









-
Many plants grow in an open environment and are exposed to multiple biotic and abiotic stresses[1]. To survive adverse conditions and minimize stress-induced damage, plants have evolved defense mechanisms[2]. Transcription factors, which play a crucial role during plant life, regulate gene transcription levels and activate the synthesis of plant components and responses to environmental changes[3].
The plant-specific TCP [i.e., TEOSINTE BRANCHED1/CYCLOIDEA/PROLIFERATING CELL FACTOR] family comprises transcription factors that participate in many developmental processes and stress responses. TEOSINTE BRANCHED1 was originally described in maize (Zea mays), whereas CYCLOIDEA and PCF (i.e., proliferating cell nuclear antigen factor 1 and 2) were first described in snapdragon (Antirrhinum majus) and rice (Oryza sativa), respectively[4−6]. The TCP gene family members encode proteins that contain an N-terminal TCP domain, a highly conserved basic helix-loop-helix (bHLH) motif consisting of 59 amino acids involved in protein-protein interactions, DNA binding, and protein nuclear localization[7]. According to the TCP domain sequence features, the TCP transcription factors have been divided into the following two subfamilies: Class I (i.e., TCP-P family), which includes PCF1 and PCF2, and Class II (i.e., TCP-C family), which includes CYC and TB1. There is a clear difference between the two subfamilies, with the Class I members missing four amino acids in the basic region of the bHLH motif. Additionally, the Class II members have been subdivided into two subclasses (CIN and CYC/TB1) according to their TCP domain differences; some Class II members contain an arginine-rich motif comprising 18–20 amino acids (R domain).
There is increasing evidence that TCP family members have versatile functions affecting multiple biological processes and traits throughout the plant lifespan, including branching[8], leaf size and curvature[9], flower development[9], circadian rhythms[10], seed germination[11], hormone pathways[12], plant immunity, and environmental stress tolerance[13,14]. The TCP genes encode plant-specific proteins that are involved in multiple abiotic stress responses. In rice, OsTCP19 influences abiotic stress (water-deficit and salt stress response) and developmental signaling by interacting with and modulating the activity of OsABI4[14]. The expression of PeTCP10 in transgenic Arabidopsis thaliana (Arabidopsis) plants reportedly enhances salt tolerance during the vegetative growth stage, but it has the opposite effect during the germination and seedling stages[15]. Additionally, TCP genes contribute to various biotic stress responses. The overexpression of
miR319a in Populus tomentosa decreases the transcription of TCP19, leading to a significant increase in leaf trichome density and a decrease in insect herbivory[16]. In Arabidopsis, a TCP directly regulates the EFR-dependent immune response to Pseudomonas syringae; the tcp8 tcp14 tcp15 triple mutant exhibits abnormal EFR-dependent PTI (PAMP-triggered immunity), with decreased production of PATHOGENESIS-RELATED PROTEIN 2 and upregulated expression of EFR after an elicitation with elf18[17]. Both MOS1 and TCP15 bind directly to the promoter of the immune receptor gene SUPPRESSOR OF NPR1-1, CONSTITUTIVE 1 (SNC1) to regulate its expression and the associated immune response[18]. Several TCP family members have recently been identified in various plants with a completely sequenced genome, including 24 TCP genes in Arabidopsis[19], 18 in Vitis vinifera[20], 34 in Pyrus bretschneideri[21], 18 in Ipomoea batatas[22], 25 in Dendrobium catenatum[23], and 32 in Petunia axillaris[24]. Rosa chinensis Jacq. is a widely cultivated ornamental plant and an important contributor to the cut-flower industry. Chinese rose (Rosa chinensis) is one of the main varieties crossed with European roses to produce modern rose varieties because of its recurrent flowering trait as well as its color and scent[25]. Rose plant growth and flowering are affected by many factors. Drought stress leads to decreased absorption of nutrients as well as leaf wilting and curling[26], while heat stress prevents plants from blooming normally and results in the production of deformed flowers[27]. Globally, black spot, botrytis, and powdery mildew are important fungal diseases that can lead to discoloration, the development of spots, the shedding of leaves and flowers, and even plant death in serious cases, thereby greatly affecting the ornamental and economic value of rose[28].
The recent sequencing of the R. chinensis genome[29] has enabled analyses of the TCP gene family, which is important for plant growth and environmental stress responses. This study provides a comprehensive analysis of the RcTCP transcription factor family in rose. Specifically, 18 non-redundant R. chinensis TCP genes were identified and then systematically analyzed in terms of their similarities to related sequences, conserved motifs, gene structures, phylogenetic analysis, cis-acting elements, and chromosomal locations. Furthermore, rose TCP gene expression patterns in response to diverse environmental stresses (fungal diseases and abiotic) were revealed. The study results may be useful for functionally characterizing the rose RcTCP gene family, which may be relevant for breeding programs interested in enhancing rose stress resistance.
-
Rosa chinensis genomic data were downloaded from the Rosa chinensis Old Blush homozygous genome v2.0 (
www.rosaceae.org/analysis/282 )[29]. The corresponding TCP protein sequences were obtained from the Pyrus bretschneideri Rehd. database (https://genome.cshlp.org/content/23/2/396 ) and the Arabidopsis Information Resource database (www.arabidopsis.org ). Candidate TCP members in R. chinensis were identified via a local BLASTP[30] search using Arabidopsis and pear sequences as queries. The TCP-containing sequences in R. chinensis were analyzed using the hidden Markov model of the TCP domain (PF03634) to screen the Old Blush homozygous genome v2.0 genome portal database to identify candidates (E-value < 1e−20). All sequences were further analyzed using online tools, including the NCBI Conserved Domain Database web server, PfamScan[31], and Pfam A[32]. Finally, sequences with complete TCP domains were named according to their locations on chromosomes (Supplemental Table S1).Sequence analysis of RcTCP
-
The TCP domain sequences were aligned using the default parameters of DNAMAN. The RcTCP protein secondary structures were examined using the Self-Optimized Prediction Methods with Alignment program (
https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html )[33]. On the basis of the combined alignments, typical conserved non-canonical bHLH domains in TCP proteins were marked in the corresponding sequences (Fig. 1).
Figure 1.
Protein sequence alignment of the RcTCP family members. Black, red and blue indicate different identities.
The RcTCP exon-intron organization was predicted using the Gene Structure Display Server (
http://gsds.cbi.pku.edu.cn ) and obtained by comparing the predicted coding sequences with the full-length sequences[34] (Fig. 2). Identified conserved motifs in RcTCP proteins were analyzed using the MEME online program (http://meme.nbcr.net/meme/intro.html )[35]. The parameters are set as: any number of repetitions; optimal motif width from 6 to 100 residues; the maximum number of motifs is set to 10 (Fig. 2 & 3).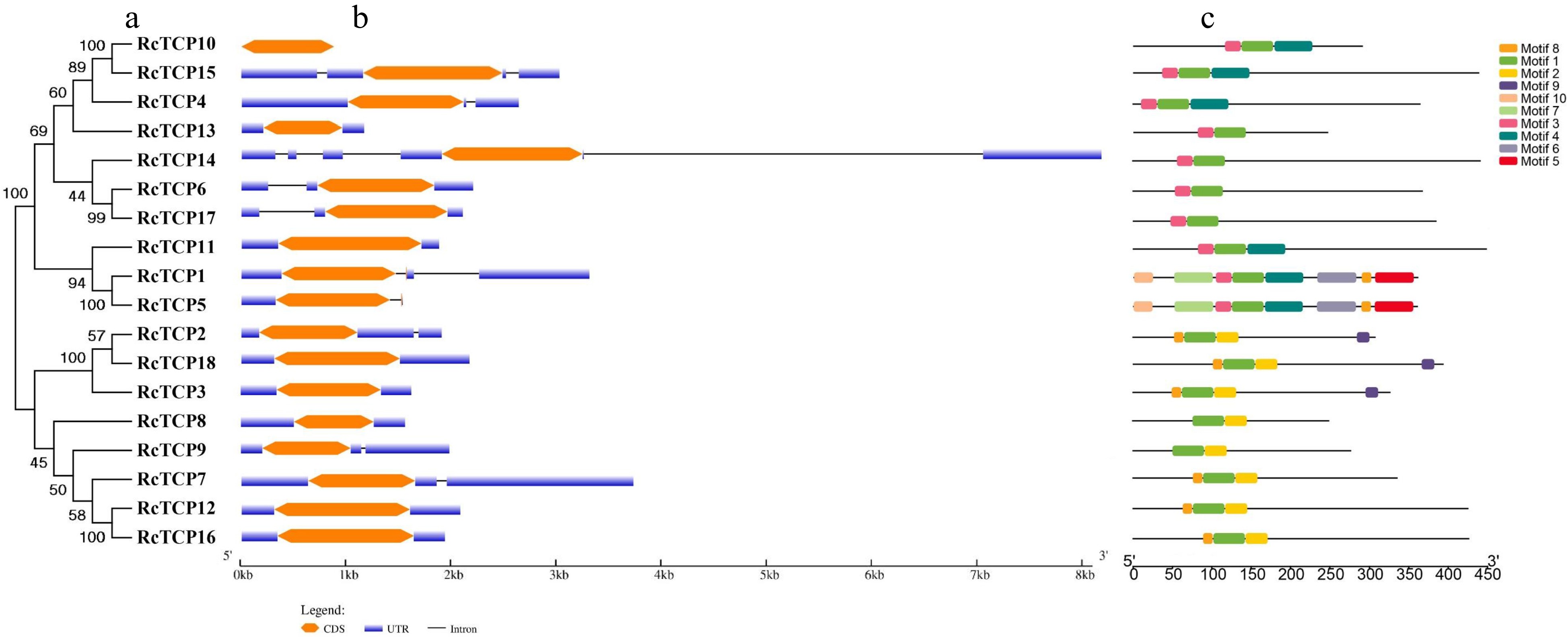
Figure 2.
Phylogenetic tree, conserved motif and gene structure analysis of TCP family members in R. chinensis. (a) A phylogenetic tree comprising RcTCP family proteins was constructed (1,000 replicates of the bootstrap test). (b) Exon–intron structures of RcTCP family members. Blue indicates 5′ and 3′ untranslated regions (UTRs), orange indicates exons, and black indicates introns. (c) Distribution of conserved motifs in RcTCP family members. Different colors and numbers represent different motifs (1 to 10).

Figure 3.
Phylogenetic analysis of the TCP transcription factors in R. chinensis, Arabidopsis thaliana, and Pyrus bretschneideri. The phylogenetic tree was constructed according to the Maximum Likelihood method (1,000 replicates of the bootstrap test) using MEGA 6.0. Blue, red, and purple indicate the PCF, CIN, and CYC/TB1 clades, respectively.
Phylogenetic analysis of the rose TCP proteins
-
The identified RcTCP genes were divided into different classes according to the AtTCP and PbTCP classification scheme and the alignment of the TCP domains in the RcTCP, PbTCP, and AtTCP proteins. The analysis was performed using the MEGA 6.0 software Maximum Likelihood (ML) method, and the phylogenetic tree was obtained by executing the parameters: BootStrap replicated 1000 times, selecting the Poisson model, and performing partial deletion (Fig. 3).
Gene localization and analysis of cis-acting elements in RcTCP promoters
-
The RcTCP genomic location data were obtained from genome annotation files. A gene location map was constructed using MapChart (
http://mg2c.iask.in/mg2c_v2.0/ ) (Fig. 4). Sequences (2.0 kb) upstream of the RcTCP coding sequences were selected from the OldBlush Hm r2.0 genome portal database and then submitted to PlantCARE online analysis software (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) to identify putative regulatory elements. The TBtools software[36] was used to plot the enrichment of each cis-acting element in the RcTCP promoters.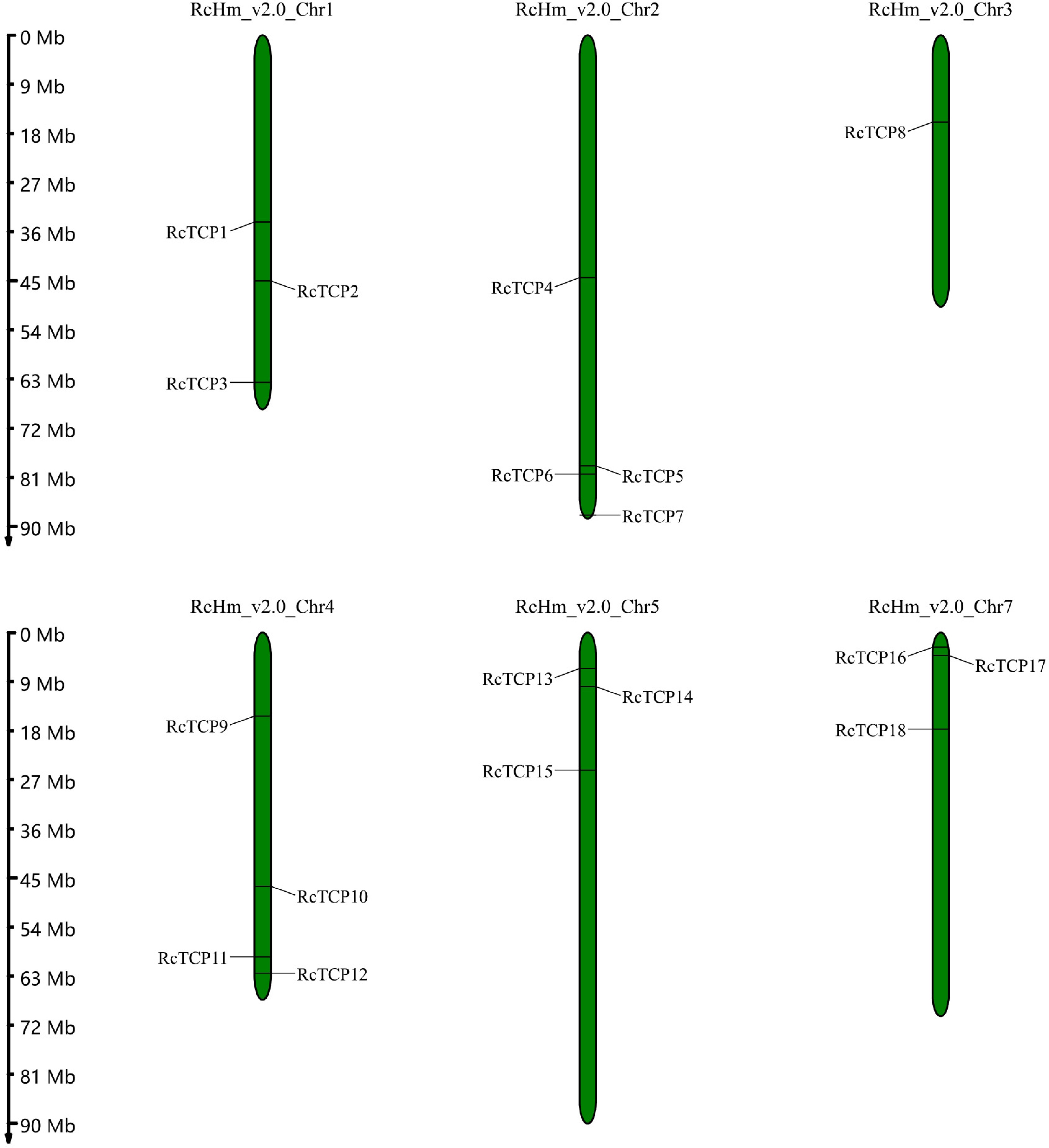
Figure 4.
Chromosomal locations of RcTCP genes. The numbers on the left side of the bars indicate the approximate physical location of the first exon of the corresponding RcTCP gene on rose chromosomes.
Transcriptomic resources
-
The RcTCP expression profiles in response to drought, salt stress and heat stress were analyzed using the SRP314899 (
www.ncbi.nlm.nih.gov/sra?term=SRP314899 ), SRP228321 (www.ncbi.nlm.nih.gov/sra/?term=SRP228321 ) and SRP150297 (www.ncbi.nlm.nih.gov/sra/?term=SRP150297 ) published data, respectively. Specifically, SRP314899 comprised data for Rosa chinensis Jacq. var. chinensis seedlings exposed to drought for 0, 30, and 60 days, SRP228321 consisted of data for R. chinensis 'Old Blush' roots exposed to salt stress for 0, 2, 24, and 48 h, and SRP150297 consisted of data for R. chinensis 'Old Blush' leaves under heat stress conditions for 0, 0.5, 2, 6, and 12 h.The RcTCP expression profiles in response to powdery mildew and Botrytis cinerea were analyzed using the SRP228524 (
www.ncbi.nlm.nih.gov/sra?term=SRP228524 ) and SRP120271 (www.ncbi.nlm.nih.gov/sra/?term=SRP120271 ). Specifically, SRP228524 comprised data for powdery mildew-infected leaves from Rosa hybrida 'Confetti' (susceptible genotype) and 'IIHRR13-4' (resistant genotype) plantlets at 0, 36, and 72 h post-inoculation, whereas SRP120271 consisted of data for R. hybrida petals infected with B. cinerea or treated with PDB medium (control) for 30 and 48 h.Plant materials and growth conditions
-
Rose cultivar 'Zhuliye', which was sensitive to Alternaria leaf spot disease obtained from Suzhou Polytechnic Institute of Agriculture (Jiangsu, China). Seedlings that were growing relatively uniformly were transplanted into a mixed matrix comprising vermiculite and perlite (3:1) and then incubated in a growth chamber at 25 °C with 40% relative humidity and a 16-h light (200 μmol m−2 s−1) : 8-h dark cycle. An Alternaria alternata strain was isolated from typical diseased 'Zhuliye' leaves and then stored in 15% (v/v) glycerol at −80 °C.
Inoculation with A. alternata
-
The isolated A. alternata strain was grown on potato dextrose agar medium in Petri dishes. To prepare the inoculum, sterile distilled water was added to the Petri dishes and the resulting liquid was further filtered. The final spore concentration was adjusted to 106 spores/mL using half-strength filtered organic grape juice as previously described[37]. Young leaves collected from rose shoots and then placed on 1% agar in Petri dishes. Each leaf was inoculated with 5 μL inoculum, after which the inoculated and control leaves were incubated in darkness at 28 °C and 90% humidity. At 6 and 24 h post-inoculation, the inoculated and control (0 h) leaves were collected and immediately frozen in liquid nitrogen for later RNA extraction experiments.
Total RNA extraction and RcTCP expression analysis
-
Total RNA was extracted from the frozen samples using the Total RNA Isolation System (Takara, Kyoto, Japan). The quality of the extracted RNA was assessed using the 2100 Bioanalyzer RNA Nano Chip device (Agilent, Santa Clara, CA, USA). The RNA samples with a λ260/280 ratio of 1.8–2.1 and a λ260/230 ratio of 2.0–2.5 were retained for the synthesis of cDNA using the ReverTra Ace-α First Strand cDNA Synthesis kit (TOYOBO, Shanghai, China).
For the quantitative real-time polymerase chain reaction (qRT-PCR) analysis, primers used were designed by Primer5 software (Supplemental Table S2), and RcGAPDH was selected as the reference gene[38]. The qRT-PCR assay was completed using the SYBR Premix Ex Taq™ Kit (Takara, Dalian, China) and the Mastercycler® ep realplex 2S device (Eppendorf, Hamburg, Germany). All samples were analyzed using three biological replicates. The reaction system was 25 μL, in which the cDNA content was 10 ng, the forward and reverse primers were both 0.2 μM, and the SYBR Green PCR master mix was 10 μL. The PCR program was set as follows: firstly, 95 °C for 2 min; then for 40 cycles (95 °C for 15 s, 60 °C for 15 s, and 72 °C for 15 s). The relative expression level was computed by the 2−ΔΔCᴛ[39] method.
-
To identify rose TCP genes, Arabidopsis and P. bretschneideri TCP protein sequences were used as queries to screen the rose protein dataset (
https://lipm-browsers.toulouse.inra.fr/pub/RchiOBHm-V2/ ) using BLASTP. The rose TCP genes were identified on the basis of sequence similarities and whether they encoded TCP domains and conserved motifs. A total of 18 non-redundant RcTCP genes on six rose chromosomes were identified and renamed as RcTCP1–18 according to their genomic distribution and relative linear order on their respective chromosomes (Supplemental Table S1). Gene characteristics, including chromosomal location, E-value, and strand, as well as the length of the encoded protein sequence, isoelectric point (pI), the protein molecular weight, and the predicted subcellular localization were analyzed (Supplemental Table S1).The RcTCP proteins varied in length from 246 (RcTCP5) to 448 (RcTCP3) amino acid residues. The proteins with the lowest and highest molecular weights were RcTCP5 (28.09 kDa) and RcTCP3 (50.04 kDa), respectively. The theoretical pI ranged from 6.32 (RcTCP3) to 9.17 (RcTCP5). The minimum and maximum aliphatic indices were 54.39 and 80.41, respectively, implying that the RcTCP proteins are rich in aliphatic amino acids. The GRAVY values for the 18 RcTCP proteins were all negative, indicative of the hydrophilic properties of the TCP proteins in rose. Additionally, the subcellular localization analysis suggested that 12 RcTCP proteins are present in the nucleus, whereas the other six RcTCP proteins are secreted extracellularly (Supplemental Table S1).
Multiple sequence alignment, classification, and phylogenetic relationship of RcTCP family members
-
The phylogenetic relationships among the RcTCP proteins were determined on the basis of a multiple sequence alignment of a conserved non-canonical bHLH domain comprising approximately 60 amino acids. The conserved domains of the TCP proteins were searched using DNAMAN, which revealed the following four conserved motifs: Basic, Helix I, Loop, and Helix II (Fig. 1). The constructed phylogenetic tree divided the TCP proteins into two categories, namely Classes I and II (Fig. 1 and 2a). Class I proteins differ from Class II proteins by the deletion of four amino acids in the basic region. In most cases, the TCP proteins in the same phylogenetic group shared similar motifs. Class I included eight RcTCP proteins (RcTCP2, 3, 7–9, 12, 16, and 18), whereas the other 10 proteins were in Class II subclass, which was further subdivided into the CYC/TB1 clade comprising three proteins (RcTCP1, 5, and 11) and the CIN clade consisting of seven proteins (RcTCP4, 6, 10, 13–15, and 17) (Fig. 1).
Gene structures and conserved motifs among RcTCP family members
-
To further structurally characterize the RcTCP proteins, we investigated the exon–intron organization of the corresponding genes and the encoded conserved motifs. Of the 18 RcTCP genes, 10 were revealed to contain introns in the 5′ or 3′ untranslated region, of which seven contain one intron, two contain two introns, and one contains four introns (Fig. 2b). Ten putative conserved motifs were detected in the RcTCP proteins using the MEME program (Fig. 2c). Motif 1, which was identified as the highly conserved TCP domain, was present in all RcTCP proteins, reflecting the reliability of our analysis. Although the functions of the remaining nine motifs are unknown, similar motifs were detected in the RcTCP proteins in the same subfamily, whereas there were significant differences in the motifs between subfamilies, suggestive of functional redundancy within subfamilies and functional diversity between subfamilies. The RcTCP proteins clustered in the same subgroup had similar motifs, which was consistent with the results of the gene structure analysis.
The TCP domain typically consists of a basic region and a helix-loop-helix structure. The basic region, which was relatively conserved between Classes I and II, included a consensus sequence (RxRRxR) (Supplemental Fig. S1a). In Arabidopsis, a glycine (G) residue at the 11th position in the RxRRxR region of Class I members corresponds to the aspartic acid (D) at the 15th position of Class II members; this amino acid difference may be related to the diversity of gene targets of the two classes[40]. We observed that the residues at these positions were highly conserved in Classes I and II. Hence, the region with these amino acid positions may be important for the differential functions among the RcTCP proteins. Previous research demonstrated that multiple TCP proteins in Arabidopsis contain R domains, such as AtTCP1, AtTCP12 and AtTCP18 in the class II CYC/TB1 clade and AtTCP2 and AtTCP24 in the CIN clade[41]. In R. chinensis, RcTCP1, RcTCP5, and RcTCP11 in the CYC/TB1 subclade encode proteins with the R domain (Supplemental Fig. S1b).
Evolutionary analysis of the RcTCP family
-
To investigate the evolutionary relationships in the RcTCP transcription factor family, we performed a phylogenetic analysis of the known TCP family members using the amino acid sequences encoded by the respective genes as the input data. The TCP proteins from Arabidopsis and P. bretschneideri were included in the analysis, which divided the TCP proteins into Classes I and II according to the divergence between their TCP domains (Fig. 3). Interestingly, for all sequences in a particular TCP group, there was significant genus-specific clustering in both R. chinensis and P. bretschneideri.
Chromosomal locations and synteny among RcTCP genes
-
The chromosomal locations of the RcTCP genes are presented in Fig. 4. The RcTCP genes were unevenly distributed on six of the seven R. chinensis chromosomes, with 0–4 RcTCP genes per chromosome. Chromosome 6 lacked RcTCP genes. Chromosome 3 contained one RcTCP genes, whereas chromosomes 1, 5, and 7 had three RcTCP genes each. Four RcTCP genes were localized to chromosomes 2 and 4. Moreover, eight Class I RcTCP genes were located on different chromosomes. In contrast, RcTCP2 and RcTCP3 (chromosome 1), RcTCP9 and RcTCP12 (chromosome 4), and RcTCP16 and RcTCP18 (chromosome 7) were detected on the same chromosomes. Additionally, three Class II CYC/TB1 subclade RcTCP genes were located on different chromosomes. Of the seven Class II CIN subclade RcTCP genes, RcTCP4 and RcTCP7 were distributed on chromosome 2, RcTCP13, RcTCP14, and RcTCP15 were detected on chromosome 5, and RcTCP10 and RcTCP17 were localized to chromosomes 4 and 7, respectively.
Stress-related cis-acting elements in RcTCP promoters
-
To clarify how RcTCP expression is regulated during biotic and abiotic stress responses, the cis-acting elements in the 2.0-kb regions upstream of the RcTCP translation start sites were identified using PlantCARE. The following nine stress response-related elements were detected: MYB, GT1-motif, ABRE, G-box, W-box, TC-rich repeats, DRE, MYC, and LTR (Fig. 5). All of the RcTCP promoters contained several stress response-related cis-acting elements, suggesting that RcTCP genes are stress-responsive genes. All 18 RcTCP promoters had at least one MYB element, and all except the RcTCP14 promoter contained MYC elements. The TC-rich repeats and DRE elements were detected in nine RcTCP promoters, whereas the W-box was identified in 12 RcTCP promoters. Moreover, 16 and 14 RcTCP promoters contained G-box and ABRE sequences, respectively. Seven RcTCP promoters consisted of 1–3 GT1-motifs, whereas six RcTCP promoters had one or two LTR elements. These findings imply that RcTCP genes are involved in plant responses to various stresses.

Figure 5.
Predicted cis-acting elements in RcTCP promoters. Promoter sequences (−2,000 bp) of the 18 RcTCP genes were analyzed using PlantCARE. The scale at the bottom presents the nucleotide positions upstream of the translation start site.
RcTCP expression profiles under abiotic and biotic stress conditions
-
Using the RNA-seq data, a heatmap for the drought-induced expression levels (i.e., FPKM values) of the 18 RcTCP genes was constructed (Fig. 6). With the exception of RcTCP10, the RcTCP genes were expressed under drought conditions. Some of the RcTCP genes had similar drought-induced expression patterns. For example, RcTCP3 and RcTCP7 expression levels were significantly up-regulated at 60 d after initiating the drought treatment. Twelve genes (RcTCP4, 6, 8, 9, 11–18) had up-regulated expression levels on day 30 and down-regulated expression levels on day 60 of the drought treatment. The expression levels of the other genes also fluctuated after 30 and 60 d of the drought treatment. The RcTCP expression patterns in response to heat stress are presented in Fig. 6b. After the heat treatment, the RcTCP1 and RcTCP5 expression levels were up-regulated, whereas the RcTCP4, 8, 9, 12, 14, and 15 expression levels were down-regulated. In contrast, RcTCP16 expression was up-regulated and then down-regulated, while the expression of RcTCP2, 3, 6, 7, 17, and 18 was down-regulated and then up-regulated. The changes in RcTCP expression levels following the salt treatment are shown in a heat map (Fig. 6c). As the salt treatment time increased, the expression of RcTCP2 gradually increased, whereas the expression of RcTCP6, 8, 12, and 13 gradually decreased. The RcTCP3 and RcTCP14 expression levels were down-regulated and then up-regulated, while the RcTCP4, 7, 9, 15, 16, and 18 expression levels were up-regulated and then down-regulated.

Figure 6.
RcTCP expression profiles in response to (a) drought, (b) heat stress and (c) salt stress. The log2-transformed FPKM values for the RcTCP genes were used to construct a heatmap with the HemI software. (a) GH1: 30 d after initiating the drought treatment; GH2: 60 d after initiating the drought treatment. (b) H 0h, H 0.5h, H 2h, H 6h, H 12h: heat treatment for 0, 0.5, 2, 6, 12 h. (c) Salt stress for 0, 2, 24, 48 h.
The RcTCP expression trends in response to fungal diseases (i.e., powdery mildew, Botrytis cinerea and black spot infections) were also analyzed. Heatmaps presenting the expression levels of the 18 RcTCP genes in leaves infected with powdery mildew were constructed on the basis of RNA-seq data (Fig. 7). In the resistant cultivar 'IIHRR13-4', the expression levels of seven RcTCP genes (RcTCP3, 4, 12, 14, 15, 16, and 17) were down-regulated and the expression levels of eight RcTCP genes (RcTCP2, 5, 6, 7, 8, 9, 13, and 18) were up-regulated after 32 h of the powdery mildew treatment. In the susceptible cultivar 'Confetti', nine RcTCP genes (RcTCP2, 3, 4, 5, 8, 12, 14, 15, and 16) had down-regulated expression levels after 32 and 72 h of the powdery mildew treatment, but the expression levels of three RcTCP genes (RcTCP7, 8, and 18) were up-regulated to varying degrees. The RcTCP6 and RcTCP17 expression levels were initially up-regulated, but were subsequently down-regulated, while the expression of the other genes did not change significantly. The RcTCP expression levels in R. hybrida petals infected with B. cinerea were analyzed. Interestingly, except for the expression of RcTCP12, which was up-regulated at 30 and 48 h post-inoculation, the RcTCP expression levels were down-regulated to varying degrees.

Figure 7.
RcTCP expression profiles in response to (a) powdery mildew and (b) B. cinerea. The log2-transformed FPKM values for the RcTCP genes were used to construct a heatmap with HemI software. (a) RPMR0, RPMR3 represent powdery mildew treatment of resistant genotype 'IIHRR13-4' for 0 and 32 h, respectively. The SCON0, SCON3, SCON7 represent powdery mildew treatment of susceptible cultivar 'Confetti' for 0, 32 and 72 h. (b) CK-30 h, CK-48 h: petal with PDB for 30 and 48 h, TR-30 h, TR-48 h: petal with B. cinerea 30 and 48 hpi.
To explore the changes in RcTCP expression induced by A. alternata, which causes black spot disease, a qRT-PCR analysis was performed to examine the transcript levels of each RcTCP gene (Fig. 8). The data revealed that the expression levels of all RcTCP genes were up-regulated at 6 h post-inoculation. Furthermore, the up-regulated expression levels were more obvious at the 12 h time-point. Specifically, RcTCP2 and RcTCP1 expression levels were up-regulated 50-times and 30-times, respectively, possibly reflecting the importance of these two genes for rose responses to black spot disease.
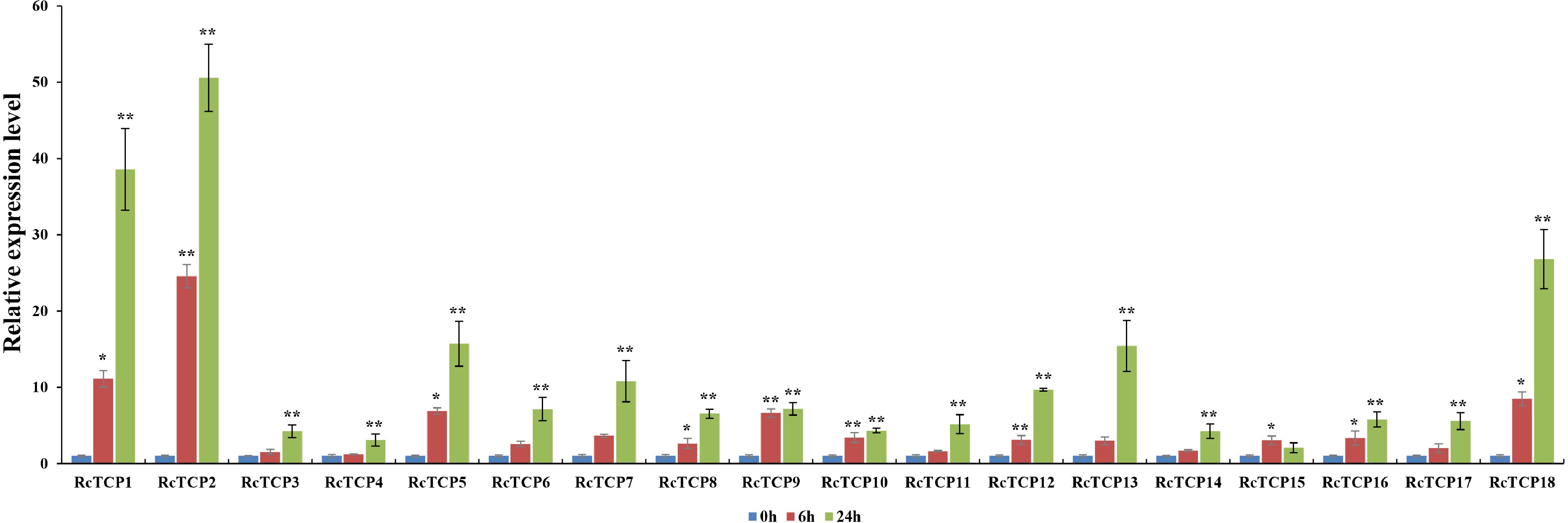
Figure 8.
RcTCP expression profiles in response to an A. alternata infection. 0 h, 6 h, 24 h: A. alternate treatment for 0, 6 and 24 h. The error bars indicate SEs (n = 3). * Significant difference: P < 0.05, ** Significant difference: P < 0.01.
-
TCP genes encode plant-specific proteins that mediate various biological activities related to plant growth and responses to environmental stimuli[42]. The sequencing of plant genomes has resulted in the identification of many TCP family members in various plants, including Arabidopsis[19], V. vinifera[20], P. bretschneideri[21], I. batatas[22], D. catenatum[23], and P. axillaris[24]. In this project, we performed a systematic analysis of the RcTCP transcription factor family in rose by exploring their sequence similarities, gene structures, phylogenetic analysis, conserved motifs, cis-acting elements, chromosomal locations, and expression patterns in response to different environmental stresses (biotic and abiotic). The results of this study will provide researchers with relevant information for future analyses of the biological and physiological functions of TCP proteins, while also enhancing our understanding of the role of TCP transcription factors in plant responses to environmental stresses.
Analysis of the RcTCP gene family and its evolution
-
In this study, 18 non-redundant RcTCP genes were identified in R. chinensis. On the basis of multiple sequence alignment and phylogenetic relationship analysis, the 18 RcTCP genes were divided into two classes (three main subgroups), consistent with the results of earlier studies on Arabidopsis and P. bretschneideri[19−21]. Each subgroup contained TCP genes from Arabidopsis, P. bretschneideri, and rose (Fig. 3), indicating the RcTCP family is relatively conserved. The number of amino acids in the RcTCP family members differed considerably, from 246 for RcTCP5 to 448 for RcTCP3 (Supplemental Table S1), reflecting the potential complexity of the origin and evolution of the rose TCP family. Additionally, the difference in the number of amino acids may be associated with the diversity in the biological functions of the TCP transcription factors in rose. The RcTCP members in the same subgroup were similar in terms of their motif composition and exon–intron organization (Fig. 2). For example, motifs 2 and 9 were present only in Class I, whereas motifs 3 and 4 were exclusive to Class II (Fig. 2). The consistency in the encoded motifs and the exon–intron structures among the RcTCP genes was suggestive of the close evolutionary relationships among these genes. The organization of introns and exons as well as the distribution of specific motifs indicated that the genes within a subgroup had similar structures. Furthermore, the similarities in the gene sequences were associated with tandem repeats that arose during evolution. These phenomena are important for gene functional diversity.
Analysis of cis-acting elements of the RcTCP genes
-
The level of transcriptional activation is coordinated by upstream cis-acting elements, which are critical for plant responses to environmental conditions[43]. In terms of cis-acting elements, the RcTCP promoters were revealed to contain some common motifs and many repetitive regions. In the current study, the 2-kb region upstream of RcTCP genes was analyzed using PlantCARE, which detected multiple cis-acting elements associated with biotic or abiotic stresses, including the light-responsive G-box element and the ABA-responsive ABRE and MYC sequences[44]. All RcTCP promoters included MYB elements, which are important for plant development and stress responses. In addition, the presence of MYC (except in the RcTCP14 promoter) suggests that the expression of these RcTCP genes may be associated with plant responses to drought conditions. The W-box was detected in the promoters of most of the RcTCP genes (e.g., RcTCP1–4, 7–9, 11, 13–15, and 18)[45], which is in accordance with the differential expression of these genes following a pathogen infection. This suggests that RcTCP expression induced by various biotic and abiotic stresses may depend on specific cis-acting elements.
RcTCP expression profiles under abiotic stress conditions
-
To clarify the functions of RcTCP proteins in plants exposed to drought stress, the RcTCP expression patterns were analyzed on the basis of RNA-seq (SRP314899) and expression-array data. There is considerable evidence that TCP genes encode crucial regulators of plant growth and development as well as proteins that confer tolerance to abiotic stresses (e.g., salinity and drought). The overexpression of the Z. mays gene TCP42 in Arabidopsis leads to ABA hypersensitivity during seed germination and enhanced drought tolerance[46]. The HrTCP20 protein positively regulates the drought resistance of Hippophae rhamnoides through its effects on the JA signaling pathway[47]. The analysis of the drought stress-related transcriptome data revealed the expression of 12 RcTCP genes was significantly up-regulated and down-regulated at 30 and 60 d after starting the drought treatment, respectively, suggesting RcTCP transcription factors contribute to the drought tolerance of rose plants. In addition, the changes in RcTCP expression differed between the heat and salt treatments. Interestingly, RcTCP14 expression was significantly affected by the drought, heat, and salt treatments, suggesting it is important for rose responses to diverse abiotic stresses. How RcTCP14 contributes to the abiotic stress resistance of rose will need to be investigated.
Fungal diseases-induced RcTCP expression profiles
-
Rosa chinensis growth and development are affected by many diseases, among which black spot, powdery mildew, and botrytis are important fungal diseases that can severely restrict the ability of R. chinensis to complete its life-cycle. In this study, RcTCP expression patterns in response to fungal diseases were comprehensively analyzed[28]. More specifically, the available transcriptome data and the results of a qRT-PCR assay involving plants infected with powdery mildew, botrytis or black spot were analyzed. Gene expression and phylogenetic analyses were conducted, and the RcTCP genes were functionally characterized according to the known functions of TCP genes from model plants. In Arabidopsis, several TCP genes encode proteins involved in plant immune responses, including TCP15, which interacts with MOS1 and binds to the SNC1 promoter, thereby regulating gene expression and immune responses[18]. The tcp8 tcp14 tcp15 triple mutant reportedly has a defective EFR-dependent PTI and an abnormal EFR-dependent immune response to P. syringae[17]. Rose is an important ornamental plant species worldwide because of its substantial economic and cultural value. Powdery mildew, which is caused by Podosphaera pannosa, is one of the most damaging and widespread fungal diseases of rose. The expression trends of most genes were similar among varieties, but the extent of the expression level changes varied. The RcTCP2 and RcTCP9 expression levels were significantly up-regulated in the resistant varieties, but they were down-regulated in the susceptible varieties. We speculated that RcTCP2 and RcTCP9 might be key factors for the resistance of different varieties to powdery mildew. This possibility will need to be verified in future studies. The infection with B. cinerea resulted in down-regulated RcTCP expression levels, with the most obvious decreases observed for RcTCP6, 15, and 18. Accordingly, these genes may be negatively regulated by botrytis. The up-regulated expression of RcTCP12 may affect the disease resistance mechanism in different ways. These findings are indicative of the extensive involvement of TCPs in rose plant responses to fungal diseases.
The cultivation of rose under greenhouse conditions is threatened by many diseases. For example, A. alternata, which causes black spot disease in temperate regions[48], is a common necrotrophic fungus that infects diverse plants and damages tissues by producing toxins[49]. Warm and humid environmental conditions lead to the spread of diseases caused by Alternaria species, which inhibit rose production throughout the year. The qRT-PCR analysis in the current study revealed that the expression of 18 RcTCP genes was up-regulated at 6 and 24 h after the inoculation with A. alternata. This finding may be relevant for future investigations on TCP-mediated plant responses to black spot disease. The RcTCP1 and RcTCP2 expression levels were substantially up-regulated after 24 h of the A. alternata infection, suggesting that these genes may encode proteins with critical functions in different stages of the black spot infection. Because of the changes in TCP gene expression induced by the exposure to biotic stress and the frequent occurrence of black spot disease in rose, the RcTCP genes will need to be more thoroughly functionally characterized.
Considered together, these findings provide clues regarding the potential functions of rose TCP proteins. According to the RcTCP expression profiles after the drought, high-temperature, salt, powdery mildew, botrytis, and black spot treatments, five RcTCP genes (RcTCP2, 4, 7, 14, and 15) had significantly up-regulated or down-regulated transcription levels in response to the different treatments, suggesting they may contribute to the tolerance or resistance to various stresses. More specifically, RcTCP14 and RcTCP2 may have important roles in rose plant responses to abiotic stress and pathogenic fungi, respectively. The results of this study may form the basis of future investigations involving the functional characterization of RcTCP genes, which may lead to the genetic improvement of rose varieties and the enhancement of agronomic traits and resistance to environmental stresses.
-
This study provides a comprehensive analysis of the rose TCP gene family at the genome-wide level. Eighteen full-length RcTCP genes were screened and divided into two main groups, with highly similar motif compositions and exon-intron structures among genes belonging to the same group or subgroup. A phylogenetic analysis of the TCP transcription factors in several plant species provided valuable insights into the evolution of rose TCP genes. Furthermore, the importance of RcTCP genes in rose responses to abiotic stresses and fungal diseases was revealed on the basis of their expression patterns following various treatments. The phylogenetic and gene expression analyses clarified the functions of the RcTCP genes. The data presented herein will be useful for future research on the biological functions of individual TCP transcription factors in rose.
This work was financially supported by the Natural Science Foundation of the Jiangsu Higher Education Institutions of China (Item No. 21KJB210006), and the 'JBGS' Project of Seed Industry Revitalization in Jiangsu Province (JBGS [2021] 091), and the Jiangsu Forestry Science and Technology Innovation and Promotion Project (LYKJ [2021] 02).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Features of RcTCPs genes identified in Rosa chinensis.
- Supplemental Table S2 The sequence of primers used for qRT-PCR.
- Supplemental Fig. S1 TCP motif sequences in R. chinensis TCP proteins. (A) Sequence logos of the RcTCP protein domains. The sequence logos were produced by MEME on the basis of the alignment of the TCP domains. The information content of each position in the sequence is expressed as a bit score. (B) Amino acid sequence alignment of the R domain of the Class II RcTCP members in R. chinensis.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Cheng P, Bi D, Chen J, Zhao M, Wang Y, et al. 2023. Genome-wide identification and analysis of TCP transcription factor genes in Rosa chinensis in response to abiotic stress and fungal diseases. Ornamental Plant Research 3:3 doi: 10.48130/OPR-2023-0003 

Genome-wide identification and analysis of TCP transcription factor genes in Rosa chinensis in response to abiotic stress and fungal diseases
- Received: 03 August 2022
- Accepted: 14 December 2022
- Published online: 24 February 2023
Abstract: The TCP family genes encode plant-specific proteins with a variety of functions in multiple processes related to plant growth and environmental stress responses. However, there is little information about the response of rose TCPs to abiotic stresses and fungal diseases. Here, 18 non-redundant RcTCP genes distributed on six chromosomes were identified in Rosa chinensis. Phylogenetic analysis showed that RcTCP genes are divided into two main groups – Class I and Class II (CIN subclass and CYC/TB1 subclass). An analysis of the RcTCP promoters (2,000 bp) revealed that some cis-acting elements were associated with the changes to RcTCP expression due to environmental stress. The RcTCP expression patterns induced by abiotic stresses (drought, heat, and salinity) and fungal diseases (powdery mildew, botrytis, and black spot) were analyzed by RNA-seq or qRT-PCR. The transcription levels of five RcTCP genes (RcTCP2, 4, 7, 14, and 15) were significantly up-regulated or down-regulated under various treatment conditions, suggesting these genes may be involved in rose responses to diverse stresses. This study will be useful for future investigations of the biological functions of TCP transcription factors, especially during plant responses to environmental stimuli.
-
Key words:
- Rosa chinensis /
- TCP /
- Genome-wide /
- Expression patterns /
- Stress
{kind=link}