









-
Plant leaf is a critical organ as it plays a pivotal role in both photosynthesis and respiration. Leaf morphology is a vital index in plant morphogenesis, where the form and size of leaves determine the plant yield and shape. However, leaf morphology is influenced by various factors such as variety, age, and environment[1,2]. By regulating leaf morphology, it is possible to optimize the plant's light energy utilization, improve its quality and yield, and alter gardening plant shape and ornamental value.
Teosinte branched1/Cycloidea/Proliferating cell factor (TCP) transcription factor family is a distinct group of proteins found in plants, characterized by a conserved structural domain consisting of atypical basic helix-loop-helix (bHLH) motifs spanning approximately 59 amino acids[3]. The TCP domain was initially identified in four genes from three plant species: Teosinte branched1 (TB1) in maize (Zea mays), which plays a crucial role in apical dominance, inflorescence development, and other biological processes[4]; Cycloidea (CYC) in snapdragons (Antirrhinum majus), which controls flower morphology[5]; and PROLIFERATING CELL FACTORS 1/2 (PCF1/2) in rice (Oryza sativa), involved in cell proliferation and growth of meristematic tissues and lateral organs[6]. TCP gene family is generally classified into two subfamilies: class I, also known as the PCF subfamily, and class II, which further branches into the CIN and CYC/TB1 subfamilies[7,8]. TCP genes are widely distributed across various plant species and play crucial roles in multiple growth and development processes. For instance, they are involved in fruit development[9], shoot branching[10], flower morphological regulation[11], leaf morphological regulation[12], hormone signaling[13] and abiotic stress response[14].
A large number of TCP genes have been identified in many plants by whole genome sequencing. In total, 23, 22, 46, and 36 members of TCP transcription factor family have been identified in Arabidopsis thaliana, rice, potato (Solanum tuberosum L.), and maize, respectively[15−18]. These TCP transcription factors play crucial roles in regulating leaf development and morphology in various species. In Arabidopsis, class I TCP genes (AtTCP7, AtTCP8, AtTCP22, and AtTCP23) exhibit mutually interactions and regulate the expression of KNOTTED1-LIKE homeobox (KNOX) genes, thus exerting critical influence over leaf development and cell proliferation[19]. Notably, the generation of a pentuple mutant of these genes leads to significant alterations in leaf development[19]. Moreover, AtTCP14 and AtTCP15 interact with SPINDLY (SPY) to modulate cytokinin (CK) biosynthesis or degradation in leaves, thereby influencing cell proliferation during leaf development[20]. Mutations in AtTCP14 and AtTCP15 genes give rise to phenotypes characterized by wider leaf bases, shorter petioles, and upwardly curved leaf edges[21]. Additionally, class II TCP genes also demonstrate involvement in regulating the cell proliferation and differentiation during leaf development. AtTCP1 plays a significant role in leaf shape regulation by participating in strigolactone signaling[13]. AtTCP4, on the other hand, inhibits the growth of leaf surface trichomes by activating GLABROUS INFLORESCENCE STEMS (GIS)[22], and AtTCP5 contributes to the regulation of leaf margin morphology by activating BEL-like transcription factors[23]. Similar observations of TCP gene involvement in leaf morphology regulation have been reported in other ornamental plants. For instance, mutations in the CIN gene in snapdragons lead to excessive proliferation of leaf marginal cells, resulting in leaf margin undulation[24]. Similarly, mutations in the LANCEOLATE (LA) gene, the tomato (Solanum lycopersicum) ortholog of Arabidopsis TCP4, result in the transformation of compound leaves into small, simple leaves[25].
Osmanthus fragrans, a plant species of significant economic and ornamental value, has been cultivated in China for centuries and holds a distinguished reputation as one of the ten most famous traditional flowers. Besides its sweet fragrance, O. fragrans exhibits an aesthetically pleasing tree shape, making it an attractive choice for landscaping purposes. Previous studies have indicated that the critical role of TCP transcription factors in the regulation of plant leaf development and morphology in most species[19]. However, there is limited research regarding the role of TCP transcription factors in the ornamental plant O. fragrans, and their involvement in the regulation of leaf morphology in this plant remains unclear. In this study, we conducted a comprehensive analysis of the TCP family at the whole genome level in O. fragrans, and examined the expression patterns of class I TCP genes in different tissues of O. fragrans. One class I TCP gene, named OfTCP13, was obtained, showing higher expression in the leaf tissue. To further explore its function, we validated the gene's role in tobacco lines. The outcomes of our investigation offer valuable insights into the understanding of leaf morphogenesis in O. fragrans.
-
The 6–7-year-old O. fragrans cultivar 'Yanhonggui' was selected and cultivated in the O. fragrans Resource Garden of Zhejiang University of Agriculture and Forestry, China (latitude: N30°15′14′′ N, longitude: 119°43′39′ E). Tissue samples, including roots, stems, mature leaves, leaf buds, were collected from different parts of O. fragrans during the vegetative stage, while the flower tissues were harvested in the full flowering stage. Approximately 0.5 g of each tissue sample was harvested and immediately frozen in liquid nitrogen. Then the frozen samples were subsequently stored at −80 °C until further analysis. Three independent biological replicates were included for each tissue sample.
Genome-wide identification of TCP gene family
-
To identify and retrieve all the OfTCP protein sequences in the O. fragrans genome, the TCP conserved domain (PF03634,
http://pfam.xfam.org/ ) was utilized and the online tool HMMER v3.3.2 (http://hmmer.org/) employed for identification with a default e-value. Subsequently, the presence of the TCP-conserved domain in all proteins was re-evaluated using the SMART (http://smart.embl-heidelberg.de/ ) and Batch Web CD-Search of NCBI (www.ncbi.nlm.nih.gov/cdd/? ). For the gene name, 12 OfTCP genes, including OfTCP1 to OfTCP5, and OfTCP7 to OfTCP13, were identified and named by Zhou et al.[26], and these names have been adopted and maintained in our study. As for the remaining OfTCP genes, they have been assigned names based on their corresponding gene IDs in a sequential order. All the OfTCP amino acid sequences were provided in the Supplemental Table S1.In addition, the molecular weight (MW) and the isoelectric point (PI) of all OfTCP proteins were obtained using the Online program ExPASY (
www.expasy.org/tools/ ). Chromosomal localization was analyzed by TBtools software[27]. The construction of the phylogenetic tree was performed using MEGA 11.0 software, employing the neighbor-joining method with a bootstrap value of 1,000, and iTOL.v6 software (https://itol.embl.de/ ) was utilized to visualize the phylogenetic tree. TCP protein sequences for Arabidopsis were downloaded from TAIR (www.arabidopsis.org/ ).Sequence characterization analysis of class I TCP genes
-
Multiple sequence alignment of the class I OfTCP genes was performed using DNAMAN software (
www.lynnon.com/dnaman.html ). To identify motifs of class I TCP proteins, the MEME online program (https://meme-suite.org/meme/tools/meme ) was utilized, with a maximum of ten output motifs selected. The NDA sequence structure of class I OfTCP genes was determined by Gene Structure Display Sever 2.0 (http://gsds.cbi.pku.edu.cn/ ), which provided visual representation of the exon-intron organization. Cis-element analysis was conducted on the 2,000 bp upstream sequences of the class I OfTCP genes using Plantcare database (https://bioinformatics.psb.ugent.be/ ). The full-length amino acid sequences of class I TCP were aligned using MEGA 11.0 software with default parameters, and neighbor-joining phylogenetic trees were constructed with 1,000 bootstrap replicates.RNA extraction and quantitative real-time PCR
-
A total of 0.5 g samples was used for RNA extraction from different tissues of O. fragrans using the RNAprep Pure Plant Plus Kit (Tiangen, China) according to the manufacturer's instructions. Then the extracted RNA samples were treated with RNase-free DNase I (TaKaRa), and their concentration was determined using a Nanodrop 2000 spectrophotometer (Thermo Fisher, USA). The OD260/280 of the RNAs ranged from 1.8 to 2.2, and the OD260/230 was > 1.8. First-strand cDNA was synthesized using HiScript III All-in-one RT SuperMix Perfect for qPCR (Vazyme, China). The expression levels of class I TCP genes in different tissues of O. fragrans were analyzed by quantitative real-time PCR (qRT‒PCR). The reaction system was SYBR Premix Ex Taq (Vazyme, China) 10 µL, forward and reverse primers (10 mM) 0.4 µL, cDNA 2 µL, double distilled H2O (ddH2O) 6.2 µL, reaction program of 95 °C for 30 s, (95 °C for 5 s, 60 °C for 30 s; 95 °C for 5 s) 40 cycles, then 60 °C for 1 min. OfACT and NtACTIN was selected as the reference gene for O. fragrans and Nicotiana tabacum, respectively[28,29]. The relative gene expression was calculated using the 2–ΔΔCᴛ method[30]. All primer sequences are listed in Supplemental Table S2.
Vector construction
-
The full-length cDNA of OfTCP13 was obtained by PCR. The PCR product was purified by a FastPure Gel DNA Extraction Mini Kit (Vazyme, China), ligated to the pMD-18T plasmid (TaKaRa, China) and then sequenced. After PCR amplification using primers with NheI and XhoI restriction sites, the complete coding sequence of OfTCP13 without the stop codon was ligated into the pORE-R4-35AA vector[31] to create a recombinant plasmid (35S::OfTCP13-GFP) for subcellular targeting and transgenic tobacco. After PCR amplification using primers with BamHI and EcoRI restriction sites, the complete coding sequence of OfTCP13 was ligated into the pGBKT7 vector to create a recombinant plasmid (pGBKT7-OfTCP13) for transcriptional activation analysis in yeast cells. The reaction systems were all 2 µL of linearized vector, 4 µL of gene fragment and 4 µL of enzyme, and the procedure used was 50 °C for 10 min and final storage at 4 °C. All primer sequences are listed in Supplemental Table S2.
Subcellular localization
-
The subcellular localization was performed according to previously described procedures[32], with little modifications. The plasmid 35S::OfTCP13-GFP was transformed into Agrobacterium rhizogenes strain GV3101. The transformed strain was mixed well with the nuclear marker 35S::D53- RFP (OD600 = 0.6 per strain, 1:1 ratio). Then the mixture was protected from light and incubated for 3 h before being injected into the leaves of N. benthamiana. As a negative control, a null plasmid (35S::GFP) was also injected . After one day of dark incubation and two days of light incubation, GFP signals were detected using confocal microscopy (Olympus Corporation, Japan).
Transcriptional activation
-
For yeast cell transcriptional activation analysis, the recombinant plasmid pGBKT7-OfTCP13 was transformed into yeast strain AH109 using the lithium acetate method and coated on selective medium plates without tryptophan (SD/-Trp). pGBKT7-OfWRKY57 was used as a positive control, and empty vector pGBKT7 was used as a negative control. Three days later, the colonies were verified by PCR and the positive bacteria were inoculated onto selective medium plates of SD/-Trp, SD/-Trp-Leu-Ade and SD/-Trp-Leu-Ade+X-α-gal. After 3 d, the yeast cells were photographed and observed for growth.
Plant transformation and transgenic tobacco identification
-
The tobacco cultivar Nicotiana tabacum L. 'NC89' was employed for genetic transformation using the method described by Sparkes et al.[33]. All the plants were grown in an incubator at 25 ± 1 °C with a 12 h light/12 h dark photoperiod. First, the recombinant plasmid 35S::OfTCP13-GFP was transformed into A. rhizogenes strain GV3101, and a bacterial solution of A. rhizogenes (OD600 = 0.6) was infected into tobacco leaves for 8 min, and the infected leaves were cultured on a medium containing 4.4 g·L−1 MS + 2.25 mg·L−1 N-(phenylmethyl)-9H-purin-6-amine (6-BA) + 0.3 mg·L−1 1-naphthylacetic acid (NAA) + 8 g·L−1 agar + 30 g·L−1 sucrose. After 2 d of dark culture, the leaves were transferred to a selective medium containing 4.4 g·L−1 MS + 2.25 mg·L−1 6-BA + 0.3 mg·L−1 NAA + 8 g·L−1 agar + 30 g·L−1 sucrose + 500 mg·L−1 carbenicillin (carb) + 100 mg·L−1 kanamycin (kana) until shoots grew. Then the obtained shoots were transferred to a medium containing 4.4 g·L−1 MS + 0.1 mg·L−1 6-BA + 0.01 mg·L−1 NAA + 8 g·L−1 agar+30 g·L−1 sucrose + 500 mg·L−1 carb + 100 mg·L−1 kana for further selection. After 30 d cultivation, the adventitious shoots were transferred to a rooting medium containing 4.4 g·L−1 MS + 8 g·L−1 agar + 30 g·L−1 sucrose until rooting occurred and transplanted into soil. Finally, a total of 1.0 g of leaf tissue was harvested for DNA extraction to identify the positive lines using the Plant Genomic DNA Kit (Tiangen, China). And 0.5 g of leaf tissue was performed for RNA extraction using the RNAprep Pure Plant Plus Kit (Tiangen, China) to analyze gene expression.
Leaf phenotype investigation
-
Three-month-old tobacco lines were selected for phenotypic observation. The lead length and width were measured by vernier scale with nine biological replicates. In order to compare the leaf thickness between transgenic tobacco and WT, the structure of leaves in cross-sections was observed using paraffin sections[33]. The method is as follows, samples were firstly fixed in FAA fixative (ethanol: acetic acid = 3:1), dehydrated in an ethanol series and embedded in paraffin. Samples were cut into 10 µm sections, dewaxed and resin-sealed, and observed using an Axio Imager A2 positive fluorescence microscope (Carl Zeiss, Germany).
Statistical analysis
-
IBM SPSS 20 software (IBM, Armonk, USA) was employed for data analysis, significant differences were determined by Student's t test, and standard error of the mean (SEM, n ≥ 3 replications) was calculated, p < 0.001 (***), p < 0.01 (**) and p < 0.05 (*).
-
A total of 39 TCP genes were identified according to the conserved bHLH domain from O. fragrans genome, and the characteristics of OfTCP amino acid sequences were presented in Supplemental Table S3. The length of 39 OfTCP proteins varied from 203 amino acids (OfTCP19) to 587 amino acids (OfTCP35), molecular weight (MW) ranged from 21.94 kDa (OfTCP19) to 65.57 kDa (OfTCP35), and theoretical isoelectric point (pI) varied from 5.88 (OfTCP7) to 10.16 (OfTCP19). According to phylogenetic tree analysis with Arabidopsis and TCP domain characteristics, OfTCP TFs were clustered into three main groups, and 16, 13, and 10 OfTCPs were contained in the PCF, CIN, and CYC/TB1 group, respectively (Fig. 1). In addition, OfTCP genes distributed on 18 chromosomes except chr8, chr11, chr14, chr17, and chr22 by chromosomal location analysis in O. fragrans (Supplemental Fig. S1).
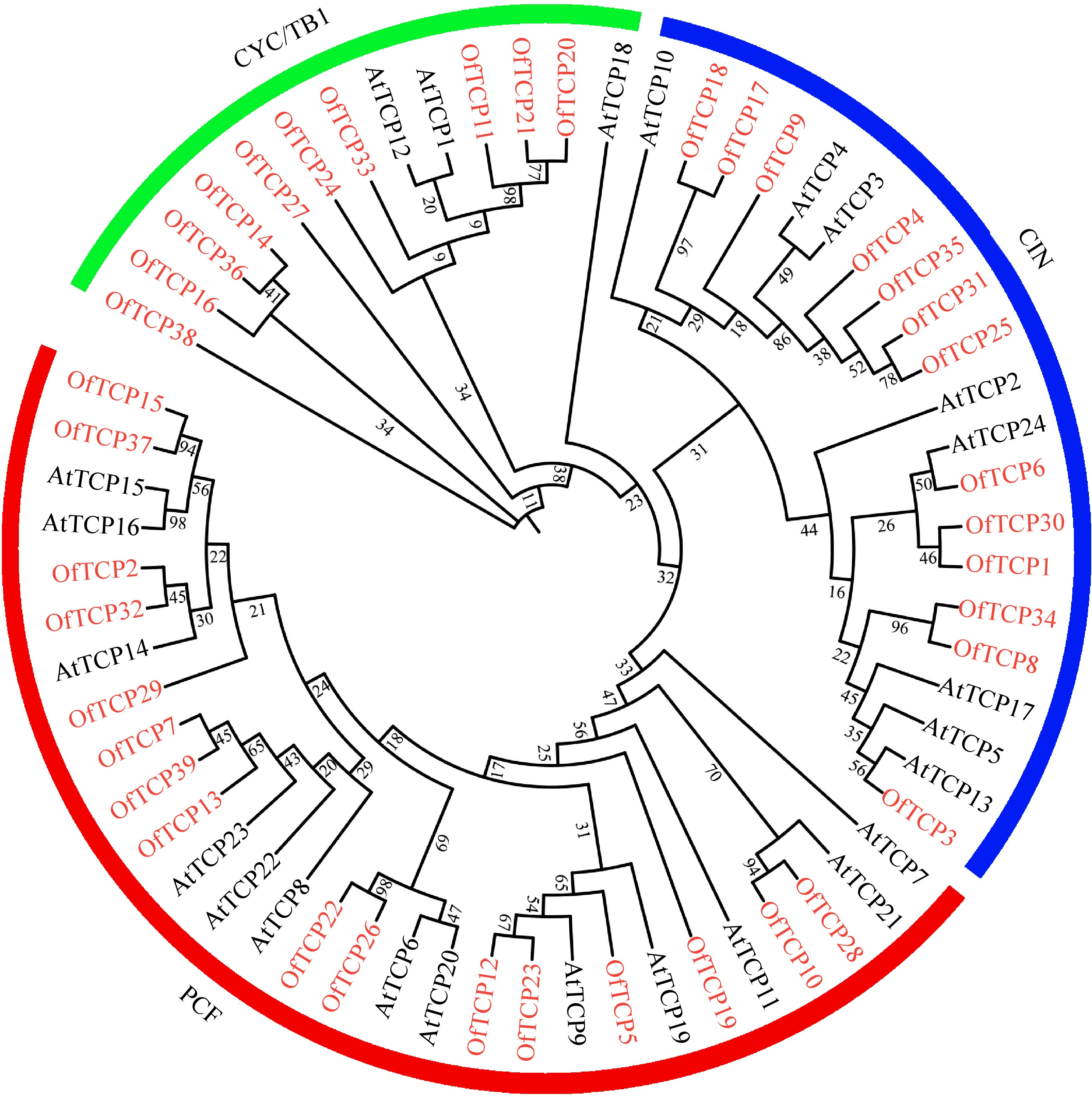
Figure 1.
Phylogenetic analysis of TCP members from Arabidopsis and O. fragrans. Note: Red font represents O. fragrans, black font represents Arabidopsis.
Class I OfTCPs sequence analysis
-
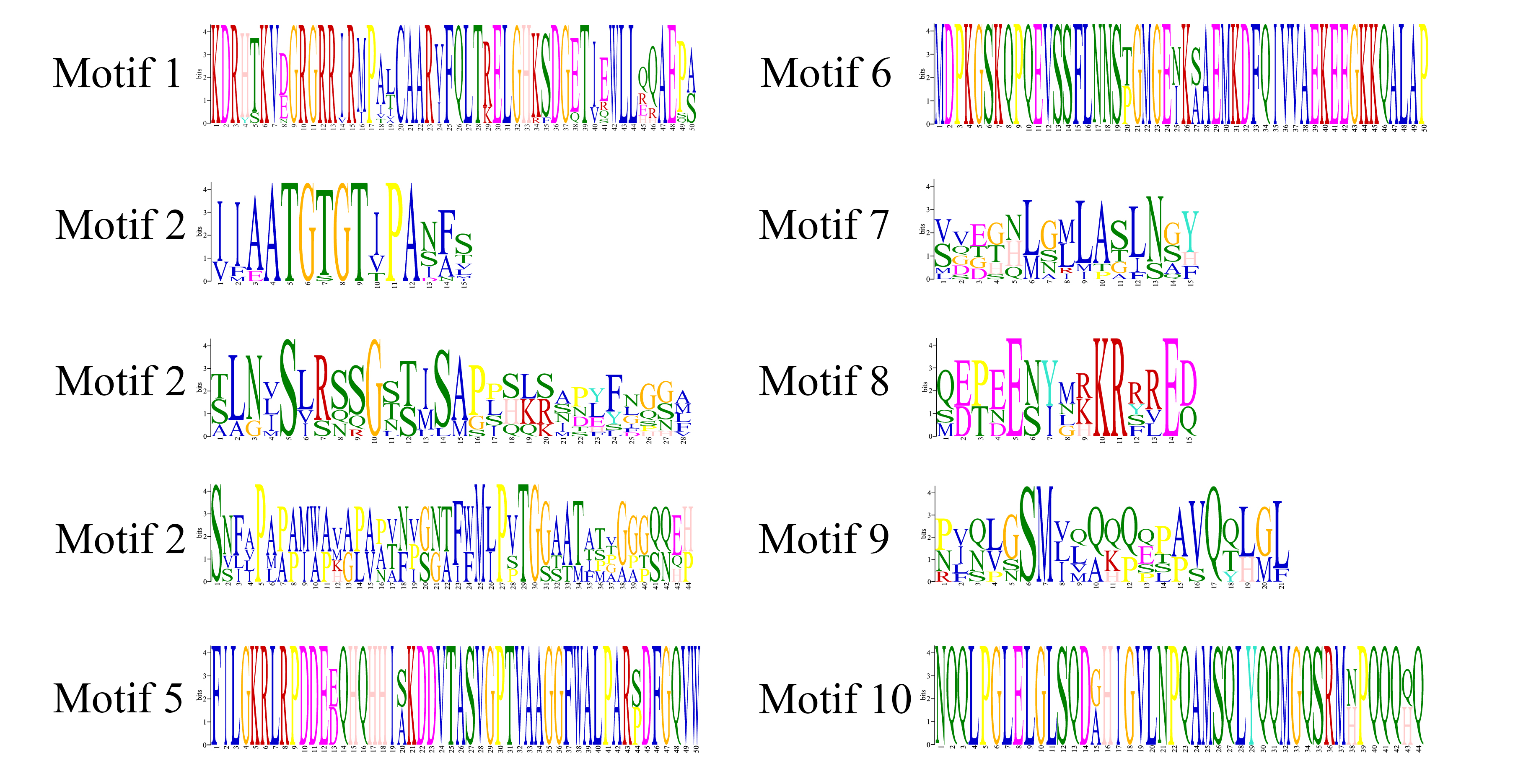
For the sequence characteristics of class I OfTCP proteins analysis, we performed multiple sequence alignment and identified the conserved TCP domain, which consists of four regions: the basic region, helix I, loop, and helix II (Fig. 2a). Analysis of motif composition revealed distinct differences among the class I OfTCP proteins. While motif 1 and motif 2 were present in the all OfTCP proteins, the distribution of the other motifs varied (Fig. 2b & Supplemental Fig. S2). Examination of the gene structures of class I OfTCP genes showed that 14 OfTCPs had no introns, OfTCP5 contained one intron, and OfTCP29 harbored two introns (Fig. 2c). These results suggest that variations in the motif composition among class I OfTCP proteins may be associated with different functional roles. Furthermore, analysis of the promoter sequences of class I OfTCP genes identified four distinct cis-acting elements within the 2,000 bp upstream regions. These elements were found to be involved in hormone response, stress response, light response, and growth and developmental regulation (Supplemental Fig. S3). Among these elements, light-responsive elements accounted for the highest proportion (45%), followed by hormone-responsive elements (34%), stress-responsive elements (18%) and growth and developmental regulatory elements (3%). Therefore, the results implied the expression of class I OfTCP genes is regulated by multiple factors and is involved in various biological processes in O. fragrans.
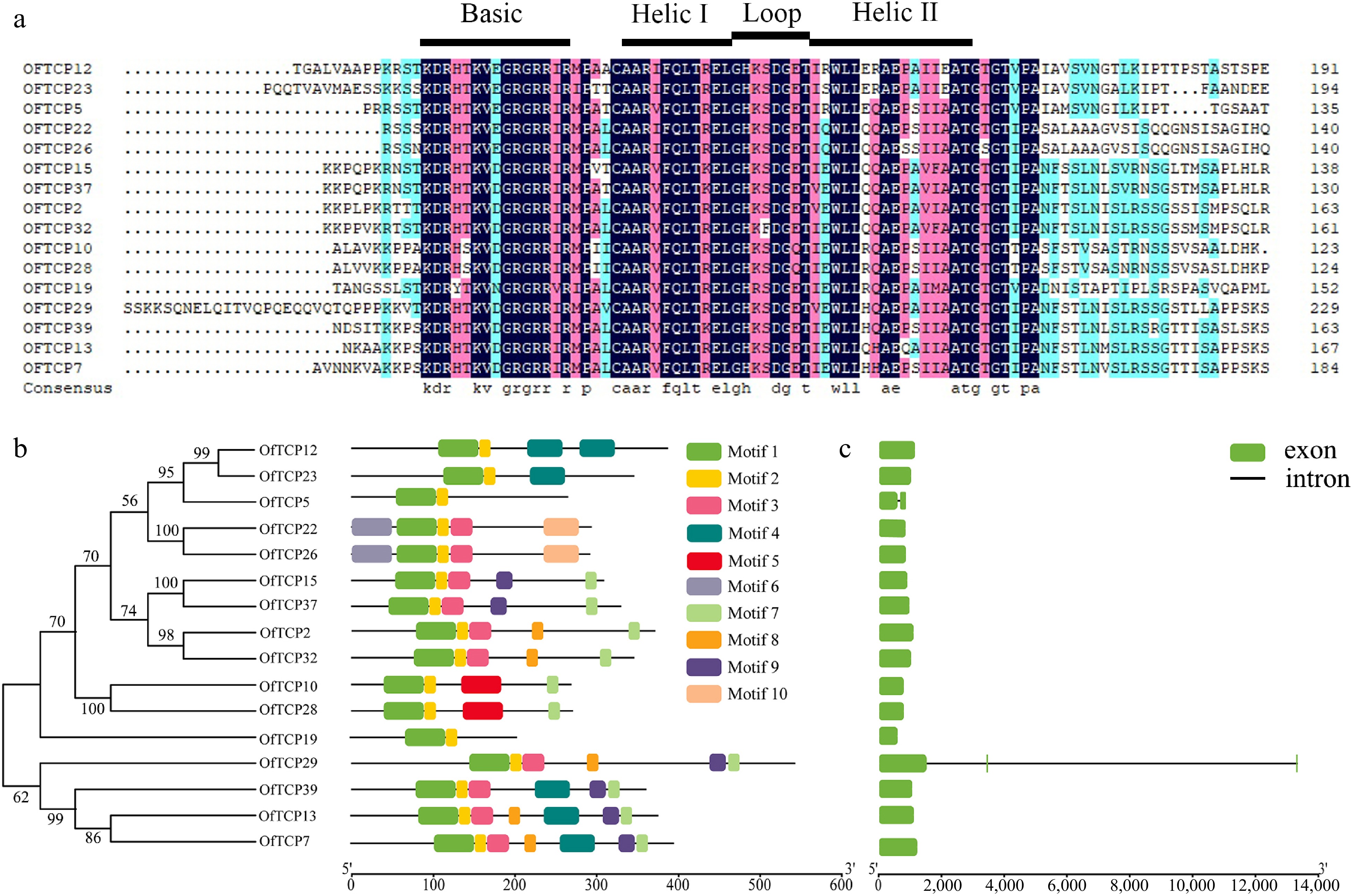
Figure 2.
Analysis of class I TCP gene sequence characteristics in O. fragrans. (a) Alignment of class I OfTCP protein sequences. (b) Motif distribution of class I OfTCPs. (c) Class I OfTCP gene structure analysis.
Expression analysis of class I OfTCPs in different tissues
-
Previous studies have demonstrated that class II TCP genes play an important role in leaf development[34]. To investigate which class I OfTCP gene is prominently expressed in leaf tissue and involved in the leaf development, we examined the expression levels of the class I OfTCP genes in various tissues (root, stem, leaf, flower, and shoot) of O. fragrans using qRT‒PCR. Our results indicated that these class I OfTCP genes are expressed in all tested tissues and could be categorized into four groups based on their expression patterns (Fig. 3). Notably, we found higher expression levels of OfTCP5, OfTCP13, OfTCP22 and OfTCP37 in leaf tissue compared to other tissues, indicating their potential critical roles in regulating of leaf development in O. fragrans. Among these genes, OfTCP13 displayed more than 8.0-fold increase in expression relative to the root tissue, drawing our attention for further investigation in subsequent experiments.
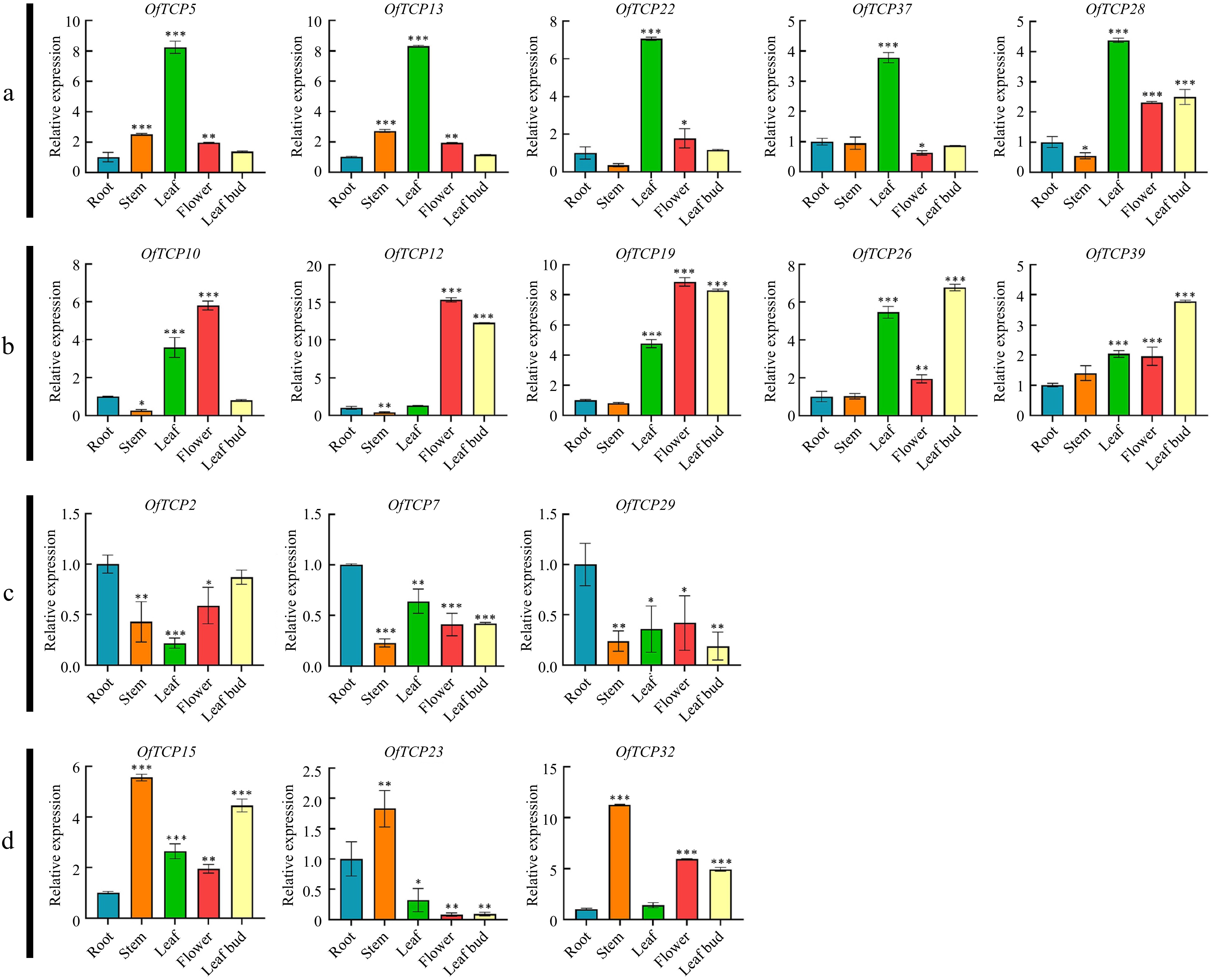
Figure 3.
Expression patterns of class I OfTCP genes in different tissues of O. fragrans. * p < 0.05, ** p < 0.01, *** p < 0.001, t test.
Analysis of OfTCP13 subcellular localization and transcription activation activity
-
To investigate the subcellular localization of OfTCP13, the 35S::OfTCP13-GFP and 35S::D53-RFP plasmids were co-transformed into N. benthamiana leaves (Fig. 4a). The 35S::D53-RFP plasmid served as a positive control for nuclear localization, which expressed a red fluorescent protein specifically targeted to the nucleus. Fluorescence imaging analysis revealed that the OfTCP13-GFP fusion protein was exclusively localized in the nucleus during transient expression. In contrast, the control GFP was evenly distributed throughout the cells. These results suggest that OfTCP13 is a nuclear-localized protein. For the transcription activation activity analysis, the full-length coding sequence (CDS) of OfTCP13 was amplified and fused into the pGBKT7 plasmid. Subsequently, we transformed this construct into the yeast strain AH109. The results demonstrated that the positive control transformed yeast cells turned blue in the presence of X-α-gal, indicating successful transcriptional activation. However, the pGBKT7-OfTCP13 transformed yeast cells, as well as the negative control transformed yeast cells, did not exhibit blue coloration in the presence of X-α-gal (Fig. 4b). These observations suggest that OfTCP13 lacks transcriptional activation activity.
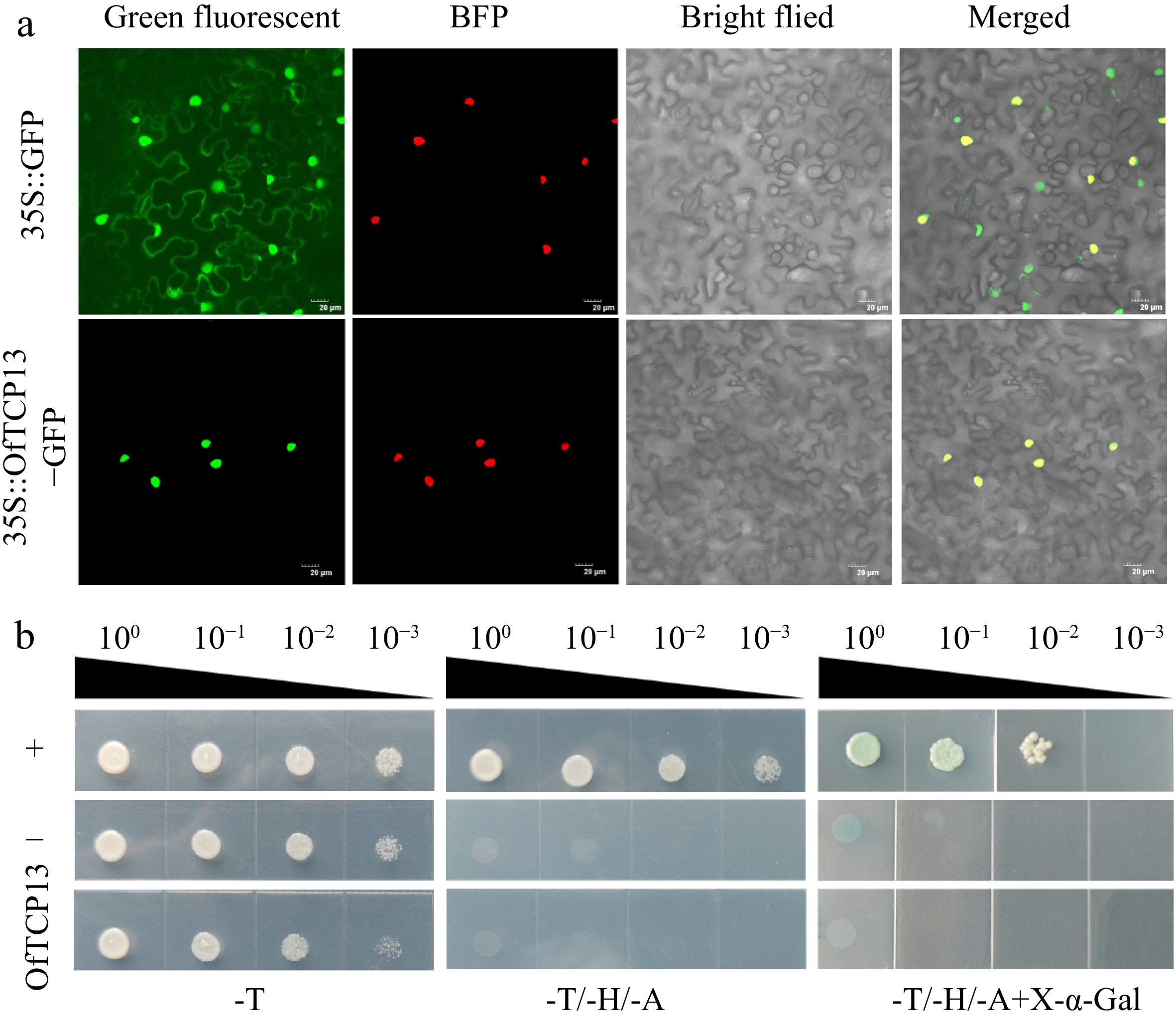
Figure 4.
OfTCP13 subcellular localization and transactivation activity. (a) Subcellular localization of OfTCP13 in N. benthamiana epidermal cells. 35S::GFP is used as a control, 35::D53-RFP is a nuclear marker. Scale bars = 20 µm. (b) Transcriptional activity analysis of OfTCP13 in yeast. Note: + represents positive control, − represents negative control.
OfTCP13 involved in the regulation of leaf shape
-
A 35S::OfTCP13-GFP plasmid was constructed and introduced into tobacco plants via leaf disc transformation (Fig. 5a). The resulting transgenic lines were initially screened by PCR to confirm the presence of the transgene, and a three-month-old transgenic plants E1 were employed for subsequent phenotypic investigation (Fig. 5b, c). Distinct differences in leaf phenotypic traits were observed between the E1 transgenic line and the wild type (WT). Notably, the E1 lines exhibited leaves with a narrowly shape and wavy serrated leaf margins, while the WT leaves displayed spreading leaf margins (Fig. 5d). Moreover, the overexpression OfTCP13 significantly influenced leaf thickness and blade length to width ratio. The leaf thickness measured 0.76 ± 0.08 mm in transgenic lines and 0.53 ± 0.06 mm in the WT, whereas the blade length to width ratios were 0.35 ± 0.24 and 0.28 ± 0.30 in transgenic lines and the WT, respectively (Fig. 5e, f). Then the cross-section of the leaves was further investigated using paraffin sections, we found an increased number of mesophyll cells in the leaf tissue of the overexpressed plants compared to the WT (Fig. 5g). These results suggest that overexpression of OfTCP13 can influence plant leaf morphology.
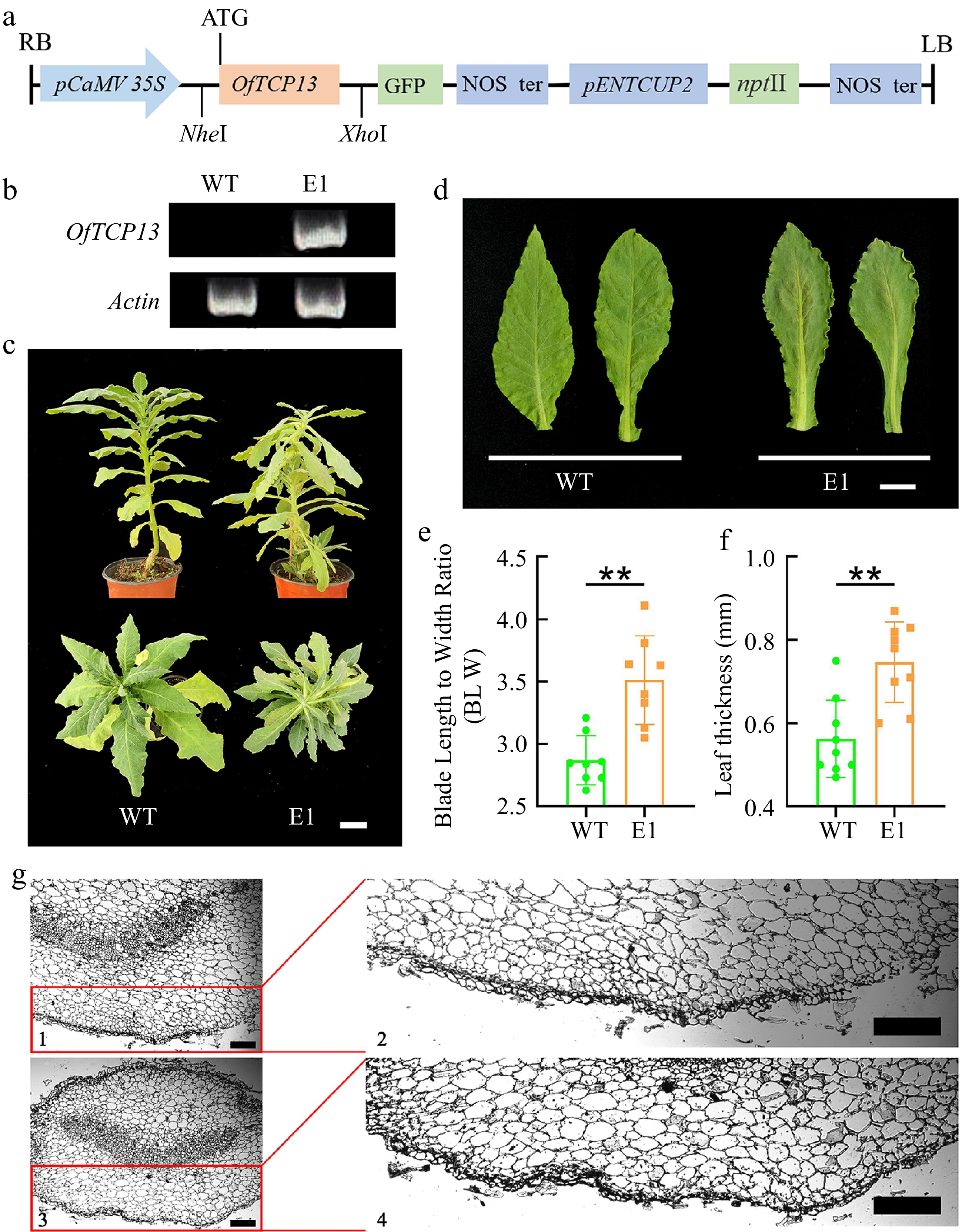
Figure 5.
Overexpression of OfTCP13 affects leaf morphology in tobacco plant. (a) Structure of pORE-R4-OfTCP13 vector for tobacco transformation. (b) Identification of OfTCP13 transgenic lines by PCR analysis (c) Phenotypic characteristics of WT and transgenic tobacco. Scale bars = 5 cm. (d) Leaf morphology of WT and transgenic tobacco. Scale bars = 2 cm. (e) Blade length to width ratio, ** p < 0.01, t test. (f) Leaf thickness, ** p < 0.01, t test. (g) Cross-sectional of leaf blade. 1 and 2 represents the leaf cross-sectional of transgenic plants, 3 and 4 are WT. Note: Three lines were used for the leaf morphology investigation, Fig. 5c & d present the typical leaf traits of line E1. Ve indicates veins. Scale bars = 200 µm.
-
TCP transcription factors are ancient proteins that have been widely identified in various plant species, such as Arabidopsis[15], rice[16] and maize[18]. Most studies have demonstrated that TCP transcription factors play critical roles in the process of plant growth and development. In this study, a total of 39 TCP genes were identified by the comprehensive analysis of the O. fragrans genome. These genes were classified into three main groups based on their phylogenetic relationships (Fig. 1). Furthermore, these genes were found to be distributed across 18 chromosomes in O. fragrans (Supplemental Fig. S1). Similar to other gymnosperms and angiosperms, such as in Populus, Vitis vinifera, and rice, we proposed that TCP genes in O. fragrans evolved into a larger family through gene duplication and diversification[7]. Among the class I OfTCP members, a highly conserved bHLH domain was identified, which is crucial for DNA binding in the basic region of the TCP N-terminal. Additionally, the C-terminal HLH region is involved in the formation of homodimer or heterodimer[34,35]. Notably, the class I OfTCP proteins exhibit distinctly differences in their motif compositions, while only motif 1 and motif 2 are present in the all class I TCP transcription factors. It is implied that the presence of different motifs in class I OfTCP proteins supports the differentiation of its functions. The phylogenetic tree analysis further revealed that class I OfTCP proteins of the same branch may be functionally similar, while those from different branches likely have complementary or different functions (Fig. 2).
Leaves serve as crucial ornamental organs for garden plants, exhibiting considerable variations in morphology across different plant species or cultivars. Furthermore, leaf morphology of leaves undergoes transformations as they age and in response to specific environmental conditions in which the plant species thrives[36,37]. In Arabidopsis, the TCP family of proteins has been extensively documented for its fundamental involvement in the regulation of leaf shape. For instance, AtTCP23 gene illustrates its influence on leaf borders, as evidenced by contrasting phenotypes observed in the overexpression and knockout lines[38]. However, there are limited reports available regarding the role of TCP transcription factors in regulating of leaf morphology specifically in O. fragrans. Therefore, in order to elucidate the potential involvement of TCP transcription factors in shaping leaves in O. fragrans, we conducted a comprehensive analysis of the expression of class I TCP family members across various tissues (Fig. 3). The outcomes revealed that all 16 OfTCP genes were expressed in the all-tested tissues, with notable higher expression levels of OfTCP5, OfTCP13, OfTCP22 and OfTCP37 observed in leaf tissue relative to other tissues. The results suggests that these four TCP genes may act as important regulators of leaf morphology in O. fragrans. As a next step, we isolated the sequence of the intriguing OfTCP13 gene from the O. fragrans cultivar 'Yanhonggui' for subsequent investigations. By means of subcellular localization analysis, we determined that OfTCP13 exclusively localized in the nucleus (Fig. 4a), consistent with previous observations in other plant species[39]. And the transcriptional activation assay revealed that OfTCP13 does not possess the capability to activate transcription on its own (Fig. 4b), indicating that it may require collaboration of other transcription factors to exert its regulatory function. TCP transcription factors have been implicated in promoting leaf growth and development by regulating processes such as cell proliferation and enlargement[40]. For instance, Arabidopsis mutants of TCP2, TCP3, TCP4, TCP10, and TCP24 result in a phenotype where the leaves exhibit an increase in both size and curvature[41]. Additionally, the transcription factors AtTCP2 and AtTCP3 have been found to synergistically activate the expression of NGATHA (NGA), which controls leaf margin formation[42]. In the case of chrysanthemum, CmTCP20 not only have effect on the regulation of petal cell growth but also induces elongation of Arabidopsis rosette leaves[43]. In our study of O. fragrans, we constructed overexpression lines of OfTCP13 in tobacco and observed that overexpressed OfTCP13 resulted in leaves with a narrower shape and wavy serrated margins compared to WT plants (Fig. 5c, d). Notably, the overexpressed lines exhibited a significant increase in the leaf thickness and blade length-to-width ratio in comparison to the WT plants (Fig. 5e, f). Moreover, the examination of paraffin sections showed a higher number of mesophyll cells in the transgenic tobacco plants (Fig. 5g). These results suggested that OfTCP13 potentially regulates leaf thickness and blade length-to-width ratio by promoting the mesophyll cell number. It is noteworthy that plant hormone auxin plays a crucial role in determining the final morphology of leaves, as it is involved in both leaf formation and the development of marginal outgrowths that contribute to the overall shape of the leaf[44]. Hence, it is plausible to speculate that there might be a correlation between OfTCP13 and auxin in promoting the proliferation of mesophyll cells in O. fragrans[45−48].
-
In summary, a total of 39 OfTCP genes were identified from the O. fragrans genome according to the conserved bHLH domain and phylogenetic relationship. Among these, the class I TCP gene OfTCP13 was found to be crucial in the regulation of leaf morphology by promoting the proliferation of mesophyll cells in O. fragrans. These findings contribute to our understanding of how TCP transcription factors govern the diversity of leaf morphology in O. fragrans and provide a solid theoretical foundation for further research in this field.
This work was funded by the National Training Program of Innovation and Entrepreneurship for Undergraduates (202110341025), College Student's Science and Technology Innovation Activity Plan of Zhejiang Province (2022R412A013), Zhejiang Agriculture and Forestry University School-level Research Training Program for Undergraduates (2021KX0055), Key Research and Development Program of Zhejiang Province (2021C02071).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Zifei Zheng, Qian Xu, Jingqian Tang
- Supplemental Fig. S1 Chromosomal distribution of 39 OfTCP genes on O. fragrans chromosomes.
- Supplemental Fig. S2 Sequence LOGOs for the motif of class I OfTCP proteins.
- Supplemental Fig. S3 Cis-regulatory element analysis of class I OfTCP promoter sequences.
- Supplemental Table S1 TCP amino acid sequences from O. fragrans and A. thaliana.
- Supplemental Table S2 Information of primers.
- Supplemental Table S3 Characterization of 39 OfTCP proteins in O. fragrans.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zheng Z, Xu Q, Tang J, Chen P, Hu Z, et al. 2023. Genome-wide analysis of TCP gene family in Osmanthus fragrans reveals a class I gene OfTCP13 modulate leaf morphology. Ornamental Plant Research 3:15 doi: 10.48130/OPR-2023-0015 

Genome-wide analysis of TCP gene family in Osmanthus fragrans reveals a class I gene OfTCP13 modulate leaf morphology
- Received: 23 April 2023
- Accepted: 25 July 2023
- Published online: 31 August 2023
Abstract: Osmanthus fragrans is a woody perennial that is cultivated in most Asia areas and widely employed for gardening and landscaping purposes. In addition to its distinctive fragrance, there has been a growing interest in the variation of O. fragrans leaf shapes. However, there are limited reports regarding the role of TCP genes regulating leaf morphology in O. fragrans. In this study, a total of 39 TCP members were identified at the genome level, and the sequence characteristics and tissue expression of class I TCP genes were analyzed. The highest expression gene OfTCP13 cloned from O. fragrans cultivar 'Yanhonggui' and transferred into tobacco plants. Moreover, the OfTCP13 protein was found to be localized in the nucleus and lacked transcriptional activation activity. Compared with wild-type (WT) plants, overexpression of OfTCP13 significantly influenced the leaf morphology of tobacco plants, resulting in significantly greater leaf thickness and blade length-to-width ratio than WT plants. Furthermore, the cross-section of the transgenic tobacco plants exhibited mesophyll cell number increased relative to WT plants, suggesting that OfTCP13 may regulate leaf morphology through promoting mesophyll cell number. Together, our results can provide a valuable insight into the improvement of diversity of leaf morphology in O. fragrans.
-
Key words:
- Osmanthus fragrans /
- TCP transcript factor /
- Leaf morphology /
- Mesophyll cell.
{kind=link}
{kind=link}