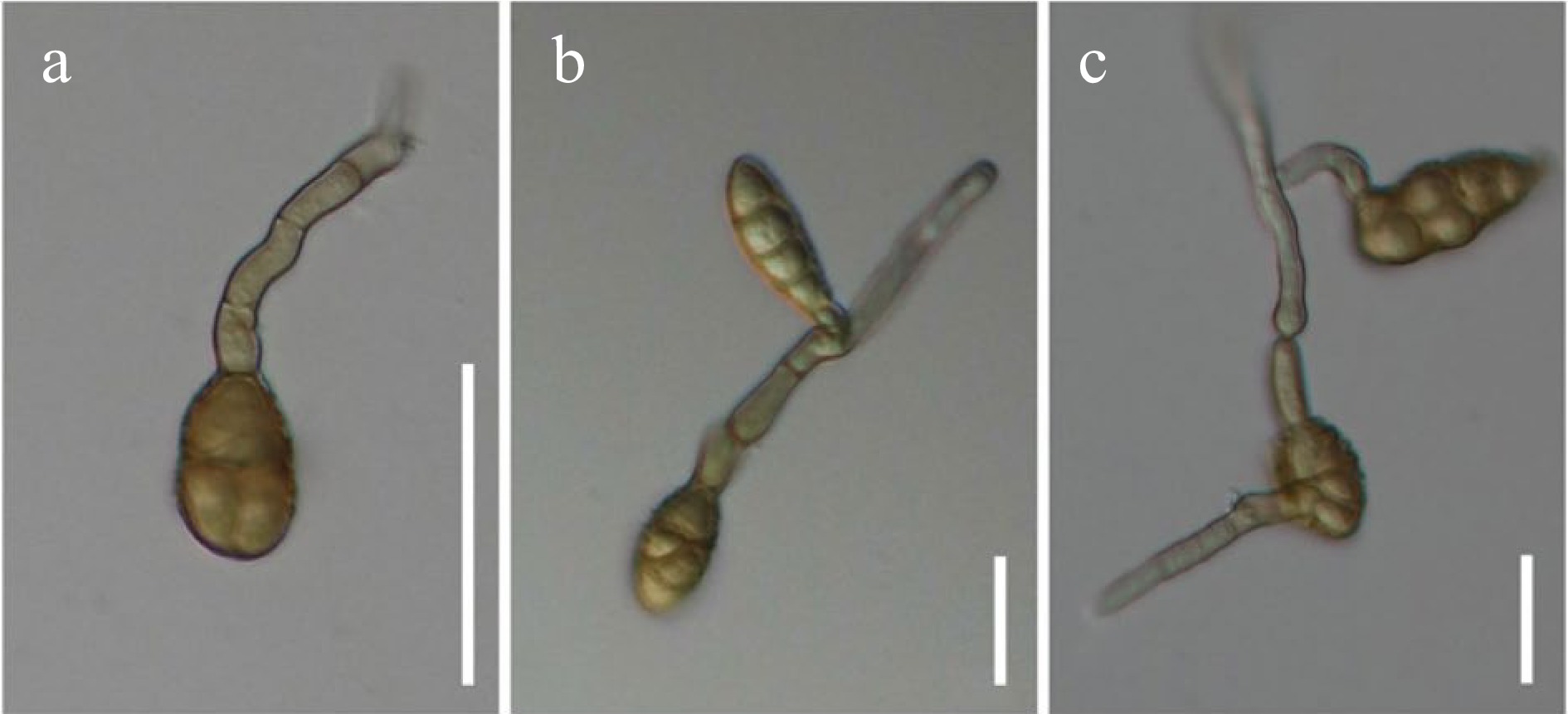
Figure 27.
Alternaria nodulariconidiophora (MFLUCC 21-0131) sporulated on PDA. (a) Conidium formed a geniculate secondary conidiophore. (b), (c) Secondary conidiophores bearing conidia with three-dimensional branching patterns. Scale bars: (a)–(c) = 30 µm.
Etymology: Referring to the geniculate conidiophores.
Holotype: MFLU 21-0315
Saprobic on stem of Heracleum sphondylium L. (Apiaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on host substrate, composed of brown to dark brown hyphae. Conidiophores (111–)119.5–183 × (12–)15–23 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, hairy or cottony, brown to dark brown, reaching 5 cm in 5 d at 25 ºC, mycelium superficial, effuse, with irregular edge, brown to dark brown hyphae. Conidia sporulated in PDA after 60 d (Fig. 27), phragmosporous to muriform, subglobose to ellipsoidal, or fusiform, yellowish brown, 1–4 transverse eusepta, with 1–2 longitudinal or oblique or Y-shaped septa in some transverse divisions, constricted at the septa, verrucose, formed apically or laterally geniculated secondary conidiophores, with 1–2 apical or lateral conidiogenous loci, with long secondary conidiophores, obturbinate to obpyriform, 50 × 30 µm.
Material examined – Italy, Province of Forlì-Cesena, Meldola, Piandispino, on dead aerial stem of Heracleum sphondylium (Apiaceae), 10 October 2017, E. Camporesi, IT2625A (MFLU 21-0315, holotype), ex-type living culture, MFLUCC 21-0131.
Notes – In multi-locus phylogeny, Alternaria nodularconidiophora formed a separate branch and clustered with Alternaria sp. (JS8-5) and A. humuli (CBS 119404) with significant support (87% ML, 1.00 PP; Fig. 5). Alternaria nodularconidiophora is characterized by subglobose to ovoid, or obturbinate to obpyriform conidia, with a rostrate apex, geniculate conidiophores with swollen knots, arising from a stomatic base, and subhyaline to pale brown conidiogenous cells, with 1–2 conidiogenous loci on bulge. While A. humuli (CBS 119404) is characterized by subhyaline to arachnoid to loosely, woolly colonies, simple to moderately, branched primary conidiophores, with 1–3 geniculate conidiogenous loci, elliptical or ovoid, 5–8 transverse septa, with or without a longitudinal septum[165]. A ITS nucleotide pairwise comparison showed that A. nodularconidiophora differs from A. humuli in 8/545 bp (1.5% difference, no gap). In GAPDH, the species differs from A. humuli in 8/521 bp (1.5% diference, no gap) and also differs from A. humuli in 11/900 bp (1.5% diference, no gap) in ATPase nucleotide pairwise comparison.
Section Porri D.P. Lawr., Gannibal, Peever & B.M. Pryor, Mycologia 105: 541. 2013.
Type species – Alternaria porri (Ellis) Cif.
Notes – The section Porri is characterized by medium to large conidia with a simple or branched, filamentous long beak, which is the second largest section in Alternaria[11,22]. This section includes many important phytopathogens, such as A. bataticola, A. porri, A. solani, and A. tomatophila. In our survey, a new species A. brevirostra is described on dead grass from Italy based on both morphological and phylogenetic evidence (Fig. 6 ).
Alternaria brevirostra J.F. Li, Camporesi, Bhat & Jeewon, sp. nov.
Index Fungorum number: IF 558450; Facesoffungi number: FoF 12668; Fig. 28

Figure 28.
Alternaria brevirostra (MFLU 21-0312, holotype). (a) Colonies on dead stem of Plantago sp. (b)–(f) Conidiophores bearing conidiogenous cells. (g)–(k) Conidia. (l) Germinated conidium. Scale bars: (a) = 100 µm, (b)–(l) = 20 µm.
Etymology: Referring to the short rostrate conidia in sect. Porri.
Holotype: MFLU 21-0312
Saprobic on stems of Erysimum sp. (Brassicaceae) and Plantago sp. (Plantagineae). Sexual morph Undetermined. Asexual morph Mycelium superficial on host substrate, composed of septate, branched, brown to dark brown, smooth, thin-walled hyphae. Conidiophores 55–72 × 10–15 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, hairy or cottony, brown to dark brown, reaching 5 mm in 20 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, brown to dark brown hyphae; conidia not sporulated in vitro within 60 d. Colonies growing on PCA, pale white to white, cottony, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, hyaline hyphae, 2–5 µm diam; conidia not formed in vitro within 60 d.
Material examined – Italy, Province of Forlì-Cesena, Tontola di Predappio, on dead aerial stem of Plantago sp. (Plantagineae), 20 October 2014, E. Camporesi, IT2195 (MFLU 21-0312, holotype), ex-type living culture, MFLUCC 21-0129; ibid., Forlì, Via Nenni, on dead aerial stem of Erysimum sp. (Brassicaceae), 28 July 2014, E. Camporesi, IT2028 (MFLU 21-0304), living culture = MFLUCC 21-0130.
Notes – Alternaria brevirostra corresponds to species in sect. Porri via being borne in chains, long and filamentous rostrate, multiple transverse and longitudinal septa conidia[11]. In phylogenetic analyses, A. brevirostra formed a sister clade with A. rostellata (CBS 117366) with significant support in BI analysis (0.98 PP; Fig. 6) and clustered with A. nitrimali (CBS 109163), A. pipionipisi (CBS 116115), A. crassa (CBS 110.38) and A. macrospora (CBS 117128). Alternaria brevirostra can be distinguished from A. rostellata in having shorter conidiophores (55–72 × 10–15 µm vs. 150–200 × 6.5 µm) and conidiophores were aggregated at the base with polytretic, integrated, terminal, determinate or percurrent conidiogenous cells, whereas A. rostellata has simple or branched conidiophores with a single dark conidium terminating each conidiophore[164]. Conidia of A. rostellata were slightly shorter (50–80 × 20–30 µm) and solitary, ellipsoidal to broadly ovoid, dilute tawny brown to dark brown in age, smooth to moderately punctate-rough, with 7–9 transverse septa and 1–3 longitudinal septa, with subhyaline, narrow beak at the apex, occasionally a filamentous beak enlarged terminally into a second conidiophore[164]. While conidia of A. brevirostra were subcylindrical to obclavate, light yellowish-brown to brown, with 5–7 transverse distosepta, with 1–2 longitudinal or oblique septa in some transverse divisions, smooth, thin-walled, with slightly long, narrow, colorless, acicular, aseptate rostrum.
An ITS nucleotide pairwise comparison showed that Alternaria brevirostra differed from A. rostellata by 12/530 bp (2.2% difference, no gap), differed from A. nitrimali (CBS 109163) by 10/530 bp (1.9% difference, no gap), differed from A. pipionipisi (CBS 116115) by 10/530 bp (1.9% difference, no gap), differed from A. crassa (CBS 110.38) by 11/530 bp (2% difference, no gap), and differed from A. macrospora by 10/530 bp (1.9% difference, no gap). In GAPDH, the species differed from A. rostellata by 8/570 bp (1.4% difference, no gap), differed from A. nitrimali (CBS 109163) by 8/577 bp (1.4% difference, no gap), differed from A. pipionipisi (CBS 116115) by 9/577 bp (1.6% difference, no gap), differed from A. crassa (CBS 110.38) by 10/577 bp (1.7% difference, no gap), and differed from A. macrospora by 11/577 bp (1.9% difference, no gap). In TEF1-α, A. brevirostra differed from A. rostellata by 11/337 bp (3.2% difference, no gap), differed from A. nitrimali (CBS 109163) by 9/337 bp (2.7% difference, no gap), differed from A. pipionipisi (CBS 116115) by 8/337 bp (2.4% difference, no gap), differed from A. crassa (CBS 110.38) by 12/337 bp (3.6% difference, no gap), and differed from A. macrospora by 10/337 bp (3% difference, no gap). In RPB2 nucleotide pairwise comparison, A. brevirostra differed from A. rostellata by 14/539 bp (2.6% difference, no gap), differed from A. nitrimali (CBS 109163) by 14/539 bp (2.6% difference, no gap), differed from A. pipionipisi (CBS 116115) by 13/539 bp (2.4% difference, no gap), differed from A. crassa (CBS 110.38) by 16/539 bp (3% difference, no gap), and differed from A. macrospora (CBS 117228) by 11/539 bp (2% difference, no gap).
Section Radicina D.P. Lawr., Gannibal, Peever & B.M. Pryor, Mycologia 105: 541. 2013.
Type species – Alternaria radicina Meier, Drechsler & E.D. Eddy
Notes – Section Radicina is a small section in Alternaria comprising only eight phylogenetic species[4,70]. Most species in this section were reported as pathogens of Apiaceae[4,15,70]. In this study, we introduced a novel species, A. phytolaccae as a saprobe on Phytolacca sp. (Phytolaccaceae) which is reported from a different host family for the first time.
Alternaria phytolaccae J.F. Li, Camporesi, Bhat & Jeewon, sp. nov.
Index Fungorum number: IF 558451; Facesoffungi number: FoF 12669; Fig. 29

Figure 29.
Alternaria phytolaccae (MFLUCC 21-0314, holotype). (a) Colonies on dead standing stem of Phytolacca sp. (b)–(e) Conidiophores bearing conidiogenous cells, with 2–3 conidiogenous loci on side of cell. (f), (g) Immature conidia. (h)–(k) Conidia. Scale bars: (a) = 150 µm, (b)–(k) = 20 µm.
Etymology: Name after the host genus Phytolacca.
Holotype: MFLU 21-0314
Saprobic on dead standing stem of Phytolacca sp. (Phytolaccaceae). Sexual morph Undetermined. Asexual morph Mycelium superficial on host substrate, with dark hyphae. Conidiophores 160–200 × 8–11 µm (
$ \bar{x}$ $ \bar{x}$ $ \bar{x}$ Culture characteristics – Conidia germinating on PDA within 14 h and germ tubes produced from all cells. Colonies growing on PDA, cottony, brown to dark brown, reaching 5 cm in 7 d at 25 ºC, mycelium superficial, effuse, radially striate, with irregular edge, grey to dark grey hyphae; conidia not sporulated in vitro within 60 d. Colonies growing on PCA, light brown, hairy, fluffy, reaching 5 cm within 7 d at 25 ºC, mycelium superficial, effuse, partly immersed on the media, radially striate, with irregular edge, composed of septate, branched, smooth, thin-walled, hyaline hyphae, 2–5 µm diam; conidia formed in vitro within 60 days, borne in chains with at least 1–2 conidia, light brown, beakless, obturbinate to obpyriform, 37 × 20 µm (Fig. 8k).
Material examined – Italy, Province of Forlì-Cesena, Forl', San Lorenzo in Noceto, on dead standing stem of Phytolacca sp. (Phytolaccaceae), 4 December 2014, E. Camporesi, IT2279 (MFLU 21-0314, holotype), ex-type living culture, MFLUCC 21-0135.
Notes – Alternaria phytolaccae morphologically corresponds with sect. Radicina in having branched, clumped, geniculate, sympodial proliferations conidiophores with several conidiogenous loci at the beakless apex. In multi-locus phylogenetic analyses (Fig. 7), A. phytolaccae formed a separate branch and clustered with A. petroselini, A. selini and A. vulgaris in section Radicina with 60% ML and 0.97 PP support. An ITS nucleotide pairwise comparison showed that A. phytolaccae differs from A. petroselini in 8/520 bp (1.5% difference, no gap), differs from A. selini in 8/520 bp (1.5% differencee, no gap) and differs from A. vulgaris 9/520 bp (1.7% difference, no gap). In GAPDH, A. phytolaccae differs from A. selini in 9/567 bp (1.6% difference, no gap) and differs from A. vulgaris in 9/567 bp (1.6% difference, no gap).
-
Alternaria species have been reported on a wide range of monocotyledonous and dicotyledonous plants worldwide[11−15,18,19,22,38,70]. In this survey carried out in China, Italy, Russia and Thailand, 65 Alternaria samples were recovered from different plant species. Morphological examinations of specimens revealed that there were 18 species that could not be ascribed to any known species within the different Alternaria sections, viz. sects. Alternaria, Infectoriae, Porri and Radicina. Given that there were some noticeable differences in their morphs and with support from phylogeny, new species are established herein following the recommendations as outlined by Jeewon & Hyde[134].
Our study also reveals other taxonomic anomalies. We note that A. oblongoellipsoidea (MFLUCC 22-0074 and MFLU 21-0310) and A. ellipsoidialis (MFLUCC 21-0132, MFLU 21-0307A and MFLU 21-0307B) share meander conidiophores apex with confidential hollow conidiogenous loci, which occur rarely in sect. Alternaria. However, these two species can be differentiated based on their morphs: A. oblongoellipsoidea differs from A. ellipsoidialis in having rather common and short conidiophores (96 × 9.5 µm vs. 145 × 6.5 µm), with rather polytretic, swollen, doliiform conidiogenous cells. Our multi-locus phylogenetic analyses (Fig. 3) also support their distinction and hence A. oblongoellipsoidea and A. ellipsoidialis should be considered as different species in sect. Alternaria.
Interestingly, Alternaria arctoseptata and A. lathyri were isolated from same host genus Lathyrus (Fabaceae). However, A. lathyri morphologically differs from A. arctoseptata in having darker brown, solitary, astomatic base conidiophores with less swollen knots, and dark brown conidia. Phylogenetic analyses with a combined seven gene loci (ITS, LSU, SSU, TEF1-α, RPB2, GAPDH and Alt-a1) positions A. lathyri in a subclade distinct from A. arctoseptata. Furthermore, we also note that A. orobanches (MFLUCC 21-0137 and MFLU 21-0303) formed a single lineage basal to sect. Alternaria. However, A. orobanches shares similar morphological characteristics to others in sect. Alternaria (e.g., phragmosporous to muriform small-spored conidia, conidiophores with monotretic or polytretic conidiogenous loci at apex), and hence A. orobanches should be considered in sect. Alternaria based on morphology and phylogeny.
This study also reveals new host records for Alternaria doliconidium and diverse hosts species for A. alternata. While A. doliconidium has been collected from Rosa sp. in Italy[66], we discovered A. doliconidium from Reseda sp. (Resedaceae) in China, which is the first record of both host species and location for this species. Another important and novel finding herein that we observed and describe for the first time, is the cultural characteristics of A. doliconidium which successfully sporulated on OA medium within 30 d (with conidia borne in chains, oblong to subglobose, verruculose to verrucose with short, branched or unbranched, aseptate apical beak, apical or lateral secondary conidiophores were formed). Moreover, in this study, 45 new collections of A. alternata were isolated from diverse plant hosts (Table 5) in China, Italy and Thailand. These 45 new collections were compared with other A. alternata strains (mainly from CBS-KNAW Fungal Biodiversity Centre, Utrecht, The Netherlands) and our phylogenetic analyses, revealed our A. alternata complex could be phylogenetically arranged in several internal subclades (Fig. 4). For example, strains HWP-01, IT2144, IT2145, IT3598, and IT3651 formed the internal subclade A with strains CBS 965.95 and CBS 966.95 (pathogen and saprobe); saprobic strains H-71, IT2143, and IT3556 formed the internal subclade with strain CBS 130254 (isolated from human sputum in India), and CBS 911.97 (isolated from Artemisia brevifolia (Asteraceae) in India) and clustered with CBS 130262, CBS 130265 (isolated from human sputum in India) and CBS 121492 (isolated from Cucumis melo (Cucurbitaceae) in China) (pathogen and saprobe). These results also indicate that A. alternata and A. doliconidium occur on a wide range of hosts as either pathogens or saprobes.
Our phylogeny also reveals higher species diversity from one particular host. For example, A. alternata have been recovered from diverse plant hosts (Fig. 4)[67] while A. doliconidium colonizes Rosa sp. and Reseda sp.[66]. It is also worth pointing out that our study here also reveals an unexpected higher diversity of Alternaria species, with over 30% (20 out of 65 specimens) from diverse host plants which are new to science and more than one species are associated with one host. This finding has important implications on nomenclature of Alternaria species and it would be erroneous to name species based on host without any additional information, especially for speciose genera, a phenomenon already reported for Pestalotiopsis-like species[166].
Phylogenetic concepts of Alternaria
-
DNA sequence data is very important in Alternaria taxonomy. Characters of conidia in some sections are not informative, as conidia are mostly dictyosporous in sects. Alternaria and Japonicae; some are mostly phragmosporous, such as in sects. Alternantherae and Nimbya. Species in sects. Alternantherae, Dianthicola and Porri have long apical narrow beaks or secondary conidiophores and these characteristics are absent in sects. Chalastospora, Gypsophilae and Ulocladium, while dictyospores and phragmospores can be found in sects. Infectoriae and Phragmosporae[18,19,38]. In our phylogenetic analyses (Fig. 3), the topology of the phylogenetic tree for the Alternaria complex corresponds with previous ones. However, many sections that even share common morphs are usually phylogenetically distinct based on DNA sequence analyses[15,25,26]. In addition, it is also noted that single gene analyses show that the ITS, RPB2 and Alt-a1 genes could resolve the taxonomy of most Alternaria sections, while TEF1-α and GAPDH genes could be informative at the species level but not for section levels.
It is interesting to note that RPB2 regions do provide reliable nucleotide differences for species comparison. However, this region should be analyzed and used properly before any taxonomic assumption is made. In our single gene phylogeny, it has been noted that it is not informative to decipher inter and intra-species relationships. It can be observed that the Alternaria alternata complex in section Alternaria was separated into at least five subclades. Species in the sects. Porri and Euphorbiicola are also dispersed in one subclade while taxa in the sects. Embellisa and Pseudoalternaria were also unresolved in their positions. We therefore recommend precautions when using this region alone to clarify species relationships.
Although Woudenberg et al.[12] assigned 35 morpho-species as synonyms of Alternaria alternata, their affinities are still unclear due to inconsistencies, lack of morphological details and a comparison of single nucleotide polymorphisms. However, further studies based on combined multi-locus phylogeny showed that recent A. alternata species may not constitute a monophyletic group in DNA sequence-based phylogenies[67,167] (present study). We compared our recent collections based on morphology and phylogeny. Interestingly, our phylogenetic analyses show that the phylogenetic strains of A. alternata can be separated to a minimum five different clades (Fig. 4) while the novel taxa from various host species are both morphologically and phylogenetically distinct from A. alternata complex and other species in Alternaria sect. Alternaria (Fig. 3).
The plasma membrane ATPase and cmdA loci are suggested to be the most informative markers for delimitation of Alternaria species in sect. Infectoriae by Lawrence et al.[14,15], while the Alt-a1 locus is not reliable to amplify some species within this section[11,14,70]. However, the cmdA locus has seldom been used for the phylogeny of most Alternaria sections and species[70]. The ITS barcode is also considered a good phylogenetic marker to define the Alternaria sections. However, it has limited discriminatory power to distinguish species[11,12,14,15,55,70].
In our study, two new taxa are accommodated in Alternaria sect. Infectoriae, based on phylogenetic analyses of a combined ITS, GAPDH and ATPase nucleotide sequences. Our new taxa A. arundinis (MFLUCC 21-0128, MFLU 21-0313A and MFLU 21-0313B) and A. nodularconidiophora (MFLUCC 21-0131 and MFLU 21-0315) have oblong-ellipsoidal or obclavate conidia can apically or laterally formed long, geniculate, multi-locus secondary conidiophores. These morphological characters typically tally with those in sect. Infectoriae. However, A. nodularconidiophora was observed as developing less secondary conidiophores on the natural substrate, but secondary conidiophores were heavily produced in culture incubated on PDA media.
Woudenberg et al.[22] indicated that ITS, Alt-a1, GAPDH, RPB2 and TEF1-α are the effective phylogenetic markers to determine relationships and species delineation for Alternaria sect. Porri. However, many strains of species in this section could not be resolved in the present study, concurring with previous studies[11,14,22]. In our study, more than 40 phylogenetic identified species in sect. Porri with one new taxon were confirmed based on phylogenetic analyses of combined ITS, Alt-a1, GAPDH, RPB2 and TEF1-α. Our new taxon A. brevirostra (MFLUCC 21-0129 and MFLUCC 21-0130) produces a relatively short filamentous beak on conidia and this warrants further verification as to whether A. brevirostra could grow longer apical beak at a mature stage or in different cultured media and also investigate the formation of short conidial filamentous beak occur on species in sect. Porri.
Species in Alternaria sect. Racidina are less phylogenetically resolved and share some similar morphological characters, with respect to conidiophores, sporulation, conidial shape and others. The recent phylogenetic studies of this section were reported by He et al.[4], Marin-Felix et al.[70] and Tao et al.[168] where they described novel species based on the multi-locus phylogenetic analyses of ITS, GAPDH, RPB2 and TEF1-α nucleotide sequences. In our survey, based on phylogenetic analyses of a combined ITS, GAPDH, RPB2 and TEF1-α nucleotide sequences, a new species A. phytolaccae (MFLUCC 21-0135 and MFLU 21-0314) is justified and correspond to the morphological and phylogenetic features in sect. Radicina in Woudenberg et al.[11] and Lawrence et al.[15]. There is a lack of taxonomic data for this section and further investigations need to be explored on fresh samples.
On the other hand, our analyses clearly show that species are phylogenetically diverse in Alternaria complex (Figs 3 & 4). The studies of Alternaria from various samples used in the present show that the genus Alternaria is speciose, which is in agreement with previous studies[4,7,11−15,18,19,22,25,26,28,40,55,66,67,70,78,108,113,165,168,169]. Additional novel species/sections are expected if more hosts/habitats are explored. However, there is a need to standardize the taxonomy at different levels. At lower taxonomic levels (intraspecies), one can compare the morph and nucleotide differences with one or two morphologically similar taxa, or phylogenetically close species. Chemotaxonomy can be recommended and can undoubtedly be useful, however, even though the discipline is still in its infancy, it could be explored for such a genus.
Application of molecular dating to solve taxonomy of Alternaria
-
Divergence time estimation of fungi at higher rank has been explored based on multi-locus analyses mainly with combined LSU, SSU and TEF1-α regions[128−130,132,170]. However, to resolve divergence time estimation of Alternaria with higher rank fungal strains, we combined ITS and RPB2 regions into our phylogenetic and divergence analyses (Figs 1& 2), which are informative markers for Alternaria species based on previous studies. The divergence time estimation in our study shows that Alternaria diverged approximately at 62 (42–85) Mya, while the crown age of Alternaria is around 53 (36–72) Mya as well as the divergence time of sect. Crivellia in Late Paleocene to early Eocene. Although this divergence time estimate corresponds to those reported by Kalgutkar & Sigler[123] where a fossil species Piriurella alternariata was obtained from late Palaeocene or early Eocene (56±5 Mya). However, affinity with Alternaria could be properly documented due to a lack of morphs and DNA sequence. On the other hand, Alternaria sections diverged early, such as species in sects. Crivellia, Phragmosporae, Ulocladium and Undifilum are morphologically beakless, round, mostly with transverse eusepta, rarely constricted at the septa, while the type section, sect. Alternaria with mostly distinct characters of Alternaria occurred with a late divergence time as 13 (6–20) Mya[11,58]. Moreover, the sect. Phragmosporae diverged with other sections in age as 43 (28–58) Mya in Eocene, and crown age of this section is around 28 Mya which is in an early crown age of Alternaria. Alternaria species in sections diverged at an early age have mostly beakless conidia with transverse septate and rarely constricted, less or 0–1 longitudinal or oblique septum in some transverse divisions and these can be considered to be the primitive structures of ancient Alternaria species.
Both phylogenetic and evolutionary estimate analyses based on multigene data show that some Alternaria sections bear close relationships and share the same divergence time. Secttion Porri and sect. Euphorbiicola diverged from a clade with a divergence time at 10 (5.6–17.4) Mya. Species in sect. Euphorbiicola are characterized with conidia with beak not distinct from the spore body, and they differ from the characters in sect. Porri. Alternaria cumini in sect. Eureka formed a separate subclade from sect. Eureka in both phylogenetic and divergence analyses (Figs 1 & 2). Secttion Eureka and sect. Embellisioides share a divergence time as 24 (14–35) Mya and these two sections and A. cumini share common morphological structures in having ovoid to subcylindrical, straight to inequilateral, transseptate and less longitudinal septa[11,38] and possibly these can be expected to be one section. Alternaria thalictrigena, sect. Panax, and sect. Teretispora share a divergence time as 27 (17–40) Mya and species in this two sections and A. thalictrigena share the same characteristics in having paler and lanky conidia, and results support their establishment as a section. Strains in sect. Pseudoalternaria in our divergence analyses lack informative DNA sequences from databases and hence we use only LSU gene of the type strain A. arrhenatheri (CBS 133068) [ITS, SSU, TEF1-α and RPB2 genes are not available] and hence the divergence results of sect. Pseudoalternaria may not be as reliable. Species in sect. Crivellia are characterized by cylindrical, straight to curved to inequilateral, with transverse eusepta, rarely constricted at septa[11], and they are morphologically distinct from species in other Alternaria sections, while sect. Crivellia is not well-resolved in ML and BI analyses (Fig. 3) despite the fact that they constitute distinct lineages from other Alternaria sections in divergence time estimation analyses.
We note that the age for the crown of most sections (24 out of 29, 85.7%) in Alternaria are between 0.1 to 20 Mya, while the genus Alternaria (53 Mya), sect. Nimbya (24 Mya) and sect. Phragmosporae (28 Mya) evolved in the range for family (20–100 Mya, crown age) as proposed by Liu et al.[129]. However, the branch length between stem ancestor and crown clades could be affected by species richness of the group, net diversification rate, timescale and model setup[129,170]. Our study only explored divergence time of Alternaria and each section (not familial relationships) but given that divergence time (stem age) of most sections in Alternaria are between 10 and 50 Mya, we can argue that these time estimates can be the supplementary evidence for acceptance of sections in Alternaria.
The authors are grateful to Yunnan Provincial Science and Technology Department (Grant no. 202003AD150004) and the Mushroom Research Foundation, Chiang Mai, Thailand for supporting this research. We also acknowledge that Biology Experimental Center, Germplasm Bank of Wild Species, Kunming Institute of Botany, Chinese Academy of Sciences provided molecular laboratory facilities for molecular work. We express our sincere acknowledgement to Emeritus Prof. Dr. Kevin D. Hyde for his valuable comments and suggestions. Rungtiwa Phookamsak is grateful to CAS President’s International Fellowship Initiative (PIFI) for young staff (grant no. 2019FYC0003), Post–Doctoral Fellowship 2022 from Chiang Mai University, Thailand (Grant No. R000031743) and Reinventing University System 2021, Mae Fah Luang University for providing visiting scholarship. Jianchu Xu thanks Yunnan Provincial Science and Technology Department, Key Project (Grant No. 202101AS070045) and NSFC-CGIAR Project 'Characterization of roots and their associated rhizosphere microbes in agroforestry systems: ecological restoration in high-phosphorus environment' (Grant No. 31861143002). Rajesh Jeewon would like to thank University of Mauritius for research support. Sinang Hongsanan would like to thank National Natural Science Foundation of China (grant no. 31950410548) for financial support. Hong-Bo Jiang would like to thank Mae Fah Luang University for Ph.D scholarship. Timur S. Bulgakov would like to thank the Federal Research Center “Subtropical Scientific Center of the Russian Academy of Sciences” (the State Task research, theme no. FGRW-2022-0006). Jun-Fu Li thanks Deping Wei, Dan-Feng Bao, Er-Fu Yang, Dr. Shaun Pennycook, Dr. Dhanushka Wanasinghe, Milan C. Samarakoon, Dr. Mingkwan Doilom, Ningguo Liu and Qing Tian for their suggestions and assistance. Austin G. Smith at World Agroforestry (ICRAF), Kunming Institute of Botany, China, is thanked for English editing.
-
Jun-Fu Li, Rajesh Jeewon, Darbhe Jarayama Bhat, Peter Edward Mortimer, Rungtiwa Phookamsak and Nakarin Suwannarach are the Editorial Board members of Journal Studies in Fungi. They are blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of these Editorial Board members and their research groups.
- Supplemental Table S1 Taxa used for phylogenetic analyses and divergence time estimation of Pleosporales in this study (excluding Alteraria species).
- Supplemental Table S2 Alternaria taxa used in the evolutionary and phylogenetic analyses and their corresponding GenBank numbers, including two outgroup taxa (Pleospora tarda CBS 714.68 and Stemphylium herbarum CBS 191.86). The type strains are indicated in ”T” and the newly generated sequences are indicated in blue.
- Supplemental Table S3 Strains used in the phylogenetic analysis of hosts and regional distribution of Alternaria alternata (sect. Alternaria) and their corresponding GenBank numbers, including two outgroup taxa (Alternaria eichhorniae CBS 489.92 and CBS 119778). The type strains are indicated by superscript “T” and the reference strains are indicated by superscript “R”. The newly generated sequences are indicated in blue.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Li JF, Jiang HB, Jeewon R, Hongsanan S, Bhat DJ, et al. 2023. Alternaria: update on species limits, evolution, multi-locus phylogeny, and classification. Studies in Fungi 8:1 doi: 10.48130/SIF-2023-0001 

Alternaria: update on species limits, evolution, multi-locus phylogeny, and classification
- Received: 08 July 2022
- Accepted: 23 December 2022
- Published online: 31 January 2023
An Author Correction to this article was published on 08 March 2024,
http://doi.org/sif-0024-0001.
Abstract: Alternaria, a genus of ascomycetes, comprises major plant pathogens, saprobes and are common allergens to humans. There are more than 360 accepted species in the genus, which are currently divided into 29 sections. This paper aims to elaborate the taxonomy of Alternaria with multi-locus phylogenetic trees derived by analyses of a concatenated DNA sequence dataset consisting of ITS, LSU, TEF1-α, RPB2, GAPDH and Alt-a1 loci. Eighteen new species viz. Alternaria arctoseptata, A. arundinis, A. baoshanensis, A. breviconidiophora, A. brevirostra, A. ellipsoidialis, A. eupatoriicola, A. falcata, A. lathyri, A. macilenta, A. macroconidia, A. minimispora, A. nodulariconidiophora, A. oblongoellipsoidea, A. orobanches, A. phragmiticola, A. phytolaccae and A. salicicola are introduced and classified in sect. Alternaria, sect. Infectoriae, sect. Porri and sect. Radicina. Alternaria alternata and A. doliconidium are also described herein with new host and geographical records, in China, Italy, and Thailand. This study further explores the utility of divergent time estimates to gain additional insights into the evolutionary relationships of Alternaria in Pleosporales.
-
Key words:
- 18 new taxa /
- Dematiaceous hyphomycetes /
- Dothideomycetes /
- Pleosporaceae /
- Pleosporales /
- Taxonomy