









-
The natural environment exposes plants to diverse abiotic stresses, including extreme temperatures, waterlogging, and high salt concentrations, with soil salinization being the worst[1]. Soil salinization worsens due to environmental degradation, climate change, poor irrigation practices, poorly regulated fertilizer use, and industrial pollution[2,3]. Salinization affects approximately 20% of irrigated land and is projected to grow about 50% by the mid-21st century[4,5]. Salt stress severely limits plant growth, quality, and yield. Two primary strategies for mitigating soil salinity include restoring damaged soil or breeding and cultivating salt-tolerant cultivars, the latter being more secure and effective[6,7].
Rosa rugosa, a diploid species belonging to the Rosaceae family that is indigenous to East Asia, has high economic value due to its multifunctional properties. R. rugosa has the potential to be utilized for various purposes, such as fragrance extraction and ornamental, medical, and edible purposes. The essential oil extracted from R. rugosa is the most precious plant essential oil in the world, known as 'liquid gold'. R. rugosa displays superior resistance to both biotic and abiotic factors when compared to other species of Rosa. Some wild R. rugosa are distributed in coastal sandy or gravel soils and a few specific cultivars have been widely cultivated as ornamental plants in coastal cities with soils with high salt content, which indicates that there are types of cultivars with strong salt tolerance. Due to its remarkable adaptability, R. rugosa is an invasive species in northern Europe.
Salinity affects the physiological, morphological and molecular processes of plants, which eventually disrupts normal growth and metabolism[8]. The process of salt stress is characterized by two stages: osmotic stress in the early phase and ionic toxicity in the later phase[9]. Prolonged saline conditions lead to an increase in active oxygen, such as mono-oxygen, superoxides, hydroxyl radicals, and hydrogen peroxide[10]. Excessive ROS damage the plant cell membranes and DNA and affect vital biological activities, such as protein synthesis and photosynthetic pigment reduction[11]. Subsequently, plants transmit specific stress signals and adopt several adaptive strategies to adapt to saline environments. The plant response to salt stress involves multiple processes, such as ion transport, osmotic adjustment, phytohormone regulation, antioxidant regulation, and salt stress-responsive gene regulation[12].
In this study, we assessed the salt tolerance of 16 R. rugosa germplasms by examining their morphological and physiological characteristics under varying levels of salinity. Then, two cultivars, i.e., 'Zizhi' and 'Fenzizhi', were selected for further study due to their significant contrast in salt resistance and similarity in terms of morphology. To investigate the mechanisms underlying salt tolerance, we performed RNA-sequencing of roots in the 'Zizhi' and 'Fenzizhi' cultivars and analysed the DEGs related to salt stress. The aim of this study was to deepen our knowledge of the mechanisms underlying salt tolerance in R. rugosa.
-
Two-year-old R. rugosa cutting seedlings were collected from the Rosaceae Germplasm Nursery located in the Forestry Experimental Station of Shandong Agricultural University, Tai'an, China (36°10'15" N, 117°09'25" E)'. A total of 16 different germplasm-cutting seedlings were planted in a plastic container with a diameter of 260 mm and 3.4 kg of garden soil.
Containers with holes at the bottom and trays were used and placed in the Shandong Agricultural University Horticultural Greenhouse in March. After three months of growth, 1,152 seedlings from 16 germplasms ('Zizhi': ZZ; 'Fenzhongguan': FZG; 'Ciguo': CG; 'Zhongke-2': ZK-2; 'Hunchun': HC; 'Tangfen': TF; 'Tangzi': TZ; 'Xizi': XZ; 'Henan-2': HN-2; 'Baizizhi': BZZ; 'Fenzizhi': FZZ; 'Saixizi': SXZ; 'Zhulongyoukong': ZLYK; 'Henan-1': HN-1; 'Tanghong': TH; 'Zhuzishuanghui': ZZSH) were selected for further experiments. Salinity stress was induced in the 16 germplasms using NaCl solutions with varying concentrations (0.3%, 0.5%, and 0.7%). The control groups were irrigated with water only, but the stress groups were irrigated with salinated water of corresponding salt concentrations. The solution was divided equally and watered over three days to avoid a salt shock reaction. Following the achievement of the final concentration, the treatment period was recorded.
Salt injury indices
-
The morphology of plants was observed at 0, 4, 7, 10, and 16 d after stress (DAS) of 0.3%, 0.5%, and 0.7% salt treatment. The salt tolerance of R. rugosa was classified into six salt injury indices (SIIs): 0 (no injury); 1 (10% of the leaves are yellow); 2 (30% of the leaves are yellow, and leaves wilting or curling slightly); 3 (50% of leaves are yellow, and leaves wilting or curling even with a few leaves falling off); 4 (80% of leaves are scorched, large numbers of leaves fall off); and 5 (the plant is dead).
Growth and physiological indices
-
Net growth of new branch length (ΔL = L10 − L0), leaf relative water content (LRWC), and plant biomass were measured at 10 DAS in the 0, 0.3%, and 0.5% NaCl treatments. ΔL (L10 − L0) is the growth of branch length at 10 DAS.
The third and fourth leaves from the top in the control groups and the 0.3% and 0.5% NaCl-treated groups were collected at 10 DAS for physiological measurements. Each sample had three replicates. The measurement of relative conductivity (REC) was carried out[13]. The leaching method was used to measure the chlorophyll (Chl) content[14]. The anthrone method was utilized to determine soluble sugar (SS) content[15]. The Coomassie brilliant blue technique was employed to ascertain the reference soluble protein (SP) level. Malondialdehyde (MDA) was calculated using the TBA technique[16]. Nitroblue tetrazolium photoreduction was used to evaluate superoxide dismutase (SOD) activity[17]. The guaiacol technique was used to determine peroxidase (POD) activity[17].
Transcriptome library construction and Illumina sequencing
-
Two cultivars, salinity-tolerant 'Zizhi'(ZZ) and salinity-sensitive 'Fenzizhi'(FZZ), were selected for transcriptome analysis. Twelve samples of roots from the control and 0.5% NaCl-treated groups at 4 DAS were collected for extracting total RNA with the Plant Total RNA Isolation Kit (Vazyme, Nanjing, China). The quality, purity, and integrity of the RNA were then assessed. cDNA libraries were prepared, normalized, and sequenced using a NovASeq 4000 (Illumina, CA, USA). The library comprised four samples (in triplicate): Z-CK (roots control in ZZ), Z-NaCl (4d salinity stress of roots in ZZ), F-CK (roots control in FZZ), and F-NaCl (4d salinity stress of roots in FZZ).
Screening and functional analysis of DEGs
-
The acquired clean reads were aligned using HISAT2 v2.1.0 to the R. rugosa genome database (
http://eplantftp.njau.edu.cn/Rosa_rugosa/ ) to comprehend their functionalities. StringTie 1.1.3b was used to accomplish new transcript prediction. The gene expression level was assessed by quantifying the reads using the fragments per kilobase of exon model per million mapped fragments (FPKM) methodology. The differentially expressed (DEGs) were identified by setting the log2|fold changes| ≥ 2 and p-value ≤ 0.05 as criteria allowed for the examination of differential expression between samples. Gene Ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses were performed to gain further insights into the functions of DEGs. Finally, we generated a protein-protein interaction (PPI) network graph by using the STRING database (https://cn.string-db.org/cgi/input?sessionId=bnwcGhIHpZEW&input_page_show_search=on ) combined with Cytoscape software.qRT-PCR verification
-
The primers of 12 DEGs were designed and are listed in Supplemental Table S1. The qRT-PCR experiments were performed using the SYBR Green qPCR kit (Accurate Biology, China), and a CFX96 Real-time System was used for all procedures. RrGADPH served as an internal control during the 2−ΔΔCᴛ analysis of the expression patterns. Each sample was biologically replicated three times.
-
We evaluated the salt tolerance of 16 R. rugosa germplasms by observing the morphological changes under varying salt concentrations (0.3%, 0.5%, and 0.7%) and determined the salt injury index (SII: 0−6). The yellowing of leaves with increasing salt concentration was accompanied by significantly different salt tolerances among germplasms. Under 0.3% NaCl treatment, 'ZZ' and 'FZG' appeared with symptoms (SII1) at 10 DAS and reached SII2 at 16 DAS; conversely, the other 14 germplasms reached SII1 at 4 DAS. Among these 14 germplasms, eight germplasms and six germplasms reached SII3 and SII4, respectively, at 16 DAS. When exposed to 0.5% NaCl, all the germplasms exhibited symptoms (SII1−4) at 4 DAS. Eventually, 10 germplasms reached SII4, of which two cultivars, 'ZZ' and 'FZG' showed symptoms (SII2) at 10 DAS, 3 d later than the other eight germplasms. Of the remaining six germplasms, three germplasms, 'ZZSH', 'ZLYK', and 'HN-1' reached SII5 at 7 DAS and were more sensitive to salt treatment. Similar to the 0.5% NaCl treatment, all R. rugosa germplasms exhibited yellow leaves (SII1−4) at 4 DAS under the 0.7% NaCl treatment. 'ZZ' and 'FZG' reached SII4 at 16 DAS, whereas the other germplasms achieved SII5 at either 7 DAS or 16 DAS (Supplemental Table S2). Through cluster analysis of SII, 16 R. rugosa germplasms were categorized into two distinct groups based on salt tolerance. The salt-tolerant group consisted of 10 germplasms that could be further divided into two subgroups. 'ZZ' and 'FZG' showed greater salt resistance than the remaining eight germplasms. The salt-sensitive group was composed of six germplasms, of which 'ZLYK', 'HN-1', 'TH', and 'ZZSH' were more sensitive (Fig. 1a).
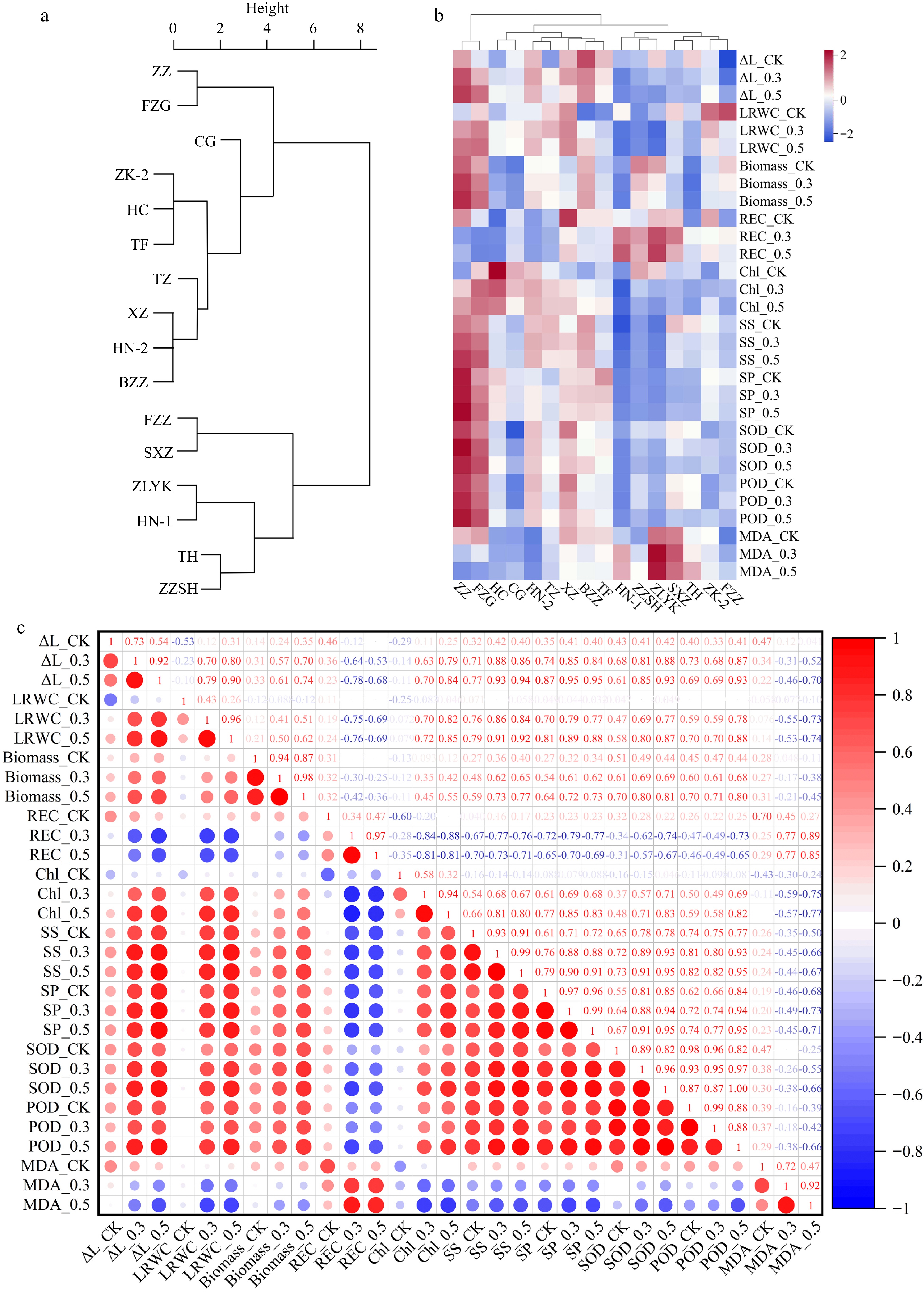
Figure 1.
Analysis of salt tolerance of 16 R. rugosa germplasms. (a) Cluster analysis of salt tolerance in R. rugosa according to salt injury index. (b) Clustering heatmap of growth and physiological indicators. (c) Correlation analysis of growth and physiological indicators.
Growth and physiological responses to salt stress
-
To make a more comprehensive evaluation of salt tolerance in 16 R. rugosa germplasms, three growth indicators and seven physiological indicators were measured at 10 DAS under 0%, 0.3%, and 0.5% NaCl treatments. As the NaCl concentration increased, a significant decrease in the three growth indicators (ΔL, LRWC, and plant biomass) and a significant increase in the physiological indicators (REC, SS, SP, MDA, SOD, and POD) were observed, except for Chl content, which displayed a decreasing trend (Supplemental Table S3).
Heatmap clustering was performed using ten indicators to evaluate the salt tolerance of sixteen germplasms (Fig. 1b). These germplasms were categorized into three distinct clusters, namely the salt-tolerant type ('ZZ' and 'FZG'), middle salt-tolerant type including seven germplasms ('HC', 'CG', 'HN-2', 'TZ', 'XZ', 'BZZ' and 'TF'), and salt-sensitive type including seven cultivars ('NH-1', 'ZZSH', 'ZLYK', 'SXZ', 'TH', 'ZK-2' and 'FZZ'). This finding is analogous to the clustering results of Fig. 1a. The strong salt tolerance of the R. rugosa germplasms showed that ΔL, LRWC, plant biomass, Chl content, REC, and MDA underwent small fluctuations, while SS, SP, SOD, and POD showed significant changes compared to the control group. The 'ZZ' and 'FZG' cultivars exhibited the strongest salt tolerance. These observations clearly demonstrate that the salt tolerance of R. rugosa germplasms varies significantly.
The findings depicted in Fig. 1a & b were consistent with the results of principal component analysis (PCA). The explained variances of PC1 and PC2 are 52.31% and 13.04%, respectively (Supplemental Fig. S1). To determine the practical physiological parameters that could pinpoint R. rugosa salt tolerance, a correlation study was carried out. All of the indicators of treatment with 0.3% and 0.5% NaCl showed a high positive connection, with Pearson correlation coefficients ranging from 0.88 to 0.99 (Fig. 1c). Hence, measuring only one concentration of the indicator in further studies would suffice. Additionally, REC and MDA exhibited inverse correlations with the rest of the indicators, whereas other indices were positively correlated among themselves.
Identification of differentially expressed genes by transcriptome sequencing
-
Transcriptome sequencing was carried out to investigate the mechanical response to salinity stress using root samples of two cultivars (salinity-tolerant 'ZZ' and salinity-sensitive 'FZZ') at 0 and 4 DAS after 0.5% NaCl treatment. The raw data are available at the NCBI Sequence Read Archive with the accession numbers SRR25020864~SRR25020875. Twelve RNA libraries were generated from the RNA sequencing data, with Q20 and Q30 values greater than 98.11% and 94.31%, respectively. The average GC content was approximately 45.14% (Supplemental Table S4). Four comparison groups were constructed, namely, Z-NaCl vs Z-CK, F-NaCl vs F-CK, F-CK vs Z-CK, and F-NaCl vs Z-NaCl, resulting in 7,806 DEGs found in at least one group. Among these groups, 795, 2,207, 4,673, and 4,914 DEGs were identified, respectively, and among them, most genes were downregulated (Fig. 2a). There were 340 genes identified to exhibit differential expression in both cultivars, while 3,129 DEGs were discovered in the comparison of different cultivars. Moreover, only 43 genes had different expression patterns in all four groups (Fig. 2b).
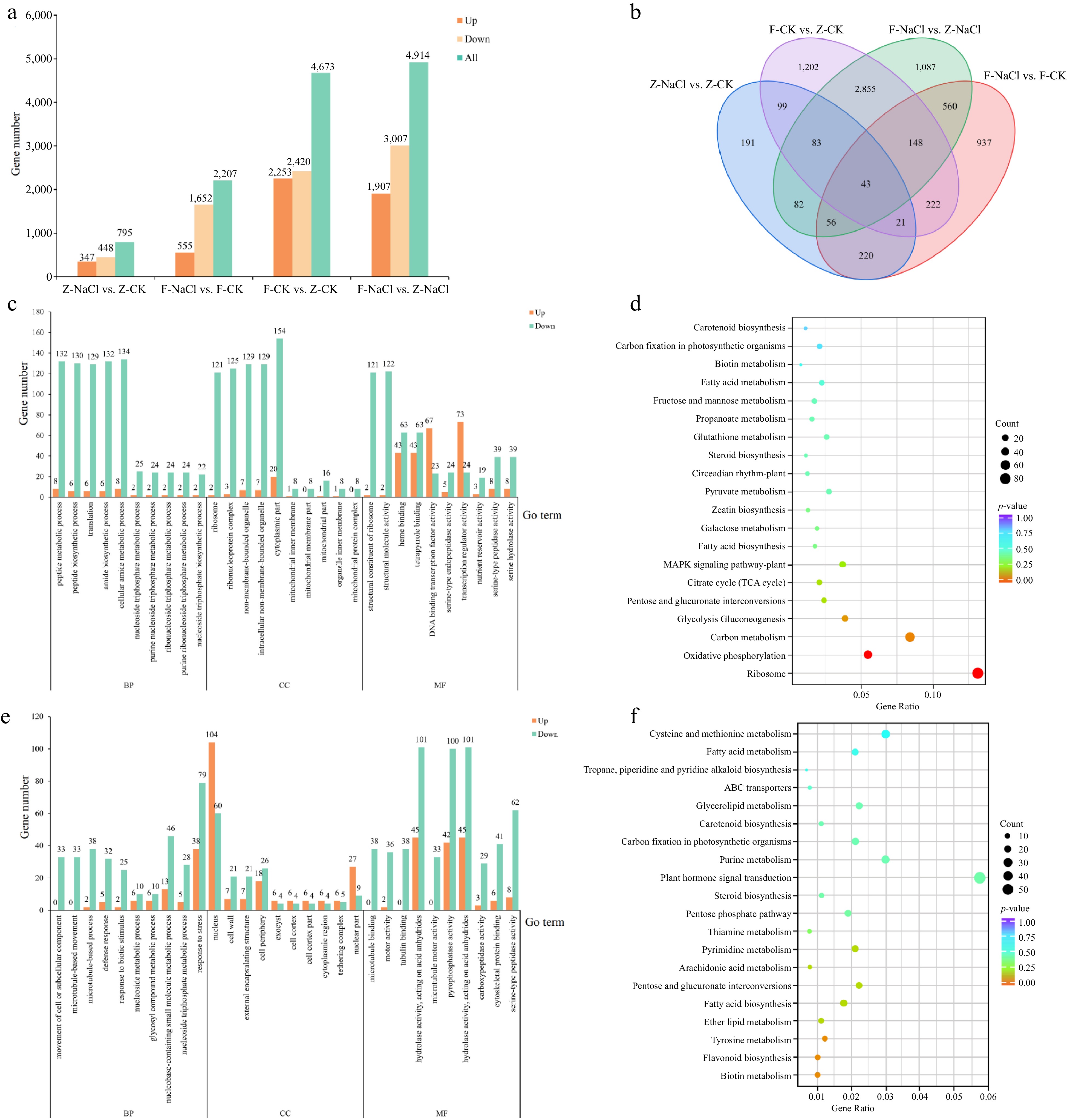
Figure 2.
DEG analysis of in the four comparison groups. (a) Total number of DEGs in Groups Z-NaCl vs Z-CK, F-NaCl vs F-CK, F-CK vs Z-CK, and F-NaCl vs Z-NaCl. (b) Venn diagram of DEGs in Z-NaCl vs Z-CK, F-NaCl vs F-CK, F-CK vs Z-CK, and F-NaCl vs Z-NaCl. (c) GO enrichment analysis of Z-NaCl vs Z-CK, and (d) F-NaCl vs F-CK. (e) KEGG enrichment analysis of Z-NaCl vs Z-CK and (f) F-NaCl vs F-CK.
GO and KEGG enrichment analysis
-
The DEGs of the Z-NaCl vs Z-CK group were categorized according to the GO database into biological processes (BP, 434), molecular functions (MF, 291), and cellular components (CC, 107) (Supplemental Table S5). On the other hand, the DEGs of F-NaCl vs F-CK were also categorized according to the GO database into BP (477), MF (302), and CC (117) (Supplemental Table S6). To further investigate the significance relevance of the salt stress-responsive DEGs, we analysed the top 10 terms that were enriched among the three groups (Fig. 2c & d). DEGs in ZZ and FZZ were mapped into 106 and 104 KEGG pathways, respectively (Supplemental Tables S7 & S8). Only two metabolic pathways (Ribosome, Ko03010, 81 genes; Oxidative phosphorylation, Ko00190, 34 genes) were significantly enriched (p-value < 0.05). There was no significantly enriched pathway in FZZ, while ribosome (Ko03010, 56 genes), carbon metabolism (Ko01200, 56 genes), and plant hormone signal transduction (Ko04075, 52) had the largest number of genes (Fig. 2e & f).
DEGs implicated in the biosynthesis, transport, and signalling of phytohormones
-
Genes involved in phytohormone biosynthesis and signal transduction showed different expression levels under salt stress in the two cultivars (Fig. 3). Notably, there were significant downregulation trends of 26 positive regulatory DEGs associated with auxin biosynthesis, transport and signal transduction in one or both cultivars, including YUC (evm.model.Chr6.539; evm.model.Chr2.1946), PIN (evm.model.Chr7.3286; evm.model.Chr7.3273; evm.model.Chr7.3546; evm.model.Chr6.1170), genes coding auxin transporter-like protein (evm.model.Chr2.6044) and auxin-induced protein (evm.model.Chr2.5904; evm.model.Chr4.2260), auxin efflux carrier component (evm.model.Chr1.4080; evm.model.Chr4.72; evm.model.Chr6.6477; evm.model.Chr6.4089), IAA14 (evm.model.Chr1.692), ARF (evm.model.Chr5.2402), GH3.17 (evm.model.Chr1.2987), ARG7 (evm.model.Chr6.4936), and eight SAUR (evm.model. Chr6.4937; evm.model.Chr2.4303; evm.model.Chr2.4302; evm.model.Chr2.920; evm.model.Chr4.2643; evm.model.Chr2.664; evm.model.Chr5.5043; evm.model. Chr7.1017). Furthermore, one auxin-repressed gene, ARP (evm.model. Chr5.6322) was upregulated under NaCl treatment in both cultivars.
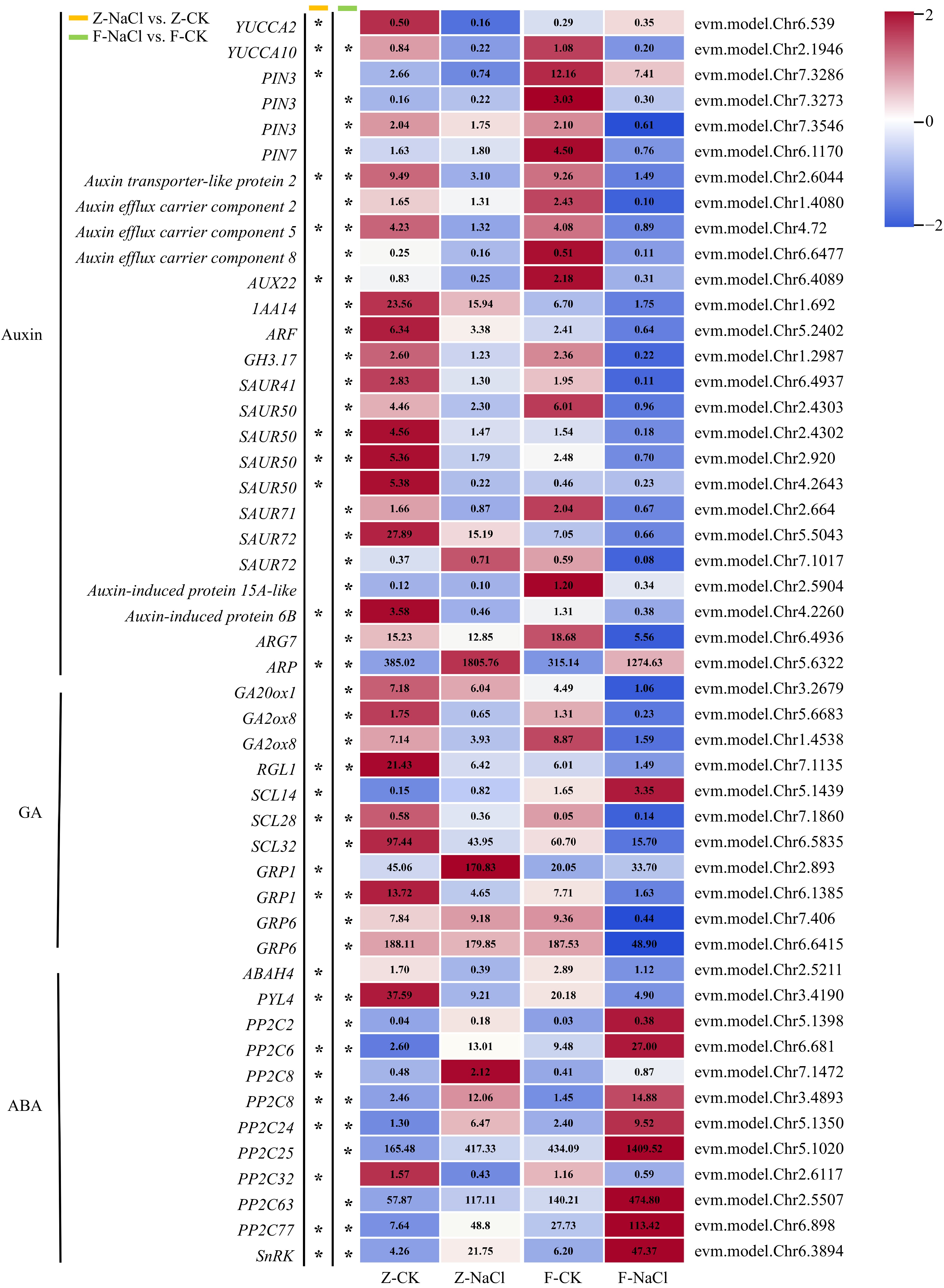
Figure 3.
Heatmap of DEGs associated with phytohormones (* log2| fold changes | ≥ 2, p-value < 0.05). ABAH: abscisic acid 8'-hydroxylase; ARF: Auxin response factor; ARG: Auxin-related gene; ARP: auxin-repressed protein; AUX: AUXIN; GA20ox: Gibberellin 20-oxidase; GA2ox: Gibberellin 2-oxidase; GH3: Gretchen Hagen 3; GRP: Gibberellin-regulated protein; IAA: Indole-3-acetic acid; PIN: PIN-formed; PP2C: Protein phosphatase 2C; PYL: Pyrabactin resistance 1-like; RGL1: RGA-like 1; SAUR: Small auxin-up RNA; SCL: Scarcrow-like; SnRK: SNF1-related protein kinase regulatory subunit gamma-like; YUC: YUCCA.
Eleven genes coding gibberellin-regulated proteins were identified as DEGs, including GA20ox (evm.model.Chr3.2679), GA2ox (evm.model.Chr5.6683; evm.model.Chr1.4538), RGL1 (evm.model.Chr7.1135), SCL (evm.model.Chr5.1439; evm.model.Chr7.1860; evm.model.Chr6.5835), and GRP (evm.model.Chr2.893; evm.model.Chr6.1385; evm.model.Chr7.406; evm.model.Chr6.6415). Similar to DEGs involved in auxin, the expression patterns of gibberellin-related DEGs were downregulated under salt stress, except for one member of GRP1 (evm.model.Chr2.893), which was upregulated after salt treatment.
Twelve genes were identified as DEGs in the abscisic acid regulation pathway. Specifically, the study found that ABAH4 (evm.model.Chr2.5211) and PYL4 (evm.model.Chr3.4190) exhibited downregulation. Moreover, nine members of PP2C (evm.model.Chr5.1398; evm.model.Chr6.681; evm.model.Chr7.1472; evm.model.Chr3.4893; evm.model.Chr5.1350; evm.model.Chr5.1020; evm.model.Chr2.6117; evm.model.Chr2.5507; evm.model.Chr6.898) and one SnRK (evm.model.Chr6.3894) gene were upregulated under salt stress, whereas only one member of PP2C (evm.model.Chr2.6117) was downregulated.
Gene expression profile associated with salinity tolerance
-
To explore the mechanism behind salt tolerance in R. rugosa, we examined DEGs of potential salinity tolerance genes in both the of F-NaCl vs F-CK and Z-NaCl vs Z-CK groups. The functional annotation showed that many DEGs may be related to salt stress and they showed interesting patterns.
In our analysis, we detected 14 DEGs related to ROS production, including eight peroxidase-encoding genes, one peroxygenase-encoding gene, three iron transporter genes, one ferritin gene, and one patatin-like protein-encoding gene. Among them, POD (evm.model.Chr6.2566; evm.model.Chr2.1569; evm.model.Chr6.5801; evm.model. Chr3.5028; evm.model.Chr1.2412; evm.model.Chr5.860; evm.model.Chr6.3517) were downregulated. PXG2 (evm.model.Chr1.280) and POD4 (evm.model.Chr5.858) were upregulated in NaCl vs CK. Additionally, PLP2 (evm.model.Chr2.486), with antioxidant activity was upregulated. Ferritin (evm.model.Chr4.1192) could maintain the dynamic balance of ROS in plants and its expression was greatly increased after treatment. In contrast, three VITs (evm.model.Chr1.4206; evm.model.Chr1.4205; evm.model.Chr6.1138) were correspondingly reduced (Fig. 4).
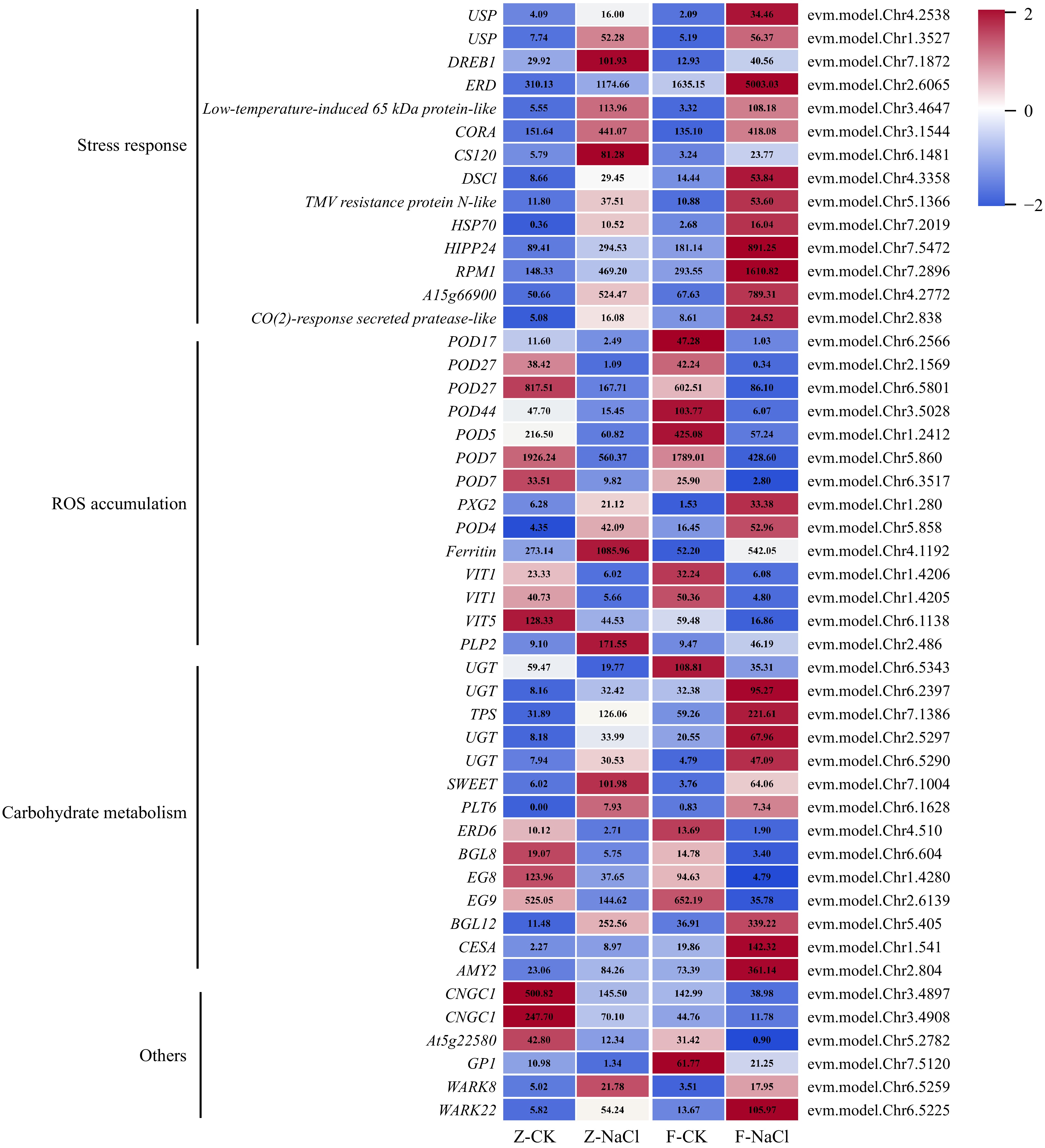
Figure 4.
Expression profile of DEGs associated with salt stress. AMY: Alpha-amylase; BGL: Beta-glucosidase; CESA: Cellulose synthase-like protein; CNGC: Cyclic nucleotide-gated ion channel 1-like; CS: Cold-shock protein; CORA: Cold-regulated protein; DREB: Dehydration-responsive element-binding protein; DSC: Disease resistance-like protein; EG: Endoglucanase; ERD: Early response to dehydration 6-like; ERD: Early-responsive to dehydration; GP1: Vegetative cell wall protein gp1-like; HIPP: Heavy metal-associated isoprenylated plant protein; HSP: Heat shock protein; PLP: Patatin-like protein; PLT: Polyol transporter; POD: Peroxidase; PXG: Peroxygenase; RPM: Resistance to pseudomonas maculicola; SWEET: Sugars will eventually be exported transporter; TPS: Trehalose-6-phosphate synthase; UGT: UDP-glucosyltransferase family protein; USP: universal stress protein; VIT: Vacuolar iron transporter 1-like; WARK: Wall-associated receptor kinase-like.
Soluble sugars can be used as osmotic regulator to maintain osmotic stability when subjected to salt stress. Four UGTs (evm.model.Chr6.5343; evm.model.Chr6.2397; evm.model.Chr7.1386; evm.model.Chr6.5290) exhibited distinct expression patterns, of which three genes (evm.model.Chr6.2397; evm.model.Chr7.1386; evm.model. Chr6.5290) were expressed at higher levels under NaCl treatment. Similarly, BGL12 (evm.model.Chr5.405), CESA (evm.model.Chr1.541), SWEET (evm.model.Chr7.1004), PLT6 (evm.model.Chr6.1628), and AMY2 (evm.model.Chr2.804) were also upregulated. However, the TPS-encoding gene (evm.model.Chr7.1386), EG8/9 (evm.model.Chr1.4280 and evm.model.Chr2.6139), ERD6 (evm.model.Chr4.510), and BGL8 were downregulated (Fig. 4).
An interesting discovery was that various stress response and resistance protein coding genes had higher expression patterns after salt stress, including USP (evm.model.Chr4.2538 and evm.model.Chr1.3527), DREB1 (evm.model.Chr7.1872), low-temperature-induced protein (evm.model.Chr3.4647), CORA (evm.model.Chr 3.1544), CS120 (evm.model.Chr6.1481), disease resistance protein (evm.model.Chr4.3358; evm.model.Chr7.2896; evm.model.Chr4.2772), TMV resistance protein (evm. model.Chr5.1366), HSP70 (evm.model.Chr7.2019), HIPP24 (evm.model.Chr7.5472) and CO(2)-response secreted protease (evm.model.Chr2.838) (Fig. 4). This finding shows that plants exhibit similar responses to different types of stressors.
Additionally, we identified additional genes that could contribute to salt tolerance. Notably, CNGC1 (evm.model.Chr3.4897 and evm.model.Chr3.4908), one stress-response A/B barrel domain-containing protein At5g22580 (evm.model.Chr5.2782) and GP1 (evm.model.Chr7.5120) were downregulated under salt treatment. Equally noteworthy, two WARK (evm.model.Chr6.5259 and evm.model.Chr6.5225) was upregulated under salt stress (Fig. 4).
Differential expression analysis of TFs
-
A total of 158 TFs were identified, and they mainly belonged to the zinc finger protein, AP2, MYB, NAM, bHLH, etc., families (Fig. 5a). Of these TFs, 46 and 125 displayed differential expression in ZZ and FZZ, respectively. In ZZ, TFs mainly belonged to the AP2, MYB, zinc finger protein, NAM, and WD40 families, while in FZZ, they belonged to the AP2, zinc finger protein, MYB, NAM, and bHLH families (Fig. 5b & c). The AP2 family had the largest number of TFs in both ZZ and FZZ. As these families are important in plant stress responses, clustering heatmaps of TFs in MYB, NAM, and bHLH were drawn. Specifically, nine, five, and seven DEGs showed downexpression patterns after salt treatment in MYB, NAM, and bHLH, respectively. Nine, nine, and three DEGs were upregulated in the three families (Fig. 5d−f).
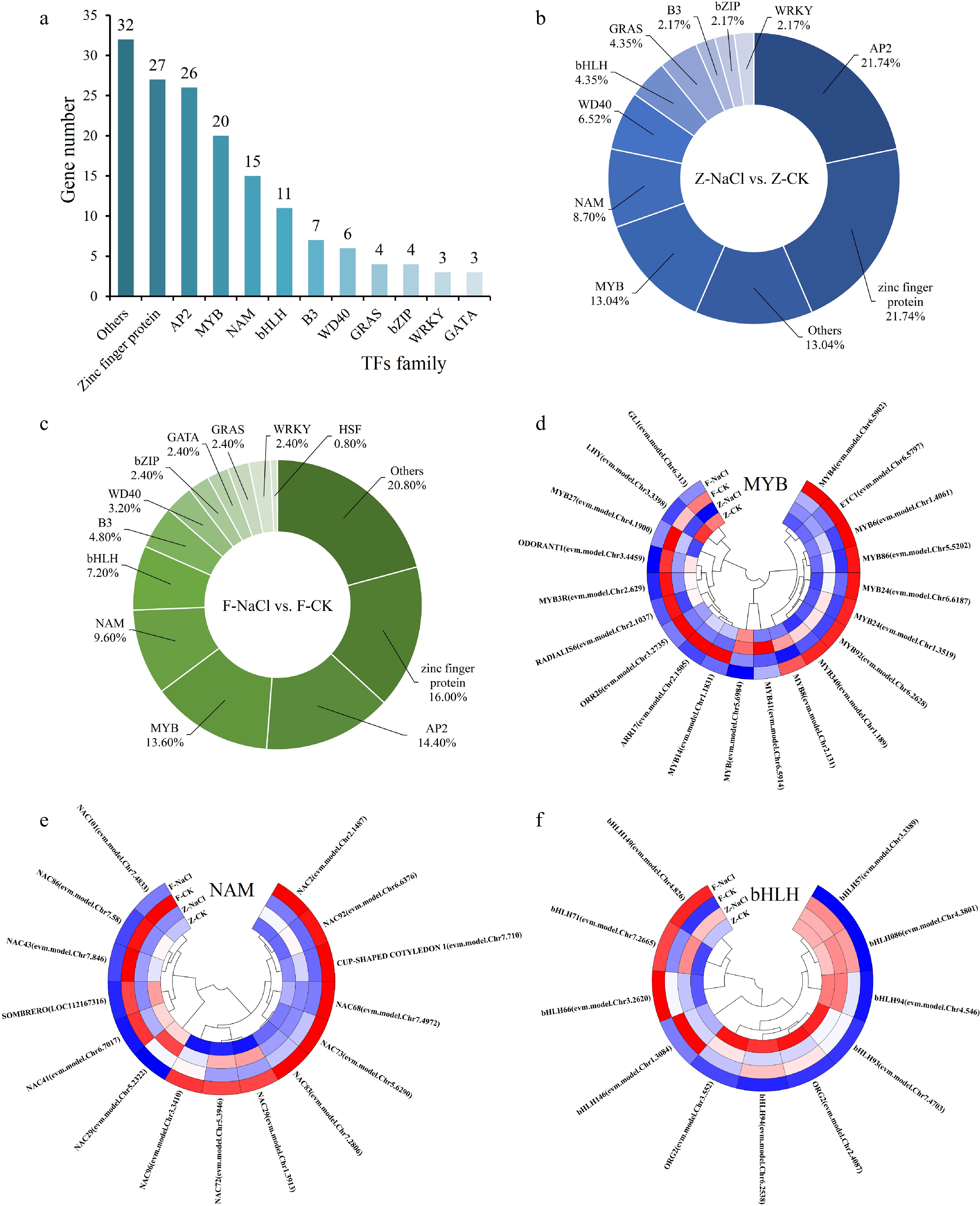
Figure 5.
Transcription factor statistical analysis. (a) Statistics on the number of TFs. (b)−(c) The percentage of TFs in Z-NaCl vs Z-CK, F-NaCl vs F-CK. (d)−(f) Clustering heatmaps of TFs in MYB, NAM, and bHLH.
PPI network of DEGs
-
To predict the potential functions and relationships, the PPI network of DEGs was utilized (Fig. 6). SCL28 and E2F1 were core regulators, as they had the most interactions with other genes. SCL28 interacted with SCL14, SR45a, and HSP70 and was further associated with ERF13 and ERF4 through HSP70. E2F1 interacted directly with MYB6, GTE3, MCM3, and MCM2 and was further related to WRKY45 and FY.
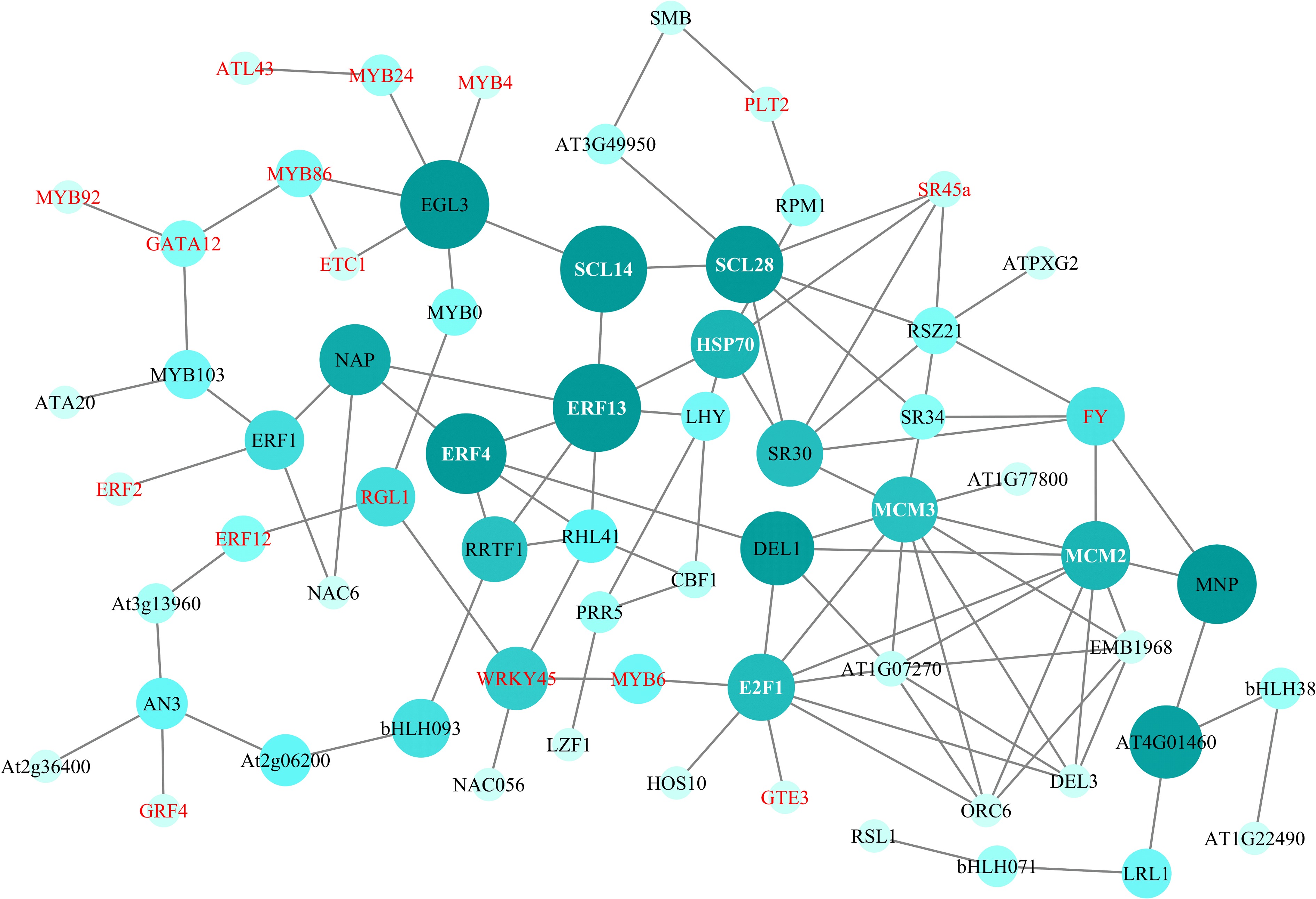
Figure 6.
PPI network of DEGs. Genes in red and white font are DEGs in the transcriptome data of R. rugosa.
qRT-PCR validation of DEGs
-
We confirmed the RNA-seq results through qRT-PCR. Therefore, we screened 12 DEGs, i.e., auxin-related DEGs (YUCCA10, SAUR50 and ARP), ABA-related DEGs (SnRK), stress response-related DEGs (USP and HSP70), DEGs related to ROS accumulation (POD27 and VIT1), DEGs related to sugar metabolism (UGT and TPS), four TFs (E2F1, SCL28, WRKY45, MYB92), and two other DEGs (CNGC1 and WARK22) (Fig. 7). Our findings consistently demonstrated the similarity of expression patterns between qRT-PCR and transcriptome data for these genes.
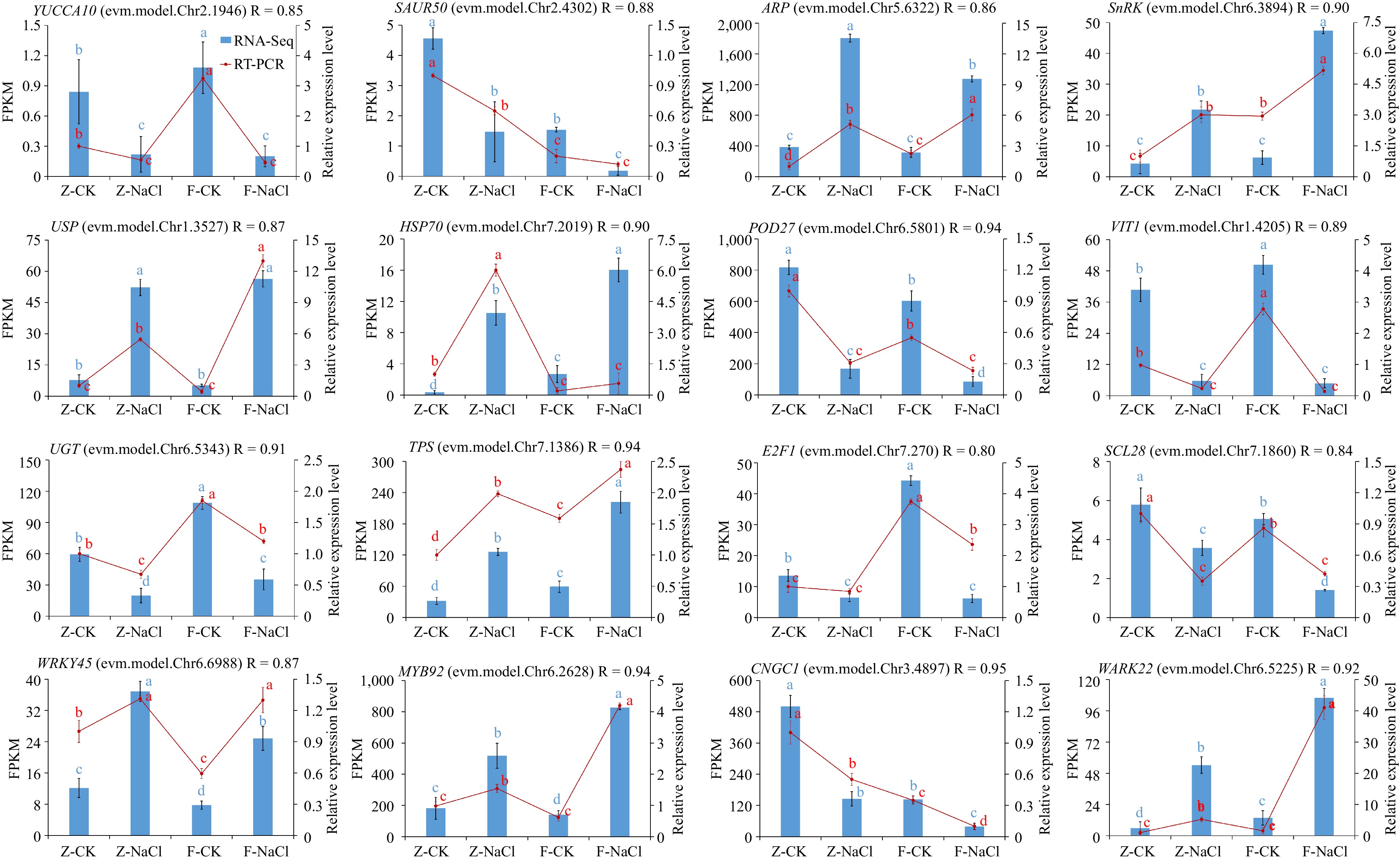
Figure 7.
Quantitative RT-PCR validation of 16 DEGs. Relative expression levels from quantitative RT-PCR and FPKM from the transcriptome of genes are shown on the left and right, respectively. The red line represents the FPKM value in the RNA-seq result. The blue bar represents qRT-PCR results. The different small letter for each expression level indicated significant difference between germplasms at α = 0.05.
-
Salt stress causes considerable damage to plant morphology, physiology, and growth. Screening salt-tolerant plant materials can be achieved via morphological characteristics, biomass, and physiological indices[18]. By observing the morphology of leaf damage in R. rugosa germplasms after salt stress treatment, salt injury was classified into six indices. Two distinct types, namely, salt-tolerant and salt-sensitive, were identified through cluster analysis. Salt stress severely inhibited plant growth, leading to reduced growth indicators. The impact on physiology was reflected in multiple factors, including soluble sugars and proteins, which promote cells to cope with osmotic stresses under salt stress conditions[19]. Salt stress affects plant photosynthesis, which is reflected in the chlorophyll content[20]. Overproduction of ROS induced by salt stress damages plant cells by destroying nucleic acids, lipids, and proteins[21]. ROS synthesis results in the accumulation of MDA, SOD, and POD, which are antioxidant enzymes that play significant defensive roles[22]. Seven physiological indicators were measured in this study, including REC, SS, SP, MDA, SOD, and POD. The physiological indicators significantly increased, except Chl, which showed a decreasing trend. Although the trends of growth and physiological changes were consistent across the six germplasms, the degree varied. The variation ranges of ZZ and FZG were evidently smaller or larger than those of other germplasms, indicating stronger salt resistance in ZZ and FZG. Heatmap clustering and principal component analysis confirmed this conclusion. The 16 germplasms were divided into three clusters: salt-tolerant type ('ZZ' and 'FZG'), middle salt-tolerant type ('HC', 'CG', 'HN-2', 'TZ', 'XZ', 'BZZ' and 'TF'), and salt-sensitive type ('NH-1', 'ZZSH', 'ZLYK', 'SXZ', 'TH', 'ZK-2' and 'FZZ').
In this experiment, roots of 'Zizhi' and 'Fenzizhi' at 0 and 4 DAS were collected for transcriptome sequencing. When comparing the salinity stress with the control, the number of downregulated DEGs was more frequent than the number of upregulated DEGs in both cultivars, indicating substantial inhibition of gene expression by salt stress. The comparison between F-NaCl and F-CK revealed a larger number of DEGs compared to Z-NaCl vs Z-CK, particularly the downregulated genes, suggesting a greater impact of NaCl in FZZ than in ZZ.
Phytohormones are essential small chemicals. Relevant research indicates that they play complex and efficient roles under varying environmental conditions[23]. The precise role of IAA metabolism in the response to salt stress is not yet fully understood. Salt stress led to a decrease in both auxin content and the expression level of auxin transporters, given that auxin is a significant growth-promoting hormone[24]. Auxin positive regulators including YUCCA, PIN, genes coding auxin transporter-like protein and auxin-induced protein, IAA, ARF, GH3, SAUR, and auxin-induced protein ARG7-like, were downregulated under salt stress in one or both R. rugosa cultivars. The YUC family is essential in auxin biosynthesis by regulating the pathway for producing indole-3-pyruvic acid (IPyA)[25]. Overexpression of YUC leads to higher capacities for maintaining low ROS levels and confers resistance to salt stress[26]. Auxin influx and efflux are severely impaired under salt stress, subsequently affecting auxin signalling. Salt stress leads to the downregulation of PIN proteins, which are essential for regulating auxin transportation[27]. Auxin signalling pathway proteins Aux/IAA were found to be downregulated due to Nitric Oxide (NO) accumulation caused by salt stress[28]. The interaction between the Aux/IAA proteins and ARFs enables the salt stress response by regulating cis-elements and posttranscriptional processes[27]. The GH3 family of enzymes conjugates IAA with amides. Interestingly, the research found that GH3 genes are induced under salt stress[29], which was different from our result. Research indicates that the SAUR gene positively influences plant root system architecture and enhances abiotic tolerance[30]. Additionally, in the context of salt stress, only one ARP gene displayed upregulation. Similarly, an investigation on Capsicum annuum revealed the induction of ARP in the presence of salt stress[31].
Five genes coding gibberellin-regulated proteins were differentially expressed under salt stress. GA20ox, GA3ox, and GA2ox are essential for the synthesis pathway and inactivation of biologically active GAs[32]. GA2oxs can enhance salt tolerance by retarding plant growth[33]. Our research revealed significant downregulation of GA20ox and GA2ox genes in R. rugosa. Reports have indicated that the overexpression of SCL can enhance salt tolerance[34]. We identified three SCL genes, of which one was induced by salt treatment, and the other two were inhibited. Both RGL and GRP proteins belong to the DELLA family. The overexpression of RGL3 can mitigate the inhibitory impact of NO deficiency on salt tolerance[28]. Moreover, the GRP protein is known to be a gibberellin repressor and has been identified as an allergen in fruits[35]. The concentration of GRP protein was influenced by salt stress[36]. According to transcriptome analysis, we found that most RGL and GRP genes were inhibited under salt stress in R. rugosa.
Salt stress induces ABA synthesis, which is the most important stress response hormone. ABAH is an essential enzyme involved in the oxidative degradation of ABA, thus negatively regulating its accumulation[37]. Salt stress was found to suppress the expression of ABAH4 in R. rugosa. Moreover, ABA signalling pathway activation results in an elevation of ABA levels[38]. In the ABA signalling pathway, PYR/PYL/RCARs receptors sense ABA, resulting in the inhibition of PP2C activity, which activates SnRK2[39]. Our transcriptome analysis showed that there were 11 ABA signalling-related DEGs, including PYL4, SnRK, and nine PP2Cs. The expression profile of SnRK in R. rugosa under salt stress conditions aligns with previous findings and acts as the most crucial node of ABA signalling[40]. Among the nine PP2Cs, only one member showed lower expression under salt treatment, possibly acting as a SnRK inhibitor.
Certain proteins exhibit responses to multiple stresses, as many features are shared across these stresses. For instance, DREB can be induced by various stressors, including both abiotic and biotic factors[41]. Moreover, pretreatment with salt could prime a plant's response to infection by biotic pathogens[42]. Salinity causes substantial stress by leading to osmotic stress and ionic toxicity, which impacts various biochemical processes and may even result in cell death. In our current research, we found that the expression of multiple stress response protein-coding genes was changed significantly by salt stress in R. rugosa, including genes coding USP, DREB1, CORA1, CS120 and so on. This observation further supports the idea that the response and adaptation mechanisms of plants to various stresses share many common elements.
As previously mentioned, excessive ROS can disrupt the permeability of the plasma membrane and damage DNA and proteins. Plant antioxidant systems consist of various enzymes, such as POD and SOD. Additionally, glutathione-S-transferase (GST) is also included in these systems[43,44]. Fourteen DEGs related to ROS production were identified, with eight POD genes being the largest group. Seven of these POD genes were downregulated, with a greater reduction in FZZ compared to ZZ. Hence, POD may serve as a pivotal regulatory gene in antioxidant system of R. rugosa, controlling its response to salt stress. In the present experiment, PXG2 and ferritin were upregulated under salt stress, which is consistent with findings in Avena chinensis and Oryza sativa[45,46]. This suggests that these two genes may play roles in the salt stress response in R. rugosa.
Soluble sugars fulfil a dual role by serving as an energy source for cellular activities and as regulators of intracellular osmotic pressure[47]. Fourteen DEGs (TPS, SWEET, PLT6, and ERD6) associated with carbohydrate metabolism and transportation were detected, among which nine genes were upregulated, which was consistent with soluble sugar content. Four UGT coding genes exhibited differential expression patterns, and three of them were upregulated under NaCl treatment. Research has demonstrated the positive impact of UGT on enhancing plant salt tolerance by promoting flavonoid accumulation[48]. In addition, salt stress has also been shown to activate the expression of TPS and sugar transporter genes (SWEET and PLT6)[49−51]. Sugar metabolism plays an important role in the response to salt stress. Five genes showed downregulation, with the expression levels of EG8 and EG9 exhibiting a significant decrease in FZZ compared to ZZ.
The PPI network indicated that the core regulators SCL28 and E2F1 had the most interactions with multiple genes. SCL transcription factors are members of the GRAS protein family and are crucial for plant stress resistance. Cruz et al. found the involvement of SCL28 in stress responses induced by ABA[52]. However, limited findings have been reported regarding their involvement in the plant's salt stress response mechanism. SCL13 can improve plant growth and increase salt tolerance when overexpressed in both Arabidopsis thaliana and Tamarix hispida[53,54]. E2F transcription factors interact with MYB-type transcription factors under salt stress conditions to coordinate cell cycles and regulate normal plant growth[55]. Previous findings suggest that these genes may interact to regulate the salt stress response mechanism in R. rugosa. However, further verification is necessary to clarify the molecular mechanism underlying these genes.
-
The authors confirm contribution to the paper as follows: research conception and design: Yu Y, Zhao F; analysis and interpretation of results: Qi S, Wang X, Li X; draft manuscript preparation: Qi S, Wang X; feedback on the analysis and manuscript: Yu Y, Zhao F, Wu Q, Xing S. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This project was funded by the Shandong Agricultural Seeds Engineering Project (2020LZGC011) and Shandong Provincial Natural Science Foundation (ZR2023MC015).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Shuai Qi, Xiaobin Wang
- Supplemental Table S1 The list of qRT-PCR primer sequences.
- Supplemental Table S2 Salt injury index of 16 R. rugosa cultivars.
- Supplemental Table S3 The data of growth and physiological indicators determination on 10 DAS of 0, 0.3%, and 0.5% NaCl treatment.
- Supplemental Table S4 Sequencing data for 12 libraries obtained by RNA sequencing.
- Supplemental Table S5 The data of GO enrichment analysis in Z-NaCl vs. Z-CK.
- Supplemental Table S6 The data of GO enrichment analysis in F-NaCl vs. F-CK.
- Supplemental Table S7 The data of KEGG enrichment analysis in Z-NaCl vs. Z-CK.
- Supplemental Table S8 The data of KEGG enrichment analysis in F-NaCl vs. F-CK.
- Supplemental Fig. S1 Principal component analysis with two factors.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Qi S, Wang X, Wu Q, Xing S, Li X, et al. 2023. Morphological, physiological and transcriptomic analyses reveal potential candidate genes responsible for salt stress in Rosa rugosa. Ornamental Plant Research 3:21 doi: 10.48130/OPR-2023-0021 

Morphological, physiological and transcriptomic analyses reveal potential candidate genes responsible for salt stress in Rosa rugosa
- Received: 01 August 2023
- Accepted: 24 November 2023
- Published online: 15 December 2023
Abstract: Rosa rugosa is a multifunctional species with various applications in fragrance extraction and ornamental, medical, and edible purposes. Compared to other species of Rosa, R. rugosa exhibits greater resistance to salt stresses; however, the mechanisms underlying its salinity tolerance are still unclear. Thus, we assessed the salt tolerance of 16 R. rugosa germplasms based on the changes in morphology and physiology under different salt concentrations. Among them, two cultivars ('Zizhi' and 'Fenzizhi') were chosen for transcriptome sequencing. The differentially expressed genes (DEGs) associated with phytohormone synthesis and signalling pathways, ROS production, carbohydrate metabolism and transport, stress response and resistance were identified as responding to salt stress in R. rugosa. PPI network analysis further identified two DEGs, SCL28 and E2F1, which exhibited the most interactions with other DEGs. Consequently, this research contributes to a greater understanding of salinity tolerance mechanism in R. rugosa and could provide gene resources for breeding salt-tolerant cultivars in Rosa.
-
Key words:
- Rosa rugosa /
- Salt stress /
- Morphology /
- Physiology /
- Transcriptome sequencing