










-
Tea plants are economically important perennial evergreen plants originating from the tropics and subtropics. Tea products have gained worldwide popularity as a beverage choice, primarily attributed to their rich flavor and significant health benefits[1−3]. Tea is cultivated worldwide, making it subject to a diverse array of different environmental stresses. Low temperature is one of the most crucial environmental factors that limit the growth and geographical distribution of tea plants[4]. Low temperature stress occurs frequently in tea-growing regions, significantly affecting both the yield and quality of tea. Research aimed at a better understanding of the molecular mechanisms underlying the response of tea to cold stress could therefore lead to the development of more tolerant tea cultivars.
Transcription factors (TFs) regulate growth, development and adaptation to stresses in plants through various signal transduction pathways[5]. The RAV transcription factor family is a plant-specific subfamily of the APETALA 2 (AP2)/ethylene-responsive factor (ERF) family, and its members typically include an AP2 domain and a B3 domain. The AP2/ERF TF family is one of the largest TF families in plants and its members play essential roles in the regulation of biotic and abiotic stress-responsive gene expression[6,7]. The AP2/ERF domains can be divided into five classes based on sequence similarity: AP2, dehydration-responsive-element-binding (DREB), ERF, related to ABA-INSENSITIVE 3/VIVIPAROUS1 (RAV), and Soloist.
Recent studies have demonstrated that the RAV subfamily is involved in many physiological and developmental pathways[8,9], including abiotic stress response[7]. Thus far, the function of RAV TFs in plants has primarily been investigated in model species such as Arabidopsis[8,10], rice[11] and others. RAV genes are found to be present in varying numbers across different plant species, with 26 found in wheat[12], 13 in soybean[13], 11 in pear[14], 15 in rice[15] and 14 in rapeseed[16]. In Arabidopsis and soybean, RAV1 has been shown to regulate seed germination, the regeneration of roots and adventitious buds, and responses to low temperature stress[17,18]. In tobacco[19], melon[20], cotton[21] and pepper[22], RAV1 enhances plant tolerance to cold, drought and high salt stress, and in Brassica campestris[23] and Galegae orientails[24], RAV expression is induced by cold, drought and high-salinity stress. PbRAV6 is also found to participate in the anthocyanin biosynthesis in Pyrus bretschneideri Rehd, which determines pericarp color[14]. Chen et al. showed that the GoRAV gene in Galegae orientalis can be induced by cold, dehydration and high-salinity and ABA treatments[24]. These findings suggest that RAV genes play conserved roles in regulating resistance to abiotic stresses across diverse plant species.
In addition, the expression levels of RAV genes are induced by the plant hormone methyl jasmonate (MeJA) in response to stress in rice. MeJA regulates various processes associated with plant growth and development and stimulates the expression of defense-associated genes, thereby mediating various plant responses to biotic and abiotic stress. Cold stress studies in Arabidopsis, peach[25,26], wheat[27], pea[28], rubber tree[29] and cherry[26] have all demonstrated the involvement of MeJA in cold tolerance. Additionally, spraying exogenous MeJA can effectively improve the cold tolerance of tea plants, and can also impact the aroma of tea leaves[30−32].
In this study, we identified all members of the RAV family in the tea plant genome. To better understand the relationship between RAV genes and the JA signaling pathway, we assessed the expression patterns of RAV genes in response to low temperature stress. This work provides a theoretical basis for creating new cold-tolerant tea plant germplasm through genetic engineering.
-
The AtRAV gene sequences of Arabidopsis were downloaded from TAIR (
www.arabidopsis.org ) and the Hidden Markov Model (HMM) profile of the conserved domains B3 (PF02362) and AP2 (PF00847) of CsRAV proteins were used to examine proteins downloaded of 'Shuchazao'[33] from the Tea Plant Information Archive (TPIA,http://tpdb.shengxin.ren , accessed on 1 July 2022). The conserved domains of candidate RAV proteins were identified using the online websites Pfam (https://pfam.xfam.org/search ) and NCBI (www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ), E value <10-5.Alignment and phylogenetic analysis of CsRAV sequences
-
A total of 13 Arabidopsis thaliana AtRAV proteins sequence and 16 rice OsRAV protein sequences were downloaded from the Arabidopsis Information Resource (TAIR) (
www.arabidopsis.org ) and Rice Genome Annotation Project (http://rice.uga.edu ). CsRAV protein sequences contained within the tea genome were obtained from the Tea Plant Genome Database. All obtained RAV proteins were aligned using DNAMAN 9. A phylogenetic tree was then constructed using maximum likelihood estimate (ML) method with MEGA 7.0 software.Gene structure, biophysical properties and protein motifs
-
The structures of the CsRAV genes were predicted using the Expasy online analysis website (
https://web.expasy.org/protparam ). We then predicted the CsRAV protein amino acid numbers, molecular weight, theoretical isoelectric point (PI), and grand average of hydropathicity. The Softberry Web Site (http://www.softberry.com/berry.phtml?topic=protcomppl&group=programs&subgroup=proloc ) was used to predict the subcellular localization of RAV proteins. Finally, the MEME suite (https://meme-suite.org/meme/tools/meme ) was used to determine the conserved motifs, with the maximum number of motifs set to 20.Gene sequence analysis of CsRAVs
-
The exon-intron structures of the CsRAV genes were identified and visualized using the TBtools. The cis-acting regulatory elements of the 2000-bp upstream promoter sequences of the CsRAV genes were analyzed using PlantCARE program (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ).Genomic location and three-dimensional structure analysis of the CsRAV genes
-
CsRAV genes were mapped on chromosomes based on the whole tea plant genome annotation by Tea Plant Information Archive (TPIA, tpdb.shengxin.ren)[34], and a map was generated using the TBtools software. Three dimensional structures of CsRAV genes were analyzed using Phyre2 (PHYRE2 Protein Fold Recognition Server (ic.ac.uk))[35], as described previously.
Plant material and treatments
-
One-year-old cuttings of the 'Longjing 43' were used as experimental materials. The tea plants were grown for two weeks at 25 °C under a 16 h/8 h day/night cycle and 75% relative humidity. The plants were then divided into seven different treatment groups (Table 1)[36−39].
Table 1. Details of each treatment.
Treatment Days 1–4 Days 5–8 Day 9 CK Room temperature and water spray Room temperature and water spray Samples collected T1 Room temperature and water spray 4 °C cold stress Samples collected T2 4 °C cold stress Room temperature and water spray Samples collected T3 Room temperature and water spray Room temperature and 100 μM MeJA spray Samples collected T4 Room temperature and 100 μM MeJA spray Room temperature and water spray Samples collected T5 Room temperature and water spray 4 °C cold stress and 100 μM MeJA spray Samples collected T6 4 °C cold stress and 100 μM MeJA spray Room temperature and water spray Samples collected After nine days of treatment, three biological replicates of one bud and two leaves were collected under six treatments and one control condition. All samples were frozen in liquid nitrogen immediately and stored at −80 °C for subsequent analysis.
Expression pattern analysis
-
Total RNA was extracted from the frozen leaf samples via the SteadyPure Universal RNA Extraction Kit (AG21019, Accurate Biotechnology, Changsha, China) according to the manufacturer's protocol. cDNA was obtained by HiScript III RT SuperMix for qPCR (+gDNA wiper) (Vazyme, Nanjing, China). The CsRAV primer pairs used for qRT-PCR were designed with Primer Premier 7.0 software (Supplemental Table S1). The qRT-PCR analyses of the seven CsRAV genes were carried out using the ChamQ SYBR qPCR Master Mix (Vazyme, Nanjing, China) and performed with Bio-Rad CFX96 fluorescence. The Csβ-actin gene was used as the reference gene[40]. The qRT-PCR reaction program was set as follows: 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s. All experiments were performed with three biological and technical replicates. Relative gene expression levels were calculated using the 2−ΔΔCᴛ method[41].
Statistical analysis
-
All data in this study were statistically analyzed using Excel (version 2019). One-way ANOVA significance analysis was performed using IBM SPSS statistics (version 26.0). Duncan's multiple range tests were used for multiple comparisons of variance analysis, with p < 0.05 as the cutoff for statistical significance. GraphPad Prism and TBtools v1.098 (version 8.0) were used for data visualization.
-
By conducting blast analysis with 13 identified Arabidopsis AtRAV family members, and using the Hidden Markov Model profile of the RAV proteins conserved domain B3 (PF02362) and AP2 (PF00847) as a query to identify the RAV family members we then used Pfam and NCBI to confirm the genes. In total, 12 putative CsRAV genes were identified in the 'Shuchazao' genome database. Five genes were excluded upon subsequent analysis. TEA021527.1, TEA006170.1 and TEA013125.1 were found to contain only the B3 (PF02362) conserved domain, while TEA011479.1 and TEA028514.1 were identical, and the TEA024560.1 has an AP2 structural domain E-value of 0.00061 which is usually considered to be a more plausible result with an E-value less than 10-5. Therefore, CSS00009470.1 does not meet the screening range criteria. The physiological and biochemical parameters of the seven remaining genes (CsRAV1-7) are summarized in Table 2. The lengths of the seven CsRAV proteins ranged from 329 aa (TEA020682.1) to 1,130 aa (TEA030533.1), while their molecular weights varied from 36.77 kDa (TEA020682.1) to 127.04 kDa (TEA030533.1). The theoretical isoelectric points (pIs) of the seven CsRAV proteins ranged from 5.2 to 9.3. The GRAVY values of all CsRAV proteins were negative, suggesting that they were hydrophilic. All seven proteins were predicted to localize to the nucleus. The genes were found to be unevenly distributed across the six tea chromosomes (Fig. 1). CsRAV1 and CsRAV4 were found to be on the end of chromosome 2, while CsRAV7 was at the beginning of chromosome 3. CsRAV6 was located at the end of chromosome 4, while CsRAV5 was at the start and end of chromosome 5. CsRAV3 was found in the middle of chromosome 9, while CsRAV2 was found at the end of chromosome 12.
Table 2. The physicochemical properties of tea plant RAV proteins.
Number Gene name Gene ID Amino acid
residuesMolecular
weight (kDa)Theoretical pI Grand average of
hydropathicitySubcellular
localization1 CsRAV1 TEA028514.1 366 40.67 8.96 −0.610 Nuclear 2 CsRAV2 TEA006376.1 375 41.78 9.30 −0.561 Nuclear 3 CsRAV3 TEA020682.1 329 36.77 8.97 −0.462 Nuclear 4 CsRAV4 TEA006600.1 344 39.68 6.20 −0.516 Nuclear 5 CsRAV5 TEA016614.1 352 39.56 8.19 −0.655 Nuclear 6 CsRAV6 TEA004203.1 359 41.07 5.61 −0.698 Nuclear 7 CsRAV7 TEA030533.1 1,130 127.04 5.20 −0.819 Nuclear 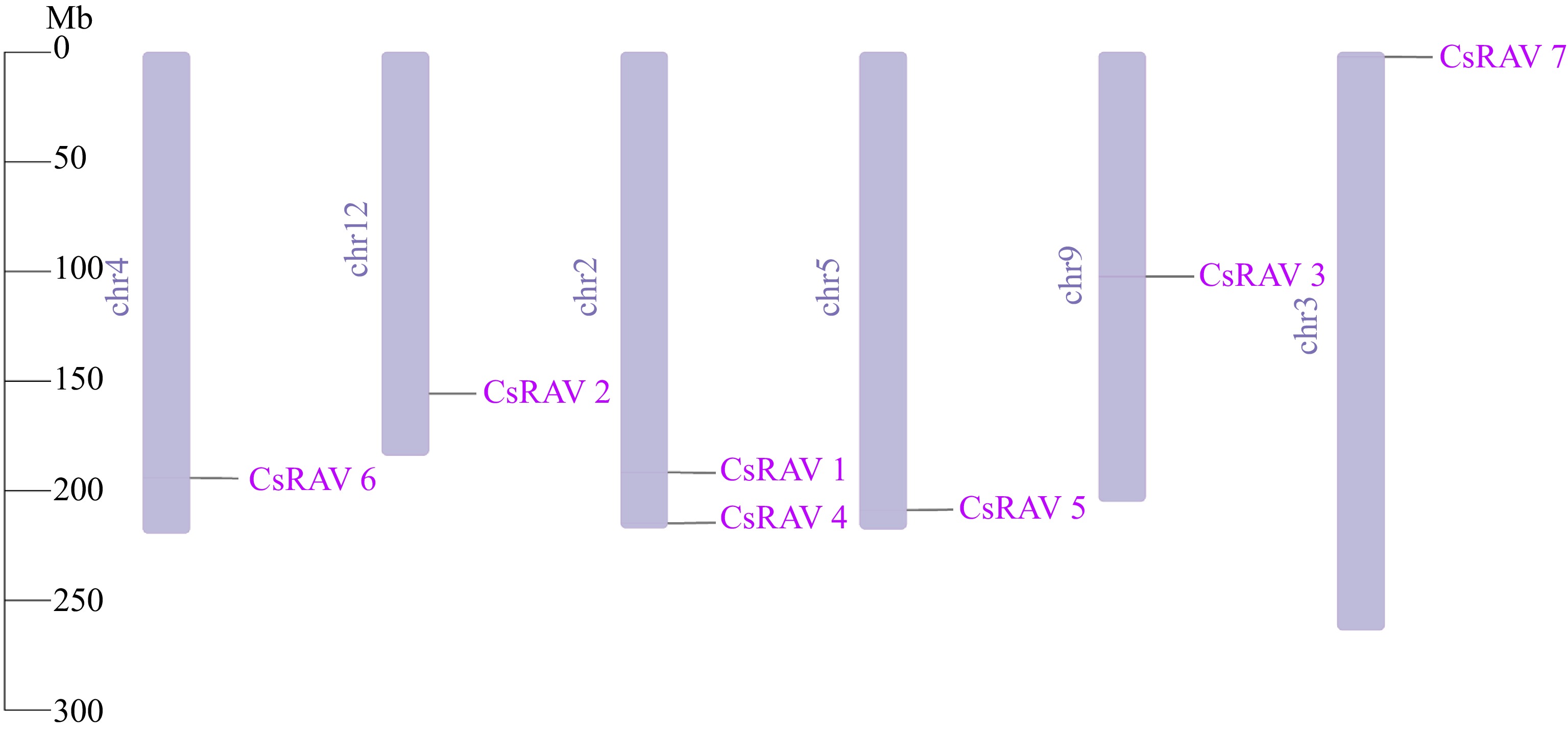
Figure 1.
The chromosomal locations of CsRAV genes.
Phylogenetic analysis and sequence alignment of CsRAV genes
-
To assess the evolutionary relationships among the RAV homologs in plant genomes, the sequence of the seven tea plant RAVs, 16 rice RAVs, and 13 Arabidopsis RAVs were used to create a phylogenetic tree (Fig. 2). CsRAV1-7 were all found to contain AP2 and B3 domains, and were further divided into three groups (Fig. 3a). CsRAV2, CsRAV4 and CsRAV5 belonged to Group I, while CsRAV1 and CsRAV3 belonged to Group II. CsRAV6 and CsRAV7 belonged to Group III. To further characterize the protein sequence features of the CsRAV family members, conserved motifs were predicted using MEME. A total of 20 conserved motifs were found and designated motifs 1 to 20 (Fig. 3b). Detailed sequence information and conserved motifs are shown in Supplemental Table S2. All seven CsRAV genes contained motif 1, motif 2, motif 3 and motif 4. Different groups of CsRAV members exhibit similar gene structures and motif composition. Group II contained more motifs than Group I and III, with CsRAV7 having the most motifs (13). CsRAV2, CsRAV3 and CsRAV5 contained similar motifs, with motif 16 found in both CsRAV2 and CsRAV5 proteins. CsRAV6 and CsRAV7 in Group III also had similar motifs, with CsRAV7 having more motifs of motif 11, 12 and 20 than CsRAV6. CsRAV genes were found to contain between 0 and 15 introns, but there were no obvious differences between the numbers of introns in different CsRAV groups. CsRAV1, CsRAV4 and CsRAV6 have no introns, while CsRAV2 and CsRAV3 have only one intron, and CsRAV5 has two introns. The CsRAV7 gene contained 15 introns. (Fig. 3c).
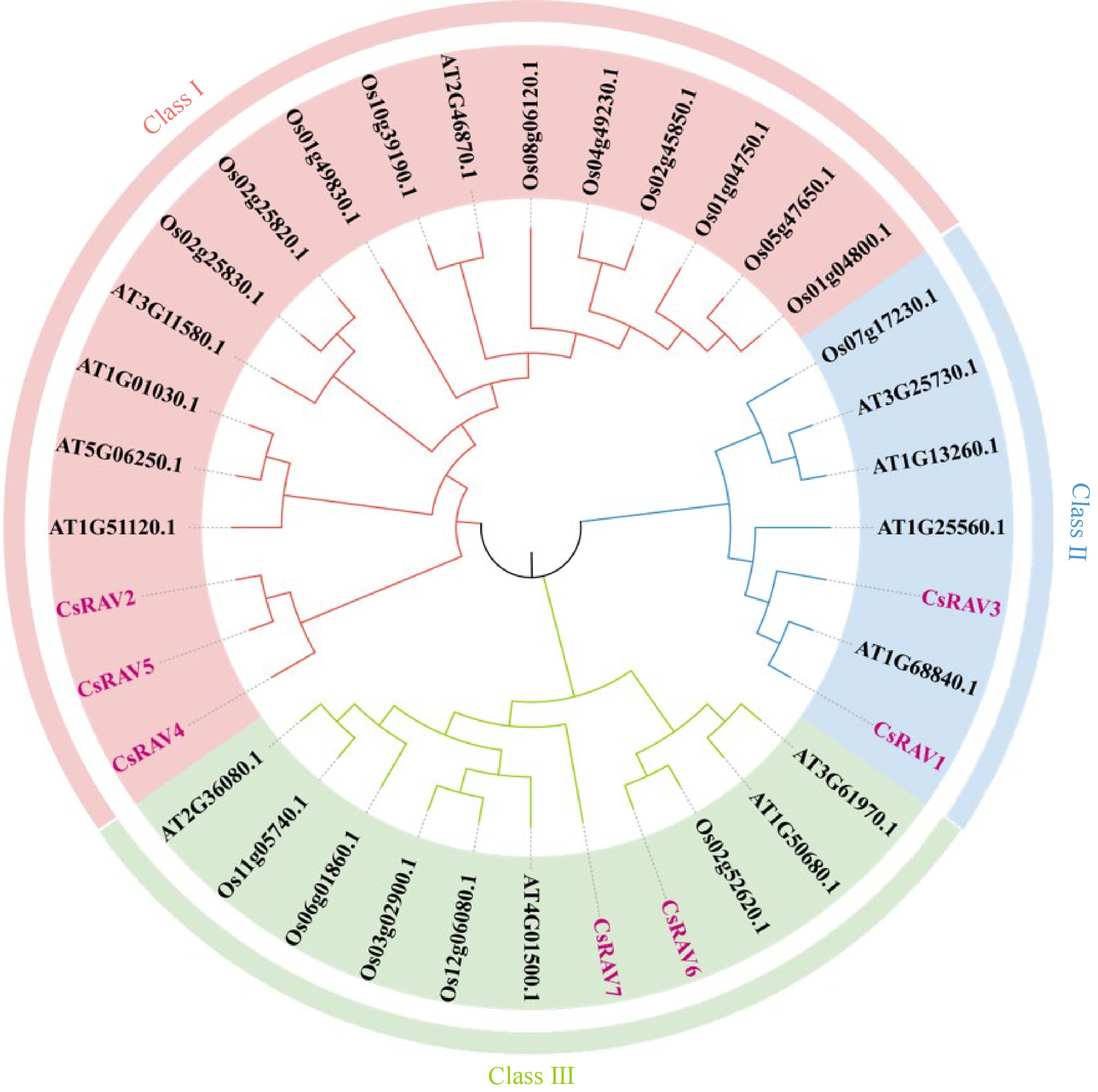
Figure 2.
Phylogenetic analysis of RAV genes from Camellia sinensis, Arabidopsis thaliana, and Oryza sativa.
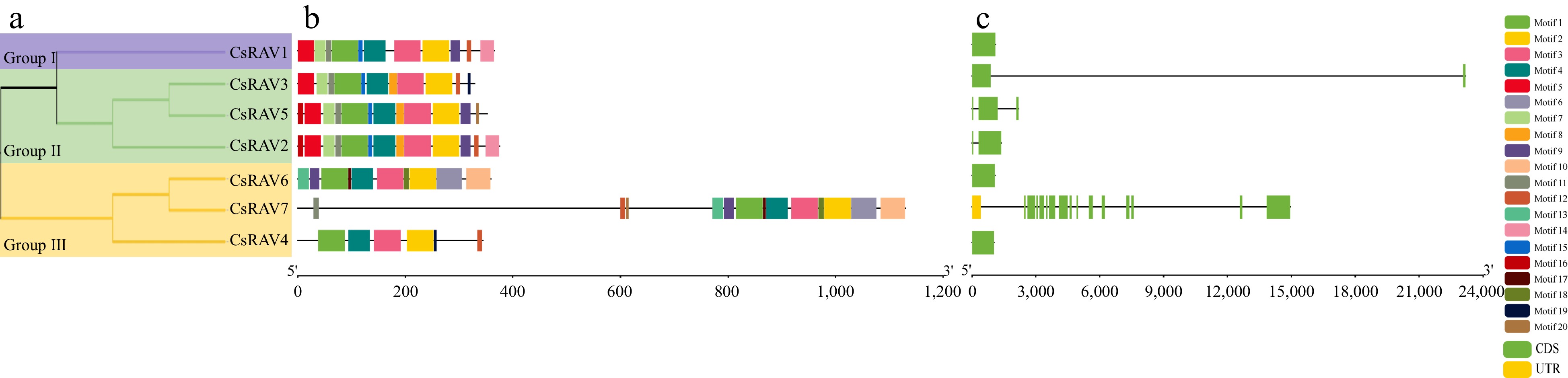
Figure 3.
CsRAV protein and gene sequence analysis. (a) Phylogenetic tree analysis, the purple line represents group I, the green line represents group II and the yellow line represents group III. (b) Motif analysis. (c) Gene structure analysis. Horizontal lines indicate introns.
To elucidate sequence characteristics of the CsRAV protein, we performed multiple sequence alignments on CsRAV1 from CsRAV7 using DNAMAN software. Results showed that the CsRAV TFs of Camellia sinensis has a core AP2 binding domain at the N-terminal and a conserved B3 domain at the C-terminal (Fig. 4).

Figure 4.
Multiple alignments of CsRAV proteins in tea plants.
Predicted structural features of conserved domains in tea RAV proteins
-
To explore the potential structural features of the conserved domains of RAV proteins in tea plants, the secondary structure prediction of CsRAV1-CsRAV7 proteins was performed using SOPMA online software. As shown in Fig. 5, the random coil structure of the seven CsRAVs proteins accounts for 42.34%−57.52% of the total protein length, the α-helix structure accounts for 26.11%−34.82%, the extended strand structure accounts for 13.01%−20.53%, and the β-turn structure accounts for 3.36%−7.95%. The prediction results of CsRAV7 protein showed that the random coil structure accounted for 57.52% of its total length, which was the highest of all seven CsRAV proteins, and the α-helix (26.11%), extended strand (13.01%) and β-turn (3.36%) accounted for the least of all CsRAV proteins.
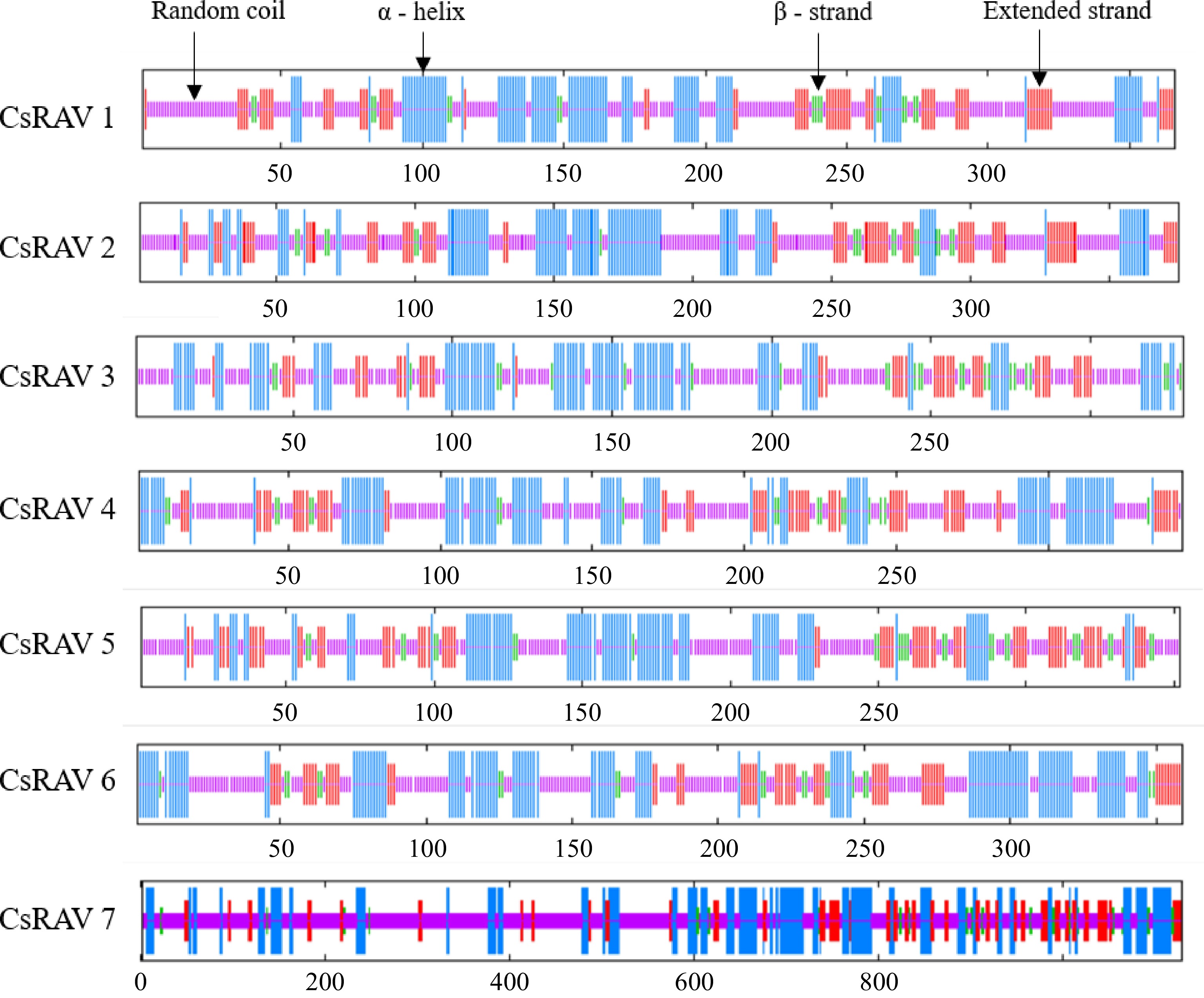
Figure 5.
Secondary structure prediction of the RAV family in tea.
The amino acid sequences of tea plant RAV family proteins were analyzed by NCBI Conserved Domain Search, which revealed that CsRAV1-CsRAV7 all contain AP2 and B3 superfamily conserved domains. Except for the CsRAV7 protein, the positions of the AP2 conserved domain ranged from 112 to 429 bp, while the B3 conserved domain ranged from 475 to 954 bp. Based on the principle of homology modeling, the three-dimensional structures of two conserved domains (AP2, B3) of tea plants RAV protein were predicted by SWISS-MODEL software (Fig. 6). The B3 domain was found to contain two α-helices, two antiparallel β-barrels and extended strands (CsRAV1/2/3/4/7) (Fig. 6a). In the AP2 domain, five (CsRAV1/2/3/4/7) of the seven CsRAV proteins had three β-barrels that were parallel with one α-helix (Fig. 6b).
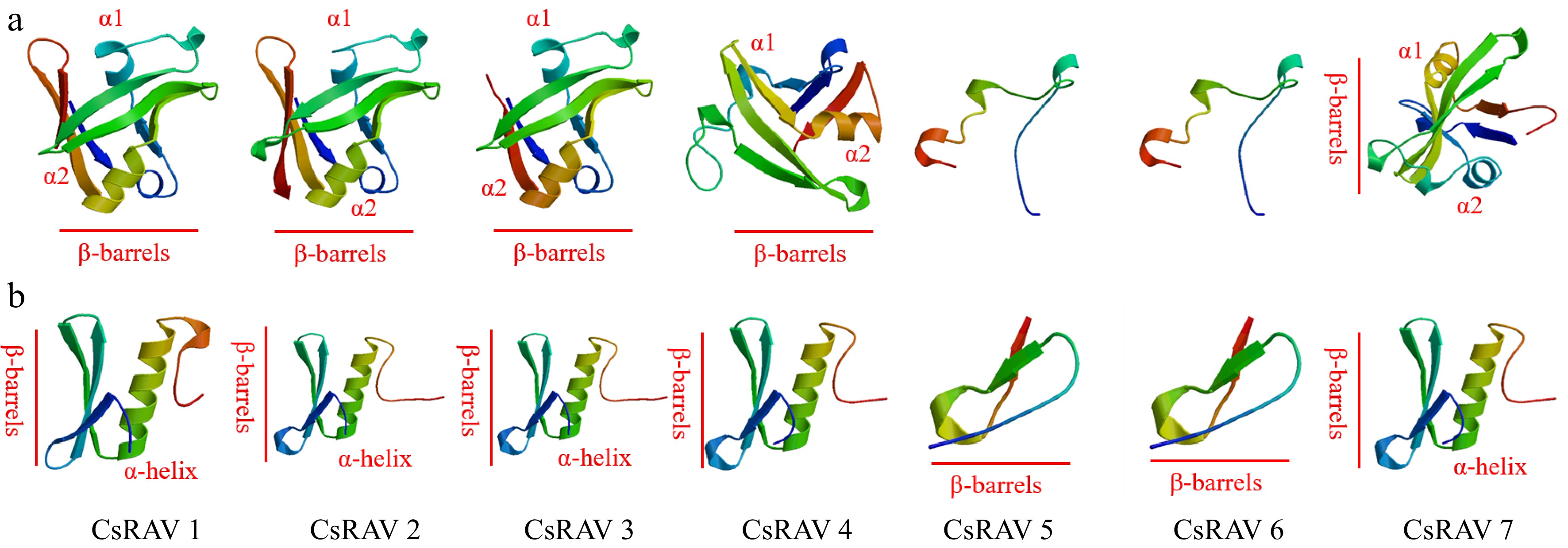
Figure 6.
Predicted structures of conserved domains in RAV proteins of tea plants. (a) Ribbon diagrams of the B3 domains of the tea plant CsRAV proteins. (b) Ribbon diagrams of the AP2 domains of the tea plant CsRAV proteins.
cis-acting elements analysis of CsRAV genes
-
To study the cis-elements in the promoters of the seven CsRAV genes, 2,000 bp of sequence upstream from the start codon of each gene were analyzed. All genes except for CsRAV5 contained 40 kinds of cis-element, which were classified into four categories related to phytohormone responses, stress responses, light response and plant growth and development (Fig. 7). Of these, seven hormone-responsive elements were found that accounted for 30.65% (42/137) of the total cis-elements, including abscisic acid responsiveness (ABRE), auxin-responsive element (TGA-element/TGA-box), gibberellin-responsive element (P-box/GARE-motif), methyl jasmonate-responsiveness (CGTCA/TGACG-motif) and salicylic acid responsiveness (TCA-element/SARE). The five most abundant cis-elements are shown in Fig. 7. The stress response category contained six cis-elements, accounting for 10.95% (15/137) of the total elements found, and included anaerobic induction (ARE/GC-motif), defense and stress responsiveness (TC-rich), low-temperature responsiveness (LTR),drought-inducibility (MBS) and wound-responsiveness (WUN-motif). All but the least abundant cis-element are shown in Fig. 7a. There were 14 light-responsive elements found, accounting for 50.36% (69/137) of the total number of cis-elements. This category included Box-4, GT1-motif, MRE and G-box, and others. Five of the cis-elements were in the plant growth and development category, accounting for 8.03% (11/137) of all cis-elements. This group included endosperm expression (GCN4_motif), meristem expression (CAT-box) and seed-specific regulation (RY-element). All but the six least abundant cis-elements are shown in Fig. 7a. The types of cis-acting elements contained in the promoter regions of different CsRAV genes varied across different promoters. The promoters of CsRAV4 contained a large number of elements. CsRAV4 was found to contain two stress-response elements and four phytohormone-response elements (MeJA response). The promoters of all CsRAV genes except for CsRAV5 contained cis-acting elements belonging to at least four different functional categories, ranging from 12 (CsRAV7) to 32 (CsRAV4). Overall, CsRAV gene promoters contained significantly more hormone-responsive and stress-responsive elements compared to elements associated with light response or plant growth and development. Some elements were only present in one or two CsRAV gene promoters, such as the TC-rich repeats (defense and stress responsiveness) that were only present in CsRAV2 and CsRAV4, LTR (low-temperature response) only found in CsRAV3, and MBS (MYB binding site involved in drought-inducibility) only present in CsRAV4 and CsRAV7. Overall, the cis-acting elements present in the promoters of CsRAV4 indicate that they are likely more responsive to plant hormones and stresses.
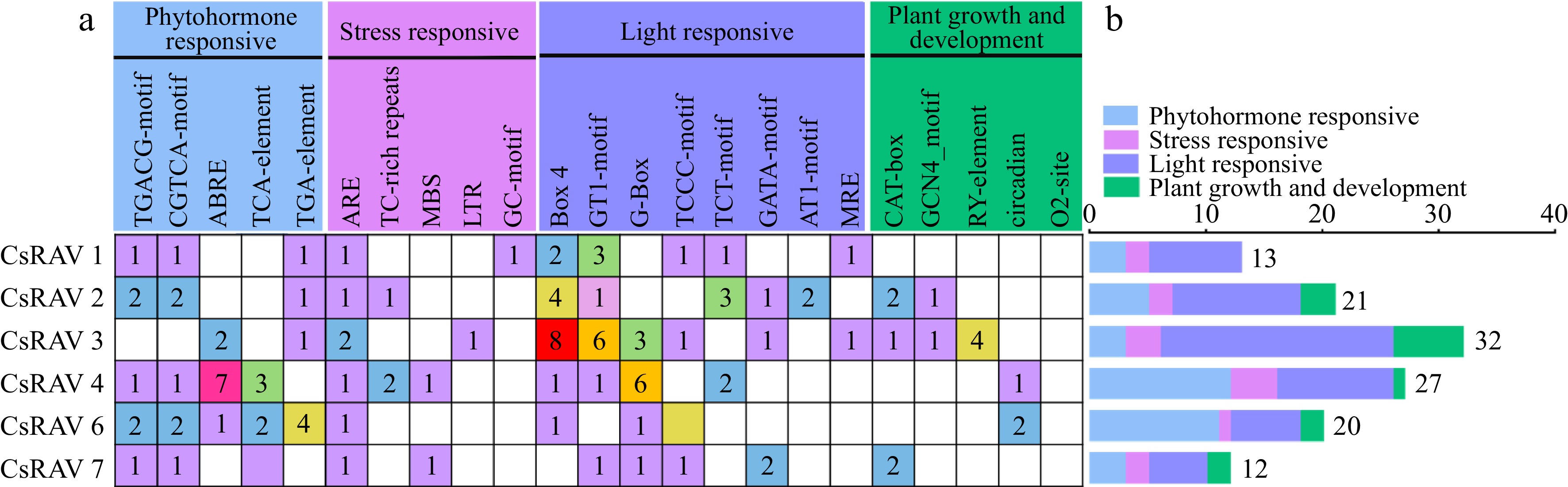
Figure 7.
Cis-promoter elements of the CsRAV genes in tea plants. (a) The numbers of different cis-elements in the promoters of CsRAV genes are indicated in different colors in the grid. (b) The numbers of cis-elements in four functional categories per gene are represented by different colored histograms.
Expression patterns of CsRAV genes
-
We next analyzed the expression patterns of all CsRAV genes in six experimental conditions (Fig. 8). This analysis revealed that the CsRAV1 gene was highly induced by exogenous MeJA treatment alone (T4), and was significantly repressed under cold stress (T1) and combined cold-stress and exogenous MeJA treatments (T5 and T6). CsRAV2 expression was significantly induced under cold stress alone (T2) and MeJA treatment alone (T4). The expression trends of CsRAV3 and CsRAV5 genes were similar to the CsRAV2 gene. CsRAV4 gene expression was significantly higher in T2, T3, T4 and T6 compared to CK, and reached the highest level in T2. The relative expression of the CsRAV6 gene was significantly elevated only under T2, while the other four treatments reduced its expression level compared to CK. Compared with the other six CsRAV genes, CsRAV7 had the most distinctive expression changes. CsRAV7 was significantly upregulated in response to six different treatments, including cold stress alone, exogenous MeJA alone, and the combined treatments.
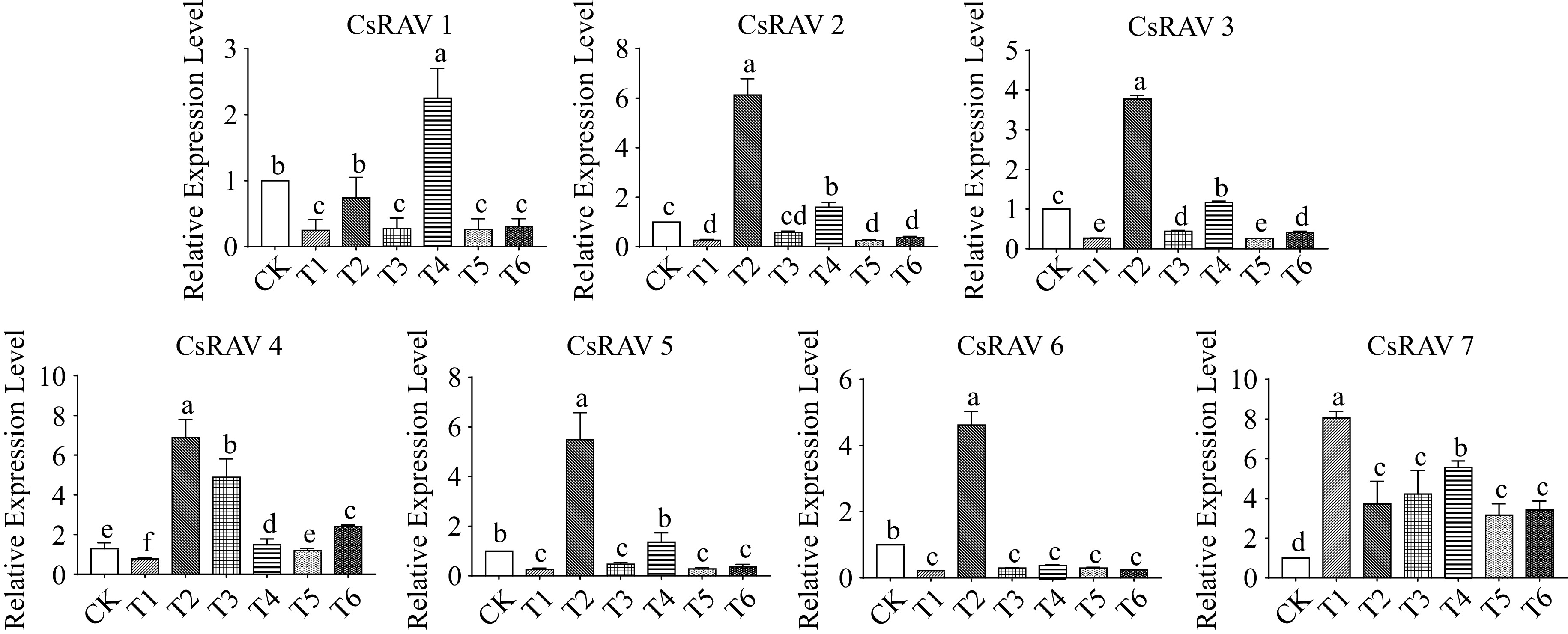
Figure 8.
The expression pattern of CsRAV genes in each sample. Error bars represent the standard error of the mean (n = 3). Different lowercase letters indicate significance at p < 0.05.
-
Plants are affected by various environmental stresses during their growth and development, which can lead to significant yield losses. Tea plants are thermophilic, and exposure to low-temperature stresses can cause significant quality and yield losses. Stress symptoms can be alleviated by the application of MeJA, which is a plant-specific signaling molecule that impacts biotic and abiotic stress responses. Previous studies have reported that RAV genes respond to both abiotic stress and hormonal signals[42−44]. RAV TFs have been found in many plant species, including soybean, rice, Arabidopsis and Brassica species. We found that the tea plant genome contains seven distinct CsRAV genes, with unique responses to low-temperature stress and MeJA treatment.
Previous studies have shown that Arabidopsis and rice possess 13 and 16 RAV genes, respectively. Although 12 putative RAV genes were initially identified in tea, only seven of these genes contained both B3 and AP2 domains. Phylogenetic tree analysis of the RAV proteins from tea, Arabidopsis and rice indicated that RAV proteins of the three species were relatively conserved during the evolutionary process. Clustering with only A. thaliana and O. sativa RAV proteins did not result in an accurate classification, but the addition of tea RAV proteins created three clear sequence homology-based groups (I, II and III). Members of these groups could therefore perform evolutionarily conserved functions. For example, the Arabidopsis genes AtRAV1 (AT1G13260) and AtRAV2 (AT1G68840)[45] that are involved in response to cold stress and leaf senescence clustered in Class II, indicating that the tea plant genes in this class (CsRAV1, CsRAV2, CsRAV3 and CsRAV5) may also be involved in these same processes. This was confirmed in our stress treatments, which demonstrated increased expression of CsRAV1, CsRAV2, CsRAV3 and CsRAV5 during cold stress. Conserved motif analysis revealed that the different subgroups of CsRAV proteins from tea plants have distinctive motifs, which correlate with functional differentiation between subgroups. This analysis also revealed that the CsRAV7 protein had the most unique motif profile, indicating that it may perform distinct biological roles compared to other CsRAV proteins.
Studies in mammals, nematodes, insects, fungi and plants have all shown that introns not only play a role in the regulation of gene expression but also participate in gene evolution[46]. Our phylogenetic analysis showed that the number and location of RAV introns were conserved within subclasses. All CsRAV genes contained less than three introns, except for the CsRAV7 gene in Group I, which contained 15. Alternative splicing enables gene families with more introns to be more flexible, leading to the acquisition of new functions during evolution. The low number of introns in most CsRAV may therefore underly their relatively high interspecies conservation.
Tea plant CsRAV proteins are hydrophilic and are predicted to localize to the nucleus. Random coils and α-helices are the main elements of the secondary structure of the seven CsRAVs proteins, all of which have the AP2 and B3 superfamily conservative domain structure. The AP2 conserved domain containing one α-helix and three antiparallel β-turns, and the B3 conserved domain containing one α-helix, two antiparallel β-turns and extended strands. It is hypothesized that they have act a synergistic role in regulating the biological properties of tea plants.
Promoter cis-acting elements interact with transcription factors to regulate gene expression at the transcriptional level and play an important role in the regulation of plant growth, development and response to stress[13]. The promoter regions of CsRAV genes contain many elements related to light response, phytohormone induction, plant growth and development, biotic and abiotic stress, indicating that CsRAV genes may play roles in all of these processes. We analyzed the cis-acting elements in promoter regions of the seven CsRAV genes, and found that they contained MeJA (TGACG/CGTCA), TC-rich repeats, ABRE, Box-4, G-Box, GT1, ARE, TGA, and LTR elements (Fig. 7a). However, the CsRAV5 gene had no predicted cis-acting elements within 2000 bp upstream of the gene. MeJA is an important signaling molecule in plants that plays a role in a variety of processes. Through the JA signaling pathway, it can regulate plant growth, development, and defense responses to various biotic and abiotic stresses. MeJA has been shown to play a positive role in alleviating cold stress in many plants. In addition, many plant bHLH proteins have been shown to contain G-Box elements[47,48]. Studies have shown that bHLH proteins are involved in the MeJA-induced responses, thereby promoting the synthesis of terpenoids and improving plant stress tolerance[49]. TCA-elements are salicylic acid (SA) response elements and studies have reported that SA can alleviate the damage of cold stress to plants by protecting the stability and integrity of plant cell plasma membrane, as well as reducing the accumulation of free radicals and lipid peroxidation in cell membranes. ABREs are a group of cis-acting abscisic acid (ABA) response elements, which are involved in regulating the expression of ABA-related genes, thereby improving plant resistance to drought, salt, cold and other stresses. It was previously reported that ABRE elements and LTR elements are critical for the cold-induced regulation of genes[50]. GT-1 cis-elements occur commonly in plant promoters and earlier work by Ning Li suggested that the interaction between OsASR2 and GT-1 plays an important role in the response of rice to pathogen infection and osmotic stress[51]. These findings, combined with our stress response testing, indicate that CsRAV genes are very likely involved in responses to phytohormones (MeJA, SA and ABA) and abiotic stresses in tea.
To further explore the potential functions of the CsRAV genes in tea plants in response to cold stress and MeJA treatment, the expression patterns of the CsRAV members in different treatments were examined. The seven CsRAV genes showed different expression patterns under individual and combined cold stress and exogenous MeJA treatment. Six CsRAV genes were significantly up-regulated in T2 and five genes CsRAV were up-regulated in T4. Additionally, six CsRAV genes were significantly down-regulated in T1. Except for the CsRAV7 gene, the other CsRAV genes were significantly down-regulated under the combined cold and MeJA treatment, while CsRAV7 had higher expression in all six different treatments. These results suggest that CsRAV7 may play a role in response to cold stress via the JA pathway in tea plants.
-
This work represents the first comprehensive and systematic analysis of the RAV gene family in tea plants. Seven CsRAV genes were identified in the genome of tea plant, and bioinformatics and expression analyses of these genes were employed to determine their potential roles in the growth, development and stress responses. The CsRAV genes were differentially expressed in response to cold stress and exogenous MeJA individual and combined treatments, implying that they are both hormone- and stress-responsive. Based on the structural analysis results of CsRAV proteins, CsRAV7 appeared to be unique within the family and requires additional research. These results lay a foundation for understanding the CsRAV gene family in tea, and further study of this family may enable the creation of more stress-tolerant tea plants.
-
The authors confirm contribution to the paper as follows: project design and management: Han Z, Ma Y, Fang W; data analysis: Han Z, Zhang H, Li P, Duan Y; manuscript preparaion and revision: Han Z, Ma Y; partial funds and consultant: Jiang J, Zhu X, Fang W. All authors read and approved the final manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This research was supported by the National Natural Science Foundation of China (31972460, 32160729), the China Agriculture Research System of MOF and MARA (CARS-19), Science and Technology Support Project of Guizhou Province (ZK[2021]154), and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primer information.
- Supplemental Table S2 The motif of CsRAV proteins.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Han Z, Zhang H, Li P, Duan Y, Jiang J, et al. 2023. Genome-wide characterization and identification of candidate CsRAV genes involved in JA pathway responses to cold stress in tea plant (Camellia sinensis). Beverage Plant Research 3:27 doi: 10.48130/BPR-2023-0027 

Genome-wide characterization and identification of candidate CsRAV genes involved in JA pathway responses to cold stress in tea plant (Camellia sinensis)
- Received: 25 July 2023
- Revised: 15 September 2023
- Accepted: 27 September 2023
- Published online: 03 November 2023
Abstract: The APETALA2/ethylene response factor (AP2/ERF) transcription factor (TF) superfamily plays an important regulatory role in the signal transduction of plant responses to various biotic and abiotic stresses, including low temperature. RAV is a subfamily of the AP2/ERF TF family, and its members contain B3 DNA binding and AP2 DNA binding domains. RAV gene family members have been studied in many species, but little is known about their function in tea plants (Camellia sinensis). In this study, seven RAV genes were identified from the tea plant genome database and named CsRAV1-7. Based on phylogenetic analysis, the CsRAV genes were classified into three groups, with genes in the same subgroup exhibiting similar structures and conserved motifs. Out of the seven genes, CsRAV7 was the most distinct and contained a large number of motifs and the longest coding region. Moreover, analysis of cis-acting elements indicated that CsRAVs are likely responsive to both abiotic stress and phytohormones. In addition, quantitative RT-PCR analysis revealed that most CsRAV genes were differentially expressed under cold stress and exogenous MeJA application as both individual and combined treatments. Only CsRAV7 was induced in all six different treatments, suggesting that it may play a key role in JA-mediated cold stress responses in tea plants.
-
Key words:
- Camellia sinensis /
- CsRAV gene /
- Cold stress /
- MeJA