









-
Lilies are well-known perennial bulbous plants that belong to the monocotyledonous family Liliaceae. The main commercial lilies can generally be grouped into several hybrid categories: Asiatic hybrids, Oriental hybrids, Martagon hybrids, Candidum hybrids, American hybrids, Trumpet lilies, Longiflorum hybrids, LA hybrids, OT hybrids, etc.[1,2]. As seed propagation is time-consuming[3], the method of asexual reproduction, particularly scale-cutting, is regarded as the most efficient and commonly used breeding approach in lily production. However, as the global demand for lilies continues to increase, improving the efficiency and quality of lily scaling propagation remains a significant challenge for the lily industry.
Bulblets, which are potentially adventitious buds, develop from the wound site at the base of mother scales during scale cutting. Bulblet formation involves two stages, namely, initiation and then the subsequent development. This process is similar to the formation of axillary buds (AXBs) in leaf axils[4−8]. Bulblet formation is regulated by numerous factors, including carbohydrate metabolism, endogenous hormones, and environmental factors[8]. Lily scales consist of reserve polysaccharides, including glucomannan and starch. Starch is the predominant type, accounting for approximately 85% of scale dry matter[9,10]. In Lycoris radiata, soluble sugars produced from the degradation of starch in the outer scales are transported to the bulblets, promoting their formation in the inner scales[8]. Starch gradually accumulates during bulblet formation, serving as a nutrient reserve during later growth stages[5,11]. Sucrose metabolism is closely related to bulblet formation. Sucrose can be transported to sink tissues through the phloem. Once unloaded, it can be converted by sucrose synthases (SUS) to uridine diphosphate glucose (UDPG) and fructose, or hydrolyzed by sucrose invertase (INV) to glucose and fructose[12]. Highly expressed genes involved in sucrose degradation, such as those encoding SUS and INV enzymes, contribute to early organogenesis[8]. In Lycoris sprengeri, the regeneration system yielding a higher number of bulblets also exhibited a significantly higher expression of LsCWIN2 (cell wall invertase, a type of sucrose invertase) compared to the system with a lower number of bulblets[13,14]. Glucose can be catalyzed by trehalose-6-phosphate synthase (TPS) to produce trehalose-6-phosphate (T6P)[15], and its content is highly associated with the number of lateral shoots[16]. Moreover, the increased content of soluble sugars in the inner scales of Lycoris radiata promotes the expression of D-type cyclin (CycD) genes and accelerates cell division during bulb development[8].
Exogenous hormones are commonly used to treat scale cuttings in order to induce bulblet formation[17−19]. 6-benzylaminopurine (6-BA), a commonly used experimental synthetic cytokinin, induces bulblet formation in Lilium lancifolium by promoting cell proliferation and activating the expression of cytokinin receptors LlAHK2/3/4[18]. In addition, cytokinins (CKs) can activate WUSCHEL-like homeobox (WOX) genes and further promote the formation of 'aerial bulblets' (bulbil) at the leaf axils of Lilium lancifolium[20,21]. Elevated levels of auxins, gibberellins (GAs), and jasmonic acid, as well as reduced levels of abscisic acid (ABA), are believed to facilitate bulbil formation in scale cuttings[22]. The initial accumulation of indole-3-acetic acid (IAA) was also observed during bulbil formation in Lilium sulphureum[23,24]. However, IAA may have a dual effect on the formation of bulbils, first promoting their initiation and subsequently inhibiting their growth[6]. In Lycoris radiata, the peak levels of ZR/GA3 and ZR/IAA occur prior to the formation of above-ground bulblets[25]. Moreover, the exogenous application of the GA biosynthesis inhibitor paclobutrazol (PBZ) exhibits inhibitory effects on bulblet development in lilies at high concentrations and promotive effects at low concentrations[26]. However, the specific effect of exogenous hormones on the initiation or development of bulblets remains unclear.
N-(2-Chloro-4-pyridyl)-N'-phenylurea (CPPU, a synthetic cytokinin analogue) is commonly used for fruit set induction and expansion[27−31]. For example, CPPU application induced sugar accumulation during grape (Vitis vinifera L.) development by up-regulating the expression of INVs[28]. It also enhanced the weight and size of kiwifruit by increasing glucose and soluble sugar levels[29]. In terms of cell development, CPPU has been reported to primarily promote hypanthium cell division and expansion. It induced the content of IAA by up-regulating the expression of indole-3-pyruvate monooxygenase (YUCCA) and reduced the content of ABA by down-regulating 9-cis-epoxycarotenoid dioxygenase (NCED)[30], while the CPPU-induced melon fruit set is dependent on GA biosynthesis[31]. In addition to its application in fruits, CPPU treatment may also promote the enlargement of bulblets by enhancing starch accumulation and amylase activity in Lycoris radiate[32,33].
Currently, there are only two active cytokinins, namely, isopentenyladenine and its hydroxylated derivative zeatin (ZT). These cytokinins can form various conjugates and involve enzymes/isozymes with different substrate specificities in their biosynthesis and interconversion[34,35]. 2-isopentenyladenine (2-iP) is one of the synthetic analogues of isopentenyladenine. Previous studies have primarily used 2-iP in culture medium to induce plant tissue regeneration and embryogenesis. For example, Murashige and Skoog (MS) medium containing silver nitrate and 2-iP was demonstrated to promote somatic embryogenesis in date palm (Phoenix dactylifera L.)[36]. Low levels of 2-iP improved the general responsiveness of root-derived microcallus tissue, which was used for shoot induction and plant regeneration[37]. Moreover, 2-iP has been reported to induce direct somatic embryogenesis in Arabica coffee (Coffea arabica L.)[38]. However, the specific effect of CPPU and 2-iP on bulblet formation in lily scale cuttings has yet to be reported.
To explore more efficient scale propagation methods, we treated lily scales with CPPU and 2-iP, and found that both treatments promoted bulblet formation by facilitating the initiation and development processes. By analyzing the changes in genes related to carbohydrate metabolism and hormone levels, this study investigated the mechanisms underlying the promotion of bulblet formation by CPPU and 2-iP, providing new insights into the formation of lily bulblets and offering an efficient method for vegetative propagation in lilies.
-
'Matrix' bulbs were harvested on October 5, 2022, subsequently stored in the National Lily Germplasm Bank at the Beijing Academy of Agriculture and Forestry Sciences (BAAFS) (S 116°17', W 39°56'), China, at a temperature of 4 °C. The experiments were conducted in December 2022 at a plant cultivation room of the Lily Research Group of BAAFS. The temperature was maintained at 23 ± 2 °C. Healthy scales without disease spots or damage were separated from the bulbs and soaked in a diluted carbendazim solution (1:500 dilution) for 30 min. Following this, the scales were washed with clean water 3–5 times, placed on perlite, and maintained in darkness with the trays covered to preserve humidity.
Treatment
-
CPPU (CF5381, Coolaber, Beijing, China) powder was dissolved in absolute ethyl alcohol and subsequently diluted with distilled water to prepare a stock solution at a concentration of 1 g/L. The powder of 2-iP (CI6611, Coolaber, Beijing, China) was dissolved in 1 M NaOH and diluted with distilled water to prepare a stock solution at a concentration of 0.05 g/L. The treatments were divided into the following seven groups for the bulblet initiation experiment: three concentrations of CPPU (10 mg/L, 50 mg/L, and 100 mg/L); three concentrations of 2-iP (0.05 mg/L, 0.1 mg/L, and 0.5 mg/L); and a control group treated with distilled water. The treatments were applied every 2 d, and samples were taken every 4 d until clear protrusions were observed at all scale bases, indicating the completion of the bulblet initiation process. The bulblet initiation rate (number of scales with basal swelling/total number of scales) for each treatment group was recorded every 4 d. A total of 30 scale bases were collected at each sampling and stored at −80 °C for further analysis. During the bulblet development experiment, three treatments (10 mg/L CPPU, 0.1 mg/L 2-iP, and distilled water) were applied. The scale, which was of a uniform size and had previously undergone 16 d of cutting, was referred to as the BD0d (bulblet development 0 days) stage. Treatments were applied every 2 d, and bulblets from 30 scale bases were sampled every 8 d and stored at −80 °C for further analysis. The scale bases were observed and photographed using a Motic microscope (SMZ-168 SERIES, China) equipped with the Zeiss Digital camera (AxioCam ICc 5, China).
RNA extraction, cDNA synthesis, and qRT-PCR
-
The samples obtained from the initiation and development of bulblets were analyzed using real-time quantitative PCR (qRT-PCR). Total RNA was extracted from the samples using an RNA extraction kit (RC401, Vazyme, Nanjing, China). First-strand cDNA synthesis was performed according to the manufacturer's instructions using a reverse transcription kit (R333, Vazyme, Nanjing, China). Specific primers for qRT-PCR analysis were designed using GenScript (
https://www.genscript.com ). The F-box family protein (FP) gene was used as the internal reference gene for normalization. Supplemental Table S1 reports the primer sequences. The qRT-PCR experiment was performed using TB Green® Premix Ex Taq™ II (RR820A, Dalian, China) on a Bio-Rad CFX96TM Real-Time System (California, USA). The 2−ΔΔCᴛ method was employed to analyze qRT-PCR expression data according to Zhang et al.[39]. Three biological replicates were performed for all qRT-PCR assays. Significant differences in the development assay were determined using Student's t-tests (*p < 0.05; **p < 0.01).Determination of endogenous hormone contents
-
Bulblets from scales treated with 10 mg/L CPPU, 0.1 mg/L 2-iP, and distilled water were harvested on the 12 d of bulblet initiation and the BD16d of bulblet development. Each biological replicate had a fresh weight of approximately 0.1 g. Sample pre-processing steps: each sample was ground in a mortar and mixed with 1 mL of pre-cooled 70%–80% methanol solution at a pH of 3.5. The sample was immersed overnight at 4 °C and then centrifuged at 12,000 g for 10 min at 4 °C. The residue was extracted with 0.5 mL of 70%–80% methanol solution at 4 °C for 2 h. After centrifugation, the supernatant was collected and combined with the supernatant obtained from the two previous extractions. The combined supernatant was evaporated to one-third of the original volume by vacuum evaporation at 40 °C, and an equal volume of petroleum ether was then added. Following phase separation, the supernatant was decolorized by repeated extraction (2–5 times) and adjusted to a pH of 8.0 using triethylamine. The mixture was incubated and shaken for 20 min after the addition of Polyvinylpolypyrrolidone (PVPP) (CP9301, Coolaber, Beijing, China) and the supernatant was collected after centrifugation. The sample was then adjusted to a pH of 3.0 with hydrochloric acid and extracted three times with ethyl acetate. The ester phase was subsequently combined with the sample and evaporated to dryness at 40 °C under reduced pressure. The combined phase was then dissolved in the mobile phase and filtered through a syringe filter for analysis. Then, they were sent to Suzhou Grace Biotechnology Co., Ltd and analyzed for endogenous hormone contents via high performance liquid chromatography (HPLC).
Determination of endogenous sugars
-
The contents of sucrose, glucose, and starch were measured using a Sucrose Content Assay Kit (BC2460, Solarbio, Beijing, China), a Glucose Content Assay Kit (BC2500, Solarbio, Beijing, China), and a Starch Content Assay Kit (BC0700, Solarbio, Beijing, China), respectively. All detection assays were conducted using a 0.1-g sample, and three biological replicates were analyzed. Significant differences were determined using Student's t-tests (*: p < 0.05; **: p < 0.01).
-
'Matrix', a lily cultivar, belongs to the Asiatic hybrid cultivars and has gained popularity due to its small size, vivid flower colors, and suitability for home cultivation (Fig. 1). Scale-cutting is commonly employed for the vegetative propagation of 'Matrix'. To improve the efficiency of scale-cutting, we performed experiments using CPPU and 2-iP. The scales were treated with three concentrations of CPPU and three concentrations of 2-iP, while distilled water was used as the control group. We observed the changes occurring at the base of the mother scales, based on morphological analysis and histological characteristics, the presence of distinct protrusions at the base of the scales was identified as evidence of bulblet initiation (Supplemental Fig. S1a & b, Fig. 2a; water treatment for approximately 16 d). Subsequently, the protuberances developed into bulblets, a process referred to as bulblet development (Supplemental Fig. S1). The results revealed that all three CPPU and 2-iP concentrations promoted bulblet initiation compared to the control group. Significant differences were observed between the control and treatment groups at 8 and 12 d, with the greatest differences noted for the latter time point (Fig. 2a, b). Compared to the control group, the CPPU treatments at 10 mg/L, 50 mg/L, and 100 mg/L resulted in bulblet initiation rates that were 4.98, 3.48, and 2.41 times higher, respectively. Likewise, the 2-iP treatments at 0.05 mg/L, 0.1 mg/L, and 0.5 mg/L led to bulblet initiation rates that were 2.80, 4.07, and 1.98 times higher, respectively. The results indicate 10 mg/L CPPU and 0.1 mg/L 2-iP as the most effective treatments under our experimental conditions, and thus further analyses were conducted using these concentrations.

Figure 1.
The lily cultivar 'Matrix'. (a) The 'Matrix' at 30 DAP (days after planting into the soil). Dormancy-broken bulbs were used for planting. (b) The 'Matrix' at 50 DAP. The first flower opens approximately 45 DAP. (c) The potted 'Matrix'. Scale bars = 5 cm.
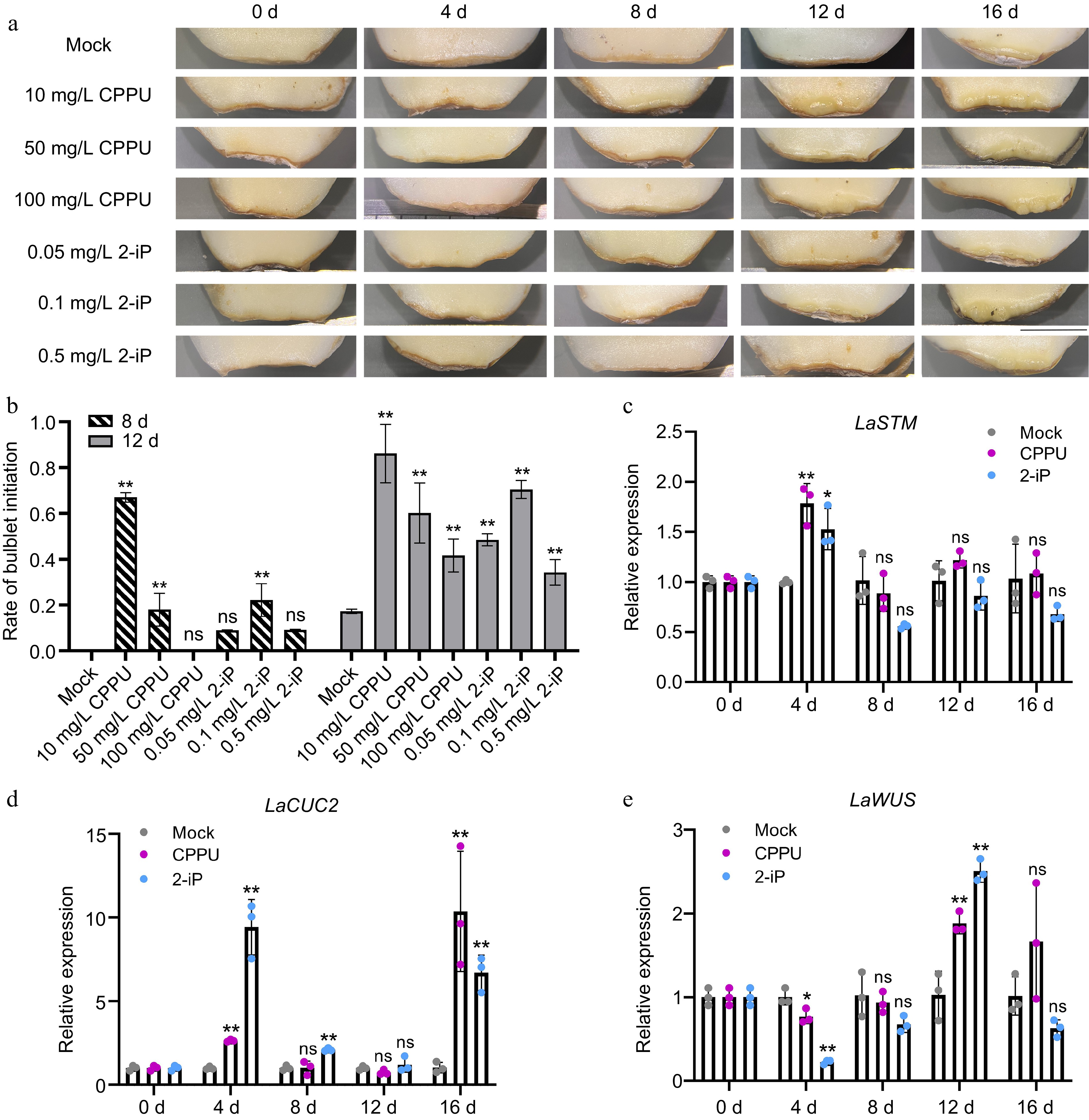
Figure 2.
Effects of CPPU and 2-iP treatments on the initiation rate of bulblets and the relative expression of organogenesis-related genes. The scales were derived from 'Matrix' bulbs stored at low temperature (4 °C) for two months. The data were collected every 4 d (0 d, 4 d, 8 d, 12 d, 16 d). (a) Morphological analysis of the initiation of the bulblet under different treatments. Bar = 0.5 cm. (b) CPPU (10 mg/L) and 2-iP (0.1 mg/L) treatments resulted in the occurrence of more bulblets at the base of scales than the control treatment (Mock). Data are from 8 and 12 d, n = 30 scales per treatment. Expression pattern of organogenesis-related genes, (c) LaSTM, (d) LaCUC2, and (e) LaWUS in the Mock, 10 mg/L CPPU, and 0.1 mg/L 2-iP treatments. qRT-PCR values were determined using the 2−ΔΔCᴛ approach. The control group for each time point was set as 1, and the treatment group was compared to its respective control group. Data are presented as the means of three biological replicates with standard deviation (SD) error bars (Student's t-test; *: p < 0.05; **: p < 0.01; ns: no significance).
The initiation process of bulblets is similar to that of axillary meristems (AXMs) in species such as Arabidopsis thaliana, Pisum sativum, and Lycoris radiata[8,40]. Genetic studies have identified several genes that specifically regulate the initiation of AXMs. During the AXM initiation process, SHOOT MERISTEMLESS (STM), a well-known marker gene, is induced by CUP-SHAPED COTYLEDON (CUCs), and subsequently activates AXMs in the leaf axil[41]. Furthermore, the expression of WUSCHEL (WUS) is de novo activated to establish a new AXM organization center. In our results, LaSTM and LaCUC2 were significantly induced by CPPU and 2-iP at 4 d (Fig. 2c, d), and there was also an interesting additional induction of LaCUC2 at 16 d (Fig. 2d). The upregulated gene expressions of LaWUS may indicate the establishment of AXMs[42], with a significant induction at 12 d during bulblet initiation (Fig. 2e). These findings demonstrate the promoting effect of CPPU and 2-iP on bulblet initiation. Further, we found a significant positive correlation between the expression of LaSTM at 4 d and the bulblet initiation rate (Supplemental File 1), indicating that LaSTM plays a key role in the process of CPPU and 2-iP-induced bulblet initiation.
CPPU and 2-iP alter the signal transduction of endogenous cytokinin and the levels of GA3 and IAA
-
The signal transduction of cytokinin involves a multi-step phosphorylation process that includes cytokinin receptors (HKs), histidine-containing phosphotransfer proteins (AHPs), and type-B response regulators (RRs)[43]. Combining previous studies[6,18,20], we examined the expression patterns of four genes by qRT-PCR (Fig. 3). The results showed that the expression of LaAHK2 was significantly induced by CPPU and 2-iP at 4 and 8 d, with no significant differences observed in the later stages compared to the control group (Fig. 3a). LaAHK3 exhibited a highly significant response to the 2-iP treatment, and mainly responded to CPPU at 16 d during bulblet initiation (Fig. 3b). In addition, LaAHK4 displayed continuous induction by CPPU, while a significant response to 2-iP was only observed at 16 d (Fig. 3c). ARR1 plays a crucial role in AXM initiation by upregulating the expression of STM[41]. However, unlike the LaAHKs, the response of LaARR1 to CPPU and 2-iP was not statistically significant (Fig. 3d), indicating the potential role of the remaining LaARRs as primary effectors in cytokinin-mediated bulblet initiation.
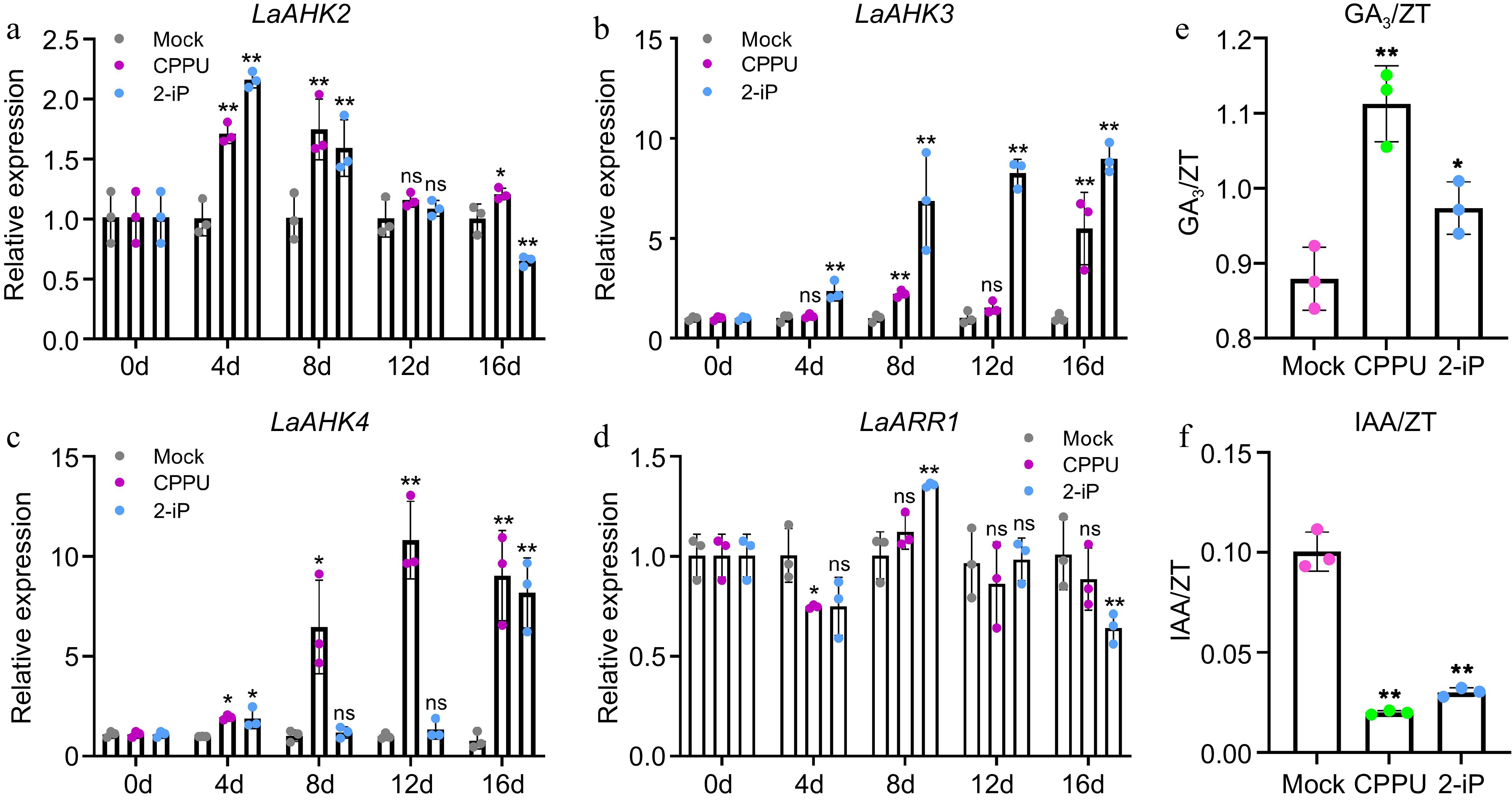
Figure 3.
Expression patterns of cytokinin signaling pathway genes and endogenous hormones with ZT in the control, 10 mg/L CPPU, and 0.1 mg/L 2-iP treatments during bulblet initiation. Relative expression of cytokinin signal genes, (a) LaAHK2, (b) LaAHK3, (c) LaAHK4, and (d) LaARR1 in the control, CPPU, and 2-iP treatments. The control group for each time point was set as 1, and the treatment group was compared to its respective control group. (e), (f) Endogenous hormones (GA3 and IAA) with ZT in the control, CPPU, and 2-iP treatments at 12 d. Data are presented as the means of three biological replicates with SD error bars (Student's t-test; *: p < 0.05; **: p < 0.01; ns: no significance).
As the process of bulblet formation is regulated by multiple hormones[18,44], the levels of three hormones in the basal part of the mother scale were measured at 12 d. The primary natural active ingredient of cytokinin, ZT, was observed to significantly increase under the CPPU and 2-iP treatments (Supplemental Fig. S2a). The trend in GA3 content was similar to that of ZT, and both were significantly induced by CPPU and 2-iP (Supplemental Fig. S2b). The role of IAA in the process of AXB initiation is complex[4,6]. However, at 12 d (during the initiation of bulblets) the IAA content was significantly suppressed by the CPPU and 2-iP treatments (Supplemental Fig. S2c). In order to investigate how GA3 and IAA interact with CKs and their effect on the bulblet initiation process, we calculated the ratios of GA3 and IAA to ZT. The ZT content of the same treatment group was used as the control. The results showed that compared to the control group, the values of GA3/ZT increased by 26.5% and 10.7% under the CPPU and 2-iP treatments, respectively (Fig. 3e), while the values of IAA/ZT were inhibited by 80.1% and 70% by CPPU and 2-iP, respectively (Fig. 3f). Moreover, there was a significant positive correlation between the accumulation of GA3 and the bulblet initiation rate, while the content of IAA showed a significant negative correlation with the bulblet initiation rate (Supplemental File 1). These findings suggest that GA3 may synergistically regulate the bulblet initiation with ZT, while IAA and ZT may have an antagonistic regulatory relationship in this process.
CPPU and 2-iP enhance the conversion of sucrose during bulblet initiation
-
The process of organogenesis is often accompanied by carbohydrate metabolism, particularly during lily scale-cutting, where sucrose and starch play critical roles as the primary sources of energy for bulblet formation[8,45]. In order to investigate the effects of the CPPU and 2-iP treatments on the sucrose conversion process during bulblet initiation, the expressions of the sucrose metabolism genes LaSusy1/2 and sucrose cell wall invertase genes LaCWIN1/3/4 were examined. Compared to the control groups at each time period, the expression of LaSusy1/2 was significantly upregulated by the CPPU and 2-iP treatments (Fig. 4a, b). The expressions of LaCWIN1 and LaCWIN4 were inhibited by the CPPU and 2-iP treatments at 4 d, but induced in the later stages of bulblet initiation (Fig. 4c, e). Additionally, after being induced by CPPU at 4, 8, and 12 d, the expression of LaCWIN3 did not significantly differ from the control at 16 d. Interestingly, LaCWIN3 was significantly induced by 2-iP in the early stage of bulblet initiation, and there was a further induction of LaCWIN3 at 16 d (Fig. 4d). These results indicate that CPPU and 2-iP induced the expression of most genes related to sucrose conversion. Furthermore, the sucrose content in the mother scale bases decreased under the CPPU and 2-iP treatments, with the exception of an increase in sucrose content induced by CPPU and 2-iP at 4 d (Fig. 4f). However, compared to 0 d, the sucrose content decreased during bulblet initiation in all treatment groups. Additionally, there was a significant negative correlation between the sucrose content at 12 d and the bulblet initiation rate (Supplemental File 1). These findings indicate potential ability of CPPU and 2-iP to promote the utilization of sucrose at the basal part of the mother scale during the bulblet initiation process.
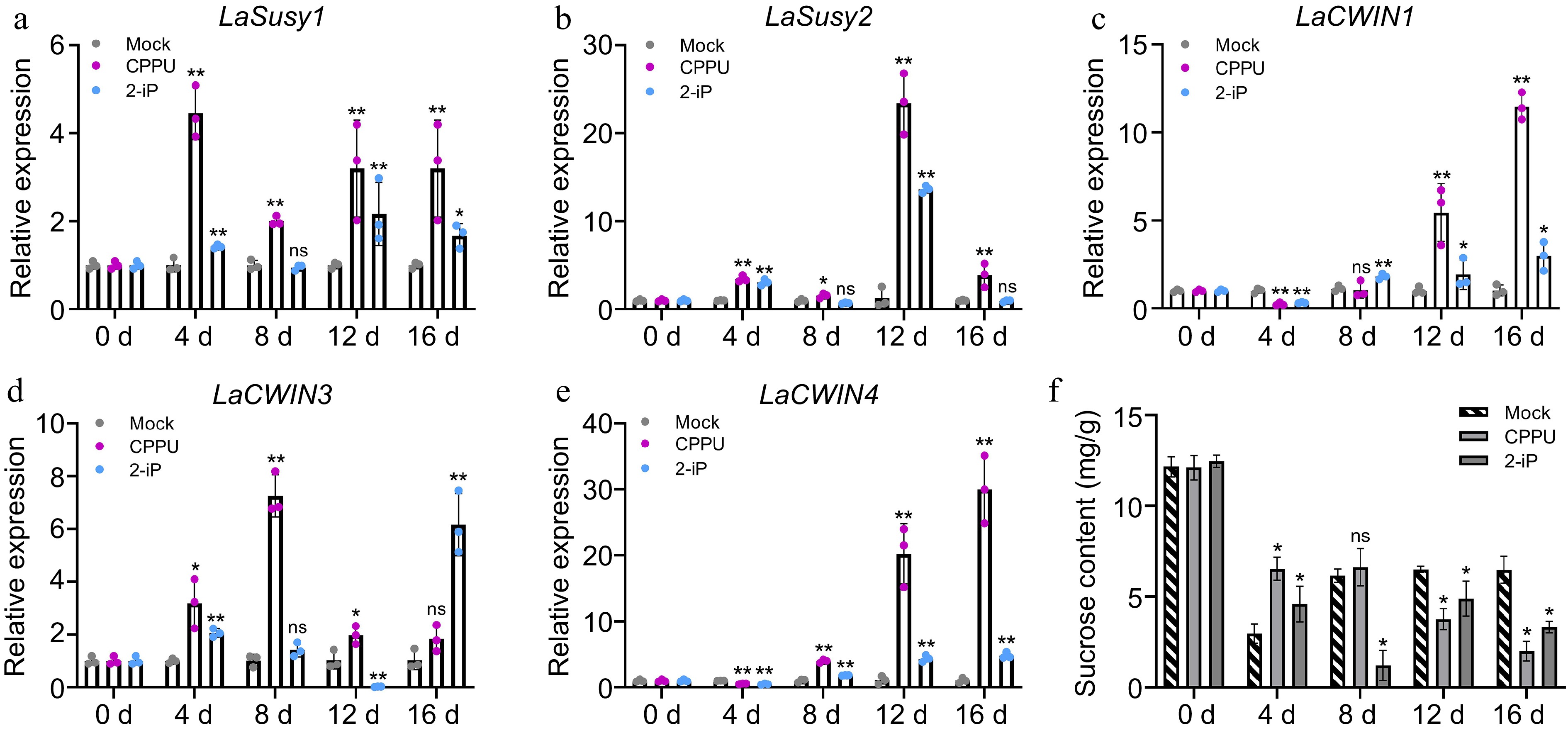
Figure 4.
Sucrose content and expression pattern of sucrose metabolism-related genes in the control, 10 mg/L CPPU, and 0.1 mg/L 2-iP treatments during the bulblet initiation process. Relative expression of (a) LaSusy1, (b) LaSusy2, (c) LaCWIN1, (d) LaCWIN3 and (e) LaCWIN4 in the control, CPPU, and 2-iP treatments. The control group for each time point was set as 1, and the treatment group was compared to its respective control group. (f) Sucrose content in the control, CPPU, and 2-iP treatments. Data are presented as the means of three biological replicates with SD error bars (Student's t-test; *: p < 0.05, **: p < 0.01; ns: no significance).
CPPU and 2-iP treatments promote bulblet development
-
The CPPU and 2-iP treatments were applied to scales treated with water for 16 d (labeled as BD0d, bulblet development at 0 d) to investigate their impact on the bulblet development process. Samples were collected at BD8d and BD16d for data collection and statistical analysis. The results indicate CPPU and 2-iP to enhance the diameter and weight of the bulblets (Fig. 5a). At BD16d, the diameter of the bulblets in the CPPU and 2-iP treatment groups increased by 54.2% and 47.8%, respectively, compared to the control (Fig. 5b). Moreover, the weight of the bulblets in the CPPU and 2-iP treatment groups was 2.8 and 1.9 times that of the control plants, respectively (Fig. 5c). Based on the promotion of bulblet diameter by CPPU and 2-iP, the upregulation of two genes related to cell expansion, LaEXPA8 and LaEXPA10, was detected (Fig. 5d, e). The expression levels of them showed a significant positive correlation with the diameter and weight of the bulblets (Supplemental File 1). These results demonstrate the significant promotion of the bulblet development by CPPU and 2-iP.
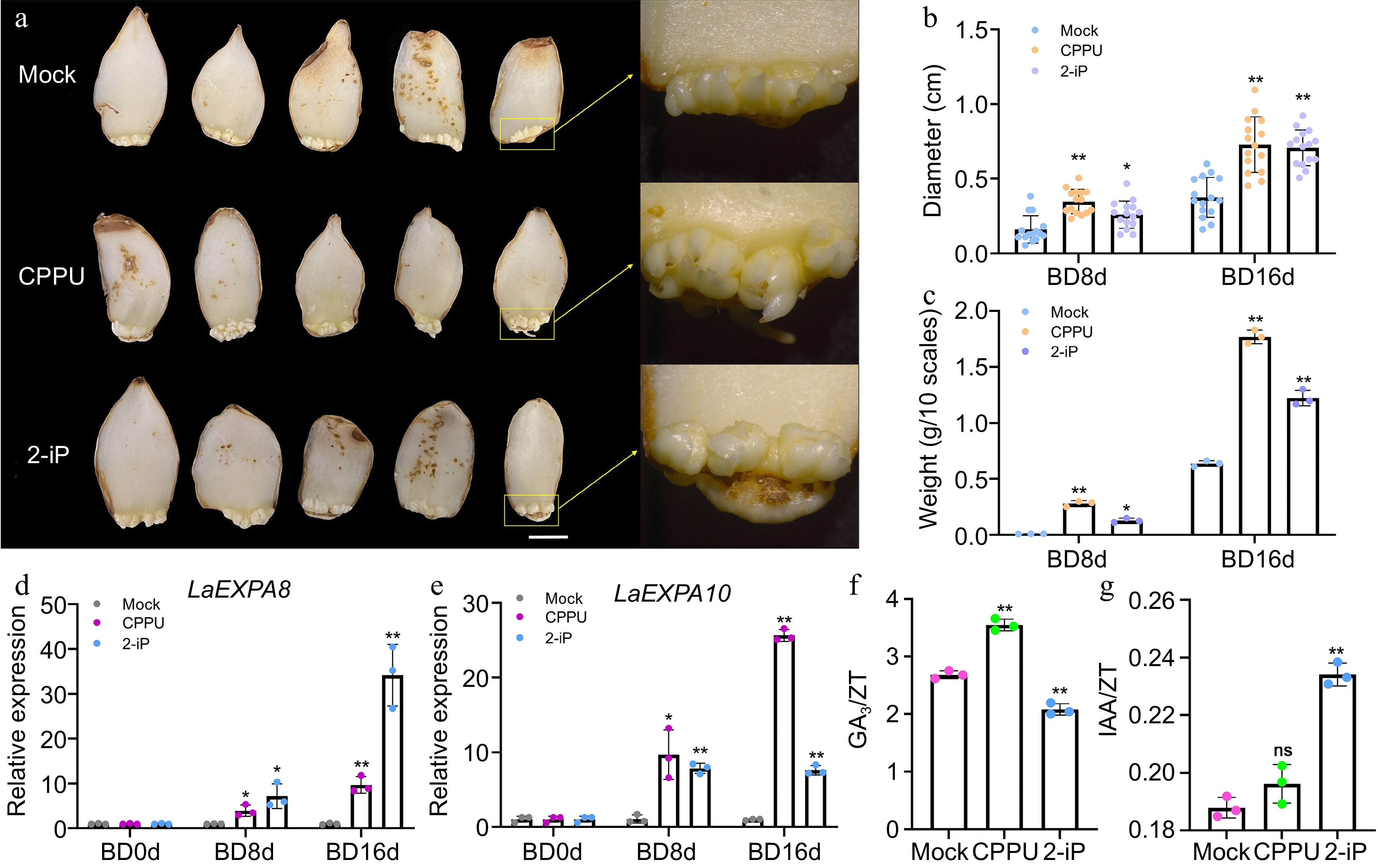
Figure 5.
10 mg/L CPPU and 0.1 mg/L 2-iP treatments promote bulblet development. (a) Morphological analysis of the development of the 'Matrix' bulblet under different treatments at BD16d. Bar = 1 cm. (b) Diameter and (c) weight of bulblets in the control, CPPU, and 2-iP during bulblet development. Each dot on panel (b) represent the average diameter of bulblets generated by a single maternal scale, n = 15. Each dot on panel (c) represents the weight of bulblets produced from 10 maternal scales. Relative expression of (d) LaEXPA8 and (e) LaEXPA10 in the control, CPPU, and 2-iP treatment groups during bulblet development. The control group for each time point was set as 1, and the treatment group was compared to its respective control group. (f)–(g) Endogenous GA3/ZT and IAA/ZT in the control, CPPU, and 2-iP treatments at BD16d during bulblet development. Data are presented as the means of three biological replicates with SD error bars (Student's t-test; *: p < 0.05; **: p < 0.01; ns: no significance).
CPPU and 2-iP treatments alert endogenous hormones during bulblet development
-
The bulblet development is also regulated by multiple hormones[8]. By quantifying the levels of endogenous hormones in the bulblets at BD16d under different treatments, we observed that both CPPU and 2-iP significantly increased the ZT content (Supplemental Fig. S3a). The CPPU treatment significantly enhanced GA3 levels, whereas the effect of 2-iP on GA3 promotion was less pronounced (Supplemental Fig. S3b). In contrast to the bulblet initiation, both CPPU and 2-iP significantly increased IAA levels within the bulblets during bulblet development (Supplemental Fig. S3c). Furthermore, we calculated the ratios GA3/ZT and IAA/ZT to evaluate the relationship of GA3 with IAA and ZT in this process. The results demonstrated that the synergistic effect of GA3 and ZT was more pronounced during CPPU-induced bulblet development, whereas the 2-iP treatment reduced GA3/ZT (Fig. 5f). Interestingly, the IAA/ZT ratio suggests that IAA may have a synergistic promoting effect with ZT during bulblet development, particularly when treated with 2-iP (Fig. 5g).
CPPU and 2-iP altered carbohydrate metabolism during bulblet development
-
As vegetative reproductive and storage organs in lilies, the development of bulblets is closely associated with internal carbohydrate changes[5,8,46]. By quantifying the sugar content in bulblets, we observed a decrease in both sucrose and glucose levels from BD8d to BD16d (Fig. 6a, b). Moreover, the sucrose content was reduced in the CPPU and 2-iP treatment groups compared to the control group (Fig. 6a). Glucose levels were induced by the CPPU treatment at both BD8d and BD16d, whereas a response to 2-iP was only observed at BD8d (Fig. 6b). No difference in glucose content was observed between the 2-iP treatment group and the control group at BD16d. Starch content increased during bulblet development, and this increase was significantly enhanced by CPPU and 2-iP, indicating that CPPU and 2-iP promote starch synthesis by influencing carbohydrate metabolism (Fig. 6c). Interestingly, CPPU and 2-iP promoted the accumulation of soluble proteins at BD8d, but decreased their content at BD16d (Fig. 6d), revealing that CPPU and 2-iP may enhance the utilization of soluble proteins during the late stage of bulblet development.
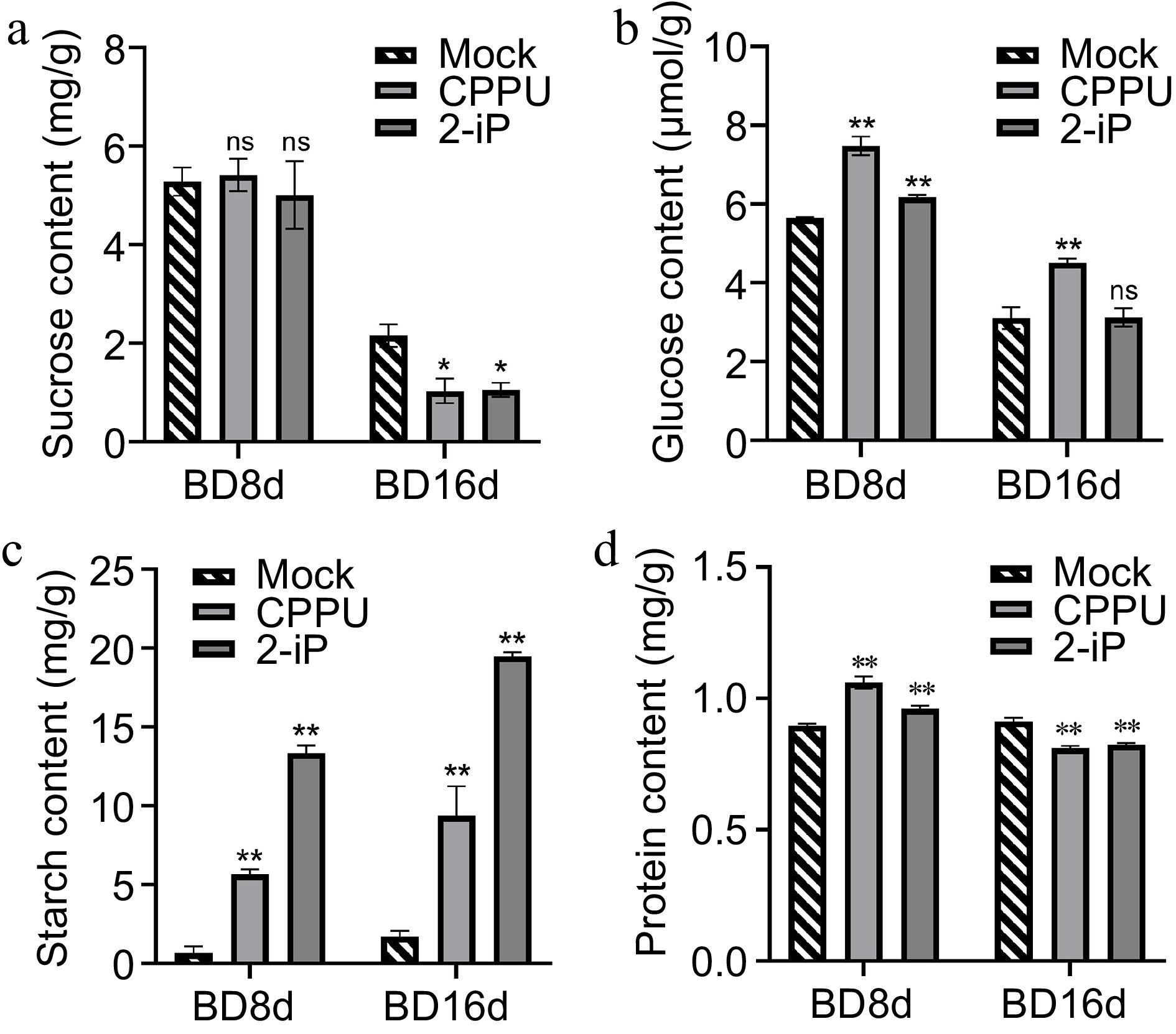
Figure 6.
Endogenous sucrose, glucose, starch, and protein content in the control, 10 mg/L CPPU, and 0.1 mg/L 2-iP treatments at BD16d. Data are presented as the means of three biological replicates with SD error bars (Student's t-test; *: p < 0.05, **: p < 0.01; ns: no significance).
Based on the above results, we conducted further examinations on the expression of genes associated with sugar metabolism. The three LaCWIN genes displayed distinct expression patterns, with LaCWIN1 and LaCWIN3 exhibiting predominant responses to the CPPU and 2-iP treatments during the late stage of bulblet development (Fig. 7a, b), whereas LaCWIN4 was induced by CPPU at BD8d (Fig. 7c). Moreover, the enzyme LaSAI, a soluble acid invertase involved in the conversion of sucrose, selectively responded to 2-iP at BD8d, but was induced by both CPPU and 2-iP during the late stage of bulblet development (Fig. 7d). LaSusy1 responded to the CPPU and 2-iP treatments at BD8d, yet no significant differences were observed compared to the control group at BD16d (Fig. 7e). LaSusy2 responded to both the CPPU and 2-iP treatments at both time points (Fig. 7f). Similar to LaCWIN1/3, LaSUT2 was exclusively induced at BD16d (Fig. 7g). In addition, we examined the expression levels of LaTPS1 and LaTPS6 to validate alterations in downstream sugar signaling. Compared to the control, both CPPU and 2-iP significantly enhanced the expression of LaTPS1 and LaTPS6 (Fig. 7h, i). Furthermore, we found a significant positive correlation between the expression levels of LaCWIN1/3, LaSAI, LaSUT2, and LaTPS1/6 with the diameter and weight of the bulblets (Supplemental File 1). Together with the sugar content observations, these findings provide evidence that the CPPU and 2-iP treatments substantially stimulate sugar metabolism during bulblet development, thereby enhancing bulblet diameter and weight.
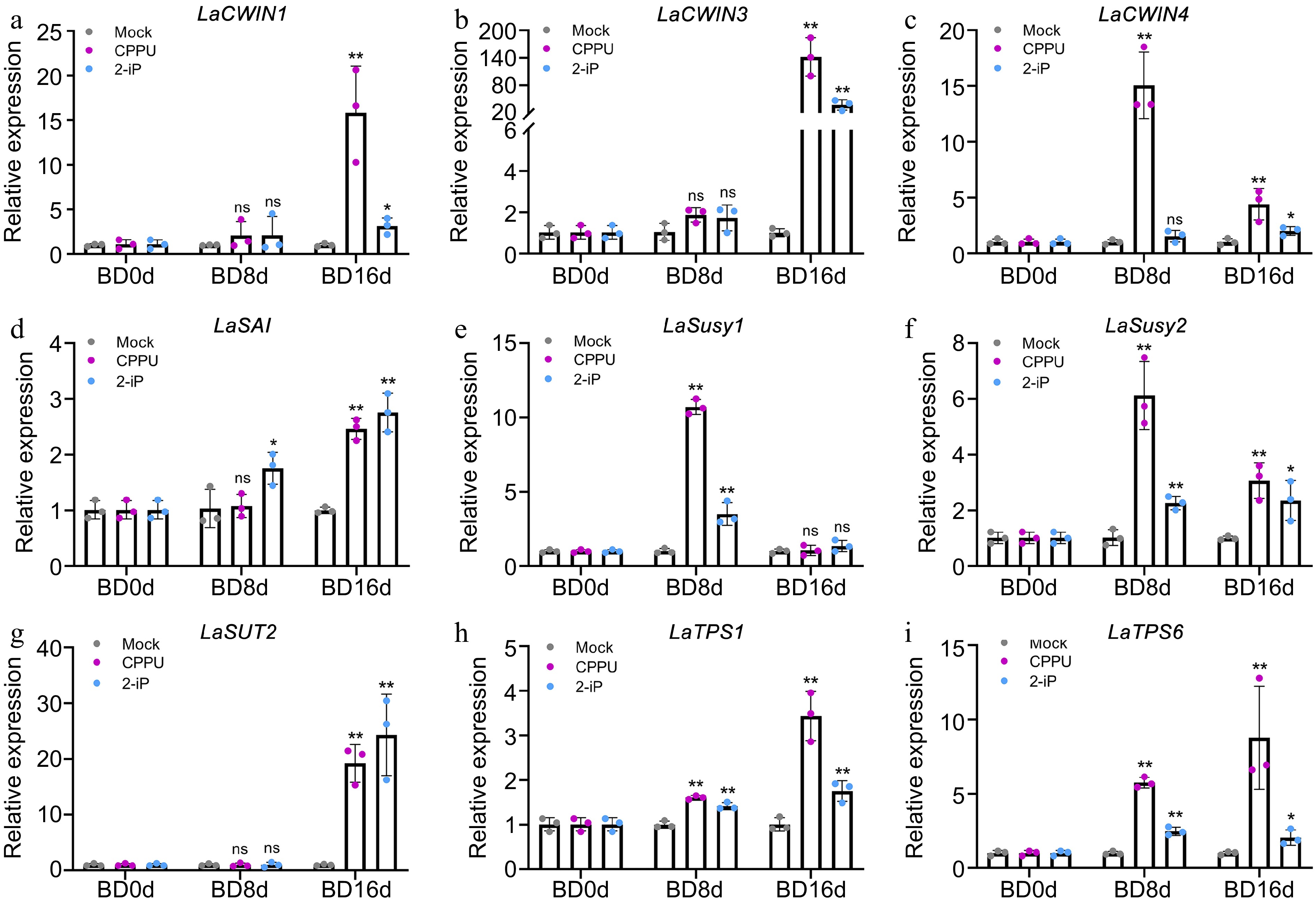
Figure 7.
Expression patterns of sugar metabolism-related genes in the control, 10 mg/L CPPU, and 0.1 mg/L 2-iP treatments during bulblet development. The control group for each time point was set as 1, and the treatment group was compared to its respective control group. Data are presented as the means of three biological replicates with SD error bars (Student's t-test; *: p < 0.05, **: p < 0.01; ns: no significance).
-
Scale cutting is the most effective and widely used technique for the reproduction of lilies worldwide. Combining it with exogenous treatments during the cutting procedure can accelerate the commercial production of lily bulbs. Nevertheless, the molecular mechanisms governing bulblet formation remain unclear, and more effective and reliable exogenous treatments need to be explored.
CPPU and 2-iP promote bulblet initiation by upregulating organogenesis-related genes, enhancing sucrose utilization and altering the ratio of GA3/ZT and IAA/ZT
-
The formation of bulblets follows a similar process to that of AXB formation[4]. This process involves two distinct stages, the initiation and subsequent development (outgrowth)[7,11]. During the initiation process, the AXM is derived from a cluster of stem cells expressing the STM gene, which originates from the leaf axil (leaf base)[41,47]. A low level of auxin is required to maintain the STM cell cluster and initiate the AXMs[4,48]. The STM gene expression is upregulated by the ATH1 (ARABIDOPSIS THALIANA HOMEOBOX GENE1)-STM complex, the LAS-REV (LATERAL SUPPRESSOR-REVOLUTA) module, and CUC, indicating the activation of AXMs[41]. The initiation of the WUS indicates the establishment of the AXM, which plays a crucial role in both adventitious bud formation and AXM initiation[42]. We found that both CPPU and 2-iP induce the expression of LaSTM, LaCUC2, and LaWUS and promote the initiation of bulblets (Fig. 2c−e). This supports the similarity between the initiation of bulblets and AXM.
As reported in the initiation process of adventitious buds, a low auxin/cytokinin ratio promotes adventitious shoot regeneration, while a high ratio promotes root induction[49]. However, the role of auxin in bulblet formation remains unclear. In scale propagation, manually separating scales from the bulb rapidly alters the distribution of endogenous auxin[50]. Auxin accumulates at the basal part of the scale close to the axis, leading to asymmetric cell proliferation and promoting the formation of bulblets. Previous studies have suggested that auxin plays a dual role in bulblet formation, with an initial increase in IAA concentration promoting the regeneration of meristematic tissue, followed by a decrease that maintains proper hormone homeostasis in the newly-formed tissues[6]. Podwyszynska[51] found that the application of exogenous auxin effectively promoted the formation of tulip bulbs. However, the formation of bulblets in Lycoris and lily is associated with relatively high levels of both CK/IAA and ZR/IAA[25,52]. Our results revealed that the rapid initiation of bulblets induced by CPPU and 2-iP is accompanied by a decrease in auxin levels and a low ratio of IAA/ZT (Supplemental Fig. S2c, Fig. 3f). Therefore, we speculate that an initial low level of auxin may be required for the initiation of bulblets. The role of CKs in promoting the initiation of bulblets and AXB formation has been well established[18,42,48]. Moreover, the degradation of starch granules in the mother scale serves as an energy source for bulblet initiation and development[5,40]. Exogenous treatment with 6-BA induces bulblet formation by activating genes related to cytokinin signaling, such as LlAHK2/3/4 and LlARR1/2/12, this activation leads to an early decrease in sucrose and soluble sugar content[13,14,18], which is consistent with our findings (Figs 3, 4). Although several reports have emphasized the role of GA in bulblet formation[25], the specific steps primarily influenced by GA remain unclear. Here, the rapid initiation of bulblets induced by CPPU and 2-iP is accompanied by an accumulation of GA3 as well as a high GA3/IAA ratio (Supplemental Fig. S2b, Fig. 3e). These findings suggest a potential positive role of GA in bulblet initiation, despite the possibility of CPPU promoting GA biosynthesis[31].
While the positive and significant relationship between sucrose metabolism and axillary organogenesis has been demonstrated[8, 13−16], the underlying mechanisms remain elusive. Through correlation analysis between the expression of sucrose metabolism-related genes at different time points and the bulblet initiation rate, we identified significant positive correlations between the expression of LaSusy1/2 and LaCWIN3 at 4 d, with the bulblet initiation rate. These correlations could be associated with the transport of sucrose towards the mother scale base, stimulated by CPPU and 2-iP treatments[12]. Given the induction of AXMs by CPPU and 2-iP, all detected sucrose metabolism-related genes exhibited significant positive correlations with the bulblet initiation rate, also showed highly positive correlations with the expression of LaSTM (Supplemental File 1). This implies that CPPU and 2-iP facilitate the formation of axillary meristems by promoting the conversion of sucrose to glucose at the scale base, thereby promoting bulblet initiation[8, 13−14].
All in all, our results suggested that CPPU and 2-iP treatments may alter the interaction between carbohydrates and endogenous hormone levels during bulblet initiation.
CPPU and 2-iP promote the metabolism of sugar and starch during bulblet development
-
Various endogenous factors, including hormones, sugars, and transcription factors, regulate the development stage of AXBs[53]. Sugar promotes the growth of AXBs, and there is increasing evidence suggesting the involvement of T6P in this process[16,54]. Unlike the outgrowth of AXBs, the development of bulblets is accompanied by the accumulation of starch and the enlargement of the bulblets (Figs 5, 6)[5,14]. Sugar and starch metabolism are tightly regulated in this process, involving various enzymes including CWIN, SUS, UDP-glucose pyrophosphorylase (UGPase), and starch synthesis enzymes (SUS, granule-bound starch synthase (GBSS), and ADP-glucose pyrophosphorylase (AGPase))[8,13]. In the process of bulblet development, the increase in soluble sugar content also promotes the expression level of CycD and accelerates cell division[8]. These processes indicate the consumption of sucrose and soluble sugars, as well as the accumulation of starch in bulblet development. The promotion of bulblet development by CPPU and 2-iP further support the observations of sugar consumption, initial accumulation, and the subsequent decrease of glucose, as well as an increase in starch content (Fig. 6a−c). Moreover, correlation analysis demonstrated a significant positive relationship between the expression of sucrose metabolism-related genes and starch content during CPPU and 2-iP-induced bulblet development. However, likely as a result of sucrose and glucose consumption, their levels exhibited a significant negative correlation with the diameter and weight of the bulblets (Supplemental File 1). Additionally, we determined the content of soluble proteins to be consistent with the changes in glucose content, suggesting that the process of bulblet development induced by CPPU and 2-iP also enhances protein utilization (Fig. 6d).
CPPU and 2-iP increase the levels of endogenous GA3 and IAA during bulblet development
-
The inhibitory mechanism of auxin on the outgrowth of AXBs is not fully understood, particularly when auxin appears to act indirectly, generally moving within the main stem rather than entering the AXBs[55]. A recent study has demonstrated that, in the early stages, AXB outgrowth induced by decapitation and CKs is independent of auxin flux in the buds[56]. Therefore, auxin may indirectly regulate branching by modulating other hormones such as CKs and strigolactones[53,55−57]. The role of auxin in the process of bulblet development is also not well elucidated. Several studies have suggested that high levels of IAA or exogenous auxin treatment may stimulate bulblet formation[22, 44]. The application of NAA enhances the number of lily bulblets[58], while in Hyacinthus orientalis, the use of IBA exerts a positive effect on bulb proliferation[59]. However, in Hippeastrum vittatum (Amaryllidaceae), the application of exogenous auxin does not result in scale propagation or bulblet development, but significantly increases the number of rotting scales[60]. Therefore, further research is required to understand the regulatory role of auxin in bulb development. We found that although auxin appears to be antagonistic to CPPU and 2-iP in the initiation process of bulblets (Supplemental Fig. S2c, f), it seemingly promotes the development of bulblets, and its level is significantly induced by CPPU and 2-iP (Supplemental Fig. S3c). Moreover, IAA may have a synergistic effect with 2-iP during the bulblet development process (Fig. 5g). We speculate that this difference arises due to the distinction between AXB outgrowth and bulblet development. AXB outgrowth is affected by apical dominance, where the accumulation of auxin within the AXB is prevented from being released to the stem, leading to failed AXB outgrowth[53,57]. In contrast, bulblet development may be independent of apical dominance, and we propose that auxin potentially plays a dual role in the process. However, further investigation is required.
The role of GAs in AXB and bulblet development is intriguing. In Rosa sp., there is a significant increase in GA biosynthesis during AXB outgrowth[61]. In Fragaria vesca, GAs plays a positive role in the outgrowth of runners (a type of AXB) without affecting their initiation[62]. In the perennial woody plant Jatropha curcas, GAs and CKs synergistically promote AXB outgrowth by inhibiting the expression of key inhibitors BRC1/2[63]. However, Zhang et al.[64] reported the GA repression of AXB formation by the modulation of DELLA-SPL9 (SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 9) complex activity. During bulblet development, GAs seemed to inhibit their formation[6,65]. In Lycoris chinensis, a high ratio of CK/GAs and ZR/GA3 favors the formation of bulblets[25,52], and exogenous GA treatment, as well as high levels of endogenous GA, appear to promote lily bulblet formation[22]. The exogenous application of the GA biosynthesis inhibitor, PBZ, is inhibited at high concentrations and promoted at low concentrations in lilies[26]. Our results indicate the upregulation of GA levels in the initiation (Supplemental Fig. S2b) and development of bulblets induced by CPPU and 2-iP (Supplemental Fig. S3b), suggesting that GA may act as a promoter in the process of bulblet formation. However, the specific regulatory mechanism of GA in this process requires further investigation.
In summary, we conducted a preliminary analysis on the effects of two plant growth regulators, CPPU and 2-iP, on the formation of lily bulblets. The results indicate their ability to promote the initiation process of bulblets through the upregulation of key gene expression and the modulation of sugar metabolism. During this process, GA3 and IAA may exhibit synergistic and antagonistic relationships with CPPU and 2-iP, respectively. Subsequently, CPPU and 2-iP significantly enhance sucrose utilization and starch accumulation, while synergistically regulating the development of bulblets along with GA3 and IAA (Fig. 8). The application of CPPU and 2-iP not only shortens the propagation cycle of bulblets but also improves their quality, highlighting the significant potential of CPPU and 2-iP for achieving high-quality and efficient production of lilies.
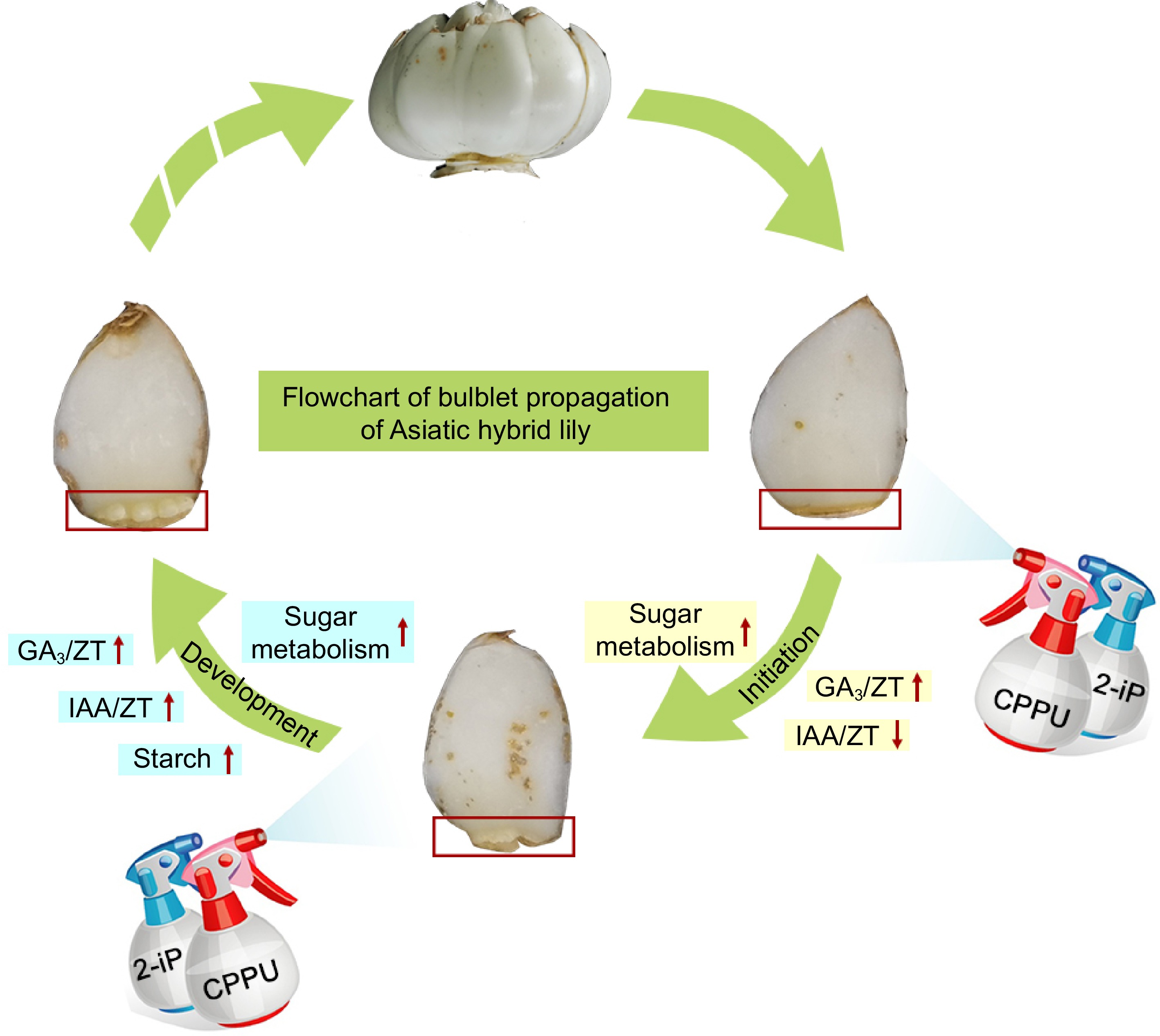
Figure 8.
Schematic model showing the promotion of bulb formation by CPPU and 2-iP in Asiatic hybrid lily.
-
The authors confirm contribution to the paper as follows: Liang J, Chen Y, Hou J, Zhang X, Wu J, Du Y conceived and designed research; Chen Y, Hou J observed the development of bulblet and performed the hormone experiments; Chen Y, Hou J, Hao J, Zuo Z, Zhang M, Liang J conducted the metabolite analysis and gene expression analysis; Liang J, Wu J, Du Y wrote the draft; Zhang X, Cao L, Wu J, Du Y reviewed the article. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This work was supported by the Excellent Youth Science Foundation of Beijing Academy of Agriculture and Forestry Sciences (YXQN202303 to Yunpeng Du), the Special projects for capacity building in scientific and technological innovation of Beijing Academy of Agriculture and Forestry (KJCX20230801 to Xiuhai Zhang), the National Natural Science Foundation of China (32171864, 32371954 to Yunpeng Du; 32172617, 32372740 to Jian Wu; 32302599 to Jiahui Liang), and the Special projects for capacity building in scientific and technological innovation of Beijing Academy of Agriculture and Forestry (KJCX20230203 to Mingfang Zhang).
-
The authors declare that they have no conflict of interest. Jian Wu is the Editorial Board member of Ornamental Plant Research who was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and the research groups.
-
# These authors contributed equally: Jiahui Liang, Yanzhu Chen, Jiaqi Hou
- Supplemental Fig. S1 Morphological and histological characteristics of bulblet formation.
- Supplemental Fig. S2 Endogenous ZT, GA3 and IAA content in the control, CPPU and 2-iP treatment at 12 days during the initiation of bulblets in lily cultivar 'Matrix'.
- Supplemental Fig. S3 Endogenous ZT, GA3 and IAA content in the control, CPPU and 2-iP treatment at BD16d during bulblet development in lily cultivar 'Matrix'.
- Supplemental File 1
- Supplemental Table S1 The primer sequences of qRT-PCR.
- Supplemental File 1 Correlation analysis.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liang J, Chen Y, Hou J, Hao J, Zuo Z, et al. 2023. Cytokinins influence bulblet formation by modulating sugar metabolism and endogenous hormones in Asiatic hybrid lily. Ornamental Plant Research 3:19 doi: 10.48130/OPR-2023-0019 

Cytokinins influence bulblet formation by modulating sugar metabolism and endogenous hormones in Asiatic hybrid lily
- Received: 24 August 2023
- Accepted: 06 November 2023
- Published online: 29 November 2023
Abstract: Lilies are bulbous flowers cherished for their aesthetic and medicinal value. Within the realm of lily cultivation, scale-cutting serves as a prevailing method for vegetative propagation, yielding diminutive bulbs termed bulblets at the base of scales. However, the burgeoning demand for lilies necessitates the optimization of propagation strategies to satisfy the surging requisites. This study centers on two synthetic cytokinin analogs, N-(2-chloro-4-pyridyl)-N'-phenylurea (CPPU) and 2-isopentenyladenine (2-iP), identified for their capacity to amplify bulblet formation within the Asiatic hybrid lily cultivar 'Matrix'. This dynamic process encompasses two pivotal phases: initiation and development. During bulblet initiation, the basal region of the maternal scale, subject to CPPU and 2-iP treatment, exhibited an elevation in the expression of cytokinin receptor genes (LaAHK2/3/4) and an augmented GA3/ZT ratio in comparison to the control, concomitant with a decrease in the IAA/ZT ratio. Moreover, CPPU and 2-iP treatments caused a notable upregulation of sucrose metabolism genes. During bulblet development stage, CPPU and 2-iP treatments invoked an augmentation in both the diameter and mass of the bulblets. The CPPU treatment accentuated the accumulation of GA3, while the 2-iP treatment effectively fostered the accrual of IAA. Additionally, both CPPU and 2-iP treatments expedited the pace of sucrose utilization and the accumulation of starch during bulblet development. This enhanced metabolic activity was conjoined by a substantial upregulation of genes linked to sucrose metabolism. Here, we underscore the proficiency of CPPU and 2-iP in elevating the formation of lily bulblets through their orchestrated impact on hormone levels and sucrose metabolism.
-
Key words:
- Lily /
- Bulblet formation /
- Cytokinins /
- Sugar /
- Hormones