









-
Passion fruit (Passiflora edulis Sims.) originates from tropical America and is grown in many countries including China, Australia, New Zealand, Ecuador, Kenya, South Africa, and India[1]. It belongs to family Passifloraceae and has more than 500 species[2,3] of which about 50 are edible[4]. Presently, yellow (Passiflora edulis f. edulis) and purple (Passiflora edulis f. flavicarpa) are two main cultivars of passion fruit being grown worldwide and appreciated due to their unique flavor and taste[5]. In Chinese culture, it is also called 'fruit of 100 fragrances' because it contains the fruit aroma of more than 100 fruits including pineapple, banana, guava, strawberry, mango, etc.[2,6]. Passion fruit is also famous due to its great medicinal value[7], such as controlling blood sugar[8], anti-hypertension[9,10], anti-inflammation and reducing fat[11], protecting liver and kidney[12], and regulating cardiac autonomic nerve functions[13].
Anthocyanins, a class of flavonoids that serve as secondary antioxidants and have a variety of biological functions[14], such as growth prevention of tumors[15], anti-inflammation and oxidation[16]. Moreover, anthocyanins serve as plant coloring agents, are essential components of defense mechanisms, and shield plants from UV deterioration[16]. The primary variables that affect the ultimate anthocyanin concentration in ripe fruits are the rate of anthocyanin production in plants, membrane transit, and utilization or degradation[17−22]. There are several enzymes which play vital role in biogenesis of anthocyanins in fruits i.e., L-phenylalanine ammonia-lyase (PAL), cinnamate 4-hydrogenase (C4H), 4-coumarate: coenzyme A Ligase (4CL), chalcone synthase (CHS), UPD-3-O- glycosyltransferase (UFGT), and glutathione S-transferase (GST) (Fig. 1a)[3,23−25]. In the anthocyanin biosynthesis pathway, the specific role of PAL is to convert the phenylalanine into cinnamic acid[26,27]. Naringin chalcone is produced by the polyketone synthase (PKS) enzymes[28], which also produce the 4CL and CHS enzymes[29]. UFGT stabilizes anthocyanin production by joining the sugar component to anthocyanin glycogens[30]. Since the absence of these proteins may result in significant alterations in plant color, the GST enzyme is crucial for the transportation of anthocyanins in plants[31].

Figure 1.
(a) Anthocyanins' biosynthesis pathway in plants. PAL: L-phenylalanine ammonia-lyase; 4CL: 4-coumarate: coenzyme A Ligase; CHS: chalcone synthase; C4H: cinnamate 4-hydrogenase; GST: glutathione S-transferase; UFGT: UPD-3-O- glycosyltransferase; CHI: chalcone isomerase; F3H: flavanone-3-hydroxylase; FLS: flavonol synthase; DFR: dihydroflavonol 4-reductase; ANS: leucoanthocyanidin dioxygenase. (b) Cross-sectional images of fruits at various phases of development of both purple and yellow passion fruits.
The amount of phenolic compounds in plant tissues is mostly determined by the genetic makeup of the plant, although extrinsic stimuli may produce qualitative or quantitative alterations in the composition of these compounds[32,33]. Flavanols and proanthocyanins are the primary flavonoids in many fruits during the start of fruit growth, and the buildup of anthocyanin pigments is often a sign of ripening[34,35]. Being a climacteric fruit, passion fruit, ethylene's burst seems to have a regulatory function in both ripening and the start of anthocyanin production[36−38]. Many studies have shown that, in addition to hormonal control, environmental factors such as temperature, light levels, nutritional status, and biotic stressors have a substantial influence in the formation of anthocyanins in fruits[39,40].
There haven't been many thorough studies stating the regulation of anthocyanin production in fruits. By integrating molecular and biochemical data, one investigation focused on the impact of genetic, developmental, and environmental variables on the production of flavonoids in strawberry fruits[41]. The findings demonstrated a distinct developmental pattern in the fluctuation of flavonoid levels, associated gene expression, and enzyme activity. Anthocyanin and flavan-3-ol levels were significantly influenced by environmental factors, but flavanol levels and proanthocyanidin-related features were significantly influenced by developmental stage and genotype. Anthocyanin production in bilberries also seems to be strongly regulated genetically[42]. Passion fruit is the richest sources of anthocyanins, containing high quantities of anthocyanin mostly in the fruit peel[43]. Without intense direct sunlight, its biosynthesis mechanism can produce large amounts of anthocyanins[44].
Although several studies have been reported on anthocyanin and flavonoid metabolism of plants such as apples[45], grapes[46,47], and mulberry[48], the in-depth mechanism of their accumulation in passion fruits is yet to be studied. In current study, we investigated the changes in anthocyanins and their derivatives during passion fruit development. Furthermore, the activities of anthocyanin metabolism-related enzymes and RT-qPCR expressions of core genes responsible for the biosynthesis or degradation of those enzymes were studied. This study provided ample information regarding anthocyanin metabolism in passion fruit and laid the foundation for further functional analysis of studied enzymes and genes.
-
A passion fruit orchard in Shaowu county, Nanping city, Fujian province, China (27°22'51.9"N 117°32'18.4"E) was the source of the plant material for two passion fruit cultivars: yellow passion fruit (Golden) and purple passion fruit (Tainong No.1). The 15 passion fruits were tested from each cultivar while they were at each of the following developmental stages: fruitlet, green, veraison, maturity, and ripening (Fig. 1b). They were then transported to the laboratory, where the pulp (separated seeds) of five passion fruits were combined to create one biological replicate, with three biological replicates for each sample. All of the samples were quickly frozen in liquid nitrogen and were stored in an ultra-low temperature refrigerator at −80 °C for subsequent use.
Determination of total flavonoids, anthocyanins, and procyanidins
-
An earlier optimized technique was used to extract the total flavonoids from passion fruit pulp[49]. Eight mL of 60% ethanol were combined with precisely weighed 1 g of frozen fruit pulp (−80 °C). The solution underwent 40 min of ultrasonic extraction, followed by 20 min of cooling and 10 min of centrifugation (12,000 g, 20 °C). The final volume of 10 mL was then obtained by drawing 5 mL of the supernatant and diluting it with distilled water. In addition, a 2 mL aliquot was separated, followed by the addition of 3 mL of 60% ethanol and 0.3 mL of 5% NaNO2. The mixture was then violently shaken for a total of 6 min to ensure that all of the components were fully combined. After that, 0.3 mL of 10% Al(NO3)3 was added, and the mixture was shaken for a total of 6 min. Next, 4 mL of 4% NaOH was added, and the mixture was shaken for a total of 12 min. After that, the absorbance was determined by measuring it at 510 nm. In order to determine the total flavonoid content, a calibration curve of rutin standard (HPLC grade, more than 98 percent purity, Solarbio Life Sciences, Beijing, China) was used (Y = 10.859X−0.0617, R2 = 0.999).
Anthocyanins in their whole form were isolated by following the procedure outlined previously[50]. After adding a 10 mL solution of 1% hydrochloric acid: methanol that included 0.2 g of plant material, the mixture was left to stand for 5 h before being centrifuged for 20 min at a speed of 1000 revolutions per minute. Ten mL of the sample's supernatant was utilized in order to get an OD reading at 530 and 560 nm. The Eqn 1 was used to determine the total amount of anthocyanins in the sample.
$\begin{aligned}&\rm Total\; anthocyanins\; (mg\cdot g^{-1}) =\\&\rm \frac{(\mathrm{O}\mathrm{D}530-0.25\times \mathrm{O}\mathrm{D}650)\times \mathrm{ }\mathrm{v}\mathrm{o}\mathrm{l}\mathrm{u}\mathrm{m}\mathrm{e}\;\mathrm{o}\mathrm{f}\;\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{ }\mathrm{e}\mathrm{x}\mathrm{t}\mathrm{r}\mathrm{a}\mathrm{c}\mathrm{t}\mathrm{i}\mathrm{o}\mathrm{n}\;\mathrm{l}\mathrm{i}\mathrm{q}\mathrm{u}\mathrm{i}\mathrm{d}\;\left(\mathrm{m}\mathrm{L}\right)}{4.62\times 104\times \mathrm{ }\mathrm{f}\mathrm{r}\mathrm{e}\mathrm{s}\mathrm{h}\;\mathrm{w}\mathrm{e}\mathrm{i}\mathrm{g}\mathrm{h}\mathrm{t}\;\mathrm{o}\mathrm{f}\;\mathrm{p}\mathrm{a}\mathrm{s}\mathrm{s}\mathrm{i}\mathrm{o}\mathrm{n}\;\mathrm{f}\mathrm{r}\mathrm{u}\mathrm{i}\mathrm{t}\;\left(\mathrm{g}\right)} \end{aligned}$ (1) Procyanidin content was determined using a slightly modified method of Jaakola[34]. After carefully measuring out 0.5 g of the sample and adding 6 mL of methanol to a centrifuge tube of 10 mL capacity, the mixture was subjected to an ultrasonic treatment at a power of 250 and a rate of 50 kHz for a period of 20 min. The absorbance at 546 nm was determined by measuring the concentration of the supernatant that was left over after the solution had been centrifuged and allowed to reach room temperature. For the purpose of determining the amount of procyanidin present, a calibration curve of procyanidin standard (HPLC grade, ≥ 95% purity, Solarbio Life Sciences, Beijing, China) was used (Y = 0.0038X+0.0202, R2 = 0.999).
Determination of flavonoids and anthocyanin metabolites
-
Five types of flavonoids (rutin, quercetin, luteolin, apigenin, and kaempferol) and three types of anthocyanin metabolites (cyanidin-3-O-glucoside chloride, peonidin-3-O-glucoside, and pelargonidin-3-O-glucoside) were quantified using slightly modified techniques as earlier described by Hellström & Mattila[51] and Henry-Kirk et al.[52]. The concentrations of various anthocyanin metabolites were calculated by using the calibration curve of the respective standard. Each measurement was based on the average of three replicates. The linearity range, limits of detection, and quantification were the validation parameters[53]. The retention times, and comparison of UV-Visible spectra and standardization correctly identified the peaks. Quantification has been done using an external standard curve with five points (Table 1).
Table 1. Parameters for the validation of the ultra-performance liquid chromatography (UPLC) technique.
Flavonoid/anthocyanin component Linearity (r2) Slope (y) Response (Sy) Sy/y LOD* (µg·mL−1) LOQ** (µg·mL−1) Rutin 0.999303 0.2737 5.2262 19.0921 63.00 190.92 Luteolin 0.999692 0.2745 4.9727 18.1111 59.76 181.11 Quercetin 0.999667 0.2756 4.6358 16.8164 55.49 168.16 Cyanidin-3-O-glucoside chloride 0.998590 0.2767 4.3319 15.6526 51.65 156.52 Peonidin-3-O-glucoside 0.999506 0.2757 4.6096 16.7147 55.15 167.14 Pelargonidin-3-O-glucoside 0.998351 0.2754 4.7720 17.3254 57.17 173.25 * Limit of detection; ** Limit of quantification. Enzyme extraction and activity assay
-
Enzymes involved in flavonoids metabolism were extracted and measured by utilizing the enzyme activity kits (Solarbio Life Sciences, Beijing, China)[54,55]. The determination of PAL, 4CL, CHS, C4H, GST and UFGT was based on earlier described methods[3,56−59].
RNA extraction and quantitative real-time PCR
-
Based on the transcriptome data of passion fruit at various stages of development, the KEGG metabolic pathway analysis of phenylalanine, flavonoids, and isoflavones found in passion fruit was used to identify differential candidate sequences of PAL, C4H, 4CL, CHS, and GST. These sequences were found to be significantly different from one another. The local BLAST screening of homologous genes was carried out using the BioEdit program (version 7.2). The genes that had been retrieved in a preliminary manner were then sent to NCBI for BLAST comparison and SMART (
http://smart.embl-heidelberg.de/ ) conserved domain analysis in order to exclude the preliminary candidate genes. The genes were compared with those that were found in the previously published genome of the passion fruit (http://ftp.cngb.org/pub/CNSA/data3/CNP0001287/CNS0275691/CNA0017758/ ). The RT-qPCR specific primers were built with the help of the online program Primer 5 by utilizing the sequence information from the unigenes in the transcriptome[60] (Supplemental Table S1). The RNA extraction from yellow and purple passion fruit was carried out in accordance with the kit's instructions (Tiangen Biotech Co., Ltd., Beijing, China). Using TaKaRa's quantitative reverse transcription kit, the first strand of cDNA was generated. Fluorescence quantitative PCR was carried out with the assistance of a LightCycler® 96 quantitative equipment (Roche Applied Science, Penzberg, Germany).In the reaction mixture, there was 0.25 µM of each primer, 1 µL of cDNA, and 10 µL of 2 × RealStar Green Fast Mixture (GenStar, Beijing, China). The total volume of the reaction mixture was 20 µL, and water was added to make it to that amount. The following describes the criteria for cycling: a temperature of 95 °C maintained for two minutes, followed by 40 cycles at 95 °C maintained for 5 seconds and a temperature of 60 °C maintained for 30 seconds. As an internal reference, the 60 S ribosomal protein was analyzed, and relative gene expression was determined by using the 2−ΔΔCᴛ technique[61]. For each sample, three independent biological replicates were analyzed.
Statistical data analysis
-
The data regarding total flavonoids, total anthocyanins, proanthocyanins, enzymes assay, and gene expressions were compared using Student's t-test between 'yellow' and 'purple' passion fruit for each developmental stage. While, the data regarding flavonoids and anthocyanin components were compared using Fisher's LSD technique through SPSS statistics 21.0 (IBM Inc., New York, USA). OriginPro2021 software (
www.originlab.com/2021 ) was utilized to create heat maps, and the 2−ΔΔCᴛ method was employed to calculate and analyze quantitative data. -
At every maturation stage that was evaluated, the total flavonoids found in purple passion fruit were considerably (p ≤ 0.05) higher than those found in yellow passion fruit, with the exception of the fruitlet stage. The ripening stage of purple passion fruit was found to have the highest concentration of flavonoids (1.44 mg·g−1 FW) (Fig. 2a). While, the anthocyanin content in the pulp of purple passion fruit experienced a sharp increase after the veraison stage and reached its maximum level (0.03 mg·g−1 FW) at the ripening stage. The anthocyanin content in the pulp of yellow passion fruit experienced an increase until the maturation stage and then a decrease after that stage (Fig. 2b). The amount of procyanidin in the fruit steadily rose as it matured, and this trend was seen in both cultivars. The largest amount of procyanidin (0.51 mg·g−1 FW) was found in purple passion fruit after it had reached the ripening stage, although it was not statistically different from the amount found in yellow passion fruit (Fig. 2c).
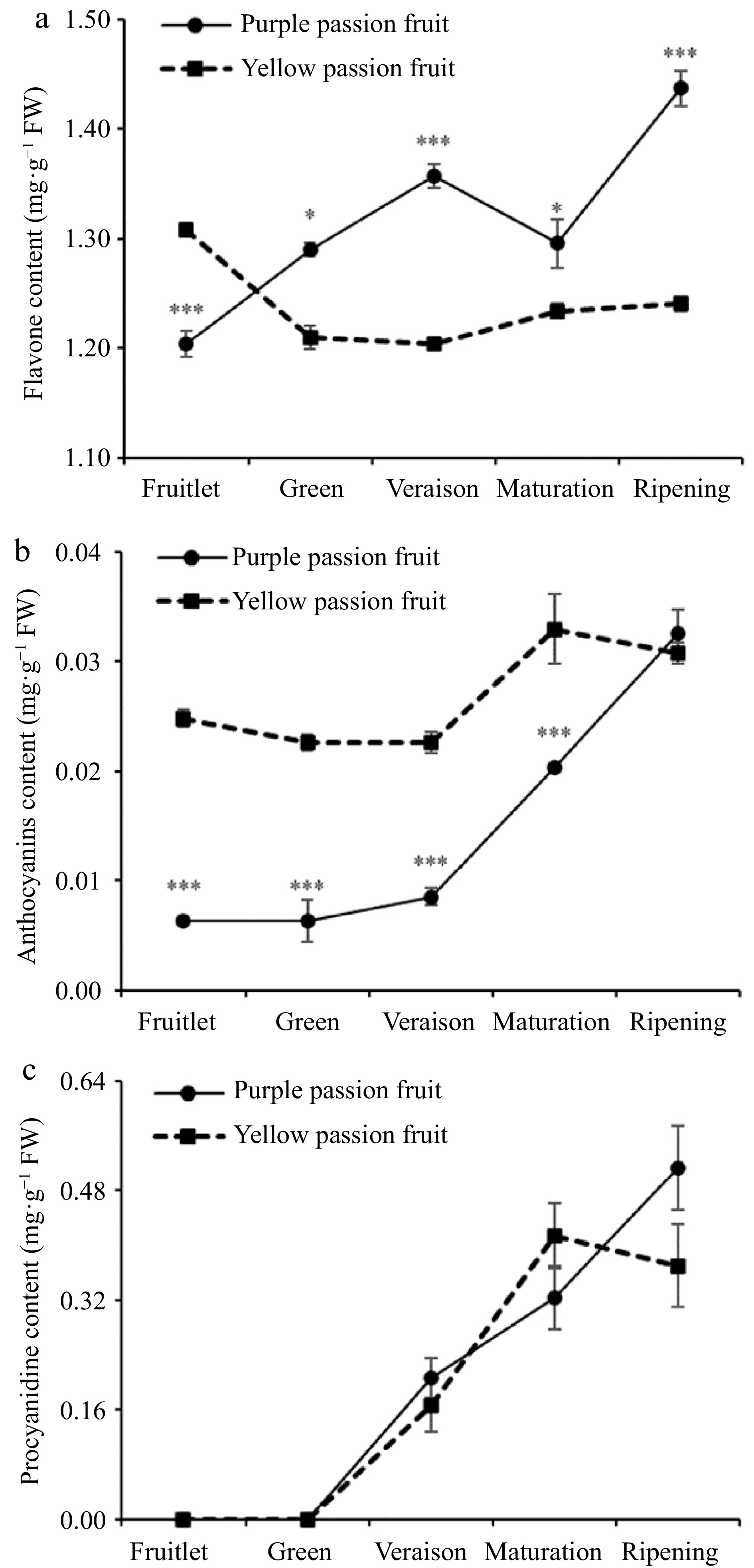
Figure 2.
Alterations in the levels of (a) total flavonoids, (b) anthocyanin, and (c) procyanidins found in the pulp of yellow and purple passion fruit as the fruit matures. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
Anthocyanin and flavonoid metabolites
-
Five types of flavonoids (rutin, quercetin, luteolin, apigenin, and kaempferol) and three types of anthocyanin metabolites (cyanidin-3-O-glucoside chloride, peonidin-3-O-glucoside, and pelargonidin-3-O-glucoside) were found in the fruits of purple and yellow passion fruits. Apigenin and kaempferol, on the other hand, were not found in every single sample of fruit; nonetheless, the other three flavonoids were found in the fruit pulp of both varieties.
The quantity of flavonoid and anthocyanin components that were measured in purple passion fruit was much greater than that of yellow passion fruit, with the exception of rutin and luteolin. The contents of six components in fruit pulp of purple passion fruit steadily rose with the growth of the fruit, with the exception of luteolin and quercetin, both of which had a minor reduction in concentration during the green fruit stage. In purple passion fruit, the quantity of rutin reached its highest level during the ripening stage, which was 77.33 ng·g−1. On the other hand, the highest amounts of luteolin (7.43 ng·g−1) and quercetin (32.47 ng·g−1) were found at maturation stage. At ripening stage, it was found that purple passion fruit had the highest levels of cyanidin-3-O-glucoside chloride (16.79 ng·g−1) and pelargonidin-3-O-glucoside (3.36 ng·g−1). On the other hand, the level of rutin and quercetin grew in the yellow passion fruit at the veraison stage after having decreased during the green stage, but the content of luteolin continued to drop throughout the growth of the fruit. There were no anthocyanin components found in yellow passion fruit, with the exception of cyanidin-3-O-glucoside chloride during the fruitlet stage, and pelargonidin-3-O-glucoside during both the fruitlet stage and the ripening stage (Table 2).
Table 2. Changes in the concentration of flavonoid and anthocyanin metabolites in developing purple and yellow passion fruits.
Cultivar Fruiting stage Cyanidin-3-O-
glucoside chloride (ng·g−1)Peonidin-3-O-glucoside (ng·g−1) Pelargonidin-3-O-glucoside (ng·g−1) Rutin (ng·g−1) Luteolin (ng·g−1) Quercetin (ng·g−1) Purple Fruitlet 3.24 ± 1.89 b 1.14 ± 0.21 a 2.63 ± 0.19 c 48.56 ± 2.12 c 9.85 ± 0.35 a 2.66 ± 0.15 c Green 0 0 0 47.43 ± 5.77 c 0 0.47 ± 0.41 c Veraison 0.84 ± 0.21 b 0 2.84 ± 0.00 bc 61.69 ± 5.42 b 4.97 ± 0.00 c 1.14 ± 0.08 c Maturation 4.95 ± 1.50 b 0 2.96 ± 0.20 b 72.57 ± 1.28 a 7.43 ± 2.03 ab 32.47 ± 1.00 a Ripening 16.79 ± 8.21 a 1.04 ± 1.05 a 3.36 ± 0.23 a 77.33 ± 9.52 a 6.23 ± 2.18 c 25.67 ± 3.06 b Yellow Fruitlet 1.60 ± 0.68 a 0 2.54 ± 0.09 b 71.14 ± 11.71 a 134.39 ± 61.28 a 2.13 ± 1.48 b Green 0 0 0 33.97 ± 3.65 b 6.54 ± 0.19 b 0 Veraison 0 0 0 41.84 ± 10.74 b 5.56 ± 0.36 b 6.61 ± 3.60 a Maturation 0 0 0 41.66 ± 5.27 b 5.12 ± 0.19 b 1.43 ± 0.12 b Ripening 0 0 2.84 ± 0.12 a 45.82 ± 9.38 b 5.25 ± 0.41 b 2.54 ± 0.12 b According to Fisher’s LSD, Same lowercase letters indicate non-significant difference (p ≤ 0.05) between different growth stages. Enzyme assay
-
The PAL activity in the pulp of both cultivars peaked during green stage (41.59 and 45.41 U·Kg−1 FW in purple and yellow passion fruit, respectively), and then decreased at veraison stage, and again surged with fruit maturity. At every stage, with the exception of the fruitlet, the PAL activity was significantly higher in yellow passion fruit than it was in purple passion fruit (Fig. 3a). Both cultivars had significantly different levels of C4H enzyme activity at each stage, as shown by a p value of less than 0.001. The C4H enzyme activity in purple passion fruit reached its highest during the maturation stage (230.18 U·Kg−1 FW), whereas yellow passion fruit reached its peak at the veraison stage (235.96 U·Kg−1 FW) (Fig. 3b). There was no noticeable change in 4CL activity between the fruitlet and maturation stage in yellow passion fruit. During fruit growth, there was an increase in 4CL activity initially, followed by a drop; this pattern continued throughout fruit development. At the veraison stage, the activity of 4CL in purple passion fruit dropped to its lowest level (58.69 U·Kg−1 FW). This activity then rose at the ripening stage. Purple passion fruit had 4CL activity that was substantially (p ≤ 0.001) greater than that of yellow passion fruit (Fig. 3c). The CHS activity of the pulp of both passion fruit cultivars varied from 69.83 to 90.80 U·Kg−1 FW. At fruitlet stage, the purple passion fruit showed the highest level of activity. Significant variation was observed in the CHS activity of both purple and yellow passion fruits (Fig. 3d). Although there was significant difference between both cultivars at fruitlet, veraison, and ripening stage (p ≤ 0.05, 0.01, and 0.001, respectively) (Fig. 3e), the shifting pattern of UFGT activity in the pulp of both cultivars was the same throughout fruit maturity. The GST activity between purple and yellow passion fruit pulp also showed substantial variations at green, maturation, and ripening stage (Fig. 3f).
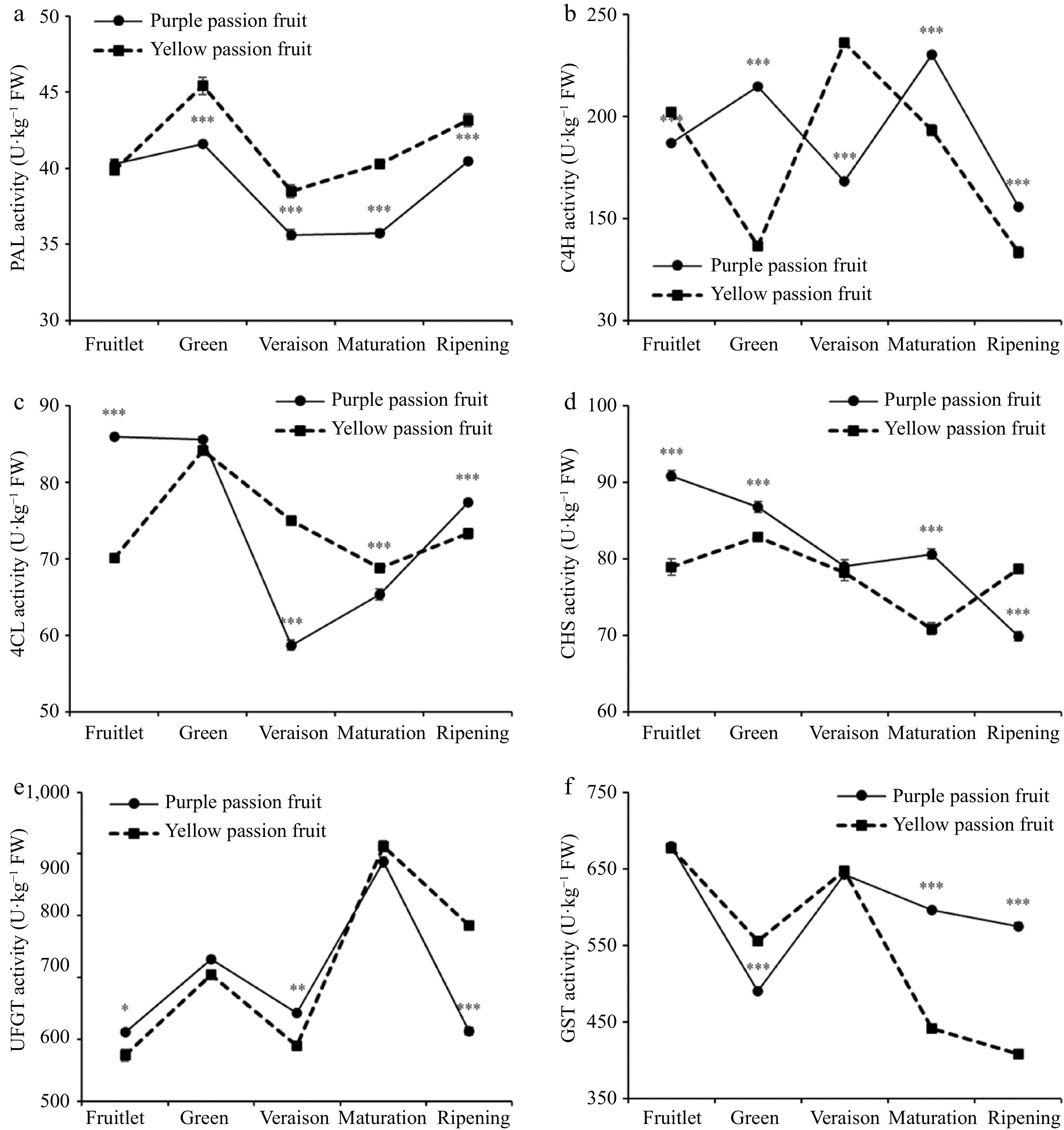
Figure 3.
Alterations in the PAL, C4H, 4CL, UFGT and GST activities of the purple and yellow passion fruits throughout the fruit's growth and development. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
Expression profiling of genes regulating enzyme activities for anthocyanin metabolism
-
The PePAL expression patterns in the pulp of purple and yellow passion fruits were different, and the difference of expression levels was also large. The PePAL1, PePAL2, and PePAL4 genes were significantly differentially expressed at different fruit maturity stages (Fig. 4). The expression level of PeC4H in purple passion fruit decreased at green stage and then increased at ripening. With exception of the fruitlet stage, the expression level of this gene in yellow passion fruit was substantially higher than that in purple passion fruit pulp (Fig. 5).
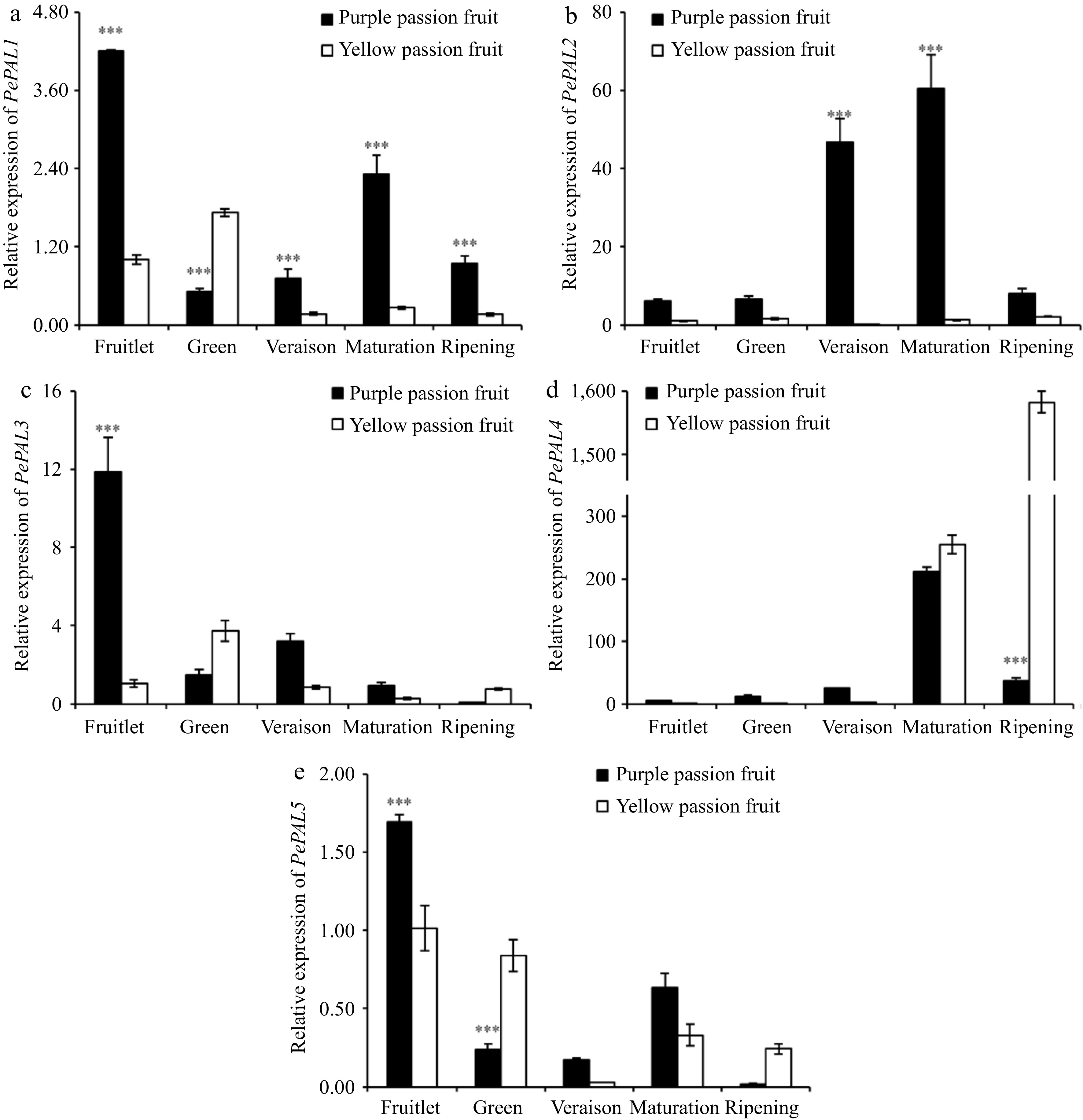
Figure 4.
Relative expressions of PAL genes in fruit pulp of purple and yellow passion fruit during fruit development. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.

Figure 5.
Relative expression of C4H gene in fruit pulp of purple and yellow passion fruit during fruit development. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
The Pe4CL1 and Pe4CL5 in purple and yellow fruits increased at green stage and then decreased, and their expressions were significantly different (p ≤ 0.05). The Pe4CL2 and Pe4CL3 showed an 'L' type change trend and were significantly differentially expressed only at the first two examined maturity stages. At ripening stage, yellow passion fruits' expression of Pe4CL4 was significantly higher than that of purple fruit. The change pattern and expression of Pe4CL4 in other stages were consistent (increased with fruit ripening). The expression patterns of Pe4CL6 and Pe4CL7 in purple and yellow passion fruits were also significantly different. At maturation stage, yellow and purple passion fruit exhibited overexpression of Pe4CL6 and Pe4CL7, respectively (Fig. 6).
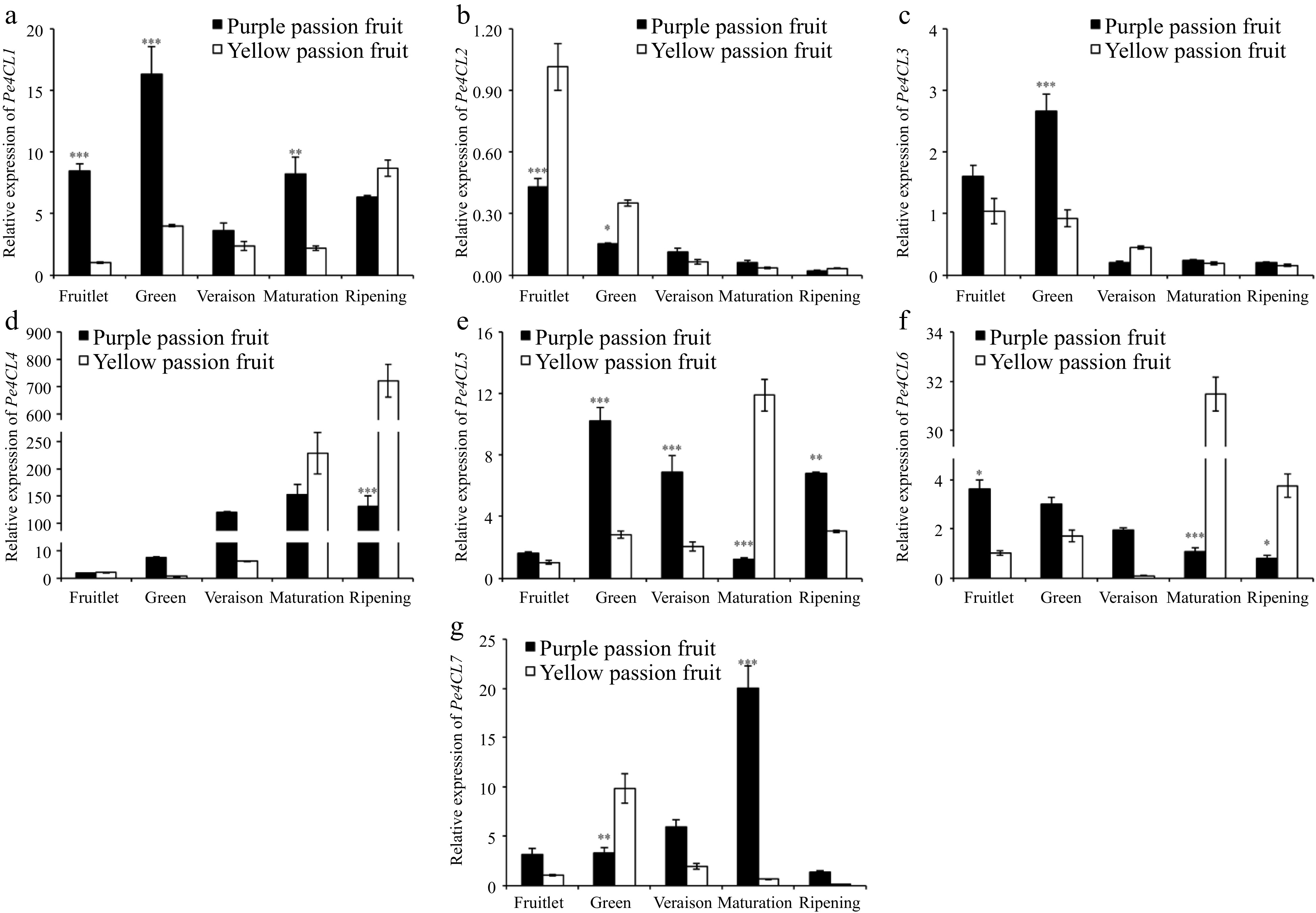
Figure 6.
Relative expressions of 4CL genes in fruit pulp of purple and yellow passion fruit during fruit development. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
In purple passion fruit, the expression of PeCHS1 surged at green stage and then declined with fruit maturity, while it continuously decreased after fruitlet stage in yellow passion fruit. The expression patterns of PeCHS2 and PeCHS3 in fruit pulp were similar to those in fruit peel[3]. Compared to yellow passion fruit, the expression level of these genes in purple passion fruit pulp was significantly higher (Fig. 7).
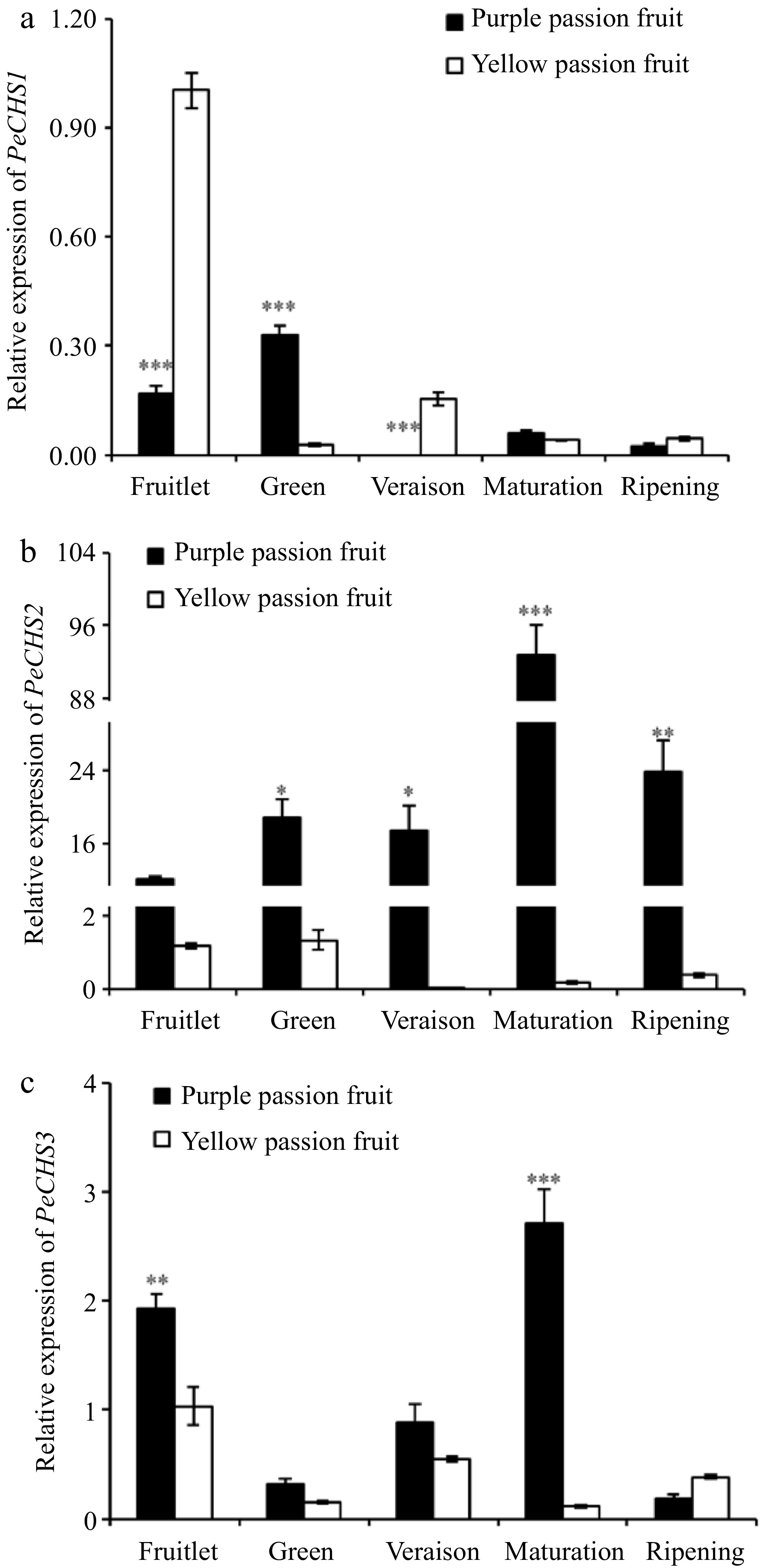
Figure 7.
Relative expressions of CHS genes in fruit pulp of purple and yellow passion fruit during fruit development. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
The expression patterns of PeUFGT1 and PeUFGT2 were significantly different in both cultivars. Purple passion fruit showed overexpression of PeUFGT1 as compared to yellow passion fruit. Compared to the yellow passion fruit, PeUFGT2 expression also remained high in purple passion fruit at maturation stage. However, during ripening stage, its expression reduced 3.5-times in purple passion fruit (Fig. 8).

Figure 8.
Relative expressions of UFGT genes in fruit pulp of purple and yellow passion fruit during fruit development. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
The PeGST1, PeGST4, PeGST6 and PeGST7 genes expression increased at earlier growth stages and then decreased during fruit development. The yellow passion fruit exhibits higher PeGST1-3 expression than the purple passion fruit, while PeGST4, 5, and 7 showed a higher expression level in purple passion fruits (Fig. 9).

Figure 9.
Relative expression of GST genes in fruit pulp of purple and yellow passion fruit during fruit development. Vertical bars indicate means ± SD (n = 3, 5 fruits per replicate). According to Student's t-test, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
Correlation between anthocyanin components and key enzymes involved in its metabolism
-
'Total flavonoids, anthocyanins and procyanidins' and 'flavonoid and anthocyanin metabolites' of the purple passion fruit pulp had a positive correlation with each other. The contents of metabolites and components were negatively correlated with CHS, 4CL, and UFGT, and positively correlated with GST, but not strongly correlated with PAL. There was a significant positive correlation between anthocyanin, rutin, and cyanidin-3-O-glucoside chloride (p ≤ 0.05), and a highly significant positive correlation between proanthocyanin and rutin (p ≤ 0.01). There was a negative association of CHS enzyme activity with anthocyanin metabolites. There was also a significant correlation between PAL and 4CL, C4H, and UFGT (p ≤ 0.05) (Fig. 10a). In yellow passion fruit, there was a highly significant positive correlation (p ≤ 0.01) between procyanidin and rutin content, while highly significant negative correlation (p ≤ 0.01) between flavone content and CHS enzyme activity (Fig. 10b).

Figure 10.
Correlation between 'total flavonoids, anthocyanins and procyanidins', 'flavonoid and anthocyanin metabolites' and 'key enzymes involved in flavonoids metabolism' in the pulp of (a) purple and (b) yellow passion fruit. Following Pearson (n) method, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
Correlation between 'anthocyanin components and key enzymes involved in its metabolism' and 'relative expressions of related genes'
-
In purple passion fruit, most genes were negatively correlated with anthocyanin components, except Pe4CL4, which was positively correlated with most components (Fig. 11a). PeCHS1 gene in yellow passion fruit pulp (Fig. 11b) was positively associated with anthocyanin components, which was contrary to the correlation analysis of purple passion fruit pulp. PeGST7 was negatively correlated with metabolite content in the pulp of both cultivars. The correlation between enzymes activities and the expression levels of corresponding structural genes was not obvious, and activities of some enzymes were significantly correlated with the corresponding genes of other enzymes in the metabolic pathway. The activity of CHS was negatively correlated with the expressions of Pe4CL1, PE4Cl2, PeCHS2, and PeCHS3, but positively correlated with PeGST4 in purple passion fruit. The negative correlation between 4CL enzyme activity and expressions of 4CL genes was obvious. Similarly, the activities of UFGT and 4CL enzymes, and expressions of their metabolic genes in both passion fruit cultivars showed a strong connection with each other (Fig. 11).
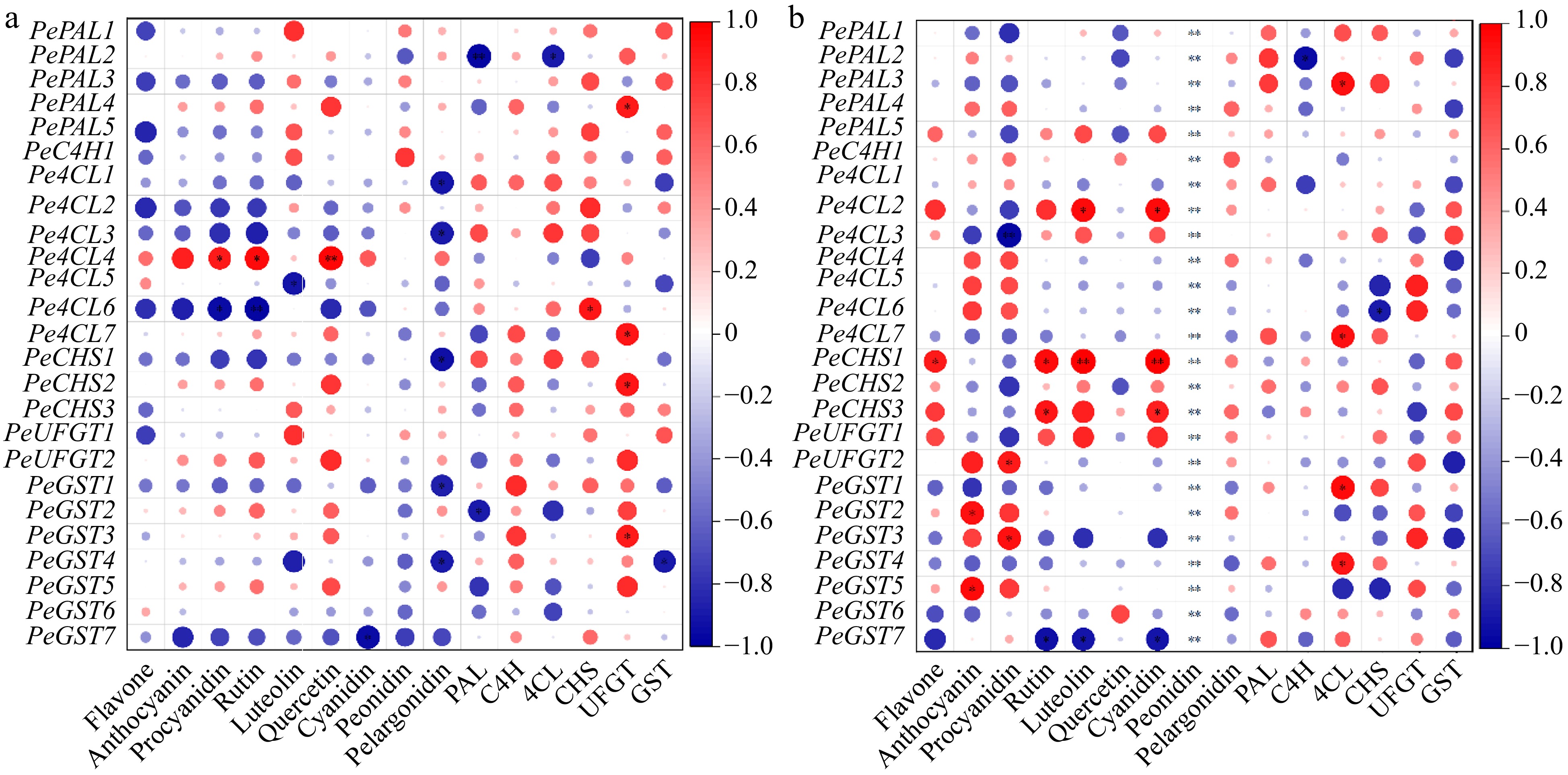
Figure 11.
The correlation analysis between 'anthocyanins components and key enzymes involved in its metabolism' and 'relative expressions of related genes' in fruit pulp of (a) purple and (b) yellow passion fruit. Following Pearson (n) method, the significance is indicated by the symbols *, **, and *** when the corresponding values for p are less than 0.05, 0.01, and 0.001, respectively.
-
Flavonoids are the widely distributed phenolic compounds that are present in almost all plant parts and the primary pigment in flowers and fruits[62−65]. In addition to this, they provide a significant contribution to the secondary antioxidant defense system, which protects the organism from a variety of biotic and abiotic challenges[66]. Flavonoids are located in the nucleus of the mesophyll cells as well as the core of the ROS generating process[67]. There have been reports of flavonoid components in leaves[68], fruit peel[69], and pulp[70] of passion fruit. In a recent study, there were evident differences in the amounts of flavonoids, anthocyanins, and procyanidins in fruit peel between purple and yellow passion fruit[3]. In the present study, purple passion fruit had a more markedly higher flavonoid content than yellow passion fruit, and the maximal difference was observed at ripening stage (Fig. 2). Total flavonoid content in peel was significantly higher than that in pulp of passion fruit[3] (Fig. 2a) during the whole development process, which was in line with the results in jujube[71,72].
In the pulp of purple and yellow passion fruits, UPLC-MS was utilized to identify three anthocyanin components (cyanidin-3-O-glucoside, peonidin-3-O-glucoside, and pelargonidin-3-O-glucoside) and three flavonoid components (rutin, luteolin, and quercetin) (Table 2). While apigenin and kaempferol were found in abundance in passion fruit leaves, they were nearly undetectable in fruits (unpublished data). When researching the antioxidant activity of passion fruit leaves, Shi et al.[3] found several types of apigenin compounds. The amount of flavonoid and anthocyanin chemicals found in the pulp of purple passion fruit throughout fruit development was much greater than that of yellow passion fruit. According to some earlier research, rutin had the greatest content of all the flavonoids that could be discovered[73,74]. The content of cyanidin-3-O-glucoside chloride has been thought to be the criteria to quantitate the anthocyanins in many crops[68,75,76], however, in current study, we found that peonidin-3-glucoside was the most abundant anthocyanin component in pulp of purple and yellow passion fruit at ripening stage (Table 2). The pulp of the fruits of both cultivars was found to have the maximum concentration of luteolin during the earliest phases of fruit ripening. Argentieri et al.[77] have found rich luteolin in the rare passion fruit variety (Passiflora loefgrenii Vitta.), making it a good choice for biopharmaceuticals. Compared to other growth stages, the luteolin content at fruitlet stage in the pulp of yellow passion fruit was much higher, which can be further studied or used as an important period for the extraction of luteolin.
It is known that different enzymes can catalyze the conversion of flavonoids into anthocyanins and other flavonoid compounds[78]. Phenylalanine is the direct precursor for the synthesis of flavonoid, and this process begins with the transformation of phenylalanine to 4-coumaryl CoA. PAL, C4H, and 4CL are the main regulatory enzymes involved in this process[79]. Subsequently, 4-coumaryl CoA and 3 malonyl CoA are converted to dihydroxyflavone, which is the pivotal step in the metabolism of flavonoids. The activities of enzymes i.e., CHS, CHI and F3H regulate this reaction[80]. Then, the unmodified anthocyanins were synthesized[81]. Finally, flavonoids would be transported into vacuole by GST after modified by glycosyltransferase (GT)[82]. During the process of passion fruit growth and development, the enzyme activity changed in a complicated manner. Although there were similarities in the variable patterns of PAL activity in purple and yellow passion fruit, there were significant differences in the degree of the variations (Fig. 3). It has been shown that the PAL activity of two different cultivars of the rapeseed (Brassica napus) plant differs. As a result of these differences in PAL activity, the biological activities that govern separate metabolic pathways are also distinct[83]. Additionally, both of the evaluated enzymes' trends and levels of activity varied in the pulp of the both varieties of passion fruit (Fig. 3). The PAL, CHS, C4H, and 4CL genes were responsible for the plant's stringent flavonoid biosynthesis regulation[84]. The RT-qPCR analysis of the structural genes of the relevant enzymes responsible for the anthocyanin metabolism reveald that they were differently expressed in the flesh of both cultivars (Figs 4−9). The maximally expressed genes in fruit pulp of both passion fruit cultivars were PePAL1,2,4,6,7, PeC4H1, PeCHS1, Pe4CL6,7, and PeGST1-5,7. Overall, the differences in the activities of these enzymes and transcript levels of corresponding structural genes at different developmental stages resulted in the differences of anthocyanin metabolites in passion fruit, and more experiments will be required to verify the specific action mechanism in the future.
In each of the examined cultivars of passion fruit, the average levels of gene transcripts, enzyme activity, and metabolites showed distinct developmental patterns, showing a coordinated developmental control of the whole system to create distinct pools of end products at each stage. Correlation analysis showed that this developmental influence predominated over the genetic in terms of gene expression and metabolite levels (Figs 10, 11). The two most prevalent types of passion fruit flavonoids, flavan-3-ols (such as rutin, luteolin, and quercetin), and anthocyanins, showed the most obvious distinction (i.e., cyanidin-3-O-glucoside chloride, peonidin-3-O-glucoside, and pelargonidin-3-O-glucoside). Later stages showed a substantial accumulation of flavan-3-ols, perhaps to shield ripe fruits from biotic and abiotic influences[41,85]. On the other hand, anthocyanin synthesis began early and peaked at the full ripening stage, acting as a visual lure for animals to eat fruit. This pattern of growth is in line with what has been seen in strawberry fruits[86,87]. The consistency between average gene and enzyme expression patterns, as well as the synchronized expression of structural genes with transcript peaks at early (PAL, C4H, and 4CL) and late (CHS, UFGT, and GST), all demonstrate the observed coordination (Figs 3−9). As in Arabidopsis, where such complexes have been shown to comprise at least the PAL, CHS, 4CL, and UFGT proteins, the encoded enzymes are hypothesized to form multi-enzyme complexes that may channel the biosynthetic precursors in an energy-efficient manner to create flavonoid end products[88,89].
According to the findings of a correlation study, the anthocyanin metabolites found in purple passion fruit had a positive correlation with GST enzyme activities and a negative correlation with C4H, 4CL, and UFGT enzyme activities. There existed positive correlation between PAL, C4H and flavonoid content in tobacco[90], while there was a substantial and positive correlation between the CHS expression and the anthocyanin levels in pomegranate[91]. The expression peaks of genes involved in flavonoid synthesis were obtained at early and late phases in grape[92], Vaccinium myrtillus[82] and wild apple (Malus Sylvestris L.)[93]. The correlation between expressions of anthocyanin metabolism-related genes and enzyme activities revealed that PePAL4, PeCHS1, and PeGST7 played a vital role in anthocyanin metabolism in fruit pulp (Fig. 11). Two passion fruit cultivars varied in the accumulation of anthocyanin-associated metabolites, and the difference was attributed not only to enzyme activity but also to the transcript levels of structural genes. According to previous studies, some versatile transcription factor such as MYB[94], BHLH[95], and WD40[96] were also involved. In addition, regulation via microRNA, ubiquitination and phosphorylation on protein expression and modification also exert important function in this process[97].
-
The amount of variation in the anthocyanin content that was assessed in this research was synergistically controlled by the activities of a number of different enzymes, including PAL, C4H, 4CL, CHS, UFGT, and GST. The enzymes i.e., C4H, 4CL, CHS, UFGT, and GST contributed significantly to the buildup of anthocyanin in passion fruit pulp. A significant contribution was made to the anthocyanin metabolism of fruit pulp by the genes PePAL4, PeCHS1, and PeGST7. These findings not only provide fresh perspective on the aspects of anthocyanin metabolism, but they also constitute a great resource for the study that will inevitably be conducted on molecular breeding in passion fruit.
The research was financed by the Department of Science and Technology, Fujian Province, China (2020N0004 and 2020S0056), and the Engineering Research Center of Fujian Province University (G2-KF2006 and G2-KF2001).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Meng Shi, Muhammad Moaaz Ali
- Supplemental Table S1 Sequences of primer pairs used in this study.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Shi M, Ali MM, Sun K, Gull S, Hu X, et al. 2023. Changes in fruit anthocyanins, their biosynthesis-related enzymes and related genes during fruit development of purple and yellow passion fruits. Fruit Research 3:17 doi: 10.48130/FruRes-2023-0017 

Changes in fruit anthocyanins, their biosynthesis-related enzymes and related genes during fruit development of purple and yellow passion fruits
- Received: 12 April 2023
- Accepted: 25 June 2023
- Published online: 13 July 2023
Abstract: Together with other polyphenols and flavonoids, anthocyanins have the capacity to serve as free radical scavengers against detrimental oxidants including reactive oxygen and nitrogen species. Moreover, the role of anthocyanin pigments as natural fruit colorings is quite common. In this study, the anthocyanin profile of purple and yellow passion fruit was determined at five developmental phases i.e., fruitlet, green, veraison, maturity, and ripening stage. Total flavonoids were abundantly found among other metabolites including anthocyanins and proanthocyanins. Purple passion fruits contained more than 2-times higher flavonoid content than yellow passion fruits at ripening stage. The findings showed that fruit maturation increased the amount of total flavonoids, anthocyanins, and procyanidins in the pulp of both varieties of passion fruit. Correlation analysis revealed that the passion fruit anthocyanin metabolism may be regulated by the enzymes C4H, 4CL, CHS, UFGT, and GST. The metabolism of anthocyanins in passion fruit may be significantly influenced by the genes PePAL4, PeCHS1, and PeGST7. New information from this work will help future research into the fundamental processes controlling the production of anthocyanins in passion fruit.
-
Key words:
- Passiflora edulis Sims. /
- L-phenylalanine ammonia-lyase /
- Rutin /
- UFGT /
- Anthocyanin /
- Cyanidin /
- Gene expression.