









-
The Brassica campestris (syn. Brassica rapa) ssp. chinensis is an economically important vegetable crop cultivated and researched worldwide. It is considered a rich source of multiple nutrients, such as vitamins, thioglycosides, carotenoids, and flavonoids[1,2]. However, little information is available about terpenes in Brassica campestris, especially terpene transport.
The genes encoding ABC proteins constitute a large gene family in plants that are categorized phylogenetically into eight subfamilies[3], and ABC transporters manage the active transport of a wide range of molecules across biological membranes, including heavy metals, secondary metabolites, and plant hormones[4−6]. The ABCG subfamily has a reverse domain including nucleotide-binding domains and trans-membrane domain (NBD-TMD) organization and plays a role in cellular efflux. In Arabidopsis, AtABCG25 and AtABCG40 function as plasma membrane abscisic acid (ABA) uptake transporters and play a role in the response to ABA[7]. CjABCB1/CjMDR1 is localized in the plasma membrane and catalyzes the import of berberine in Coptis japonica[8]. The ABCC transporters ZmMRP3 and ZmMRP4 are required for anthocyanin accumulation in maize kernels[9]. NtPDR1 and AaPDR3 participate in the transport of diterpenoids of Nicotiana tabacum and β-caryophyllene of Artemisia annua, respectively[10]. To date, there have been few studies on the transport of terpenoids in Brassica campestris.
Complete genome sequences of several organisms have revealed that genes encoding ABC transporters comprise a large gene family in plants; there are more than 131 members in Arabidopsis thaliana[11], 121 members in Oryza sativa[12] and 130 members in Zea mays[13]. The basic ABC transporter consists of four core structural domains, two transmembrane domains (TMDs) and two nucleotide-binding domains (NBDs), which provide a translocation pathway and hydrolyze ATP to provide the energy needed for ABC proteins, respectively[14].
In our previous study, it was found that β-caryophyllene content varies greatly among different varieties of Brassica campestris[15], but little is known about β-caryophyllene transport. In this study, we first performed genome-wide identification of ABC family members in Brassica campestris. Then, eight differentially expressed genes were screened using RNA-seq data, and their phylogenetic relationships, conserved motifs, and gene structure characteristics were characterized. Finally, the spatiotemporal expression patterns of eight ABC genes were explored in pak-choi. Overall, this work provides new insights into the transport of β-caryophyllene and its response to abiotic stress in Brassica campestris.
-
Brassica campestris, Suzhouqing and Xiangqingcai ('XQC' and 'SZQ'), used for transcriptome sequencing and Gas Chromatography-Mass Spectrometry (GC-MS) were harvested at 30, 60, 90, and 120 d after germination in a greenhouse located in Wu Jiang City, Jiang Su, China. At 60 d, 'XQC' samples were treated with 100 mmol·L−1 NaCl, 100 µmol·L−1 ABA, 100 µmol·L−1 MeJA, 4 °C/25 °C (low temperature, LT), and 35 °C/25 °C (high temperature, HT) in a growth chamber with light for 16 h per day and the relative humidity was 70%. Samples were collected from three seedlings and pooled together as one biological repeat at 0, 4, 8, 12, 24, and 48 h after three abiotic stresses and two hormone treatments. The collected leaves were immediately frozen in liquid nitrogen and stored at −80 °C for subsequent total RNA extraction and expression profiling. Three biological repeats were performed for each treatment at each time point.
Identification of ABC transporter genes from Brassica campestris
-
Information about ABC proteins in Arabidopsis thaliana was obtained as previously described[16] and used as a probe to execute an HMMER (
http://hmmer.org/ ) search of putative ABC proteins in the non-heading Chinese cabbage 001 (NHCC001) (Brassica campestris) reference genome[17]. After obtaining the full-length protein sequences, the putative Brassica campestris ABCs were verified with an additional online domain analysis tool, the Conserved Domain Database (CDD) (www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ).Phylogenetic analyses for ABC genes
-
Using the integrated tool MEGA7, protein sequence alignment for ABC members and AtABCs was carried out with the MUSCLE program, and a phylogenetic tree was then built with the maximum likelihood algorithm, wherein the bootstrap replicate value was set as 1000. Thereafter, the physicochemical properties of the ABC genes were analyzed with the online tool ExPASy (
http://www.expasy.org ).Motif, structure prediction, and cis-regulatory element analyses for ABC genes
-
The genome of 'NHCC001' was used as a reference genome, and the exon‒intron distribution of ABC genes was visualized with Tbtools. Cis-regulatory elements (CREs) in ABC promoters, which were defined as 2.0-kb genomic regions upstream of translational start codons, were detected with an online tool, PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ). The CREs were identified by PlantCARE and visualized with Tbtools. Motif analysis for ABC proteins was carried out using the Multiple EM Motif Elicitation (MEME) program (http://meme-Suite.org/ ), of which the maximum motif number was set as 10 under default mode.Sample preparation and GC–MS analysis
-
To further explore the content of β-caryophyllene in Brassica campestris, 2 g of fresh leaves was ground to a powder in liquid nitrogen. All samples were detected with a gas chromatograph (TRACE 1310, Thermo Scientific, America) coupled to a triple quadrupole mass spectrometer at 250 °C for 3 min in split mode (flow rate 1 mL/min). Three replicates of each treatment were performed. Subsequently, 2 g of the powder was transferred to a 20 mL headspace vial (Agilent, CA, USA) containing 2 mL of NaCl-saturated solution to inhibit the enzyme reaction. An 85-µm CAR/PDMS fiber was exposed to the headspace of the sample for 40 min at 35 °C for solid-phase microextraction (SPME) analysis[15].
RNA extraction and RNA-seq analysis
-
Total RNA was extracted from Brassica campestris using a plant RNA isolation kit (Mage, Beijing, China) according to the manufacturer's instructions for RNA library preparation and sequencing. After verification of the total RNA samples, the library was constructed with the MGIEasy RNA library. The library was constructed with three biological replicate samples at each time point. Raw reads were obtained from each sample after sequencing. After removing primer sequences, adaptor sequences, and low-quality reads, clean reads consisted of more than 80% base pairs with a Q-value ≥ 30. High-quality reads were compared with the spliced transcriptome. The transcripts per million (TPM) value was calculated for each transcription region to quantify expression abundance and variations, and differentially expressed genes (DEGs) were identified using DESeq2. DEG screening criteria were |log2FC| ≥ 1 and p < 0.05.
Expression analysis and subcellular localization of BcABC genes
-
Total RNA was extracted using a HiPure Plant RNA Mini Kit (R4151-02, MEGA) for qRT‒PCR. Then, the RNA was reverse transcribed to first-strand cDNA using the Reverse Transcription Kit (PrimeScriptTM RT Master Mix, Takara). The relative expression level at different treatments was calculated according to the 2−ΔΔCᴛ method. The tubulin gene was used as the reference gene. The complete CDs sequence of ABCG18 without a stop codon was subcloned into PJX003 to generate 35S::ABCG18 effector constructs. Agrobacterium solution containing PJX003-BcABCG18-GFP was injected into tobacco leaves. Green fluorescent protein (GFP) fluorescence (LSM780, Zeiss) was observed at 3 d after injection.
Statistical analysis
-
All data were analyzed using SPSS statistical software, and performed with three biological replicates as the average of the RNA-seq results or the average of the three biological repeats ± standard deviation (SD) for the qRT‒PCR. The differences between Bssraica campestris samples were evaluated by one-way ANOVA at a significance level of 0.05.
-
The morphology differed between Brassica campestris cultivars (XQC and SZQ) in shape, texture, and size of leaves at 30 d (Fig. 1a). The 'XQC' and 'SZQ' materials at 30, 60, 90, and 120 d were used to measure β-caryophyllene. The results showed that β-caryophyllene could be detected only in 'XQC', and the content of β-caryophyllene was highest at 60 d (Fig. 1b & c). The GC‒MS peak indicated a difference in the β-caryophyllene content in four stages (Fig. 1b).
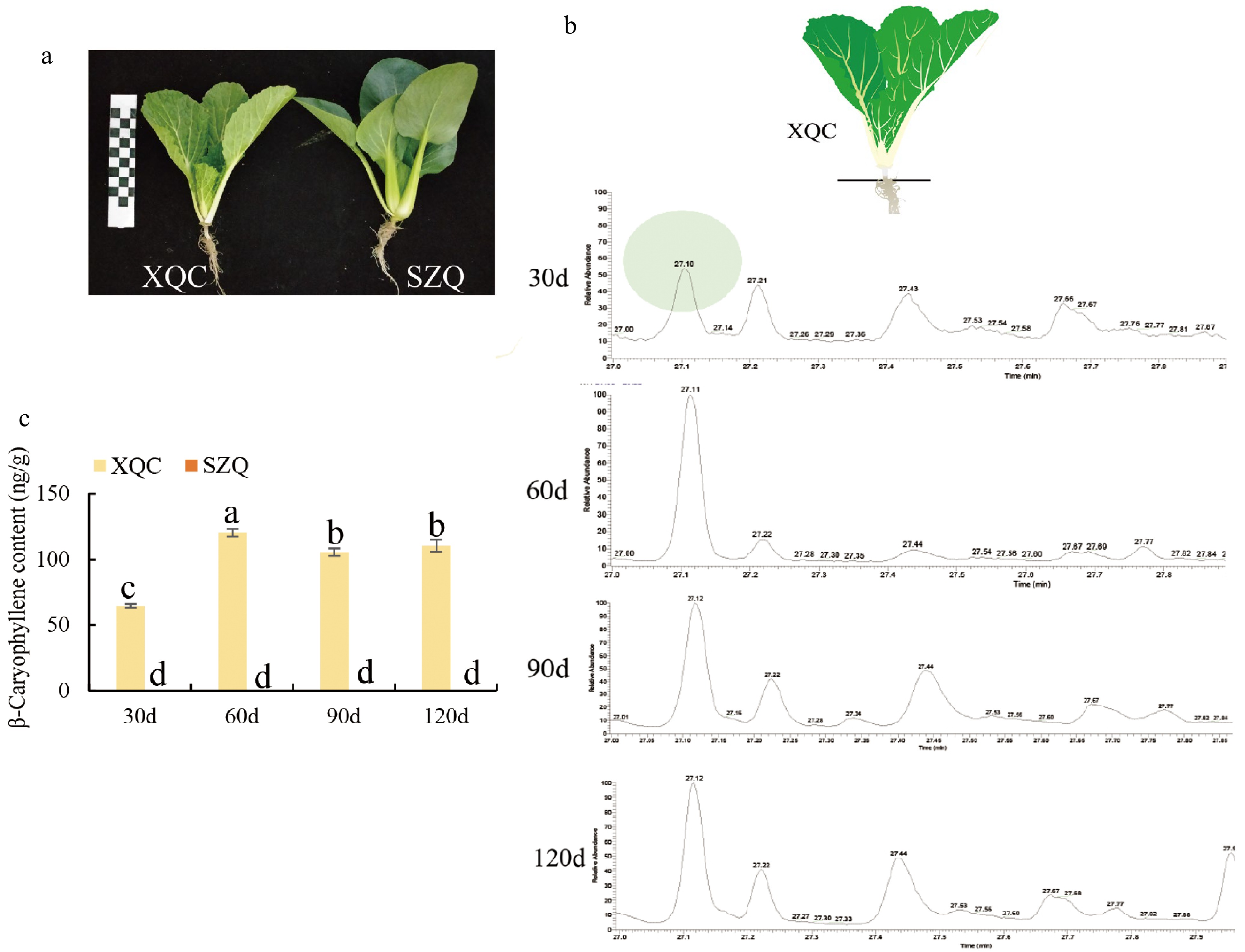
Figure 1.
β-Caryophyllene content in pak-choi cultivars 'XQC' and 'SZQ'. (a) Phenotypes of two pak-choi cultivars, 'XQC' and 'SZQ', at the 30 d stage. (b) Retention time of β-caryophyllene volatiles in GC-MS at 30, 60, 90, and 120 d. (c) β-caryophyllene content at 30, 60, 90, and 120 d in 'XQC' and 'SZQ'.
Identification of ABC transporter genes related to β-caryophyllene
-
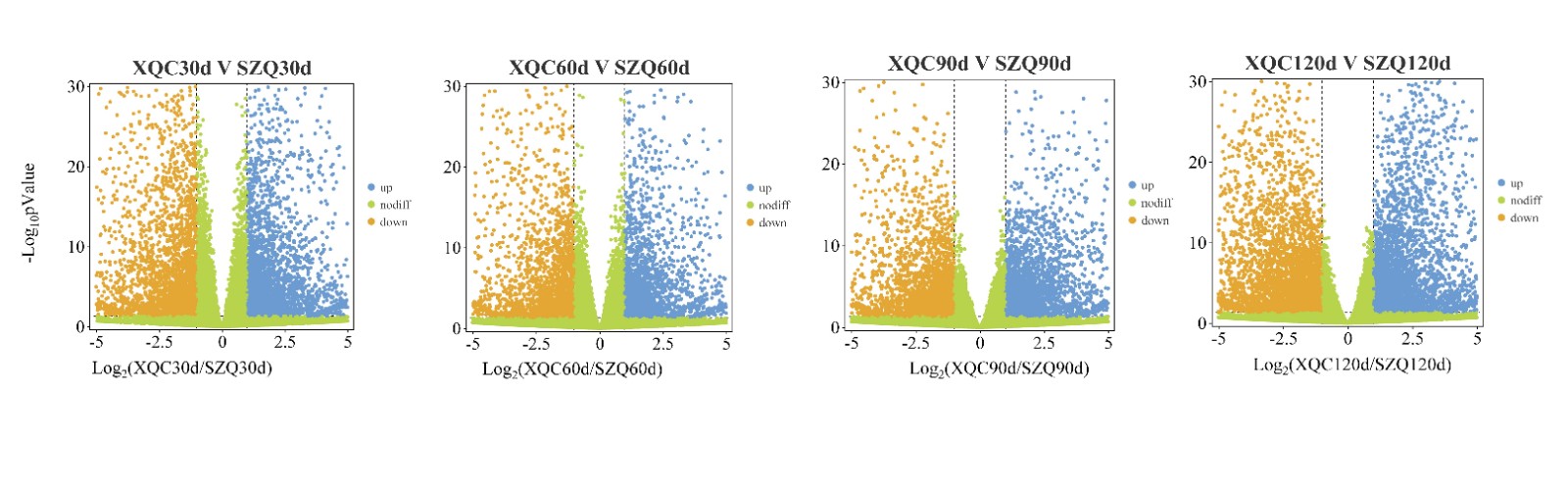
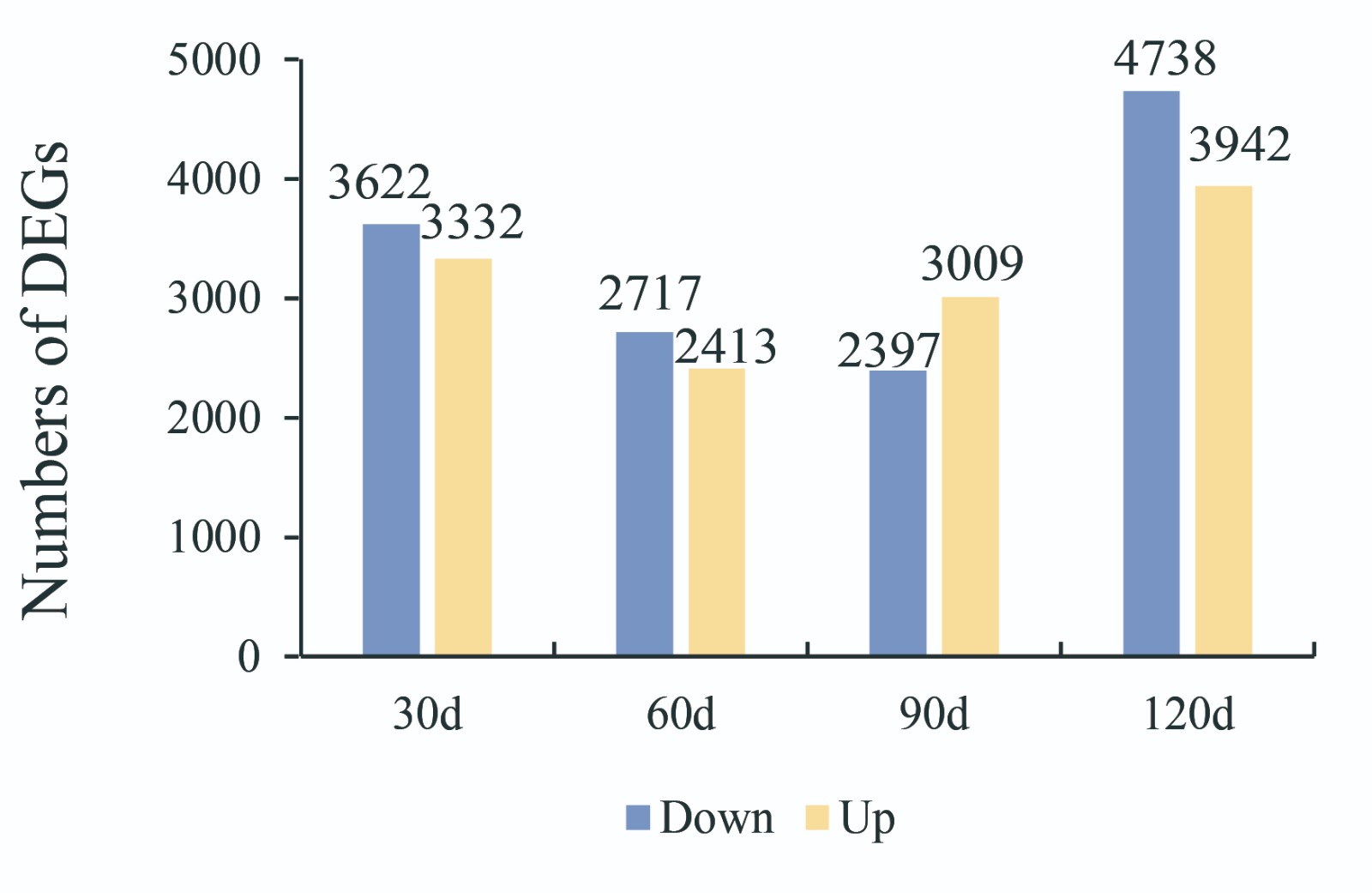
To investigate the transport of β-caryophyllene, transcriptome sequencing was carried out at 30, 60, 90, and 120 d after germination, with three replicates at each time point. From these samples, approximately 1.13 billion raw reads were produced. Following filtering, 1.12 billion high-quality-filtered (clean) reads proceeded to the next step, and the average read count for each sample ranged from approximately 40 to 61 million (Supplemental Table S1). The resulting clean reads were aligned against the Brassica campestris genome[17]. Differential expression results were generated using DESeq2, and gene expression was quantified using transcripts per million (TPM). Genes with expression |fold change| > 2 (p < 0.05) were considered to be differentially expressed, and they were visualized through volcano plots (Supplemental Fig. S1). In total, 6,954 (3,332 were upregulated and 3,622 were downregulated), 5130 (2717 were upregulated and 2413 were downregulated), 5406 (3009 were upregulated and 3,797 were downregulated), and 8680 (3942 were upregulated and 4738 were downregulated) differentially expressed genes (DEGs) were identified at 30, 60, 90, and 120 days, respectively (Fig. 2a & Supplemental Fig. S2). In this study, nine unigenes that encode enzymes associated with terpene biosynthesis were identified based on the enriched KEGG pathways and GO functional analysis. The terpene synthase gene (TPS), the last key enzyme in the synthesis of β-caryophyllene, is related to the content of β-caryophyllene. The correlation heatmap of DEGs involved in the terpene biosynthetic pathway shows the importance of the terpene synthase gene (Fig. 2b). Moreover, 426 transport protein genes were identified as DEGs, and among them, sixty transport proteins were identified with positive correlation coefficients with TPS21, including one equilibrative nucleotide transporter 3 (ENT3), two ABCs, one sulfate transporter 3.4 (SULTR3, 4), and one putative auxin efflux carrier component 5 (PIN5) (p > 0.8) (Fig. 2c & Supplemental Table S2).
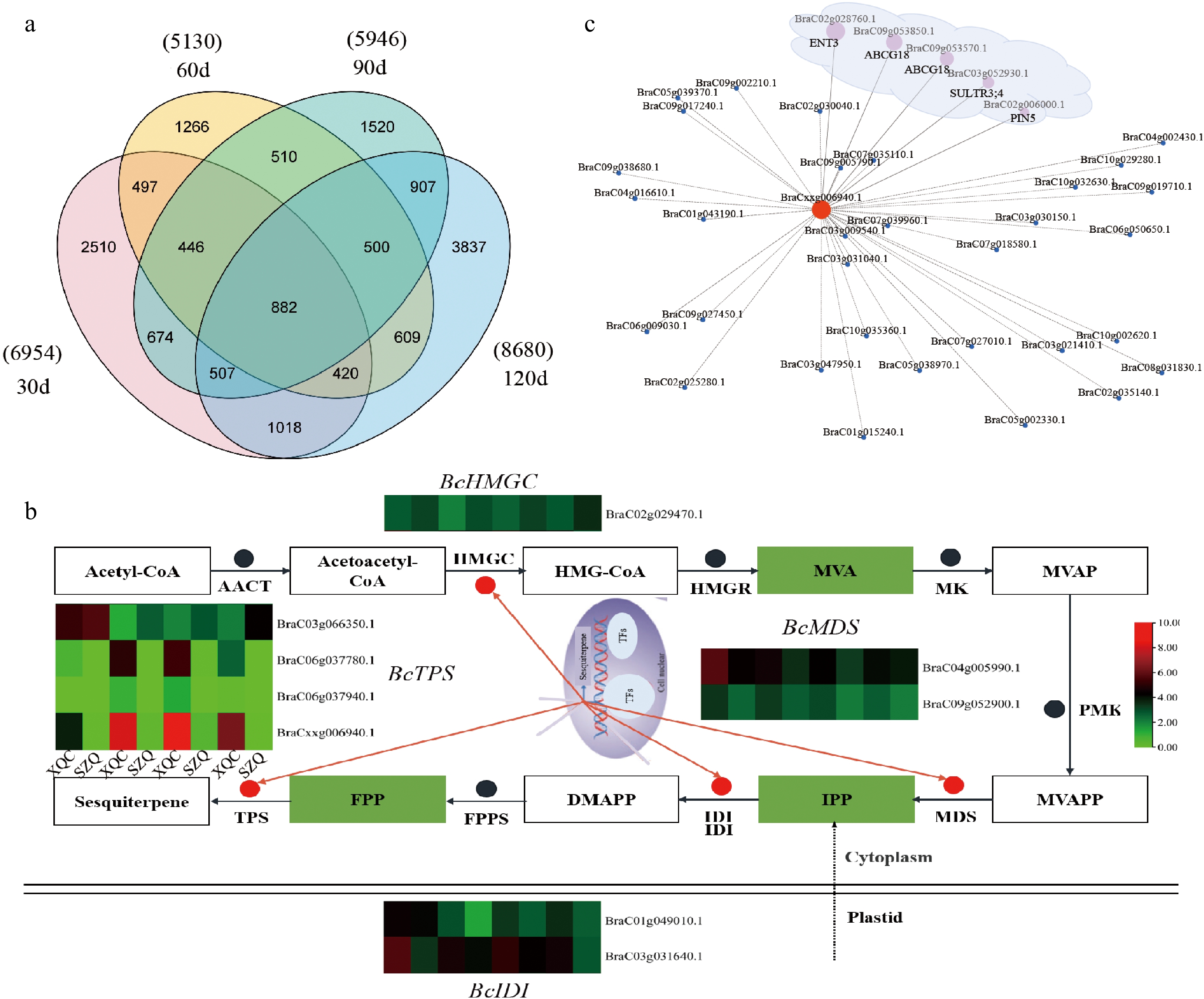
Figure 2.
Transcriptomic analysis of 'XQC' and 'SZQ' at 30, 60, 90, and 120 d. (a) Venn diagram showing the number of DEGs identified between 'XQC' and 'SZQ' at 30, 60, 90, and 120 d. (b) DEG heatmap of the RNA-Seq expression involved in the sesquiterpene pathway. (c) Network between TPS21 and the differential transporters. Circle size indicates Person correlation.
Structures and motifs of BcABC genes
-
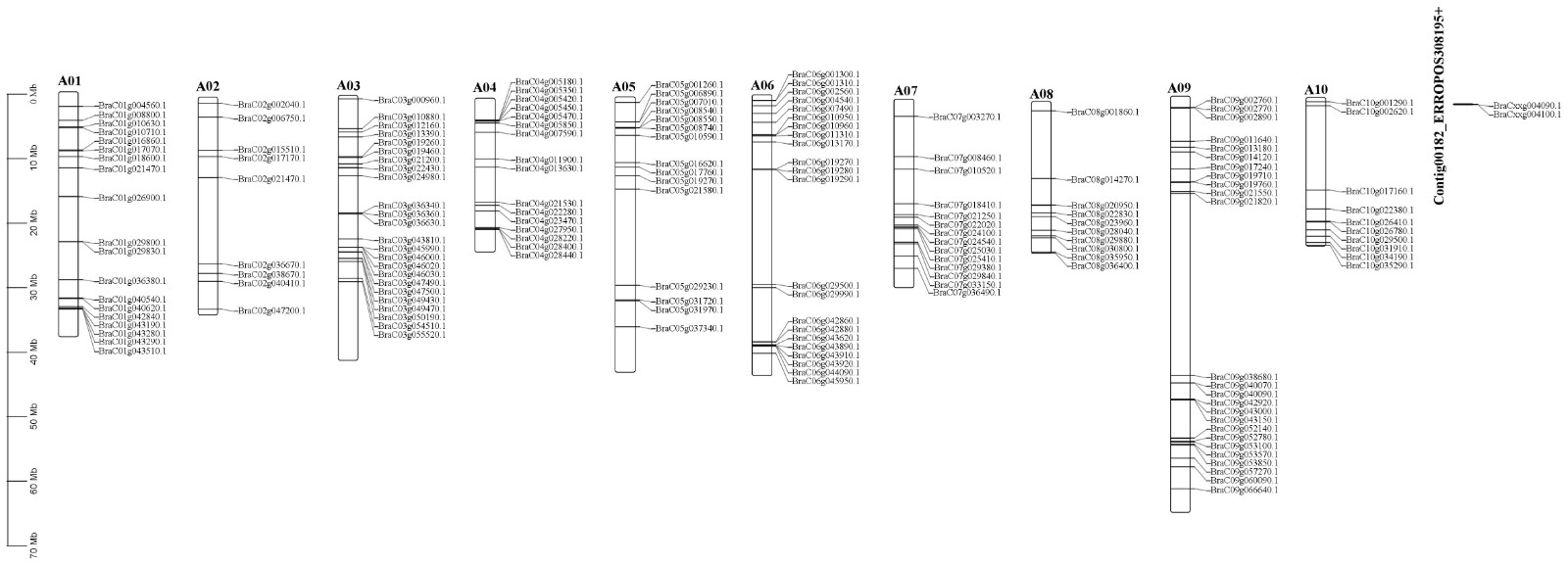
The ABC transporter, which is highly correlated with TPS21, is thought to be an important carrier for the transport of metabolites in plants. According to the genome, a total of 169 ABC proteins, listed in Supplemental Table S3, were recognized as members of the Brassica campestris ABC family, and their physical locations are shown in Supplemental Fig. S3 in the Brassica campestris genome. The phylogenetic tree was constructed with ABC proteins from Brassica campestris and Arabidopsis, and proteins of Arabidopsis are listed in Supplemental Table S4. Phylogenetic analysis of ABC proteins from Brassica campestris and Arabidopsis indicated that among the eight subfamilies, subfamily G had the most genes, with 55, followed by subfamily B, with 37 (Fig. 3a). In order to identify the role of β-caryophyllene-related transporters and analyze ABC transporters in abiotic stress, seven differential ABC transporters were identified through the transcription data of 30, 60, 90, and 120 d of material grown in greenhouses. Meanwhile, according to a previous study, the ABCG33 gene may respond to stress. Eight ABC genes, five members belonged to ABCG, two members belonged to ABCC, and only one member belonged to ABCE. Eight ABC proteins were named according to homology (BraC05g008540.1, ABCG33; BraC05g010590.1, ABCC2; BraC05g037340.1, ABCG29; BraC06g010960.1, ABCG35; BraC08g014270.1, ABCE2; BraC08g023960.1, ABCC1; BraC09g053570.1, ABCG18; BraC10g031910.1, ABCG22). The results showed that eight ABC proteins were divided into four subgroups. The first was composed of two BcABCs (BraC08g23960.1; BraC05g10590), the second consisted of BraC08g14270, the third consisted of three BcABCs (BraC05g008540.1; BraC05g037340.1; BraC06g010960.1), and the fourth consisted of two BcABC (BraC10g031910.1; BraC09g053570.1) (Fig. 3b-X). Using the MEME program, we further performed motif prediction for ABC proteins, and 10 different motifs (motifs 1–10) were identified (Fig. 3b). Motif 1 was identified in all ABC proteins (Fig. 3b). Structural features were then characterized for eight ABC genes, and a large divergence in exon number was observed (Fig. 3b-Z). The length, molecular weight, and theoretical isoelectric point of eight ABC proteins ranged from 605 to 1626 aa with an average of 540 aa, from 68 to 182 kDa with an average of 135 kDa, and from 5.9 to 9.37 with an average of 7.39, respectively (Supplemental Table S5). To understand their transcriptional regulation patterns, CREs were identified in ABC gene promoter regions. A total of 16 CREs, namely, anaerobic induction-, meristem expression-, abscisic acid-responsive element-, anoxic inducibility-, circadian control-, endosperm expression-, gibberellin responsive element, light responsive element, low temperature responsive element, MeJA responsive element, MYB binding site, MYBHv1 binding site, MYC binding site, salicylic acid responsive element, sugar metabolism and plant defense signaling, and wound responsive element types, were identified (Fig. 3c). Light-responsive elements were detected in eight ABC promoters, and MeJA-responsive elements were detected in six ABC promoters except for ABCG33 and ABCG18. In addition to ABCE2, the ABA motif was detected in seven ABC promoters.
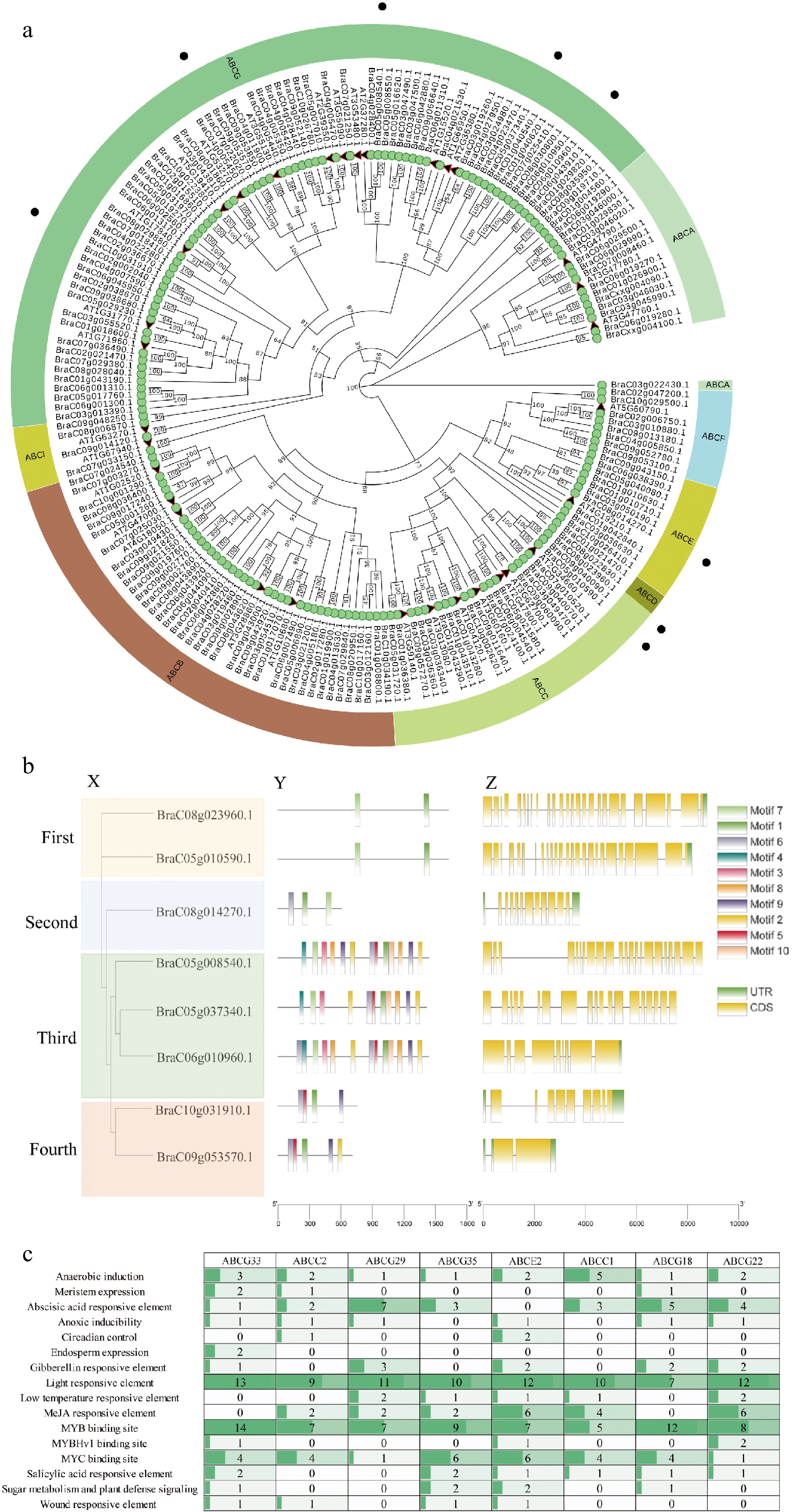
Figure 3.
Phylogenetic analysis, gene structure, conserved domains, and cis-acting elements of ABC proteins from Brassica campestris and Arabidopsis. (a) Phylogenic relationship of the identified ABC genes in the Brassica campestris genome. (b) Eight ABC transporter motif distributions and exon-intron distribution in Brassica campestris. The motifs identified in the BcABC proteins are indicated by different colored boxes and named motifs 1–10. The closed yellow boxes and black lines represent exons and introns, respectively. (c) The number and type of existing putative cis-acting elements in two kb upstream regions of the eight ABC genes.
Transcriptomic analysis and β-caryophyllene content analysis
-
The expression levels of the seven transporters (except ABCG33) in different stages of development were observed, and the results indicated that they could be classified into two groups, and ABCC1 and ABCG18 were clustered with TPS21 (Fig. 4a). We further analyzed the correlation between seven ABC transporters and TPS21 and found that ABCG18 was significantly correlated with TPS21 (p > 0.85) (Fig. 4b). A potential regulatory network of ABCG18 is shown in Fig. 4c, and MYB12 is the most closely related to ABCG18 and may be involved in the transcription of ABCG18 (Fig. 4c).
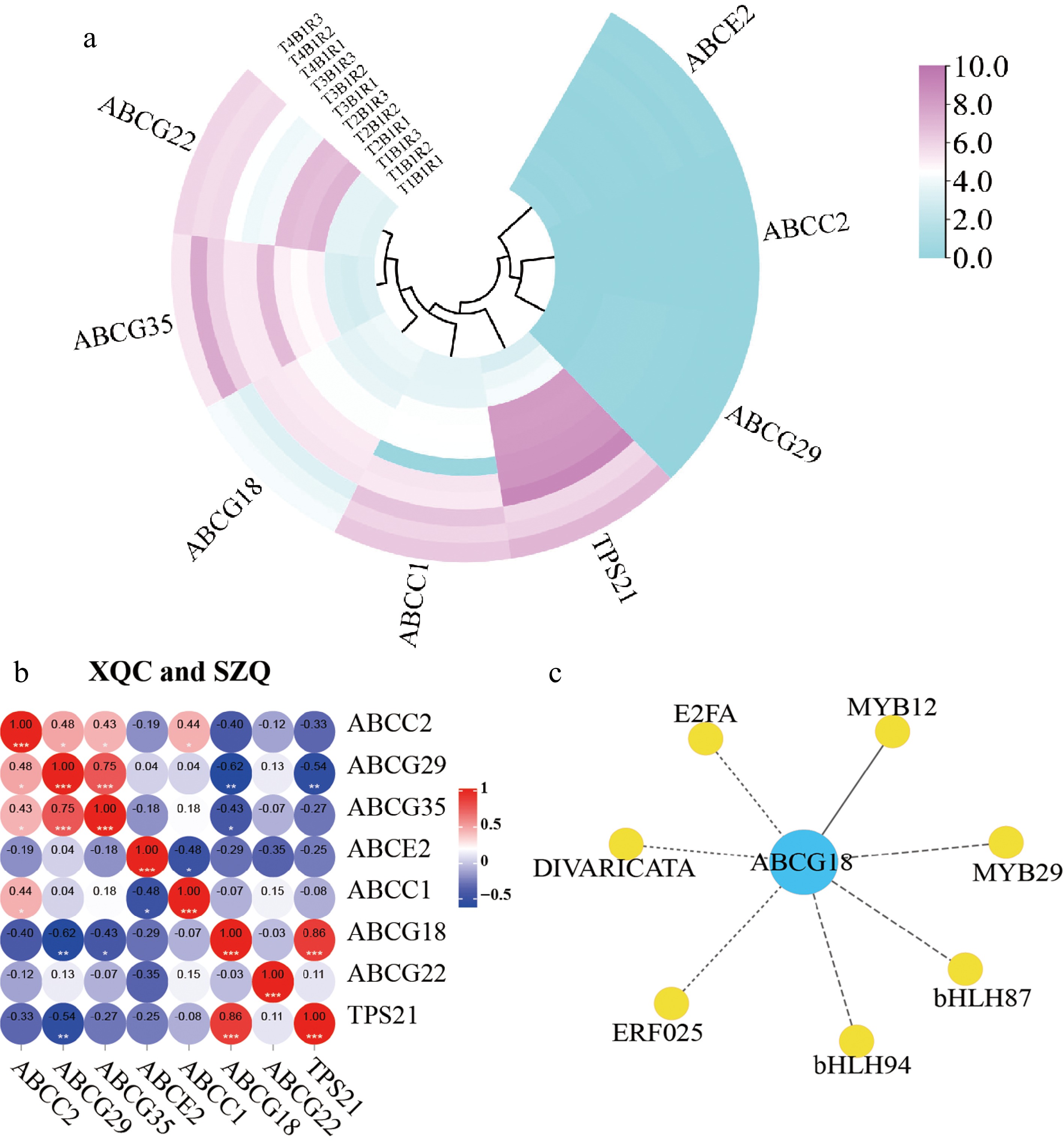
Figure 4.
Pearson analysis of 'XQC' and 'SZQ' at 30, 60, 90, and 120 d. (a) Heatmap of RNA-Seq expression of seven ABC differential transporters. (b) Correlation among the ABC proteins in 'XQC' and 'SZQ'. (c) The correlation between transcription factors and ABCG18 of 'XQC' and 'SZQ' transcriptome data, solid line -> 0.8.
Expression profiles of the ABCs and subcellular localization
-
To cope with environmental changes, the transport of materials in plants is often dependent on carriers. We further examined the expression profiles of eight ABC genes and TPS21 using qRT‒PCR under three kinds of abiotic stress and two hormonal treatments (Fig. 5a). After 12 h of MeJA or ABA treatment, two ABCs (ABCG33 and ABCC1) were significantly upregulated. Interestingly, the ABCC1 transcription level had a similar expression pattern to TPS21 under all five treatments. Under low temperature, ABCG22 and ABCG35 increased markedly, while ABCG22 and ABCG35 decreased under the other treatments. Our results show that TPS21 is expressed in two ways: one is first increased and then decreased and was lower than CK under salt stress, low temperature treatment and ABA treatment, and the other is always higher than CK under MeJA and high temperature treatment. Furthermore, TPSa21 and ABCG18 transcription patterns were investigated in different tissues (Fig. 5b). TPS21 was mainly expressed in the leaf, while the expression of TPS21 in the root and petiole was almost zero. ABCG18 was mainly expressed in leaves and petioles but not in roots. These results suggest that ABCG18 may play an important role in the transportation of β-caryophyllene under normal conditions, while ABCG35 may be an important transport protein under low temperature. BraC09g053570.1 had the best homology with ABCG18 (AT3G55110.1) (Fig. 5c). The membrane localization ABCG transporter plays a key role in the export of β-caryophyllene and other secondary metabolites. A green fluorescent protein-fused ABCG18 reporter gene was constructed and used for transient transformation experiments in tobacco leaves, and it was found to be a membrane localization protein (Fig. 5d).
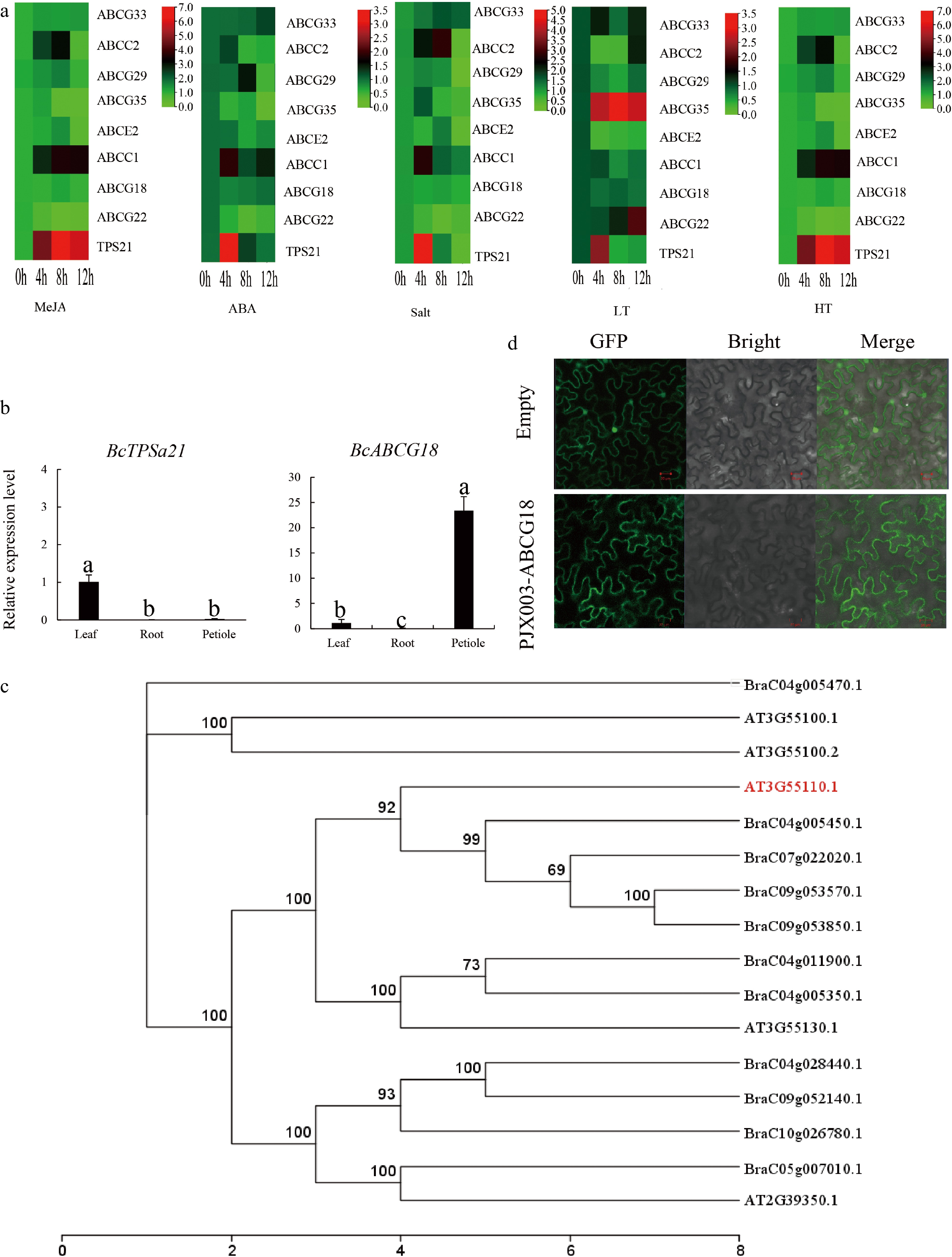
Figure 5.
Expression profiles of BcABC genes and subcellular localization. (a) Expression profiles of BcABC genes under different conditions (MeJA, ABA, salt, high temperature, low temperature). (b) Transcript levels of ABCG18 and TPS21 in leaf, stem, and root tissue. (c), (d) Finding the best homology of the ABCG18 gene and subcellular localization.
-
Plants are frequently exposed to various stresses that affect their growth and development. To survive, plants have evolved sophisticated response systems to cope with a variety of stressful environments. Among these responsive networks, ABC-mediated transport is a representative system that has attracted increasing attention from researchers. ABC transporters are a family that can take part in the transport of a broad range of terpenoids. Genome-wide analyses of ABC transporters in Solanum lycopersicum[18], Arabidopsis[16], Oryza sativa[12], Zea mays[13], and Brassica rapa[19] have already been performed. However, little is known about ABC transporters related to β-caryophyllene in Brassica campestris. To address this, we performed comparative transcriptome analysis in Brassica campestris to investigate the potential function of the ABC transporter in β-caryophyllene transport and response to abiotic stress in Brassica campestris.
Terpenoids show a wide array of pharmacological activities and are one of the most extensively studied and structurally diverse classes of natural compounds. Terpenoids can be classified based on the number of isoprene units in the parent structure, such as monoterpenoids (C10), sesquiterpenoids (C15), diterpenoids (C20), sesterterpenoids (C25), triterpenoids (C30), tetraterpenes (C40) and polyterpenes. AaPDR3 (ABCG40) is involved in sesquiterpene beta-caryophyllene transport in Artemisia annua[10]. This study indicated that TPS21 was one of the most important enzymes in the synthesis of caryophyllene (Fig 2d). Five highly correlated transporters with TPS21 were identified (p > 0.8), including nucleotide transport and ABC transporters. Thus, ABCG18 may become a potential caryophyllene transporter according to correlation. Our results indicate that MYB12 is a potential transporter regulating ABCG18. MYB12 is involved in the regulation of metabolism of various substances. High temperature enhances anthocyanin coloration in Asiatic hybrid lily flowers via upregulation of the MYB12 positive regulator[20]. COP1 is involved in the dark-dependent repression of MYB12 expression and flavonol accumulation[21]. Under abiotic stress conditions, ABCG17 and ABCG18 are transcriptionally repressed, promoting active ABA movement and response[22]. ABA homeostasis under normal and abiotic stress conditions can be regulated by ABCG18 protein. This is similar to our results with ABA treatment and provides a new perspective for the study of caryophyllene transport. WGDs (whole genome duplications) are strongly supported by evidence commonly found in many species-rich lineages of eukaryotes and thus are considered a major driving force in species diversification. In Arabidopsis, a total of 129 ABC transporter genes were identified. This study showed that a total of 169 annotated genes were identified as Brassica campestris ABC family members, which was more than that in Arabidopsis but similar to that in Brassica rapa (Chiifu-401)[19].
Evolutionary analysis showed that seven subfamilies of Brassica campestris and Arabidopsis ABC transporters (ABCA, ABCB, ABCC, ABCD, ABCE, ABCF, ABCG, and ABCI subfamilies) were generated. ABCG had the most genes in subfamilies. This phenomenon was consistent with observations in Brassica rapa and Arabidopsis[19]. Eight differentially expressed ABC protein genes were screened based on transcriptome data, and eight ABC proteins were predicted to be located in the plasma membrane, which may be related to their transport functions. Variations in structure or motifs can be relatively reliable parameters to evaluate the evolution of a gene family. In this study, eight ABC proteins were classified into four groups, and similar gene structures and motifs were commonly shared by most members in the same group.
Functional conservation and diversification can also be uncovered to a great extent by the expression profiles. It is clear that the expression of many ABCs in different plant species can be stimulated by various hormones or stresses, such as ABA, methyl jasmonate (MeJA), and low temperature[23−25]. Fifteen differentially expressed ABC genes exhibited diverse expression responses to stress treatments, including drought, cadmium, and salt stress[26]. We investigated the temporal and spatial expression profiles of these ABC transporters in different developmental stages and under different treatments. Expression analysis results showed that the transcription of ABCs was differentially regulated by various abiotic stresses and hormone treatments. ABCC1 is a vacuolar membrane-localized transporter; ABCC is responsible for the transport of the phytochelatins (PCs) and cadmium (PCs–Cd) complex into the vacuoles[27]. Notably, ABCC1, which is a membrane-bound, energy-dependent efflux transporter, belonging to the superfamily of ABC transporters, has a similar expression pattern under MeJA treatment and low temperature. Moreover, ABCG22 and ABCG35 were significantly induced under low temperature stress. The effect of ABCG35 at low temperature is still unclear, our results indicated that it might be a key gene for participating in low temperature.
In conclusion, this study provides a genome-wide analysis of ABC transporters from the Brassica campestris genome. The expression profiles of eight ABC transporters were validated using both RNA-seq analyses and qRT‒PCR. Furthermore, this study provides insights into ABC transporter gene expression after exposure to stress. These combined results may offer a basis for future studies on the transport of β-caryophyllene by ABC transporters in Brassica campestris.
-
Taken together, this study represents the characterization of eight different ABC gene spatiotemporal expression patterns according to the genome database and transcriptome of Brassica campestris for the first time. Our results indicate that ABCG18 is a potential carrier of β-caryophyllene, which might participate in substance transport in leaves and petioles, and MYB12 may be involved in the regulation of ABCG18 in Brassica campestris. ABCG35 and ABCG22 may be important for low temperature. These results are helpful for understanding the mechanism of β-caryophyllene transport metabolism in Brassica campestris and provide a theoretical basis for further research on the physiological function of ABC transporter genes in Brassica campestris.
This work was supported by the Jiangsu Seed Industry Revitalization Project [JBGS (2021)015], the National vegetable industry technology system (CARS-23-A-16), the National Key Research and Development Program (2018YFD1000805), and a project funded by the priority academic program development of Jiangsu higher education institutions (China).
-
Dr. Ying Li is the Editorial Board member of journal Vegetable Research. She was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of the Editorial Board member and her research group.
- Supplemental Fig. S1 Volcano plot of DEGs (differentially expressed genes) between ‘XQC’ and ‘SZQ’ at 30, 60, 90, and 120 days.
- Supplemental Fig. S2 The number of DEGs in different comparison groups. Oranges indicate upregulation, and blue indicates downregulation.
- Supplemental Fig. S3 Distribution of ABC genes on Brassica campestris chromosomes.
- Supplemental Table S1 RNA-seq data used in this study.
- Supplemental Table S2 425 transport proteins ID.
- Supplemental Table S3 169 ABC protein sequence in Brassica campestris.
- Supplemental Table S4 ABC proteins of Arabidopsis.
- Supplemental Table S5 Basic information on ABC genes identified in Brassica campestris.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang H, Zong C, Bai Y, Yuan S, Li Y, et al. 2023. Comparative transcriptome-based identification and expression analysis of ATP-binding cassette (ABC) transporters reveals a likely role in the transport of β-caryophyllene and response to abiotic stress in Brassica campestris. Vegetable Research 3:13 doi: 10.48130/VR-2023-0013 

Comparative transcriptome-based identification and expression analysis of ATP-binding cassette (ABC) transporters reveals a likely role in the transport of β-caryophyllene and response to abiotic stress in Brassica campestris
- Received: 21 January 2023
- Accepted: 13 March 2023
- Published online: 18 April 2023
Abstract: β-caryophyllene has physiological activities such as antibacterial, antiedemic, anti-inflammatory, antitumor, and fungicidal. Previous research has focused on the synthesis of β-caryophyllene, however little information is available about its involvement in β-caryophyllene transport. ATP-binding cassette (ABC) transporters are involved in the transport of multiple substrates such as amino acids, terpenes, and heavy metals. Herein, we present a genome-wide comparative transcriptome analysis between non-β-caryophyllene cultivars ('SZQ') and high β-caryophyllene cultivars ('XQC') to identify potential ABC functional gene activities that occur during plant development. This article analyzes potential ABC transporters that may transport β-caryophyllene by comparative transcriptome and GC-MS, and explores the genetic structure, evolutionary relationship, cis-acting element analysis, and transcription patterns of these transporters under different treatment conditions. Phylogenetic and cis-acting element analysis indicated that eight genes in 'SZQ' and 'XQC' belonged to the ABCC, ABCG, and ABCE subfamilies and had the highest number of light-responsive elements and MYB binding sites. The RNA-seq and qRT-PCR results showed that eight ABC genes were expressed in a tissue- and development-specific manner, while their abundance apparently varied when plants were exposed to different treatment conditions. Pearson correlation analysis showed that ABCG18 is a potential carrier of β-caryophyllene, which might participate in substance transport in leaves and petioles. In summary, these observations open up new future research directions for β-caryophyllene transport and provide insight into ABC family genes in Brassica campestris.
-
Key words:
- β-caryophyllene /
- ABC proteins /
- Expression profile /
- Transport
{kind=link}
{kind=link}
{kind=link}