









-
Hydrogen sulfide (H2S) has been identified as a new gasotransmitter after NO and CO, and it plays multiple physiological roles in all living organisms. Accumulating evidence unveiled that H2S participates in seed germination, root morphogenesis, stomatal movement, and photosynthesis[1]. Moreover, H2S serves as a signal to enhance plant acclimation to various abiotic and biotic stresses[2, 3]. H2S could be generated during sulfur assimilation and cysteine decomposition. In sulfate assimilation pathway, H2S is produced mainly through sulfite reductase (SiR), then sulfide is integrated into the first organic sulfur-containing molecule cysteine by O-acetylserine thiol lyase (OAS-TL)[4]. In the other route, H2S is generated from L-cysteine by the catalyzation of L-cysteine desulfhydrase (L-CDes), or D-cysteine by D-cysteine desulfhydrase (D-CDes)[5−7]. Exploration of endogenous H2S-producing enzymes in plants dates back to the 1960s, and after decades of exploration, D-cysteine desulfhydrase was found in Escherichia coli, where it decomposes D‐cysteine into pyruvate, H2S and ammonium[8]. At present, DCD has been found in the study of a variety of plants such as Arabidopsis, Spinach, Chlorella, Zucchini and Tobacco, where it can only use D-Cys as the specific substrate instead of L-Cys[5, 9] A previous study suggested that Cd-induced WRKY13 activates the expression of AtDCD, increasing the production of H2S, thereby improving Arabidopsis tolerance to Cd[10]. And the SlDCD2 mutant exhibited higher ethylene content, enhanced chlorophyll degradation and increased carotenoid accumulation. Additionally, the expression of multiple ripening-related genes, including NYC1, PAO, SGR1, PDS, PSY1, ACO1, ACS2, E4, CEL2 and EXP was enhanced during the dcd2 mutant tomato fruit ripening.[11] In addition, DCD could also improve the ability of Eruca sativa to respond to drought stress[12]. However, whether DCD is involved in biotic stress response in tomato remains to be further studied.
Over the past decades, the role of various types of sulfur-containing compounds in plant defense and resistance to microbial pathogens has been widely discovered[13], and in addition to the recognized role of glutathione and cysteine, the role of H2S in plant disease resistance has gradually been confirmed. Exogenous H2S protected pear fruit from the infection of the pathogens Aspergillus niger and Penicillium expansum, suggesting that H2S could be developed as an effective fungicide for postharvest storage[14]. H2S fumigation was found to alleviate the decay symptoms of peach fruit inoculated with Pseudomonas and Monilinia fructicola by inhibiting spore germination and hyphal development of Pseudomonas and M. fructicola[15]. Also, H2S application reduced the rotten rate of tomatoes, citrus, apples and kiwifruit inoculated with A. niger and Italian penicillium through disturbance on spore germination and hyphal elongation of the pathogens[16]. H2S donor NaHS significantly inhibited Botryosphaeria Dothidea mycelial growth and enhanced the disease resistance of kiwifruit after harvest[17]. Previous research indicated that the transcription levels of LCD and DCD1 in Arabidopsis increased significantly after 6 h of treatment with Pseudomonas, and the production of endogenous H2S increased by 1.2−1.3 times. Overexpression of AtLCD and AtDCD1 showed increased endogenous H2S production and enhanced resistance to Pst DC3000 increase, while treatment with taurine (H2S scavenger) resulted in decreased resistance to the pathogen, suggesting the potential role of H2S in biotic stress response[18].
Plants have been endowed with sophisticated barriers to prevent pathogen invasions[19]. Accumulation of reactive oxygen species (ROS) has been observed in a wide range of plant-pathogen interactions[20−22]. For instance, after inoculation with B. cinerea, rapid increase of ROS was found around the penetrated cell wall as well as in the plasma membrane[23]. In the process of maintaining the homeostasis of ROS, the credit of antioxidant enzymes are indispensable[24]. B. cinerea, which is a necrotrophic pathogen, prefer dead cells for nutritional purposes, and thus tissue necrosis caused by ROS during pathogen infection increased plant susceptibility to necrotrophic. Therefore, antioxidant capacity of plants including antioxidative enzymes and molecular antioxidants would be motivated to scavenging excessive ROS. To alleviate necrotrophic pathogens induced ROS stress, increased activity of antioxidant enzymes superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT) were widely observed[15]. Accumulating reports suggested that H2S acts as a signal to alleviate postharvest senescence of multiple fruits and vegetables by maintaining balanced ROS homeostasis through activating antioxidant enzymes. Thus, it is speculating that H2S might attenuate the accumulation of ROS. However, whether and how endogenous H2S interferes with ROS metabolism during tomato infection by B. cinerea is still unknown. In the present study, the gene SlDCD1 encoding D-cysteine desulfhydrase 1 in tomato was mutated by CRISPR/Cas9, and the effect of SlDCD1 mutation on plant susceptibility and ROS metabolism to B. cinerea was evaluated. Besides, the difference in plant response to B. cinerea infection during different fruit ripening stages at green, breaker or red were investigated.
-
Tomato (S. lycopersicum, Micro Tom) plants were cultured under the following conditions: 16 h day/8 h night cycle, 25 ± 2 °C/20 ± 2 °C day/night temperature, 65 % relative humidity, and 250 μmol·m−2·s−1 light intensity.
B. cinerea was maintained on potato glucose agar medium in the dark at 25 °C. Conidia of B. cinerea strain were harvested as described by Asselbergh et al.[25]. The conidial suspension was centrifuged for 10 min at 10,000 g. After removal of the supernatant, the conidia were resuspended in inoculation buffer (containing 16.7 mM KH2PO4 and 25 mM glucose) at a concentration of 106·mL−1. Conidia pregerminated for 2 h in the inoculation suspension at 22 °C. Fifty μL conidia suspension was injected into the flesh of tomato fruit at mature green, breaker or red stages, and the leaves were infected by vacuuming (0.8 kg·cm−2, 1 min). Subsequently, fruit and leaves were stored on wet sterile filter papers in petri dishes at 23 °C for 5 d.
Determination of the amount of H2S release in tomato leaves and gray intensity analysis
-
As mentioned previously[26], the release of H2S in 0.2 g of tomato leaves was determined using lead acetate test strips (cat. number WHA2602501A, Sigma, Darmstadt, Germany). The amount of H2S release is measured according to the color of zinc acetate test strips. Gray intensity analysis of the zinc acetate test strips was performed using ImageJ software.
Phylogenetic analysis
-
Putative D-cysteine desulfhydrase proteins in S. lycopersicum, A. thaliana, V. vinifera, P. patens, N. tabacum, O.sativa, T. aestivum, P. bretschneideri, Z. mays, C. annuum, and M. acuminata were obtained by the BLASTP tool in the NCBI (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ) database with AtDCD1 (NP_001319174.1) as a query. The amino acid sequences of SlDCD1 (NP_001234368) and SlDCD2 (XP_004228490.1) from S. lycopersicum; AtDCD1 and AtDCD2 (NP_001327499.1) from A. thaliana; VvDCD1 (XP_002263358.2) and VvDCD2 (XP_002282104.1) from V. vinifera; PpDCD1 (XP_024396726.1) and PpDCD2 (XP_024361159) from P. patens; NtDCD1 (XP_016466777.1) and NtDCD2 (XP_016500459.1) from N. tabacum; OsDCD1 (XP_015626189.1) and OsDCD2 (XP_015621767.1) from O.sativa; TaDCD1 (XP_044421034.1) and TaDCD2 (XP_044357483.1) from T. aestivum; PyDCD1(XP_009349823) and PyDCD2 (XP_048427768.1) from P. bretschneideri; ZmDCD1 (NP_001130254.1) and ZmDCD2 (NP_001353762.1) from Z. mays; CaDCD1 (XP_016563957.1) and CaDCD2 (XP_016577814.1) from C. annuum; and MaDCD1 (XP_009417666.1) and MaDCD2 (XP_009409959.1) from M. acuminata were selected to construct a phylogenetic tree by the neighbor-joining method according to the parameters previously reported by Saitou & Nei[27].Generation and genotyping of the dcd1 mutant by CRISPR/Cas9
-
CRISPR/Cas9 mutagenesis of DCD1 in tomato was performed as previously described[28]. The primers for sgRNA are listed in Supplemental Table S1. For confirmation of the dcd1 mutant, we amplified a fragment of the sgRNA target sequence using genomic DNA from the dcd1 mutant. The amplified fragment was further used for DNA sequencing, and the genotyping of tomato plants was analyzed on the website DSDecodeM (
http://skl.scau.edu.cn/dsdecode/ )[29].Visualization of defense responses of tomato to B. cinerea
-
To compare the defense responses of wild-type (WT) and dcd1 leaves, trypan blue staining was used to detect necrosis of tomato leaf cells infected with B. cinerea[30]. Leaves were incubated in petri dishes containing staining solution (containing 10 mL 85% lactic acid, 10 mL saturated phenols, 10 mL glycerol, 10 mL distilled water, 0.4 g trypan blue) at 25 °C for 1 h and then were decolorized with alcohol at 25 °C for 12 h. The distribution of H2O2 in tomato leaf cells infected with B. cinerea was detected by DAB (3,3'-Diaminobenzidine) staining[30]. The leaves were soaked in staining solution (containing 0.5 g DAB, 25 μL TWEEN-20, 2.5 mL 200 mM Na2HPO4, 45 mL H2O, pH 3.0) and then vacuumed 2−3 times for 1 min at 0.8 kg·cm−2. Then chlorophyll was removed using ethanol, and plant leaves were photographed.
RNA extraction and RT-qPCR
-
RNA was extracted from 0.2 g leaf or fruit and the first strand cDNA was synthesized following the method reported previously[31]. Tomato Tubulin was used as an internal reference. Gene-specific primers for RT-qPCR are listed in Supplemental Table S1. The injected fruits or leaves were sampled within 10 mm diameter of the lesion and RNA was extracted, the actin gene transcript levels of B. cinerea were used as an indicator of B. cinerea growth[30].
Assay of antioxidant enzyme activities
-
Tomato tissue (0.5 g) was extracted using 10 mL of 50 mM phosphate buffer (pH 7.8) at 4 °C. Then samples were centrifuged at 10,000 g and 4 °C for 15 min. Supernatant is the crude antioxidant enzyme solution[32]. CAT, APX, SOD and POD (peroxidase) activity were measured and calculated spectrometrically[33−35]. An absorbance increase of 1.0 × 10−5 OD470 nm·min−1 was considered 1 U of POD activity, a decrease in absorbance of 1.0 × 10−3 at OD240 nm·min−1 was considered 1 U of CAT activity, the amount used to inhibit 5% of the photochemical reduction of NBT was considered 1 U of SOD activity, and a decrease in absorbance of 1.0 × 10−4 at OD290 nm·min−1 was considered 1 U of APX activity. The results are expressed on a FW (Fresh Weight) basis as U·g−1.
Malondialdehyde content
-
As mentioned previously[36], 0.5 g of plant sample was homogenized, incubated, and then centrifuged to collect the supernatant. The absorbance was measured at 450, 532 and 600 nm.
H2O2 content
-
A 0.5 g sample of plant material was homogenized and centrifuged to collect the precipitate. Then, the precipitate was added to 1.5 mL of 2 M H2SO4. The absorbance of the mixture was measured at 412 nm, and the content of H2O2 was calculated[37, 38].
The production rate of O2.−
-
The reaction buffer was composed of 50 mM phosphate buffer (pH 7.8) containing 17 mM sulfanilic acid, 1 mM hydroxylamine hydrochloride, 7 mM 1-naphthylamine, and 50 μL sample solution. The absorbance of the mixture was measured at 530 nm, and the production rate of O2.− was calculated using previously described formulas[39].
Principal Components Analysis
-
Principal Components Analysis (PCA) of SOD, POD, APX, CAT enzyme activities, H2O2 and MDA contents and O2.− production rate was processed using the OmicShare website (
https://www.omicshare.com ).Statistical analysis
-
Data were based on three replicates in each experiment, and the experiments were repeated independently three times. Statistical significance was assayed using a one-way analysis of variance with IBM SPSS Statistics (SPSS version 20.0; Armonk, NY, USA), and the results are expressed as the means ± SDs. Significant differences were calculated by a t test (p < 0.01 or p < 0.05).
-
To study the potential role of H2S in response to B. cinerea infection, we measured endogenous H2S production in infected wild-type leaves by lead acetate strips (Fig. 1a). As strips shown in Fig. 1b and gray intensity analysis in Fig. 1c, the leaves produced more H2S with D-Cys as the substrate when infected with B. cinerea for 1, 2 and 3 d compared with control leaves. Subsequently, we examined the transcript levels of DCD1/2 and LCD1/2 in the infected leaves and found that after infestation the transcript level of DCD1 significantly increased, especially at 3 DPI was three times that of 2 DPI, and the transcript level of LCD1 slightly increased, whereas the transcript levels of LCD2 and DCD2 did not change significantly (Fig. 1d). Therefore, we hypothesized that DCD1 might affect the resistance of tomato to B. cinerea. To investigate the phylogenetic relationships between DCD proteins in tomato and other plant species, the gene encoding AtDCD1 (AT1G48420) in Arabidopsis was searched in the NCBI database, and the homologous-proteins were searched in S. lycopersicum, A. thaliana, V. vinifera, P. patens, N. tabacum, O. sativa, T. aestivum, P. bretschneideri, Z. mays, C. annuum, and M. acuminata using the AtDCD1 protein sequence as the query. As shown in the phylogenetic tree in Supplemental Fig. S1, the identified DCDs could be classified to two groups. The I subfamily contained DCD1 in the above species, and the Ⅱ subfamily contained all DCD2 proteins formed a single branch. The results indicated that DCD1/2 in tomato showed higher homology with homologs in chili pepper, both belong to the Solanaceae family.
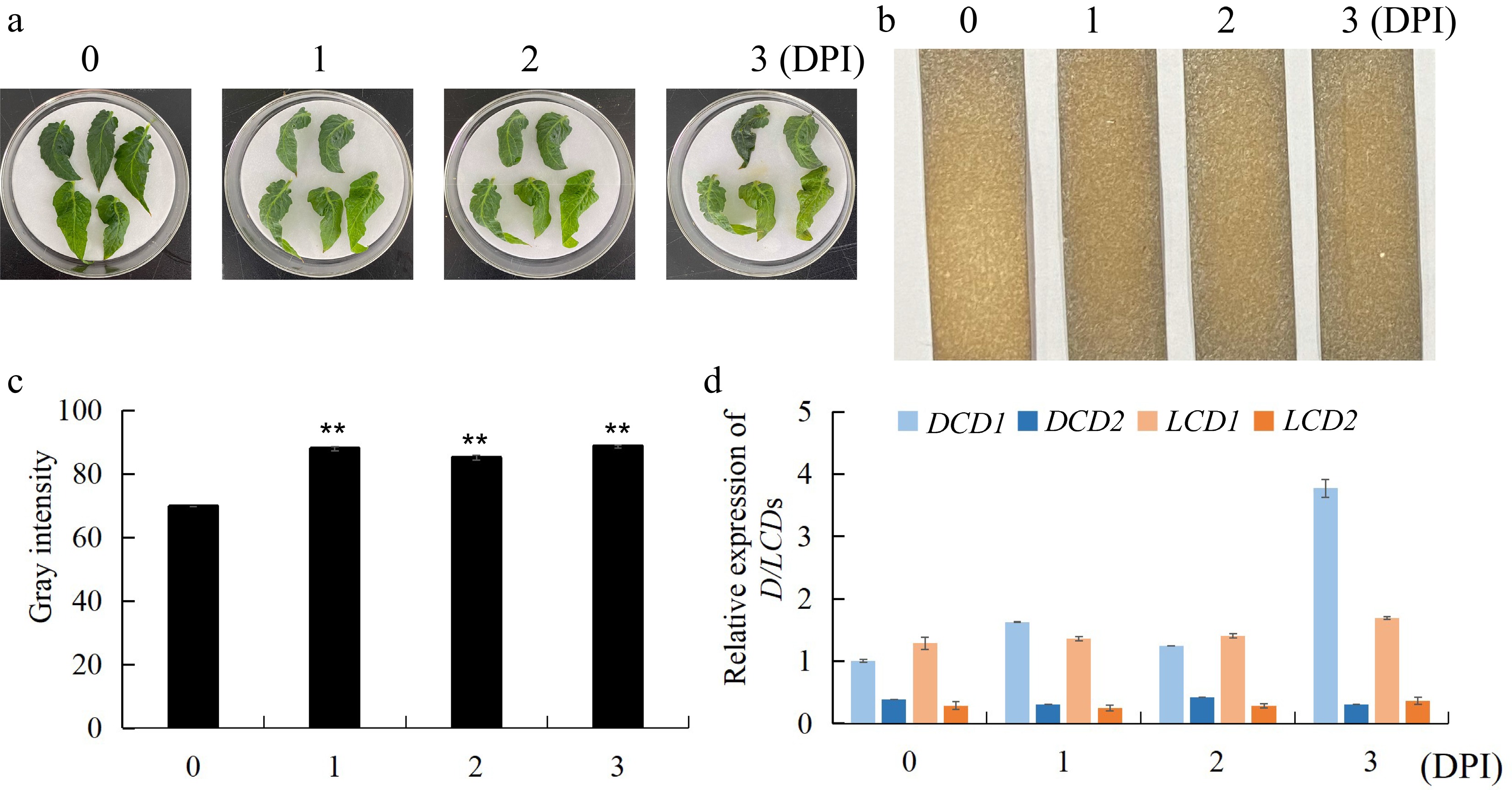
Figure 1.
Response of H2S and L/DCDs expression of tomato leaves to B. cinerea infection. (a) Phenotypes of WT leaves infected by B. cinerea for 0, 1, 2, 3 DPI. (b) The endogenous H2S production in infected WT leaves, was measured by lead acetate H2S detection strips. (c) The gray intensity analysis of strips in (b). (d) Gene expression of DCD1, DCD2, LCD1, LCD2 in WT leaves infected with B. cinerea. Data indicate mean ± SD (n = 3). The symbol ** stands for p < 0.01.
Construction of DCD1 gene-edited plant by CRISPR/Cas9
-
The sgRNA target of DCD1 was integrated into the CRISPR/Cas9 vector which was further transformed into tomato using Agrobacterium-mediated transformation. For the genotyping of positive T2 plants (Fig. 2a), the gene fragment flanking sgRNA target of DCD1 was amplified from genomic DNA of dcd1 mutant. Figure 2b indicated that 128 bp deletions in dcd1-1 near the PAM destroyed the transcription start site of DCD1, while there are 2 bp deletion in dcd1-2 which leaded to frame shift mutation, specifically, the translation stopped after the 70rd amino acid residue.
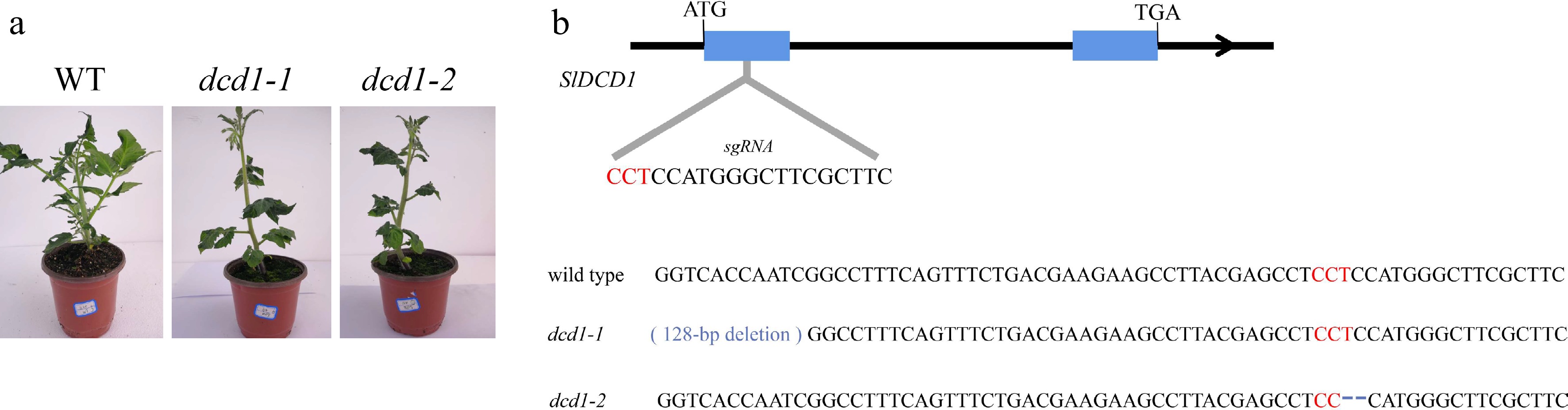
Figure 2.
The overall phenotype of the two mutant lines dcd1-1 and dcd1-2 at 45 d of growth. (a), (b) Generation of dcd1 tomato lines by CRISPR/Cas9. The protospacer-adjacent motif (PAM) is indicated in red and the dashes mean deletions of bases.
dcd1 mutant leaves showed increased susceptivity to B. cinerea infection
-
To investigate the effect of dcd1 mutation on resistance against B. cinerea, WT and dcd1 tomato leaves were inoculated with the fungal pathogen (Fig. 3a & b). Firstly, the relative expression of actin gene in B. cinerea, an index of pathogen growth, were determined at 3 and 5 DPI. Fig.3c showed that the expression actin gene increased significantly in dcd1 mutant leaves compared with WT leaves, suggesting that B. cinerea propagated more in dcd1 mutant. Moreover, trypan blue staining and DAB staining were used to observe dead cells distribution and H2O2 distribution in leaves, respectively. As shown in Fig. 3a & b, it was observed that more death cells were accumulated in dcd1 mutant leaves than WT leaves based on the trypan blue staining, and higher levels of H2O2 was found in mutant leaves according to DAB staining.
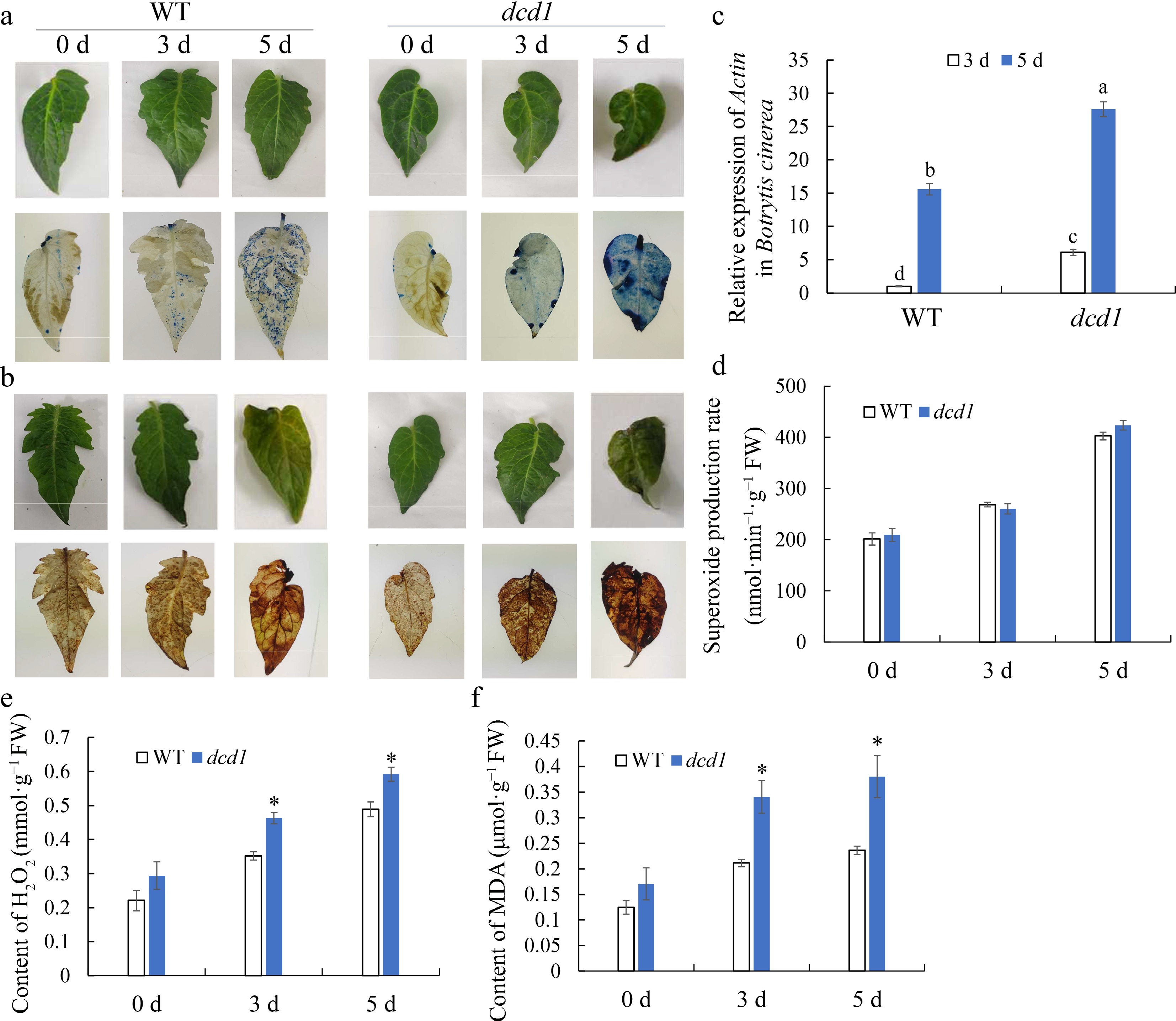
Figure 3.
Effect of dcd1 mutation on the ROS metabolism in tomato leaves infected with B. cinerea. Visualization of dead cells stained by (a) trypan blue and (b) H2O2 accumulation by DAB in tomato leaves of wild-type and dcd1 mutation. The expression of (c) B. cinerea actin gene, (d) O2.− generation rate, (e) H2O2 content and (f) MDA content in wild-type and dcd1 mutant leaves after infection with B. cinerea for 0, 3 and 5 d. Trypan blue staining and DAB staining of leaves infected with a conidial suspension were performed at different time points post inoculation (0, 3, and 5 d). The results of (c) - (f) are expressed as the mean values ± SD, n = 3. The symbols * or letters above the bars stands for student's t-test at p < 0.05.
ROS are the key feature of plant defense against invading pathogens[20]. As shown in Fig. 3d−f, O2.− generation rate, H2O2 content and MDA content displayed an increasing trend for 5 d of infection in both WT and dcd1 mutant leaves. The production of O2.− was not significantly different between WT and dcd1 (Fig. 3d), while H2O2 and MDA level in dcd1 leaves were significantly higher than that of WT leaves at 3 DPI and 5 DPI (Fig. 3e & f). These results imply that the tomato leaves defense to B. cinerea was largely suppressed in dcd1 leaves, and the dcd1 mutation caused excessive accumulation of H2O2 and MDA, suggesting that the lower level of H2S in dcd1 may lead to an imbalance in ROS metabolism and that excessive ROS may weaken the disease resistance of tomato leaves.
Effect of dcd1 mutation on antioxidant enzymes and transcription of defense-related genes in tomato leaves infected by B. cinerea
-
Antioxidant enzymes protect plants from oxidative stress and maintain redox homeostasis through scavenging of ROS produced during pathogen attack[40]. Then, antioxidative enzymatic activities, including SOD, CAT, APX and POD, in the leaves of WT and dcd1 were determined to assess the dynamics of the antioxidant system following challenge with B. cinerea. Figure 4b & c show that CAT and APX activity in dcd1 were always lower than that in WT infected by B. cinerea, and CAT activity decreased obviously in dcd1 leaves at 3 DPI and was just one-forth of that in WT leaves. At 5 DPI, CAT activity in WT was 1.5 times that in dcd1. Compared to WT leaves, at 3 DPI and 5 DPI, the SOD (Fig. 4a) and POD (Fig. 4d) enzyme activities in dcd1 leaves were also lower than that of WT. Overall, DCD1 mutation led to decrease in antioxidant enzyme activities, suggesting that a lower level of H2S in dcd1 may lead to excessive ROS accumulation which weaken the disease resistance of tomato leaves.
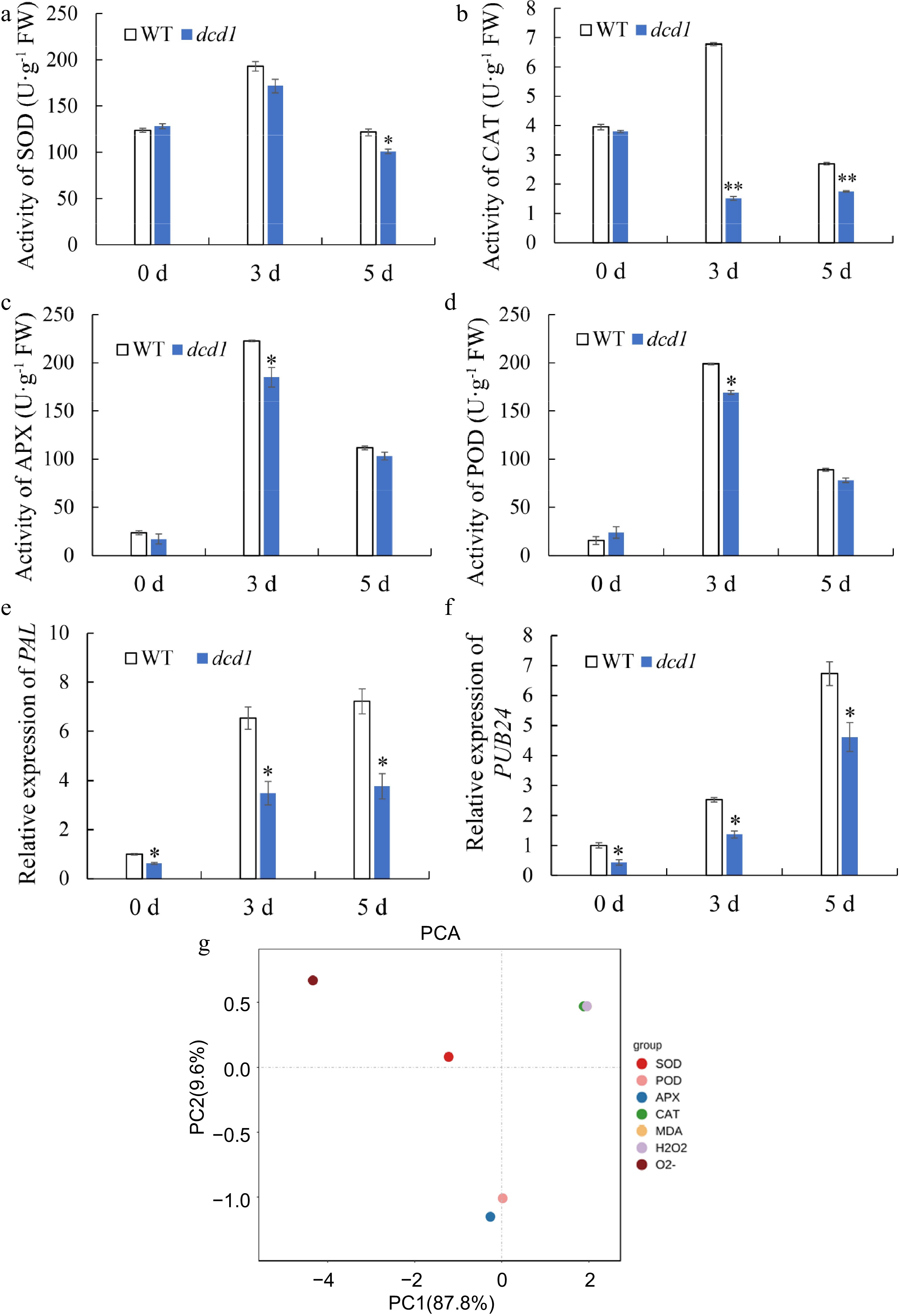
Figure 4.
Changes of antioxidant enzyme (a) CAT, (b) SOD, (c) APX, (d) POD activities and PCA analysis of the parameters of antioxidant enzyme activities and MDA content, content of H2O2, production rate of O2.− in tomato leaves after inoculating with B. cinerea for 0, 3 and 5 d. (g). The expression levels of pathogenesis-related genes (e) PAL and (f) PUB24 by RT-qPCR in tomato leaves after inoculating with B. cinerea for 0, 3 and 5 d. Data indicate mean ± SD (n=3). The symbols ** and * stand for p < 0.01 and p < 0.05, respectively.
In order to explore the clustering among different parameters mentioned above in leaves and determine their effect on plant disease resistance, we conducted PCA (Fig. 4g). The PCA score plot showed the total variance (97.4%) of the two main principal components, of which 87.8% accounts for principal component one (PC1) and 9.6% is responsible for principal component two (PC2). According to the scoring plot, the CAT activity, H2O2 content and MDA content on PC1 are the key factors affecting ROS metabolism in leaves.
Then, defense-related genes, including PAL (Fig. 4e) and PUB24 (Fig. 4f) were selected to investigate the transcript responses to B. cinerea in WT and dcd1 leaves. Phenylalanine ammonia-lyase (PAL) is a rate-limiting enzyme for the metabolism of phenylpropane substances in plants, and the infection of pathogens could induce enhanced activity of PAL, and the enhanced enzyme activity is positively correlated with disease resistance[41]. PUBs belong to U-box type E3 ligases functioned in plant defense responses[42]. As shown in Fig. 4e & f, the expression of defense-related genes PAL and PUB24 were remarkably induced by B. cinerea, while B. cinerea-triggered transcript induction of these genes were significantly depressed in dcd1 leaves. The relative expression of PAL in dcd1 leaves remained low at 3 DPI and 5 DPI compared to wild-type tomato leaves. At 0 DPI, the transcript level of PUB24 in dcd1 leaves was slightly lower than WT leaves, but the difference between the two widened significantly at 3 DPI. At 5 DPI, the transcriptional level of PUB24 in WT leaves was about 1.3 times that in dcd1 leaves. Overall, DCD1 mutation caused decreased expression of defense-related genes and excessive ROS accumulation.
Effect of dcd1 mutation on defense to B. cinerea in tomato fruit at different ripening stages
-
B. cinerea is a major threat to the production and storage life of tomato fruit around the world[43]. To investigate whether the DCD1 gene affect fruit defense against the fungal pathogen, WT and dcd1 tomato fruits were inoculated with B. cinerea, and the relative expression of actin gene of B. cinerea were determined at 3 and 5 DPI, and meanwhile the growth diameters of B. cinerea are recorded. As shown in Fig. 5a & b, at 3 and 5 DPI, the infection process of WT and dcd1 mutant fruit at mature green stage were not obvious. When the fruit were infected for 3 or 5 d, obvious infection lesions appeared on the surface of dcd1 breaker fruit, while WT breaker fruit did not display obvious lesions. As for the red fruit of WT and dcd1 mutant, both fruit developed obvious lesions at 3 DPI, but the diameter of the lesions in dcd1 mutant was significantly larger than that of WT, as was the case on the 5 DPI. Moreover, there was little difference in B. cinerea actin transcript levels at the sites of WT and dcd1 mature green fruit lesions, while the transcription level of B. cinerea actin on the dcd1 breaker fruit surface was about 2-fold that of WT, whether at 3 DPI or 5 DPI (Fig. 5c). The above results showed that the mutation of DCD1 reduced the resistance of breaker and red tomato fruit to B. cinerea largely. Besides, fruit at more mature stages were more susceptible to fungal pathogen infection.
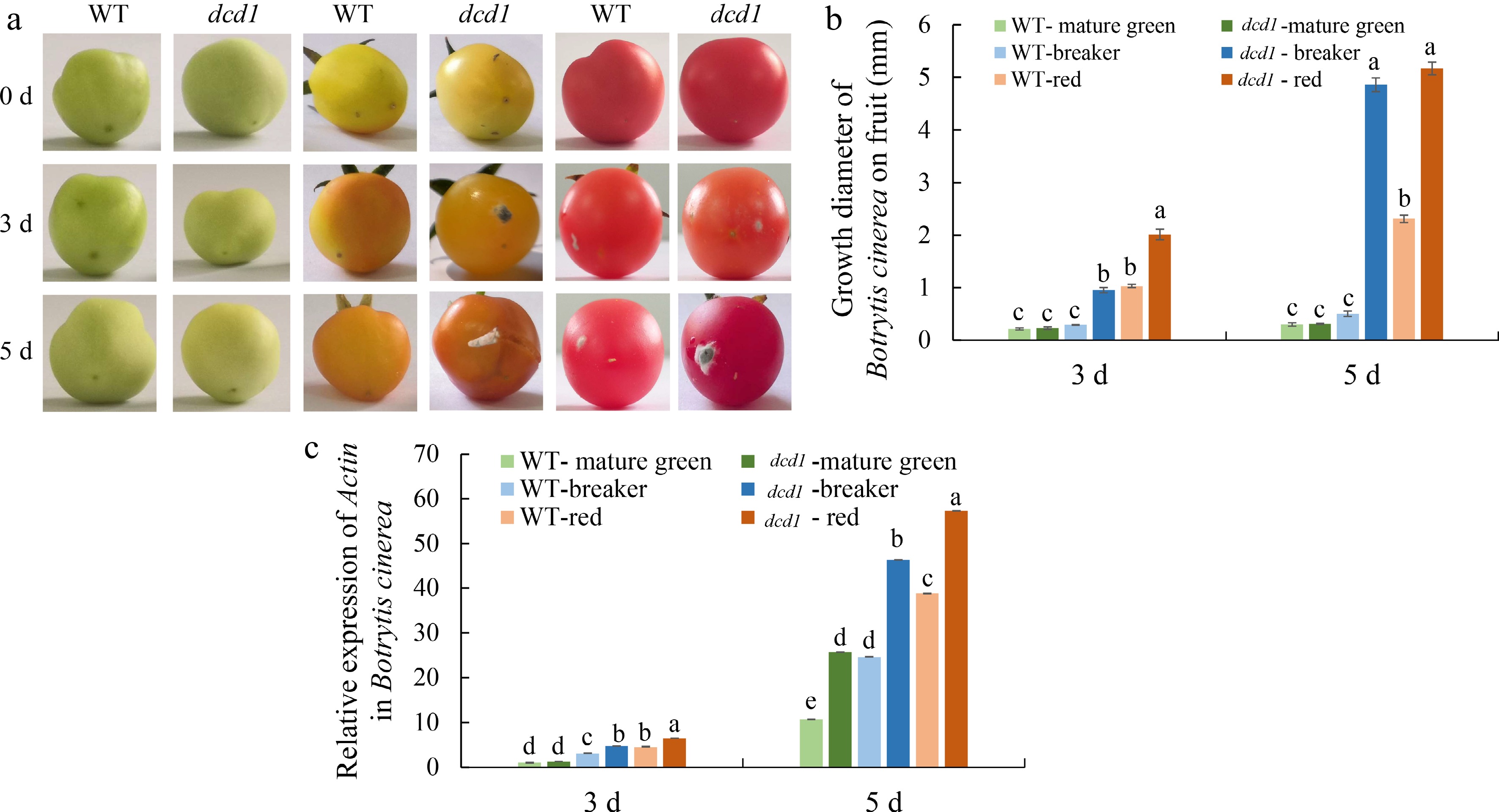
Figure 5.
Effect of WT and dcd1 mutation on defense to B. cinerea in tomato fruit at different ripening stages. (a) Images of WT and dcd1 fruits at mature green, breaker or red stages inoculated with B. cinerea for 0, 3 and 5 d. (b) Growth diameter of B. cinerea growing on the surface of WT and dcd1 fruits and (c) B. cinerea actin gene expression were detected after inoculating with B. cinerea. Values are the means ± SDs of three replicates. The letters above the bar indicate statistical significance determined by a student's t-test at the p <0.05 level.
Effect of dcd1 mutation on the accumulation of ROS in tomato fruit at different ripening stages infected by B. cinerea
-
The fruit tissues near the lesions were sampled for ROS determination. As shown in Fig. 6, O2.− production rate, H2O2 content and MDA content generally showed an increasing trend during B. cinerea infection for 0, 3 and 5 d in both WT and dcd1 mutant fruit at mature green, breaker or red stages. The production of O2.− was not significantly different between WT and dcd1 fruit, but fruit at red stage produced more O2.− than mature green and breaker fruit (Fig. 6a). At 3 DPI, the content of H2O2 in the mature green, breaker and red fruit of dcd1 were not significantly different from that of WT, however, H2O2 content in dcd1 mature green, breaker and red fruit was 1.4, 1.2, 1.3 times that in the counterpart WT fruit at day 0, respectively, suggesting that DCD1 mutation caused excessive H2O2 accumulation compared with WT (Fig. 6b). After infection for 5 d, dcd1 mutant fruit at all ripening stages showed significantly higher levels of H2O2 in comparison to WT. MDA content in dcd1 breaker fruit was significantly higher than that in WT fruit at 5 DPI, while the difference was not obvious between WT and dcd1 fruit at red stage at 5 DPI. MDA content in dcd1 red fruit was 1.4 times that in the counterpart WT fruit at 3 DPI (Fig. 6c). The results indicated that the deletion of DCD1 accelerated the accumulation of ROS in tomato fruit and fruit at red stage accumulated more ROS compared with un-ripened fruit.
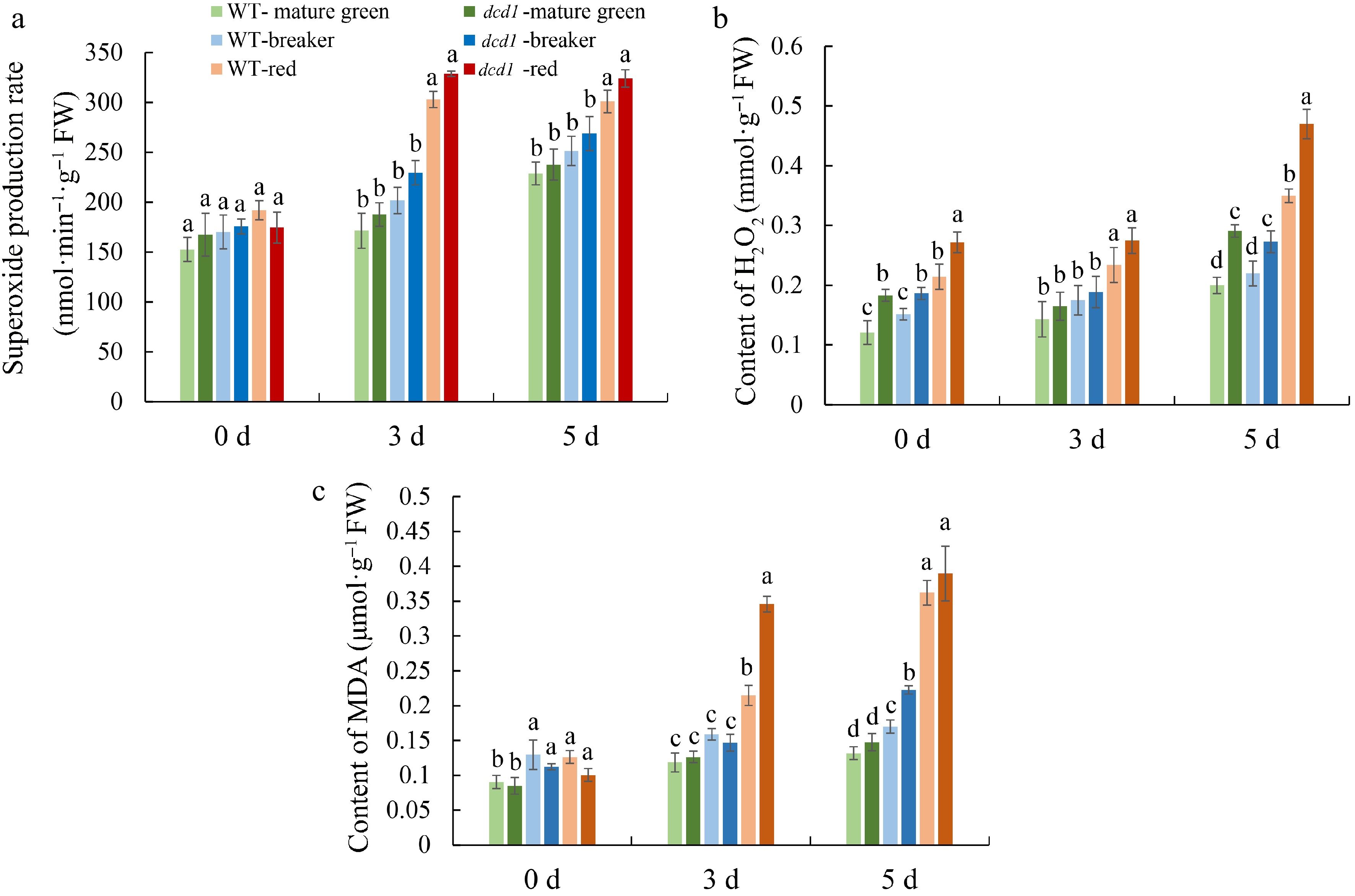
Figure 6.
Changes of (a) MDA content, (b) content of H2O2, (c) production rate of O2•− in tomato fruit of WT and dcd1 mutant at mature green, breaker and red stages after inoculating with B. cinerea at 0, 3 and 5 d. Data indicate mean ± SD (n = 3). Letters indicate statistical significance determined by a student's t-test at the p < 0.05 level.
Deletion of DCD1 affects antioxidant enzyme activities and expression of defense-related genes in tomato fruit at different ripening stages
-
To further investigate ROS metabolism in dcd1 mutant fruit, the enzyme activities of SOD, CAT, APX and POD were determined. There was minor difference in the enzyme activity of SOD in WT and dcd1 fruit, and at 5 d after infection, the activity in dcd1 mutant was lower than that of WT at different ripening stages (Fig. 7a). In both WT and dcd1 at breaker as well as red stages, CAT activity first rose at 3 DPI and then decreased at 5 DPI, and the activity in dcd1 was significantly lower than that in WT (Fig. 7b). At 5 DPI, CAT activity in dcd1 mutant was 92.1%, 80.3%, 41.7% of that in WT at mature green, breaker and red stages, respectively, suggesting that dcd1 mutant decreased CAT activity in fruit. and dcd1 mature green fruit, gradually increased, the activity in dcd1 was significantly lower than that in WT (Fig. 7b). Similarly, dcd1 mutant also caused decreased APX and POD activity in fruit at 3 and 5 DPI compared with the counterparts of WT (Fig. 7c, d). Overall, DCD1 mutation caused excessive accumulation of H2O2 and MDA, also led to decrease in antioxidant enzyme activities, suggesting that a lower level of H2S in dcd1 may lead to an imbalance in ROS metabolism and that excessive ROS may weaken the disease resistance of tomato fruit especially red and breaker fruit. As shown in Fig. 7g, the PCA score plot showed the total variance (96.4%) of the two main principal components, of which 90.0% accounts for principal component one (PC1) and 6.4% is responsible for principal component two (PC2). According to the scoring plot, the CAT activity, H2O2 content and MDA content on PC1 contributed more for ROS metabolism in fruit.
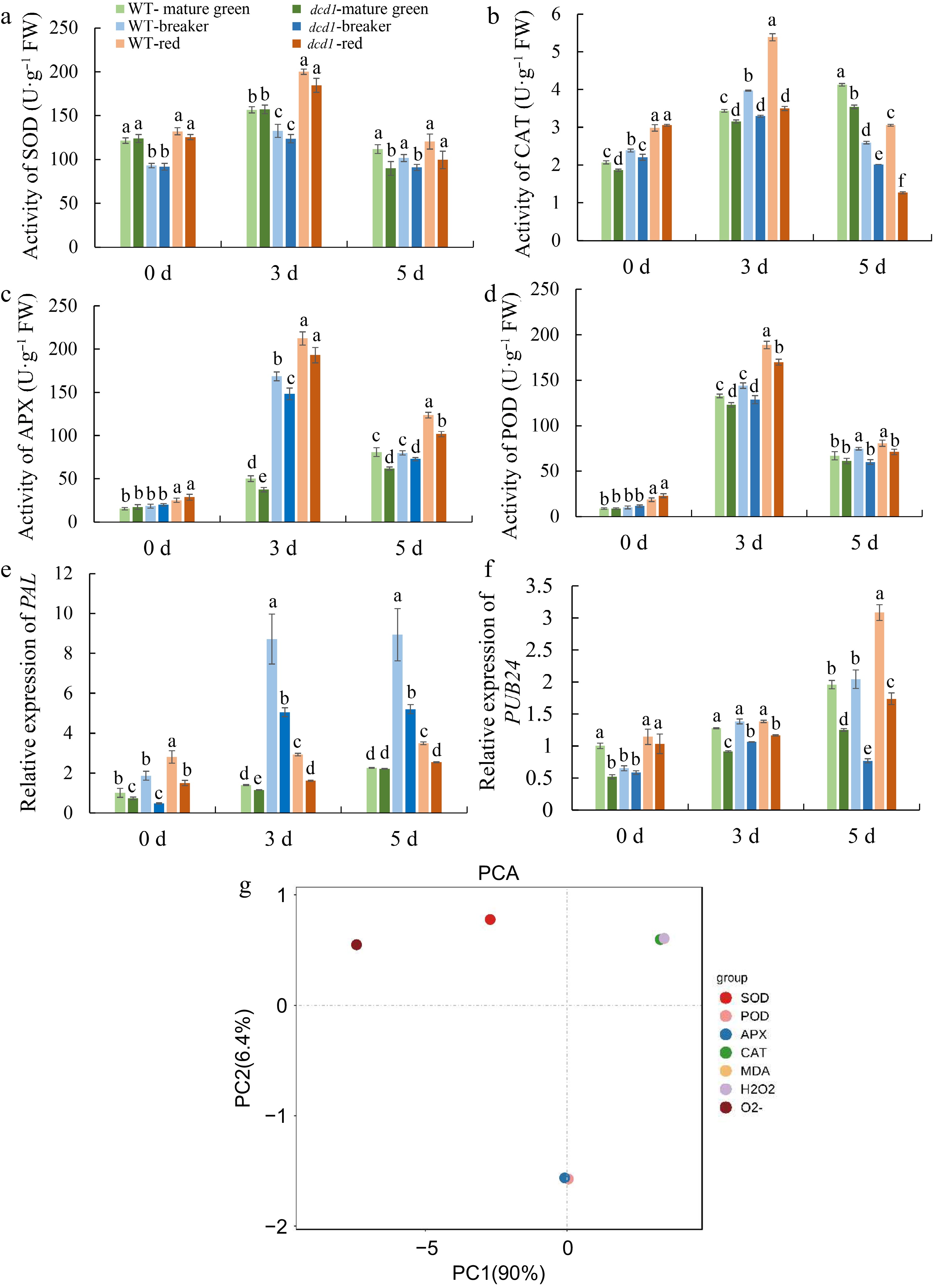
Figure 7.
Changes of antioxidant enzyme (a) CAT, (b) SOD, (c) APX, (d) POD activities and PCA analysis of enzyme activities and MDA content, content of H2O2, production rate of O2.− in tomato fruit of WT and dcd1 mutant at mature green, breaker and red stages after inoculating with B. cinerea at 0, 3 and 5 d. (g) The expression levels of pathogenesis-related genes (e) PAL and (f) PUB24 determined by RT-qPCR in WT and dcd1 tomato fruits after inoculating with B. cinerea. Data indicate mean ± SD (n = 3). Letters indicate statistical significance determined by a student's t test at the p < 0.01 level.
Then the expression of the marker genes for evaluating plant resistance to pathogen infection were determined. In WT fruit infected with B. cinerea, the expression of PAL was always higher than that in dcd1 (Fig. 7e). As shown in the Fig. 7f, the expression of PUB24 in WT fruit at all ripening stages were generally higher than that in dcd1 fruit. At 5 DPI, the expression of PUB24 in WT breaker fruit was approximately 1.8 times that in infected dcd1 breaker fruit, and that in WT red fruit was nearly 1.5 times that in dcd1 red fruit. Generally, dcd1 mutation caused attenuated expression of defense-related genes and this was consistent to the higher sensitivity of dcd1 fruit to B. cinerea infections compared with WT.
-
The role of H2S in plant disease resistance has gradually been confirmed. Exogenous application of H2S helped protect pear fruit from the invasion of the fungal pathogens Aspergillus niger, Penicillium expansum by inhibiting the growth of pathogens[6]. Besides, the transcript level of DES1 was elevated after pathogen infection, and DES1 overexpressing plants showed fewer of Magnaporthe oryzae in infected tissues compared to wild-type plants, whereas DES1 mutant plants showed increased bacterial growth[44]. D-cysteine desulfhydrase (EC 4.4.1.15), which catalyzes the conversion of D-cysteine to H2S, represent a completely different enzyme both in protein structure and biochemical properties[45]. In the present work, we found that B. cinerea infection of tomato leaves resulted in a significant increase in the release of H2S from the leaves with D-cysteine as the substrate and an increase in the expression of DCD1 was observed, suggesting the potential role of DCD1 in plant response to fungal pathogen infections. To further explore the function of DCD1 in tomato resistance to B. cinerea, we constructed T2 generation of dcd1 mutant tomato plant. The results showed that dcd1 mutant increased the susceptibility of leaves to B. cinerea and more B. cinerea reproduced evidenced by the higher actin expression in dcd1 mutant leaves. It was observed that more dead cells were accumulated in dcd1 mutant leaves than WT leaves, and higher levels of H2O2 in dcd1 mutant leaves. Besides, the resistance of tomato fruit was studied at mature green, breaker and red stages. At 3 and 5 DPI, the infection of B. cinerea on WT and dcd1 mutant fruit at mature green stage are not obvious. However, dcd1 mutant at breaker and red stages showed strong B. cinerea infection and the growth diameter B. cinerea was increased when infected for 3 or 5 d. There was little difference in B. cinerea actin transcript levels at the sites of WT and dcd1 mature green fruit lesions, while the transcription level of B. cinerea actin on the dcd1 breaker fruit surface was about 2-fold that of WT, whether at 3 DPI or 5 DPI. The above results showed that the mutation of DCD1 largely reduced the resistance of tomato leaves and breaker and red fruits to B. cinerea. Moreover, the infection data indicated that fruit at ripened stage is more susceptible to fungal infections compared with green fruit, suggesting that senescence of plant tissues is more favorable to fungal infection. Consistently, senescent tobacco leaves were more sensitive to necrotrophic pathogens including B. cinerea and Alternaria alternata[46].
Excessive accumulation of ROS has toxic effects on plants, leading to cell death and making plants more susceptible to diseases[47, 48]. It has been shown that ROS not only have direct antimicrobial activity, but also can act as a signal for defense response, causing upregulation of resistance-related genes and participating in the plant disease resistance process[49]. In the present study, we showed that the content of H2O2 in the leaves and fruit of dcd1 mutant at different ripening stages was higher than that of WT under B. cinerea infection. Besides, increasing trend of H2O2 was observed during B. cinerea infection, suggesting H2O2 is the key type of ROS that plant responses to fungal pathogen infection. Previous studies showed that H2S treatment greatly reduced H2O2 and MDA contents, and enhancing antioxidant enzyme activities and relative expression levels of defense-related genes, which in turn alleviated Fusarium head blight of wheat seedlings[50]. Previous reports suggested that H2S could delay postharvest senescence of multiple fruit and vegetables by maintaining balanced ROS homeostasis through activating antioxidant enzymes[51−53]. In the present study, significantly higher MDA and H2O2 levels were observed in dcd1 mutant fruits and leaves after B. cinerea infestation compared with WT. These data support that H2S generated by DCD1 appears to be an antioxidant signaling molecule involved in tomato resistance to B. cinerea.
To further investigate the role of DCD1 and H2S in mitigating ROS toxicity, we determined the activities of various antioxidant enzymes, including SOD, POD, CAT and APX. Figure 4b & c show that APX and CAT activities in dcd1 leaves were always lower than that in WT leaves infected by B. cinerea, and CAT activity decreased obviously in dcd1 leaves at 3 DPI and was just one-forth of that in WT leaves (Fig. 4b). Compared to WT leaves, at 3 DPI and 5 DPI, SOD, APX and POD showed lower activity in dcd1 mutant leaves compared with control. For tomato fruit, DCD1 mutation caused decreased SOD activity at 5 DPI, and decreased APX and POD at 3 and 5 DPI in different ripening stages of tomato fruit. In both WT and dcd1 of green, breaker as well as red fruit, CAT activity rose at 3 DPI and then decreased at 5 DPI, and the activity in dcd1 was significantly lower than that in WT. Previous studies have shown that the activities of POD, APX and CAT collectively regulated ROS homeostasis in slnpr1 mutants[40]. Among the antioxidative enzymes, SOD catalyzes the reaction of O2.− to H2O2 and O2[54], then CAT and APX are responsible for the decomposition of H2O2. In the present study, the decreased activity of antioxidative enzymes especially CAT, may contribute to excessive accumulation of H2O2 as observed in leaves and fruit infected with B. cinerea. Consistently, lower CAT activity in slnpr1 mutant leads to higher H2O2 levels compared to WT[40]. By PCA, we suggest that the CAT activity, H2O2 content and MDA content are the key factors affecting ROS metabolism in tomato leaves and fruit. Due to the attenuated antioxidative enzymes in dcd1 mutant, more ROS accumulated in dcd1 mutant leaves and fruits. For necrotrophic fungal pathogens such as B. cinerea, pathogen-induced formation of cell death and ROS accumulation normally promotes pathogen growth and lesion development[55]. Therefore, more dead cells and excessive ROS observed in dcd1 mutant may facilitate the infections by B. cinerea.
PAL is a key enzyme of phenylpropanoid metabolism and overexpressing PAL in tobacco decreased the susceptibility to fungal pathogen[56]. In our study, the relative expression of PAL in dcd1 leaves and fruit remained lower at 3 DPI and 5 DPI compared to WT leaves and in infected WT fruits, suggesting that mutation of DCD1 resulted in decreased PAL expression and diminished resistance in dcd1 mutant. PUBs belong to U-box type E3 ligases function in plant defense responses[42]. Therefore PUB24 was determined as the marker gene for disease response. In the present work, PUB24 increased significantly in leaves and fruits infected with B. cinerea, and the transcriptional level of PUB24 in dcd1 leaves or fruits was significantly lower than WT at 3 and 5 DPI. Thus we proposed that DCD1 and H2S are required for normal expression of PAL and PUB24 in response to fungal infections.
In conclusion, the present work indicated that DCD1 plays an essential role in tomato in response to B. cinerea. The mutation of DCD1 largely reduced the resistance of tomato leaves and breaker and red fruits to B. cinerea accompanied with increased ROS accumulation. DCD1 mutation caused decreased activity of antioxidative enzymes especially CAT, which may contribute to excessive accumulation of H2O2 as observed in mutant leaves and fruits infected with B. cinerea. Moreover, DCD1 mutation caused decreased expression of defense-related genes PAL and PUB24. Thus the work emphasizes that DCD1 and H2S are required for the activation of antioxidant enzymes and for ROS homeostasis in plant response to necrotrophic fungal pathogens. In addition, the work first provides strong evidence that fruit at ripened stage is more susceptible to fungal infections compared with green fruit, suggesting that senescence of plant tissues is more favorable to fungal infection.
This research was supported by the National Natural Science Foundation of China (31970312, 31970200, 32170315, 31901993), the Fundamental Research Funds for the Central Universities (JZ2021HGPA0063) and the Natural Science Foundations of Anhui Province (1908085MC72).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Yuqi Zhao, Kangdi Hu
- Supplemental Fig. S1 DCDs from S. lycopersicum, A. thaliana, V. vinifera, P. patens, N. tabacum, O.sativa, T. aestivum, P. bretschneideri, Z. mays, C. annuum, and M. acuminata were analyzed in the phylogenetic tree. The phylogenetic tree was constructed, the Neighbor-Joining was adopted using MEGA 7.0 software.
- Supplemental Table S1 Primers used in the present study.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhao Y, Hu K, Yao G, Wang S, Peng X, et al. 2023. D-cysteine desulfhydrase DCD1 participates in tomato resistance against Botrytis cinerea by modulating ROS homeostasis. Vegetable Research 3:21 doi: 10.48130/VR-2023-0021 

D-cysteine desulfhydrase DCD1 participates in tomato resistance against Botrytis cinerea by modulating ROS homeostasis
- Received: 01 February 2023
- Accepted: 16 May 2023
- Published online: 03 July 2023
Abstract: Tomato is one of the most popular horticultural crops, and many commercial tomato cultivars are particularly susceptible to Botrytis cinerea. Hydrogen sulfide (H2S) is an important gaseous molecule in various plant stress responses. In this study, it was found that endogenous H2S increases in tomato leaves in response to B. cinerea infection, along with a 3.8-fold increase in gene expression of DCD1 which encodes a H2S-generating enzyme D-cysteine desulfhydrase 1 in tomato at 3 DPI. Then we investigated the role of DCD1 in resistance of tomato leaves and fruits to B. cinerea. The mutation of DCD1 by CRIPSR/Cas9 greatly reduced the resistance of tomato leaves and breaker and red fruits to B. cinerea accompanied with increased reactive oxygen species (ROS) especially hydrogen peroxide (H2O2) and malondialdehyde (MDA) content increased by 1.2 and 1.4 times respectively at 5 DPI of leaves. Further investigation showed that DCD1 mutation caused decreased activity of antioxidative enzymes superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT) in both leaves and fruits, in particular, CAT activity in dcd1 mutant was 25.0 % and 41.7 % of that in WT at leaves and red fruits at 5 DPI. DCD1 mutation also caused decreased expression of defense-related genes PAL (encoding phenylalanine ammonia-lyase) and PUB24, and their expression in the dcd1 red fruit is approximately 1.3 and 1.8 times higher than in wild-type red fruit at 5 DPI, respectively. Thus, the work emphasizes the positive role of DCD1 and H2S in plant responses to necrotrophic fungal pathogens. In addition, the work provides strong evidence that fruit at ripened stage is more susceptible to B. cinerea infection compared with green fruit, suggesting that senescence of plant tissues is more favorable to B. cinerea infection.
-
Key words:
- H2S /
- Tomato (Solanum lycopersicum) /
- ROS /
- D‐cysteine desulfhydrase /
- Botrytis cinerea