









-
Strawberry (Fragaria × ananassa Duch.) is planted worldwide due to its unique flavor, high nutritional and economic value. With the rapidly developing modern agricultural economy, the research on fruit ripening and quality has been paid increasing attention. The sugar content, sugar-acid ratio, anthocyanins, and aroma are important indicators of fruit ripening and quality[1, 2]. In general, with the ripening of strawberry fruits, the content of sugar, soluble solids, aromatic volatile components, and anthocyanin components are significantly increased, while the content of total acids, phenols, flavonoids, and antioxidant capacity are decreased[3−5].
Macromolecular phosphorus is an important component of nucleic acid, phospholipid, enzyme, and ATP in cells, and has an irreplaceable role in life activities. So, phosphorus is one of the key factors of fruit quality regulation[6−9]. When strawberry plants lack phosphorus, especially in the flowering and fruiting period, this will affect the fruit setting rate, and the fruit flavor is poor and prone to deformity[7]. Cao et al. reported that the content of soluble solids was positively correlated with the content of phosphorus in ripe strawberry fruits[8]. Vacuolar phosphate transporter VPT1, also known as PHT5; 1, belongs to the PHT5 family and is mainly responsible for phosphate transport from cytoplasm to vacuoles[10, 11]. In strawberry, FaVPT1 affects fruit phosphorus content and sugar accumulation promoting fruit ripening, which is related to its SPX and MFS domains directly transporting soluble sugar and phosphate to vacuoles[12, 13].
The ripening and the quality of strawberry fruits include a series of significant changes in color, texture, taste, and aroma, which is a highly complex developmental process regulated by plant hormones and many transcription factors (TFs). In the early development stage of strawberry fruit, auxin and gibberellin (GA) produced by achenes promote ABA breakdown and fruit growth. During late fruit development, auxin and GA content decreased, while abscisic acid (ABA) content increased rapidly, which acts as the key signal molecule that promotes strawberry ripening and the formation of fruit quality[14, 15]. NAC family TFs FaRIF and FcNAC1, TCP family TF FvTCP9, FaWRKY71, ABA-stress-ripening (ASR), and auxin response factors (ARFs) are all key regulators of fruit ripening[13, 16−22]. A series of TFs can regulate anthocyanin biosynthesis, including R2R3-MYB TFs FaMYB10 and FaMYB1, FaRAV1, FaBBX22, and FvbHLH9[23−29]. TFs such as FaEOBII, FaDOF2, FaMYB63, and FaERF9 promote the production of eugenol, furanone, and other aroma substances by regulating the expression of related genes[30−33]. Sugar accumulation in fruits is also regulated by TFs, FvbZIP11 induces sugar accumulation in tomato, on the contrary, FaMYB44.2 and FvERF2 negatively regulates sucrose accumulation in strawberry[20, 34, 35]. Sucrose functions as an important signaling molecule which regulates anthocyanin accumulation in strawberry fruit through FaSUT1-mediated ABA signaling[36].
The plant-specific GRAS superfamily TFs are named after the first three identified members of Arabidopsis: the GAI (gibberellic acid insensitive), the RGA (repressor of GAI), and the SCR (scarecrow). They were initially divided into eight subfamilies: DELLA, LISCL, PAT1, HAM, LS, SCR, SHR, and SCL3, and more recently, into 10−17 subfamilies[37]. LRI-VHIID-LRII motif in the GRAS domain constitutes the DNA binding domain or the protein interaction binding domain[37]. To date, the number of GRAS family members has been described in several plants, such as 54 in strawberry, 53 in tomato (Solanum lycopersicum L.), and 50 members in Citrus reticulata Blanco[37−40]. Many GRAS superfamily TFs are reported to be involved in plant development and response to plant hormone signaling, especially GA and ABA. GRAS family DELLA proteins (GAI and RGA) are negative regulators in GA signal transduction, while SCL3, also a GRAS protein, acts as a positive regulator by attenuating the DELLA repressors in the root endodermis[41, 42]. It suggested that the GRAS TFs of the DELLA and SCL proteins integrate GA responses into ABA-controlled abiotic stress tolerance[43]. Overexpression of GRAS TF SlFSR leads to plant dwarf and reduced fruit weight, while SlFSR RNAi inhibits cell wall degradation and extends fruit preservation time[44, 45]. SlGRAS4 promotes tomato ripening by controlling ethylene synthesis-related genes and SlMADS1[46]. The FveRGA1 RNAi promotes the growth of strawberry stolon[47, 48]. PAT1 subfamily FaSCL8 is increased by ABA and mainly expressed in ripe fruit[49, 50].
Previous studies showed that the expression of FaVPT1 was precisely regulated in fruit development. In this study, starting with the screening of upstream regulators of FaVPT1, combined with expression analysis of strawberry at all stages of development, GRAS TF FaSCL8 may possess a key function in the strawberry fruit ripening mediating the transcript expression of FaVPT1. Revealing the regulatory relationship between FaSCL8 and FaVPT1 will lay the foundation for molecular breeding research related to improving the efficiency of phosphorus absorption and utilization in strawberry and improving fruit quality.
-
In order to find the TFs related to the regulation of FaVPT1, we used FaVPT1 promoter as bait to conduct yeast one-hybrid assay to screen the cDNA library derived from strawberry fruits (Fragaria × ananassa Duch.). A total of 15 putative candidate genes were found, one of which was a TF of the GRAS superfamily. According to our transcriptome data, the GRAS superfamily TF was highly expressed in ripe flesh (Supplemental Fig. S1), which suggested a positive role in strawberry fruit ripening. The GRAS superfamily TF encoded the homologous protein of AtSCL8 (AT5G52510), therefore, it was named FaSCL8. The full-length cDNA sequence of FaSCL8 contained an open reading frame of 2028 bp (Supplemental Data 1), which encoded a polypeptide of 676 amino acid. FaSCL8 had a calculated molecular mass of 73.8 kDa and a conserved GRAS domain as predicted (Supplemental Fig. S2) (
https://www.ncbi.nlm.nih.gov/cdd ). We made a systematic evolutionary tree which passed the full-length protein sequence of FaSCL8 and its homologous proteins in Fragaria vesca, Arabidopsis thaliana, Rose (Rosa chinensis), jujube (Ziziphus jujuba), mulberries (Morus notabilis), grape (Vitis vinifera), castor (Ricinus communis), apricot (Prunus mume), crab apple (Malus baccata), and pear (Pyrus bretschneideri) (Supplemental Fig. S3). The results showed that SCL8 is highly conserved in plants (Supplemental Fig. S3). Sequence comparative analysis showed that FaSCL8 had high similarity with the GRAS TF AtSCL8 in Arabidopsis and FveGRAS54 in Fragaria vesca[38, 51].To determine the intracellular localization of FaSCL8, a construct of FaSCL8 fused with GFP or the empty vector as control were separately infiltrated into the leaves of N. benthamiana for imaging. The laser confocal microscopy results showed that FaSCL8-GFP signals specifically overlapped with the 4',6-diamidino-2-phenylindole dihydrochloride (DAPI) nucleus staining (Fig. 1a).
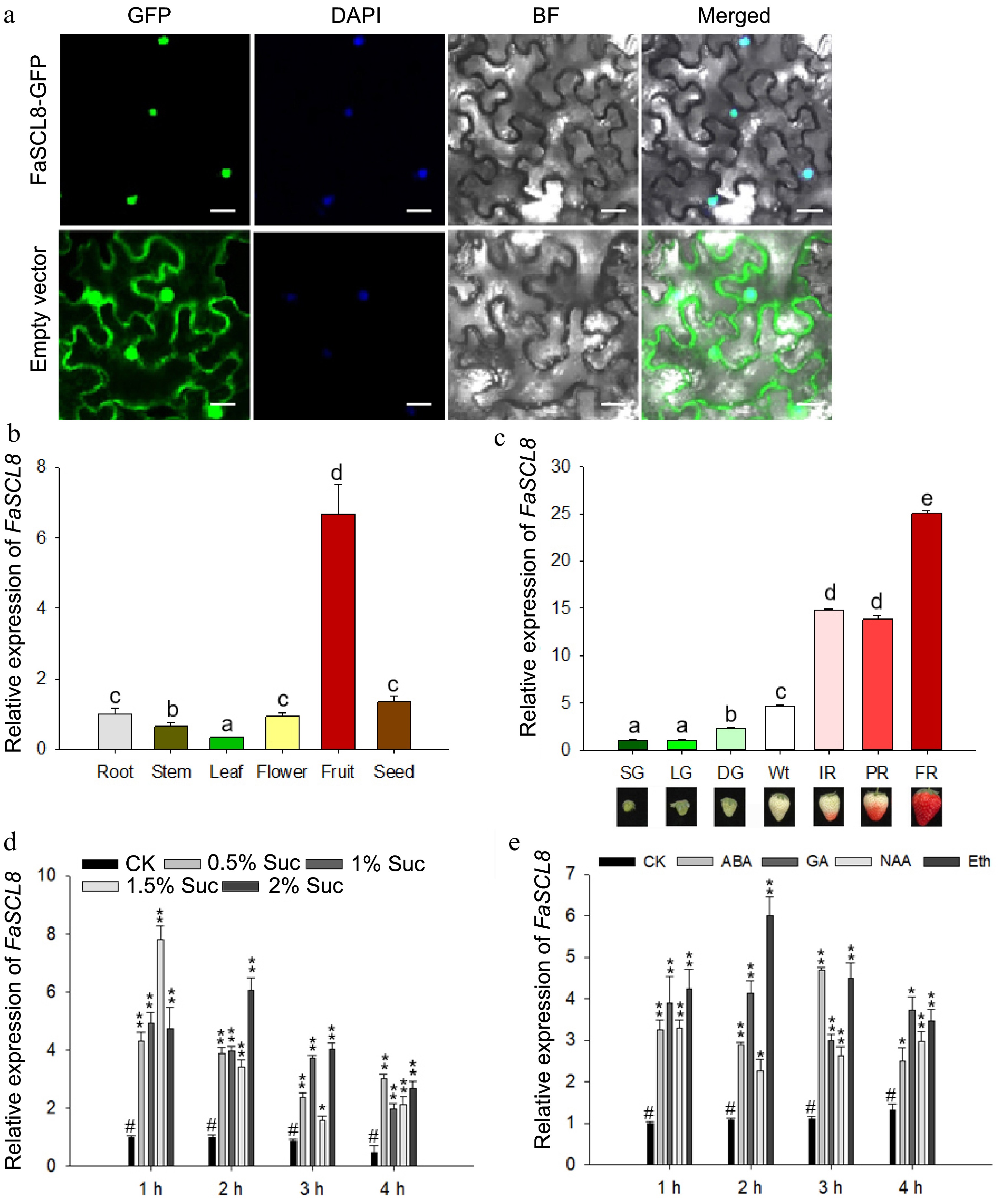
Figure 1.
FaCL8 localization and expression pattern at different stages and under different treatments. (a) Subcellular localization of FaSCL8-GFP fusions in transiently transformed N. benthamiana leaves. All experiments were assayed 48 h after infiltration. The same N. benthamiana leaves were stained with DAPI to show the locations of the nuclei. Bars = 20 um. (b) The transcript level of FaSCL8 in different tissues of strawberry. (c) FaSCL8 transcript levels in fruit receptacles at different developmental stages of Fragaria × ananassa cv. 'Benihoppe'. SG, small green fruit stage; LG, large green fruit stage; DG, green-white fruit stage; Wt, white fruit stage; IR, initial red fruit stage; PR, partial red fruit stage; FR, full ripening fruit stage. Relative expression values were relative to receptacles at the SG stage in all cases, which was assigned an arbitrary value equal to one. (d) The transcription level of FaSCL8 treated with different concentrations of sucrose. (e) The transcription level of FaSCL8 in fruit discs after exogenous plant hormones incubation. Bars are means SEs of three independent experiments (Student's t-test; * p < 0.05; ** p < 0.01).
The expression pattern of FaSCL8
-
Through RT-qPCR, the expression of FaSCL8 was tracked in different organs of strawberry and seven development stages of strawberry fruit. FaSCL8 transcripts level was higher in the ripe flesh than in the root or red seeds (Fig. 1b). When the strawberry fruit is mature, the expression of FaSCL8 increased rapidly and achieved the highest at the full red stage (Fig. 1c). Therefore, it is speculated that FaSCL8 may take effect in the process of fruit ripening.
Given that sucrose significantly promoted FaVPT1 expression, the effect of different concentrations of sucrose on the mRNA expression level of FaSCL8 were determined with fruit disc incubation in vitro. The results showed that compared with the control condition, sucrose significantly promoted FaSCL8 expression, especially 1.5% sucrose treated for 1 h (Fig. 1d), confirming a potential relationship of FaSCL8 with sucrose.
As a non-climacteric fruit, abscisic acid (ABA) and auxin are key regulatory components for strawberry development and signaling[52]. To evaluate whether FaSCL8 could regulate by ABA, gibberellin (GA), IAA, and ethylene. The fruit discs of Wt stage strawberry fruit were used for in vitro incubation with different plant hormones. These results showed that the transcript level of FaSCL8 was upregulated by phytohormones like ABA, GA, IAA, and ethylene (Fig. 1e).
FaSCL8 binds to and transactivates the FaVPT1 gene promoter
-
The 1,521 bp region upstream from the transcription start site (TSS) of FaVPT1 from 'Benihoppe' strawberry was amplified using specific primers (Supplemental Table S1), and it was divided into six fragments (P1−P6, Fig. 2a). The results of Y1H suggested that FaSCL8 could bind to the P6 fragment of the promoter (Fig. 2b). According to previous studies, GRAS protein can adjust downstream genes by identifying and binding to the GT cis-element (G(A/G)(A/T)AA(A/T))[37]. For this reason, we supposed that FaSCL8 functions through a similar mode of action. In order to analyze the direct binding of FaSCL8 to FaVPT1 promoter in vitro, an electrophoretic mobility shift assay (EMSA) was conducted using the purified recombinant protein FaSCL8-His.
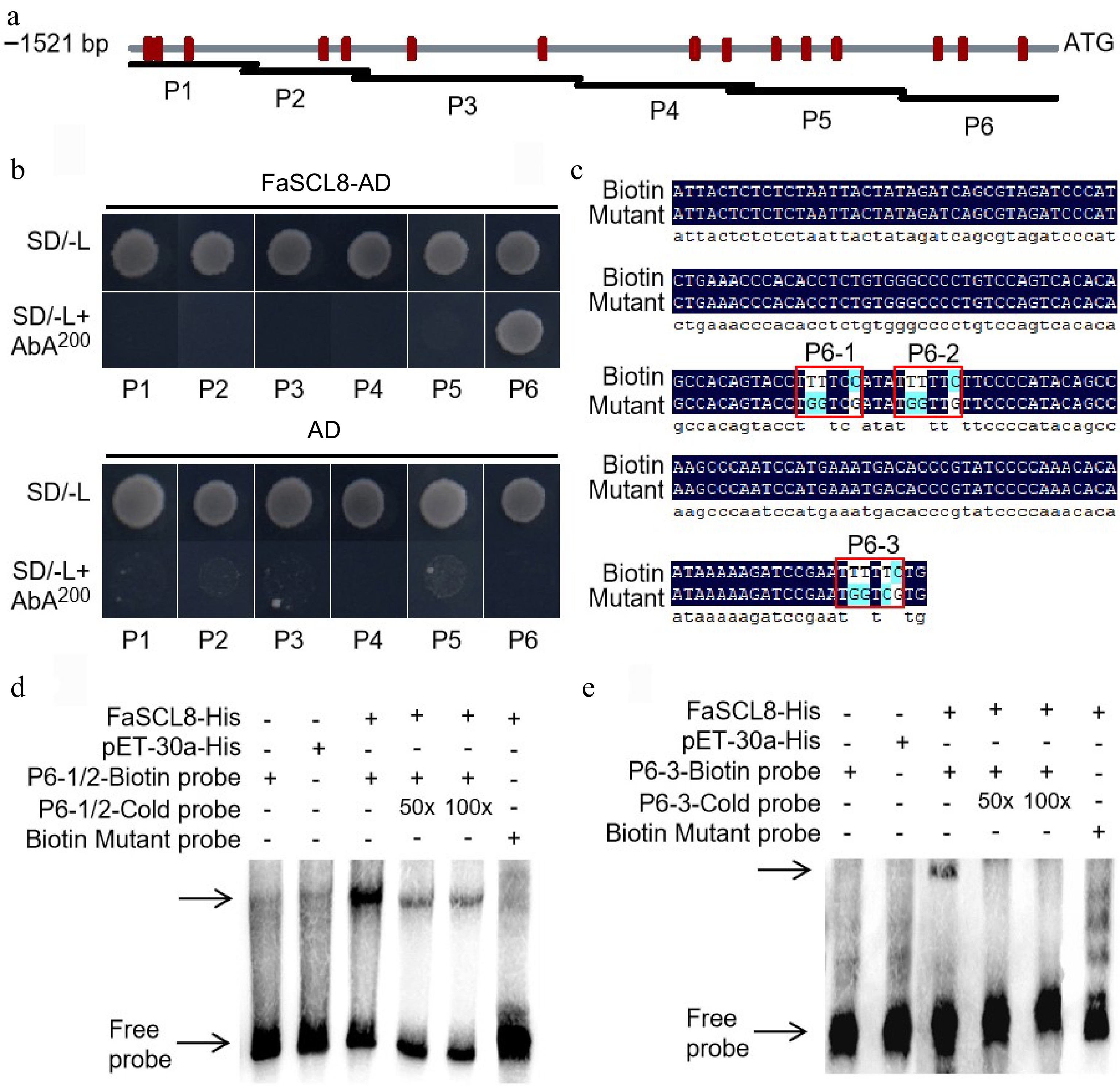
Figure 2.
FaSCL8 binds to FaVPT1 promoter. (a) Schematic diagram of FaVPT1 and its promoter, promoter area (gray line), GT element (red vertical line). (b) Yeast one-hybrid experiment verified that FaSCL8 combined with P6 promoter of FaVPT1 could grow on SD/-Leu medium with 200 ng/mL AbA. (c) P6-1/2 and P6-3 were shown in the figure, where P6-1/2 was −118 to −183 bp, and P6-3 was −36 to −105 bp, the binding of biotin labeled probe to its protein was a negative control, and the position of free probe band was shown by the arrow. EMSA between FaSCL8 and the promoters of (d) FaVPT1-P6-1/2 and (e) P6-3.
In order to determine whether FaSCL8 has transactivation activity on the FaVPT1 gene promoter in vivo, using dual-luciferase (LUC) assays, FaSCL8 was overexpressed in N. benthamiana leaves in the presence of FaVPT1 promoter (Fig. 3a). The coding region of FaSCL8 was driven by the CaMV 35S promoter, resulting in the effector plasmid 35S:FaSCL8. Infiltration of Agrobacterium tumefaciens harboring ProFaVPT1:LUC and empty vector into tobacco leaves produced only basal luciferase activity. When ProFaVPT1:LUC was co-transfected with 35S:FaSCL8, a significant increase in luciferase activity was detected (Fig. 3b), indicating the activation of FaVPT1 gene promoter by FaSCL8. When ProFaVPT1:LUC was co-expressed with 35S:FaSCL8, compared with the cells co-transfected with empty vector, the LUC/REN ratios were significantly increased (Fig. 3c). The above results demonstrated that FaSCL8 could bind to the promoter of FaVPT1 and activate the expression of the downstream protein.
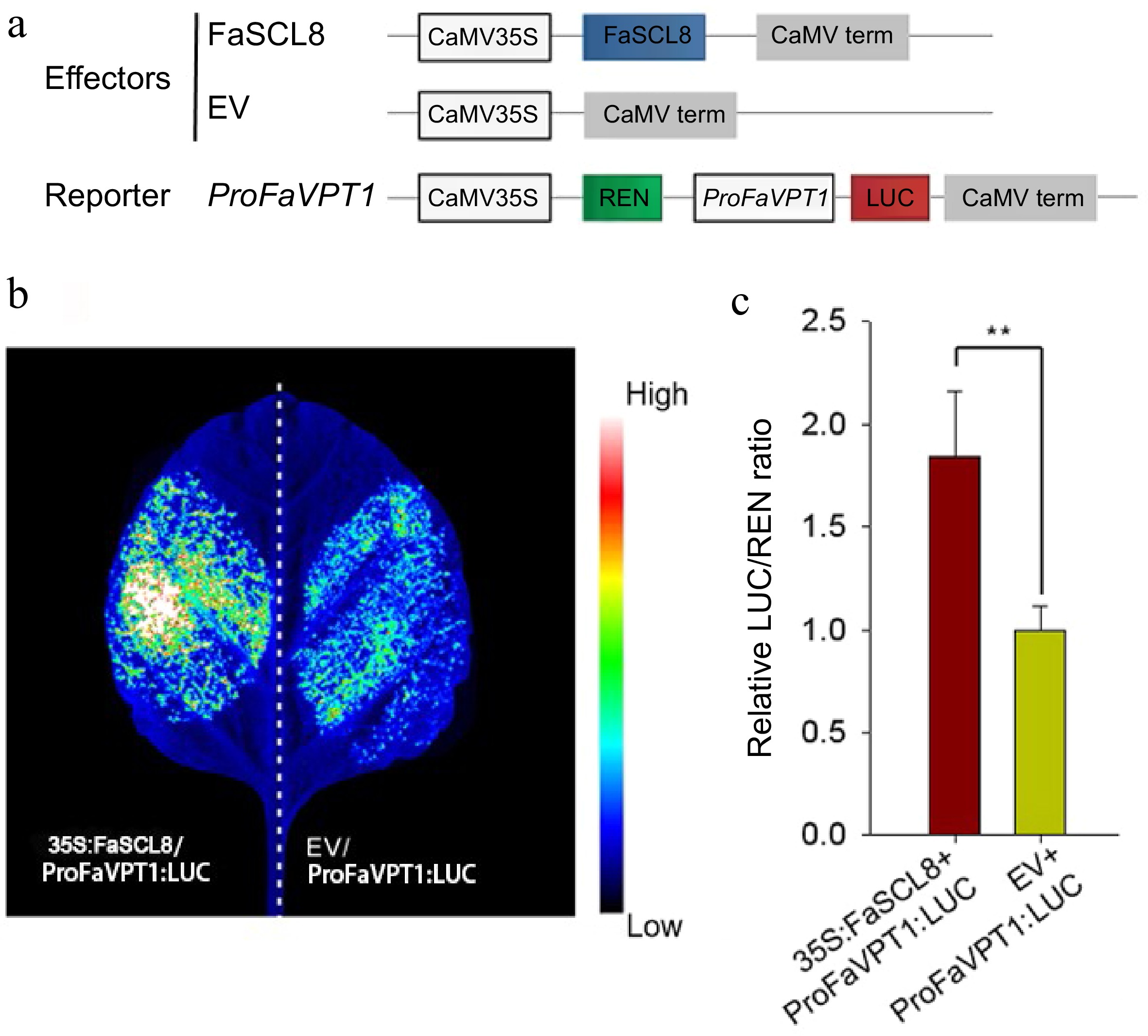
Figure 3.
FaSCL8 regulates the transcription of FaVPT1 in tobacco. (a) Schematic diagrams of the effector (35S:FaSCL8) and reporter vectors (ProFaVPT1:LUC) that were used in the dual-LUC assay. (b) Transactivation of the FaVPT1 promoter by FaSCL8. A representative image of an N. benthamiana leaf 48 h after infiltration is shown. Empty vectors: EV. (c) Promoter activities of FaVPT1 activated by FaSCL8 in dual-LUC assays were expressed as the ratio of LUC to REN in N. benthamiana leaves co-transformed with the effector and the reporter combinations. Data are means ± SE of five biological replicates. Statistical significance was determined using Student’s t test: ** p < 0.01.
FaSCL8 positively regulated FaVPT1 expression and fruit ripening of strawberry
-
In order to prove the role of FaSCL8 in strawberry, octoploid strawberry fruits were instantly transformed with an overexpression or RNA interference construction. Phenotypic observations showed that, overexpression of FaSCL8 could promote strawberry fruit ripening (Fig. 4a). RT-qPCR analysis showed that the FaSCL8 mRNA level in the fruits of the RNAi group with slower coloring was about 50% lower than that of the control fruits. While the mRNA level of FaSCL8 in the fruits of OE group, which showed the fastest coloring, was about 200% higher than that of the control (Fig. 4b).
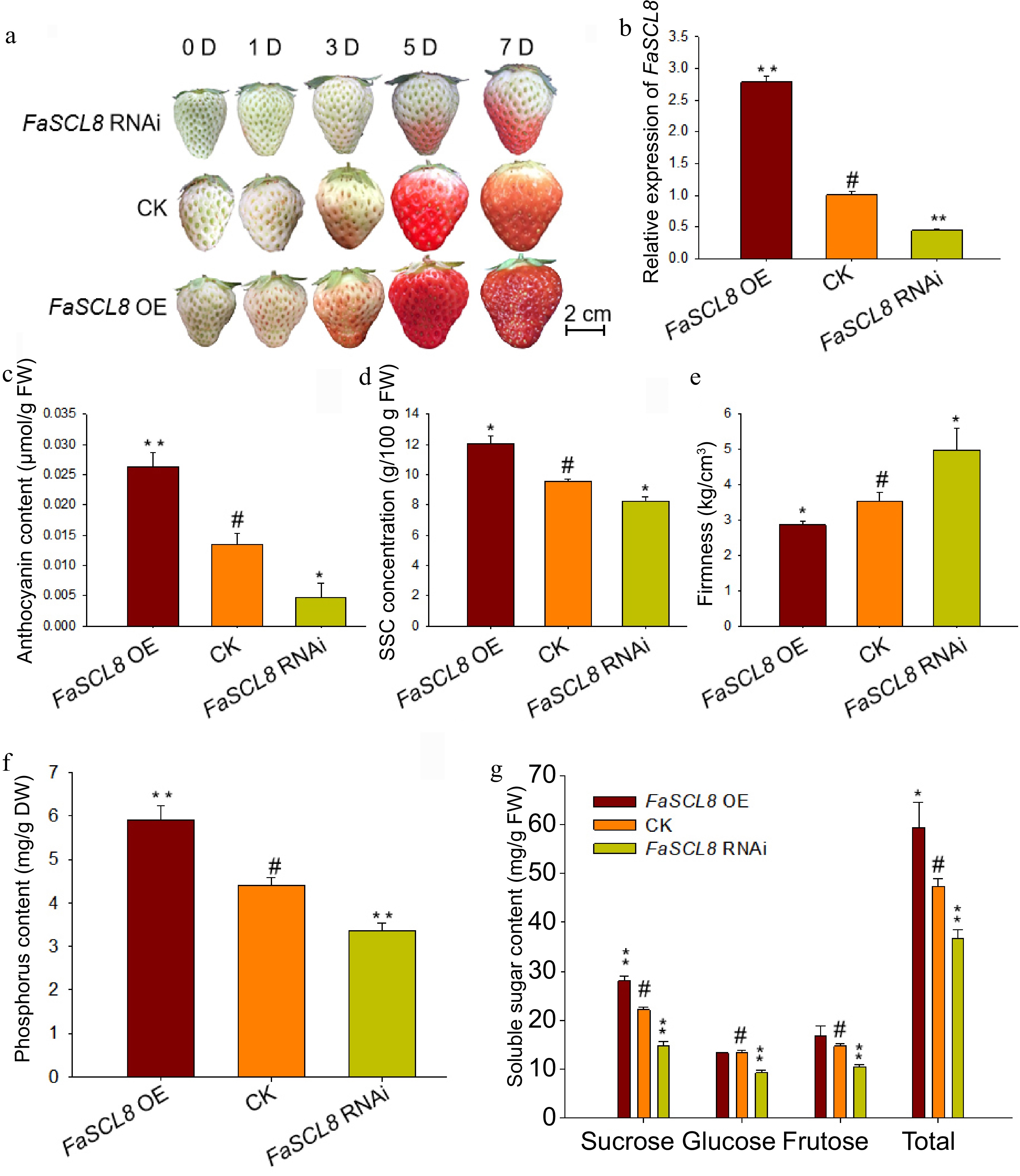
Figure 4.
Phenotype and physiological changes of FaSCL8 transgenic fruits. (a) Agrobacterium tumefaciens containing empty vectors, Super1300-FaSCL82028 or pk7GWIWG2(II)RR-FaSCL8226 constructs were injected into strawberry fruits at the de-green stage. After injection, photos were taken at 0, 1, 3, 5, and 7 d respectively to record the phenotype. (b) RT-qPCR was used to analyze the expression of FaSCL8 in OE, control and RNAi fruits, and three biological replicates were tested for all samples using the Actin gene from strawberry as an internal control. The relative expression levels were calculated using the 2−ΔΔCᴛ method. (c) Anthocyanin content, (d) SSC content, (e) firmness, (f) phosphorus, and (g) soluble sugar content of FaSCL8 transgenic and WT fruits. Bars are means SEs of three independent experiments (Student's t-test; * p < 0.05; ** p < 0.01).
The content of anthocyanin and Soluble Solids Content (SSC) were higher in FaSCL8 overexpression fruits than that of the control, while those in the RNAi fruits were lower (Fig. 4c & d). The firmness also showed that FaSCL8 promoted strawberry fruit ripening (Fig. 4e). It was reported that FaVPT1 positively regulates the phosphorus and sucrose accumulation[12]. Herein, the content of phosphorus and soluble sugar in the transgenic fruits were further determined. The phosphorus, sucrose, and total soluble sugar content were improved in FaSCL8 OE fruits, but receded in FaSCL8 RNAi fruits (Fig. 4f & g). The above results showed that FaSCL8 could positively regulate the accumulation of phosphorus, promoting strawberry fruit ripening and fruit quality.
In addition to the physiological indexes related to maturity detected above, we tested the relative expression of FaVPT1 in transiently infected fruits. The results suggested that the expression level of FaVPT1 was evidently up-regulated in FaSCL8 OE fruits and down-regulated in FaSCL8 RNAi fruits (Fig. 5a).
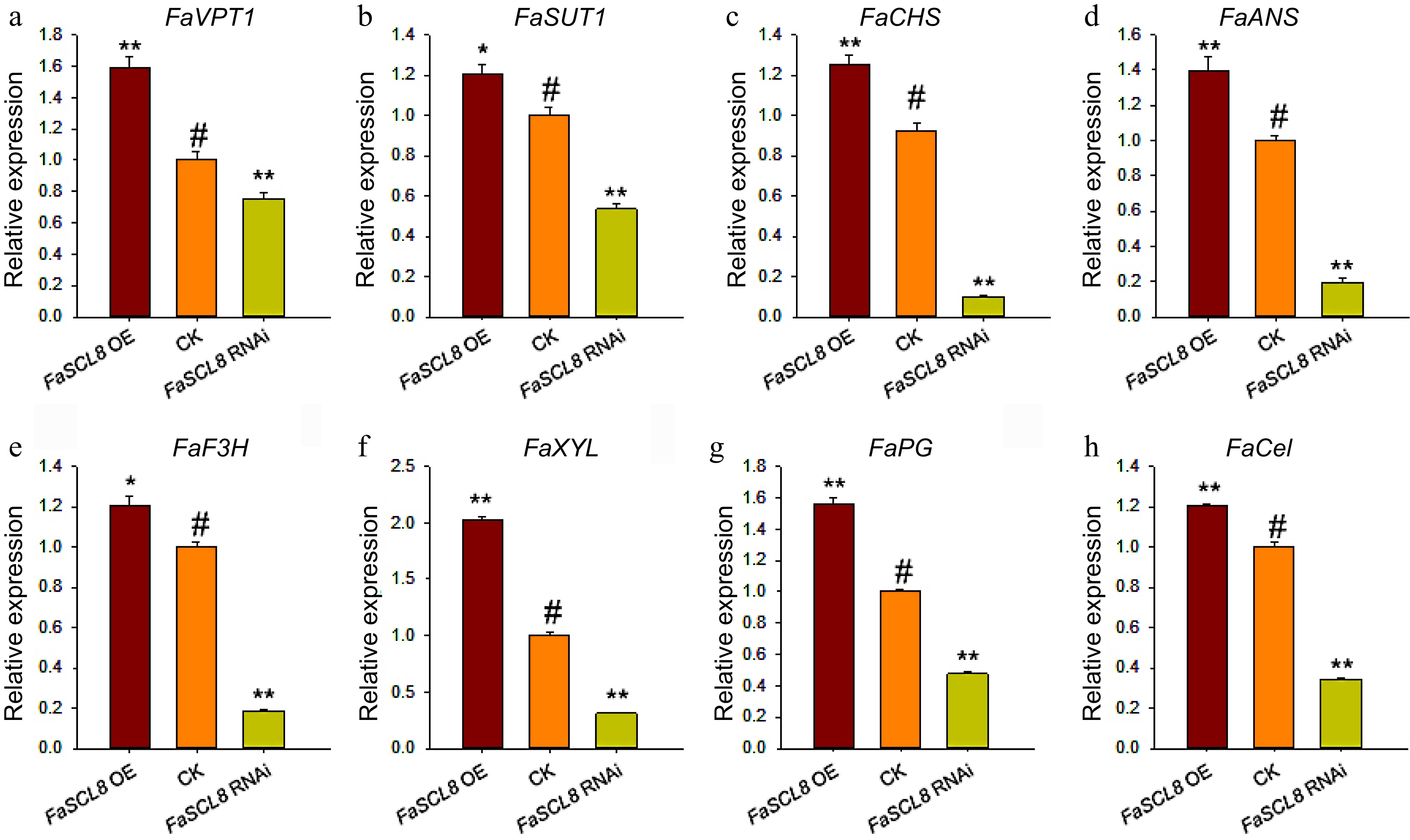
Figure 5.
Genes expression in FaSCL8 transgenic fruit. (a) Relative expression of FaVPT1 in the FaSCL8 transgenic and control lines. (b) Relative expression of FaSUT1 in the FaSCL8 transgenic and control lines. (c)−(e) Relative expression of anthocyanin biosynthesis genes in the FaSCL8 transgenic and control lines. (f)−(h) Relative expression of several genes associated with firmness in the FaSCL8 transgenic and control lines. All gene expression data were verified by RT-qPCR. The gene IDs of all verified genes are listed in Supplemental Table S1. Bars are means SEs of three independent experiments (Student's t-test; * p < 0.05; ** p < 0.01).
Based on RT-qPCR, the expression of marker genes related to fruit maturity and quality in transgenic strawberry fruits was detected to determine whether FaSCL8 is involved in the regulation of fruit ripening and quality at the molecular level. The result suggested that the expression of sugar-related marker gene FaSUT1, coloring-related marker genes FaCHS, FaF3H, and FaANS, firmness-related marker genes FaXYL, FaPG, and FaCel were significantly higher in the FaSCL8 OE fruits, but significantly lower in the RNAi fruits (Fig. 5). The expression of these marker genes was consistent with the physiological results and phenotypic observations in previous experiments, indicating that FaSCL8 promoted fruit ripening and fruit quality, partially through regulating the expression of FaVPT1.
-
Our previous reports revealed the role of FaVPT1 and its critical SPX and MFS domains in fruit ripening. FaVPT1 could transport soluble sugar and phosphate into vacuoles. Many TFs have been identified as important regulators that affect strawberry quality and fruit ripening[14−36], but few TFs, except FvPHR1, were reported to be involved in phosphate accumulation[9]. In this study, the GRAS TF FaSCL8 but not FvPHR1 was identified by screening with the promoter of FaVPT1. We found that FaSCL8 functions in positively regulating phosphate and sugar accumulation by directly activating the transcription of FaVPT1 during fruit development (Figs 2−5).
FaSCL8 was also shown to activate the transcription of anthocyanin biosynthesis related genes FaCHS and FaANS and anthocyanin content significantly increased and decreased in FaSCL8 OE and RNAi fruits, respectively (Figs 4 & 5). These results suggested FaSCL8 would activate the genes' transcription to regulate anthocyanin biosynthesis. It was reported that silencing FaSCL8 significantly inhibited the transcript accumulation of anthocyanin biosynthesis related genes PAL2, CHS1, CHS2, CHI, F3H, UFGT and FaMYB10, a TF that regulates the expression of most of the genes involved in anthocyanin production in ripened fruit receptacles[22, 50]. Whether FaSCL8 could control the transcription level of genes related to flavonoid/anthocyanin biosynthesis directly or by affecting the expression of FaMYB10 needs further study in the future.
FaSCL8 belongs to the Phytochrome A signal transduction 1 (PAT1) subfamily from GRAS superfamily[38], and potentially modulates flavonoid or anthocyanin biosynthesis[50]. A few genes from PAT1 subfamily have been reported to be involved in the regulation of fruit maturation, quality or plant stress response. Silencing SlGRAS2 reduces fruit weight in tomato[53]. VaPAT1 regulates jasmonic acid biosynthesis in grape cold stress response[54]. SCL21 is a member of the GIBBERELLIN-INSENSITIVE, REPRESSOR of ga1-3, which controls the periclinal division of the endodermis/cortex initial daughter cell[55]. The heterodimeric transcription factor complex ETHYLENE RESPONSE FACTOR115 (ERF115)–PAT1 maintains meristem function by promoting cell renewal after stem cell loss[56]. It is necessary to better understand the role of GRAS genes in fruit development and ripening regulation.
During fruit ripening, the expression of FaSCL8 is induced by hormone and sucrose
-
The expression of FaSCL8 was much higher than that in the other tissues in ripened strawberry fruit (Fig. 1b). FaSCL8 expression increased significantly during maturation and reached the highest in the full red period (Fig. 1c). Meanwhile, external application of ABA and sucrose could up-regulate the transcription level of FaSCL8 in strawberry fruit when Wt stage fruits were treated (Fig. 1d & e). Hormones have key effects on fruit ripening and quality control, especially, ABA is an important hormone involved in fruit ripening[14]. Sucrose is not only a major photosynthate but also serves as a signal of fruit ripening via ABA-dependent pathways, which appears to play a major role in the regulation of fruit ripening[36]. Therefore, FaSCL8 may be involved in the ABA and sucrose related fruit ripening regulation network. Similarly, the expression of VmSCL8, a homologous gene of FaSCL8 in bilberry (Vaccinium myrtillus L.) was significantly elevated in ripe fruit and up-regulated by ABA indicating a potential role in ABA-regulated fruit ripening processes in bilberry[57]. However, the upstream regulatory factors of FaSCL8 and the mechanisms of molecular interaction between FaSCL8 and ABA are still unclear.
It is interesting that FaSCL8 acted as a positive regulator of ripening, but was also induced by NAA, GA, not only induced by ABA. RGA, the GRAS family DELLA protein, was accumulated in root cell nuclei and disappeared from those nuclei within a few hours of GA treatment, which was a negative regulator in GA signal pathway[58]. Meanwhile, the GRAS TF SCL3 expression was reduced by exogenous bioactive GA (GA3), and by contrast, its expression was up-regulated by the GA biosynthesis inhibitor paclobutrazol (PAC), acting as a positive regulator of GA signal[42]. As a member of the GRAS family, FaSCL8 is considered to have general roles during development, particularly in response to GA through protein or transcription level regulation[51].
-
Previous studies showed that the expression of FaVPT1, which transports phosphate and sucrose to regulate strawberry fruit ripening, was precisely regulated during fruit development. In this work, we found that GRAS superfamily TF FaSCL8 can positively regulate the transcription of FaVPT1 by directly binding to its promoter. The transient transformation indicated that FaSCL8 could improve phosphorus and sugar accumulation in fruit and promote strawberry fruit ripening. This study reveals a new regulatory mechanism of strawberry fruit ripening and provides a theoretical basis for molecular breeding research.
-
Strawberries (octoploid cultivar, Fragaria × ananassa 'Benihoppe') were planted in greenhouses of Beijing University of Agricultural, with a circadian rhythm of 10 h darkness and 14 h light. The environment of these greenhouses is maintained at 20−30 °C, humidity 70%. Strawberry fruit was divided into seven developmental stages: Small green (SG, 7 d), large green (LG, 14 d), de-greening (DG, 17 d), white (Wt, 24 d), initial red (IR, 26 d), partial red (PR, 28 d), and full red (FR, 30 d). We used liquid nitrogen to quickly freeze all fruits and tissues at different stages and stored them in a refrigerator at −80 °C. Nicotiana benthamiana plants used for dual-LUC in this study were planted in a light culture room with a light/dark cycle of 16 h/8 h at 24 °C.
RNA extraction and RT-qPCR
-
RNA extraction was carried out as described by the plant RNA extraction kit of Beijing Biotechnology Co., Ltd. (Beijing, China) for detailed operation steps. Total RNA was separately isolated and reversed transcription refers to kit instructions. Quantitative real-time PCR (RT-qPCR) was performed in accordance with the manufacturer's protocols. Using the Actin gene in strawberry as the internal control, three biological repeated tests were carried out on all samples. The primers involved in the test are listed in Supplemental Table S1. The relative expression levels were calculated using the 2−ΔΔCᴛ method[59].
Incubation of fruit discs in vitro
-
The fruit disc tissues of strawberry fruit were treated with sucrose and hormones respectively in vitro and incubated as described by Hou et al.[60]. Strawberry fruits in the Wt stage were cut into 1 mm slices and immersed in different concentrations of sucrose or different kinds of hormones (100 μM ABA; 150 μM GA3; 500 μM NAA; 100 μM Ethephon; a treatment including six fruits per replication), and basic medium was used as the control. The fruit discs were placed in a 250 mL flask with basic medium and shaken at 25 °C, after washing with ddH2O, the tissues were frozen in liquid nitrogen and kept at −80°C. The expression level of FaSCL8 was measured by quantitative real-time PCR (RT-qPCR). The experiment was performed with three replications.
Full-length CDS cloning and sequence analysis of FaSCL8
-
A partial GRAS sequence was acquired in the octoploid strawberry cDNA library by yeast one hybrid. The sequence was used as a query in a BLAST search against National Center for Biotechnology Information (NCBI) databases. Candidate genes are the genes with the highest matching degree with the partial sequences, and primers (Supplemental Table S1) were designed to amplify the full-length CDS from 'Benihoppe' strawberry fruits cDNA. The conserved domains of FaSCL8 protein were analyzed by NCBI Conserved Domains Search (
www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ).The SCL8 homolog protein sequences queried from NCBI were used to construct the systemic development trees. Multiple sequence alignment was processed into a maximum likelihood systemic development tree using MEGA5.1.
FaSCL8 OE and FaSCL8 RNAi strawberry fruits were transiently transformed by Agroinfiltration
-
The full-length CDS of FaSCL8 was isolated by PCR from strawberry fruit cDNA using primers as described in Supplemental Table S1. To generate the FaSCL8 OE construct, the full-length CDS sequence of FaSCL8 was cloned into the pSuper1300-GFP vector. To generate the FaSCL8 RNAi construct, the fragment of FaSCL8 (137-362 bp) was ligated into pk7GWIWG2(II)RR vector. The Super1300-FaSCL82028, pk7GWIWG2(II)RR-FaSCL8226 or empty vectors were transformed into Agrobacterium strain GV3101.
Agrobacterium containing pSuper1300-FaSCL8 or pk7GWIWG2(II)RR-FaSCL8226 was injected into de-greening strawberry fruits. The injection site was cut 7 d after injection, then quickly frozen with liquid nitrogen, and stored at −80 °C for subsequent experiments.
Subcellular localization experiment
-
The full-length CDS of FaSCL8 was cloned into the overexpression vector pSuper1300-GFP. Agrobacterium infection experiments were carried out on N. benthamiana leaves. The Agrobacterium strain GV3101 (carrying P19) containing either pSuper1300-FaSCL8-GFP or pSuper1300-GFP was grown at 28 °C. Cells were collected and resuspended with infiltration buffer (10 mM MgCl2, 10 mM MES, and 0.1 mM acetosyringone). The OD600 of the bacterial suspension was adjusted to a final concentration of 0.6−0.8. The transformed cells were kept away from light for 2−3 h at room temperature. The suspensions were injected into the leaves from the abaxial leaf surface with a syringe, and material was collected after 3 d.
The leaf tissues of N. benthamiana were cultured with DAPI and observed by Leica confocal laser scanning fluorescence microscope with a 40× objective. When detecting GFP fluorescence, the excitation wavelength is 488 nm (10 % intensity) and the emission wavelength is 500−550 nm. DAPI signal was determined using an excitation wavelength is 405 nm and the emission wavelength range is 445−460 nm.
Y1H assay
-
The Y1H screening was conducted using the MatchmakerTM Gold Yeast One-Hybrid Library Screening System (Takara, Kyoto, Japan) and Yeastmaker Yeast Transformation System 2 (Takara, Kyoto, Japan). The screening was carried out according to the manufacturer's instructions. The 1,521 bp promoter of FaVPT1 was divided into six fragments and connected to pAbAi vector as the bait construct. In the end, positive clones were obtained by DNA sequencing.
The Y1H assay was also used to detect the ability of FaSCL8 to bind to FaVPT1 promoter. The six promoter fragments of FaVPT1 were amplified by PCR using specific primers (Supplemental Table S1) and ligated into the pAbAi vector to obtain bait vectors. The full-length CDS of FaSCL8 was fused in frame with the GAL4 activation domain (AD) in the pGADT7-AD vector to generate the prey vector (pGADT7-FaSCL8). The prey vector was transformed into the bait-reporter strain. Yeast cells co-transformed with the prey and bait vectors were plated on synthetic dropout SD/-Leu medium supplemented with or without Aureobasidin A (AbA) and incubated for 2−5 d at 30 °C.
EMSA
-
The full-length of FaSCL8 was inserted into the pET-30a vector to obtain the FaSCL8-His recombinant plasmid. The recombinant plasmid was introduced into BL21 (E. coli) cells, and FaSCL8- His fusion protein expression in E. coli cells was induced with 1 mM IPTG at 25 °C for 5 h. Generally, each treatment used approximately 200 ng purified recombinant Fa SCL8-His.
Conserved cis-element motifs of the 1,521 bp FaVPT1 promoter were predicted using PLACE (
www.dna.affrc.go.jp/PLACE/signalscan.html ) and PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ). Seventy bp oligonucleotide probes containing GRAS-binding sites were synthesized and labeled with biotin at the 5'-hydroxyl end of the sense strand according to the manufacturer's instructions. All primers involved in EMSA were shown in Supplemental Table S1. EMSA was conducted according to the reported method[60]. To confirm the specificity of the shifted band, 50-fold or 100-fold amount of unlabeled identical or mutated oligonucleotides were incubated with nuclear proteins 20 min before the addition of labeled oligonucleotides. The signal was detected using LightShiftTM Chemiluminescent EMSA Kit (Thermo Fisher, USA).Dual-LUC activity assay
-
The 1,521 bp FaVPT1 promoter was fused into pGreen000-LUC vector as reporter plasmids. In the same vector, Renilla LUC (REN) under the control of the empty 35S vector was used for ratiometric analysis. The plasmids including 35S:FaSCL8 effector, empty 35S vector, and ProFaVPT1:LUC were transfected into A. tumefaciens strain GV3101 separately. A. tumefaciens carried different constructs were mixed in a certain proportion. The mixture (1.2 mL) for detecting the regulatory effect of transcription factors on their downstream genes were 300 μL P19 + 300 μL ProFaVPT1:LUC + 600 μL 35S:FaSCL8. 300 μL P19 + 300 μL ProFaVPT1:LUC + 600 μL empty vector was used as the control group. After mixing, it was allowed to stand at 28 °C for 2 h; and coinfected into N. benthamiana leaves. Dual-Luciferase® Reporter Assay System (Promega, Madison, WI, USA) was used to detect the ratio of LUC to REN activity after 3 d of agrobacterium infection. Each combination includes at least six determinations.
Determination of total phosphorus content
-
To measure the Pi content, three transiently transformed strawberry fruits were used. After grinding in liquid nitrogen, 0.5−1 g powder each replicate. was used for the determination of Pi content following the reported method[61]. This experiment was repeated three times.
Determination of fruit firmness and soluble solids content
-
The firmness and soluble solid content of strawberry fruit were measured with three instantaneously transformed strawberry fruits (n = 3). Fruit firmness was measured using a fruit hardness tester (FHM-5, Takemura Electric Works Ltd, Tokyo, Japan). Sugar analysis instrument was used to determine the soluble solids content of receptacle (MASTER-100H, ATAGO, Tokyo, Japan), and the specific operation was referred to the method of Huang et al.[12]. The experiments were repeated three times.
Determination of anthocyanin content
-
Three transiently transformed strawberry fruits (n = 3) were used to determine anthocyanin content. Anthocyanin content was measured by reverse-phase HPLC using a ZORBAX Eclipse XDB-C18 column (4.6 × 150 mm, 5 µm; Agilent). Grinding 3 g fruit flesh sample into powder in liquid nitrogen, then adding 2.5 mL of methanol (1% HCl) to 0.5 g of powder. Specific operations of extraction, centrifugation, and content determination of anthocyanins were carried out according to the methods of Huang et al.[12]. Pelargonidin-3-O-glucoside was used as a standard. The entire process was repeated three times.
Determination of soluble sugar content
-
Transiently transformed strawberry fruits (n = 3) were used for the soluble sugar content determination. The soluble sugar content was determined using reverse-phase HPLC (1,200 Series, RID1 A detector; Agilent Technologies, Santa Clara, CA, USA). The supernatant was fractionated using a Sugar-Pak™ I column (6.5 × 300 mm; Waters, Milford, MA, USA) with 100% MilliQ water for 25 min at a flow rate of 0.4 mL·min−1. The column temperature was 80°C, and the injection volume was 20 µL. The standard samples used were D-(+)glucose, D-(−)fructose, and sucrose (Sigma-Aldrich)[12]. The entire process was repeated three times.
This work was supported by Beijing Natural Science Foundation (6222004), the National Natural Science Foundation of China (Project 32102362, 32030100).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Zhenzhen Zheng, Yanjun Zhang
- Supplemental Fig. S1 RPKM of FaSCL8 in transcriptome data.
- Supplemental Data 1
- Supplemental Fig. S2 Conserved domain analysis of FaSCL8.
- Supplemental Fig. S3 Phylogenetic analysis of FaSCL8 and its homologs in plants.
- Supplemental Table S1 Primer sequences used in the experiments.
- Supplemental Date 1 The full-length CDS sequence of FaSCL8.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng Z, Zhang Y, Gao Y, Shen Y, Huang Y. 2023. GRAS family transcription factor FaSCL8 regulates FaVPT1 expression mediating phosphate accumulation and strawberry fruit ripening. Fruit Research 3:15 doi: 10.48130/FruRes-2023-0015 

GRAS family transcription factor FaSCL8 regulates FaVPT1 expression mediating phosphate accumulation and strawberry fruit ripening
- Received: 02 February 2023
- Accepted: 29 May 2023
- Published online: 27 June 2023
Abstract: Strawberry is an extensively planted horticulture crop with multiple economic values. Vacuolar phosphate transporter (FaVPT1) can promote phosphate and sugar accumulation and improve strawberry fruit quality, however, its transcription regulatory mechanism remains unknown. Here, we report a GRAS superfamily transcription factor (TF) SCARECROW-LIKE8 (FaSCL8) which regulates FaVPT1 expression to control strawberry fruit ripening and quality. The promoter of FaVPT1 was used as a bait to screen the octoploid strawberry cDNA library by yeast one hybrid to obtain the candidate TFs including FaSCL8. FaSCL8 was located in the nucleus and was mainly expressed in fruit, its expression level was increased rapidly during ripening, and induced by exogenous phytohormones including ABA, GA, IAA, and ethylene, as well as sucrose. FaSCL8 can directly combine with the FaVPT1 promoter and regulate its expression in vitro and in vivo. The results of Agrobacterium-mediated transient infection of strawberry fruits showed that overexpression of FaSCL8 could increase soluble sugar, anthocyanin, and phosphorus content, promoting fruit ripening, while decreasing expression of FaSCL8 showed the opposite phenotype. The expression levels of fruit ripening and quality-related genes including FaVPT1, were improved in the over-expressed fruits of FaSCL8, while receded in the FaSCL8-silenced fruits. In conclusion, this study found that FaSCL8 can regulate FaVPT1 expression to improve phosphorus and sugar accumulation in strawberry fruits and promote fruit ripening, providing a new regulation mechanism of strawberry fruit ripening by phosphorus, a macroelement key to fruit quality improvement.
-
Key words:
- Strawberry /
- FaSCL8 /
- FaVPT1 /
- Phosphate accumulation /
- Fruit ripening