









-
Carotenoids, a group of widely distributed natural pigments, influence the color development of petals in many plants[1,2]. The final accumulation of carotenoids, determined by a series of biosynthetic genes and degradation genes, has a great influence on petal coloration[3]. Since the C-C double bond on the carotenoid skeleton is vulnerable to oxygen attack, non-enzymatic or enzymatic oxidative cleavage occurs, resulting in various forms of carotenoid derivatives produced by the enzymatic cleavage of the carotenoid cleavage oxygenases (CCO) family. CCO family composed of multiple members in plants can be divided into two main categories: carotenoid cleavage dioxygenase (CCD) and 9-cis-epoxy carotenoid dioxygenase (NCED)[4].
Among different CCD gene members, CCD1 and CCD4 participating in the degradation of carotenoids, are regarded as the key factor to regulate carotenoid accumulation in several plant species[5−7]. In plants, the CCD1 enzyme only exists in the cytoplasm due to the lack of plastid localization peptides. Since carotenoids are mainly produced in the plastids, CCD1 cannot directly act on the C40 carotenoid substrates in the plastids[8,9]. Only C27 apocarotenoids, the product of C40 carotenoids cleaved by CCD4, can be further cleaved by CCD1 into C13 and/or C14 small-molecule volatiles, such as α-ionone and β-ionone[10−12]. But in the bacterial pigment complementation system, when the CCD1 prokaryotic expression vector and the engineering plasmid capable of producing carotenoids are expressed in Escherichia coli, CCD1 can contact and cleave several C40 carotenoids, including lycopene, α-carotene, β-carotene, ε-carotene and zeaxanthin[13,14]. Based on these results, it is presumed that CCD1 can directly act on both apocarotenoids produced by other CCD proteins and C40 carotenoids under certain conditions.
In contrast, the plastid localization of CCD4 allows access to carotenoid substrates that were synthesized and stored in plastids[15]. CCD4 cleaves the same position (9, 10 and 9', 10' double bond) of different C40 carotenoids to produce C13 apocarotenoids, but CCD4 exhibited different cleavage activity for diversiform C40 carotenoids, among which β-carotene was considered to be the preferred substrate for CCD4[16]. It has been reported that the transcriptional level of CCD4 is a key factor affecting the total carotenoid content, which leads to color differences such as white, pale yellow, and dark orange in plant petals[17−19]. For example, the transcription level of IbCCD4 was negatively correlated with the total carotenoid content during flower coloration changes from yellowish-cream to white in Lilium brownie var. colchesteri[20]. More importantly, in chrysanthemum (Chrysanthemum morifolium) petals, there was a significant negative correlation between CmCCD4a expression level and total carotenoid content of petals, and white petals turned yellow due to carotenoid accumulation after the RNAi construct of CmCCD4a was introduced[21]. It is suggested that some carotenoid cleavage dioxygenase genes, rather than carotenoid biosynthesis genes, are largely responsible for carotenoid accumulation in many plant species.
Zinnia elegans is an annual herbaceous ornamental flower, which is widely used as summer and autumn flower in domestic and foreign gardens due to its diversity in flower types, color, and versatility of landscaping. Petal color of Zinnia elegans ranges widely from red to orange and yellow, which is mainly caused by different content of carotenoids and anthocyanins[22,23]. β-carotene is the main carotenoid in the petals, accounting for 65.89% and 82.25% of the total identified carotenoids in red and yellow petals, respectively[24]. What’s more, the cyclizing function of lycopene cyclase genes intimately associated with β-carotene synthesis, has been verified from Zinnia elegans[24] by bacterial pigment complementation system[25]. However, how the carotenoid cleavage dioxygenase genes, which are closely related to β-carotene degradation, participate in the carotenoid metabolism in the ray florets of Zinnia elegans and regulate the molecular mechanism of petal coloration in different color cultivars of Zinnia elegans is still unclear and requires further study.
In this study, the red, orange and yellow flower cultivars of Zinnia elegans, with different carotenoid content in the petals, were used as study materials. Three CCD genes (one CCD1 and two CCD4) from Zinnia elegans were cloned, and further characterized the cleavage ability of these genes on four kinds of C40 carotenoids using the bacterial pigment complementation system[25]. In addition, the carotenoid content and the expression patterns of three CCD genes were also analyzed among the red, orange and yellow cultivars. Finally, the subcellular localization of ZeCCDs was analyzed. This study will help to further explore the molecular mechanism of carotenoid metabolism in Zinnia elegans.
-
The experimental materials were three cultivars from Zinnia elegans 'Dreamland' series: Zinnia elegans 'Dreamland Red' (DRE), Zinnia elegans 'Dreamland Coral' (DC) and Zinnia elegans 'Dreamland Yellow' (DY). These plants were cultured in the climate room (photoperiod: 13 h light and 11 h dark, temperature cycle: 23 °C day and 18 °C night) of the College of Landscape Architecture, Zhejiang Agriculture and Forestry University (China). In accordance with previous studies[22−24], the developmental stages of petals were divided into four stages: the elongation stage (S1), the coloring stage (S2), the extension stage (S3) and the blooming stage (S4). The roots, stems, leaves, phyllary and petals of DY at S4 stage were collected for gene expression pattern analysis, and the outermost ray florets at four different developmental stages of DRE, DC and DY were collected to determine total carotenoid content and gene expression patterns. All samples were rapidly collected, frozen in liquid nitrogen, and subsequently stored at −80 °C until analysis.
Extraction and determination of total carotenoids
-
Total carotenoid content in petal was determined by spectrophotometric according to Lichtenthaler[26]. Fresh petals samples (0.2 g) stored at −80 °C were fully ground, mixed with 95% (v/v) ethanol solution, vortexed, and extracted for 15 min away from light, and centrifuged at 2,500× g for 10 min. The absorbance at 470 nm was measured with the supernatant, and the total carotenoid content of the petals at different developmental stages of different cultivars were calculated using Lichtenthaler's formula[26].
Molecular cloning of ZeCCDs
-
The petal samples of four developmental stages of DY were mixed, and the total RNA in the samples was extracted using UltraClean Polysaccharide and phenol Plant RNA Purification Kit (DNA free) (Novogene, Beijing, China). One μg of high-quality RNA was reverse transcribed into cDNA using the PrimeScript RT Master Mix (Perfect Real Time) Kit (TaKaRa, Dalian, China) for cDNA library construction and sequenced with DNBSEQ platform (Beijing Genomics Institute, Shenzhen, China). CCD1 and CCD4 proteins from Helianthus annuus (HaCCD1 XP_022023037.1; HaCCD4 XP_021977754.1) was used as bait, and the single gene sequence containing BLASTx (E value < 10−5) was searched in the NCBI non-redundant (Nr) protein database (
www.ncbi.nlm.nih.gov ). Finally, three CCDs (one CCD1 and two CCD4) were obtained, named ZeCCD1, ZeCCD4-1 and ZeCCD4-2, respectively.The full-length open reading frames (ORFs) of the three ZeCCDs amplified from the petals of DRE, DC, and DY were cloned into the vector pMAL-c5X (New England BioLabs, NEB) using homologous cloning technique (ClonExpress® II One Step Cloning Kit, Vazyme, Nanjing, China) with cloning primers (Supplemental Table S1) to identify gene sequence of each ZeCCD from different cultivars. The recombinant plasmids pMAL-ZeCCDs were transformed into E. coli Top10 competent cells and coated on Luria-Bertani (LB) solid medium containing 100 µg/mL ampicillin for screening. Single bacteria were selected for Polymerase Chain Reaction (PCR) positive identification and cultured overnight in LB liquid medium containing 100 µg/mL ampicillin. The plasmids were extracted and sequenced by Tsingke Biotechnology Co., Ltd (Hangzhou, China).
In silico analysis
-
Using the ProtParam tool (
http://web.expasy.org/protparam/ ), we analyzed the biochemical properties of the proteins encoded by ZeCCDs from DY, such as protein length, molecular weight (MW), isoelectric point (pI), instability index, aliphatic index and average hydrophilicity (GRAVY). The amino acid sequences of CCDs from Arabidopsis thaliana, Vitis vinifera, Triticum aestivum, Nicotiana tomentosiformis, Zea mays, Crocus sativus, Crocus angustifolius, Crocus chrysanthus, Rosa rugosa, Osmanthus fragrans, Lycium chinense, Helianthus annuus, Nicotiana tabacum and Solanum lycopersicum were downloaded from GenBank. The GenBank accession numbers of the downloaded sequences are shown in Supplemental Table S2. The sequence alignment was performed with ZeCCDs using MEGA X software[27], and a maximum likelihood phylogenetic tree was constructed based on the JTT matrix-based model (JTT) with 500 bootstrap replicates.Functional characterization of ZeCCDs
-
In order to characterize the cleavage function of ZeCCDs, the above pMAL-ZeCCD1, pMAL-ZeCCD4-1, pMAL-ZeCCD4-2, and the empty vector pMAL-c5X, were respectively co-transformed into E. coli Top10 competent cells[28−30] with four engineering plasmids (including pAC-β, β-carotene; pAC-ε, ε-carotene; pAC-ZEA, zeaxanthin; pAC-LYC, lycopene). The transformed E. coli cells were coated on LB solid medium containing 34 μg/mL chloramphenicol and 100 μg/mL ampicillin, and cultured overnight at 37 °C. Positive single colonies were inoculated in 2 mL LB liquid medium containing corresponding antibiotics. After 12 h of shaking culture at 37 °C, 1 mL of bacterial culture was expanded to 100 mL of medium containing the same antibiotics, and cultured again at 37 °C for 12 h. The bacterial cells were collected by centrifugation, and carotenoids were extracted with 1 mL of acetone, and shaken until the sediment was colorless. The supernatant was carefully aspirated after centrifugation at 2,500× g for 5 min, filtered through an organic filter membrane with a pore size of 0.22 μm, and stored in a brown glass sample bottle for subsequent carotenoid analysis.
Expression analysis
-
In order to quantitatively analyze the expression of ZeCCDs genes, AceQ® qPCR SYBR Green Master Mix (Vazyme, Nanjing, China) was used, and the primers for quantitative real-time PCR (qRT-PCR) are available in Supplemental Table S3. The reaction system and conditions were set according to our previous studies[24]. The qRT-PCR analyzes of the target genes were performed on the LightCycler® 480II (Roche Diagnostics, Germany) instrument. With ZeACT as the internal reference gene[22−24], Cq values were automatically calculated by LightCycler® 480II software and the relative expression of target genes was quantified by the 2−ΔCᴛ[31]. All qPCR reactions were set up with three biological replicates.
The total carotenoid content and the expression levels of ZeCCDs in the petals of different cultivars at different developmental stages were subjected to Person correlation analysis in SPSS 20.0 software (SPSS Inc., Chicago, IL, USA).
Subcellular localization analysis
-
The coding sequences of ZeCCDs from DY without stop codon were inserted into pCNHP-eYFP vector, so that each of the three ZeCCDs was fused with the eYFP protein to produce the 35S: ZeCCDs-eYFP fusion construct (primers used are shown in Supplemental Table S1). The recombinant plasmids verified by sequencing were transferred into Agrobacterium tumefaciens GV3101. The Agrobacterium tumefaciens GV3101 containing the target plasmid was respectively injected into 4-week-old tobacco (Nicotiana benthamiana) leaves. After 48 h of infection in the dark, the fluorescence signal was observed by confocal fluorescence microscope (ZEISS, Germany) with chloroplast autofluorescence as the marker.
-
Total carotenoids were extracted from fresh petals of three Zinnia elegans cultivars DRE, DC and DY (Fig. 1a) at four developmental stages (Fig. 1b) and quantified by spectrophotometry. The results showed that the total carotenoid content of DY petals was consistently higher than that of DRE and DC at different developmental stages, while that of DRE was always the lowest. Especially at S4, the total carotenoid content in DC petals was 2-fold higher than that of DRE, and the DY content was almost 10-fold higher than that of DRE (Fig. 1c).

Figure 1.
The total carotenoid content in the petals of Zinnia elegans. (a) Picture of flowers from DRE, DC and DY, and at (b) different developmental stages. (c) Total carotenoid content in petals from three cultivars and different developmental stages. These are mean ± standard errors, three biological replicates. Different letters indicate significant differences in the one-way ANOVA Duncan test, p < 0.05.
Molecular cloning of ZeCCDs
-
Carotenoid cleavage dioxygenase can breakdown the different sites of different carotenoids, cleave carotenoids, decrease the content of carotenoids in plants, and thus change the plant coloration. Therefore, the isolation, identification and characterization of ZeCCDs are essential for understanding carotenoid metabolism in the petals of Zinnia elegans. In the current study, the petal transcriptome was used for homology-based search to identify the carotenoid cleavage dioxygenase of Zinnia elegans[24]. Using HaCCD1 and HaCCD4 from Helianthus annuus as bait, three homologous genes with the highest sequence identity were identified in Zinnia elegans, which were named ZeCCD1, ZeCCD4-1 and ZeCCD4-2 respectively. Sequence analysis of ZeCCDs in DRE, DC and DY showed that sequence identity of ZeCCD1, ZeCCD4-1 and ZeCCD4-2 in three different cultivars were 100%, 98.21% and 99.78%, respectively. More importantly, we found four histidine residues binding to Fe2+ cofactors conserved in the amino acid sequences of ZeCCDs from three cultivars, which are essential for carotenoid cleavage dioxygenase activity, and their sites in the three cultivars are consistent (Supplemental Fig. S1). Subsequent analysis was carried out with ZeCCDs from DY, and coding regions of ZeCCD1, ZeCCD4-1 and ZeCCD4-2 were 1629 bp, 1734 bp and 1791 bp, respectively (Table 1).
Table 1. Biochemical properties of ZeCCDs.
Protein ORF length
(bp)Protein length
(aa)Molecular weight
(Da)Isoelectric
pointAliphatic
indexGrand average of
hydropathyInstability index ZeCCD1 1,629 542 61,102.98 5.85 78.54 −0.267 33.71 ZeCCD4-1 1,734 577 64,307.79 6.99 88.01 −0.109 31.13 ZeCCD4-2 1,791 596 65,784.95 6.05 79.13 −0.196 45.14 The amino acid sequences of the ZeCCDs were compared with protein sequences of Helianthus annuus, Osmanthus fragrans, Lycium chinense, Arabidopsis thaliana and other species. It was found that four histidine residues binding to Fe2+ cofactors, which are essential for carotenoid cleavage dioxygenase activity, were well conserved in the amino acid sequences of CCDs in the above different specie (Fig. 2a).
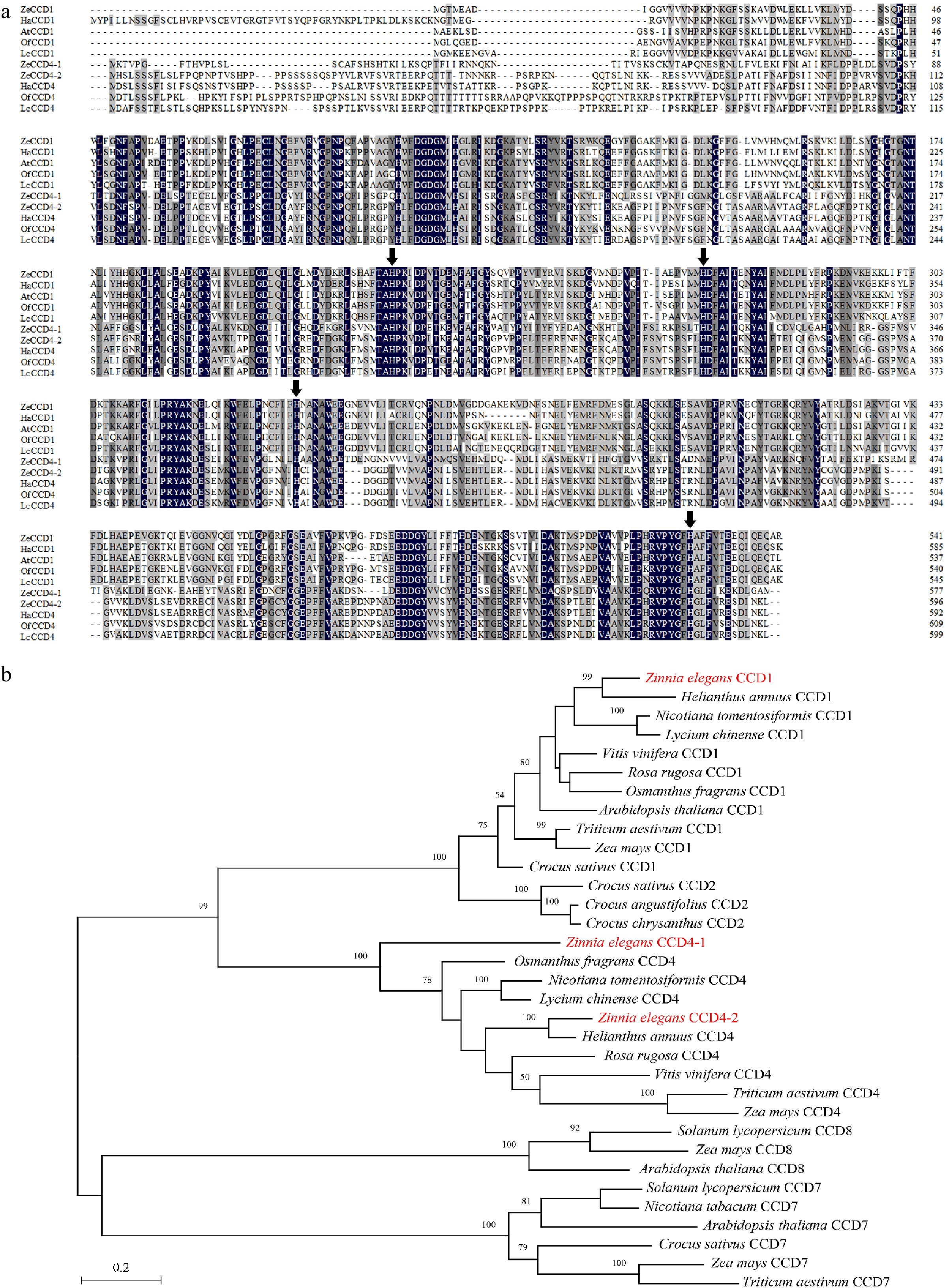
Figure 2.
Sequence alignment and phylogenetic analysis of CCDs proteins. (a) Alignment of the deduced amino acid sequences of CCDs homologs. Black represents the 100% homology level of the sequence, dark gray represents the ≥ 75% homology level, and light gray represents the ≥ 50% homology level. The black arrow refers to the histidine residue that binds to the Fe2+ cofactor. Sequences of functionally characterized HaCCD1 (XP_022018126.2) and HaCCD4 (CTP93722.1) from Helianthus annuus, OfCCD1 (AXQ60417.1) and OfCCD4 (ABY60887.1) from Osmanthus fragrans, LcCCD1 (AIY62758.1) and LcCCD4 (AIY62809.1) from Lycium chinense, AtCCD1 (NP_191911.1) from Arabidopsis thaliana. (b) Phylogenetic analysis of CCDs homologs from Zinnia elegans and other species (see Supplemental Table S2 for their GenBank accession numbers). The phylogenetic tree was built using MEGA X software with 500 bootstrap replicates based on the JTT matrix-based model (JTT). The scale bar represents the phylogenetic distance calculated based on the number of differences.
Phylogenetic analysis showed that CCDs were mainly divided into two major branches, one of which consisted of CCD1, CCD2 and CCD4. In this branch, CCD1 and CCD4 were clustered into a small branch (Fig. 2b).
Functional characterization of ZeCCDs
-
The pAC plasmids, which can provide the genes essential for efficient production of C40 carotenoids in E. coli, were used to establish a system based on the bacterial pigment complementation system[25] to verify the cleavage activity of three CCDs on four C40 carotenoid substrates. Due to the accumulation of carotenoids, cells transformed with different pAC plasmids showed diversiform coloration, which is a convenient system for characterizing enzymes that cleave carotenoids.
The results showed that all three ZeCCDs could make E. coli cells, which could produce β-carotene and other carotenoids, show different degrees of discoloration. Compared with pMAL-c5X, the cells co-transformed with four pAC plasmids (pAC-β, pAC-ε, pAC-ZEA, pAC-LYC) and pMAL-ZeCCD1 were the lightest in coloration and close to white, while co-transfected with pAC plasmids and pMAL-ZeCCD4 were lighter in coloration (Fig. 3). Carotenoids extracted from E. coli cells were examined by spectrophotometer. The results showed that the content of total carotenoids in E. coli, which transformed plasmids expressing ZeCCD1 with pAC, was significantly reduced compared to the control, and the content in cells transformed plasmids expressing ZeCCD4-1 and ZeCCD4-2 was also reduced and not significantly different between each other. The above results indicated that these three ZeCCDs in E. coli cells showed different cleavage activity for four carotenoids (β-carotene, ε-carotene, zeaxanthin, lycopene), among which ZeCCD1 had the strongest cleavage activity (Fig. 3).
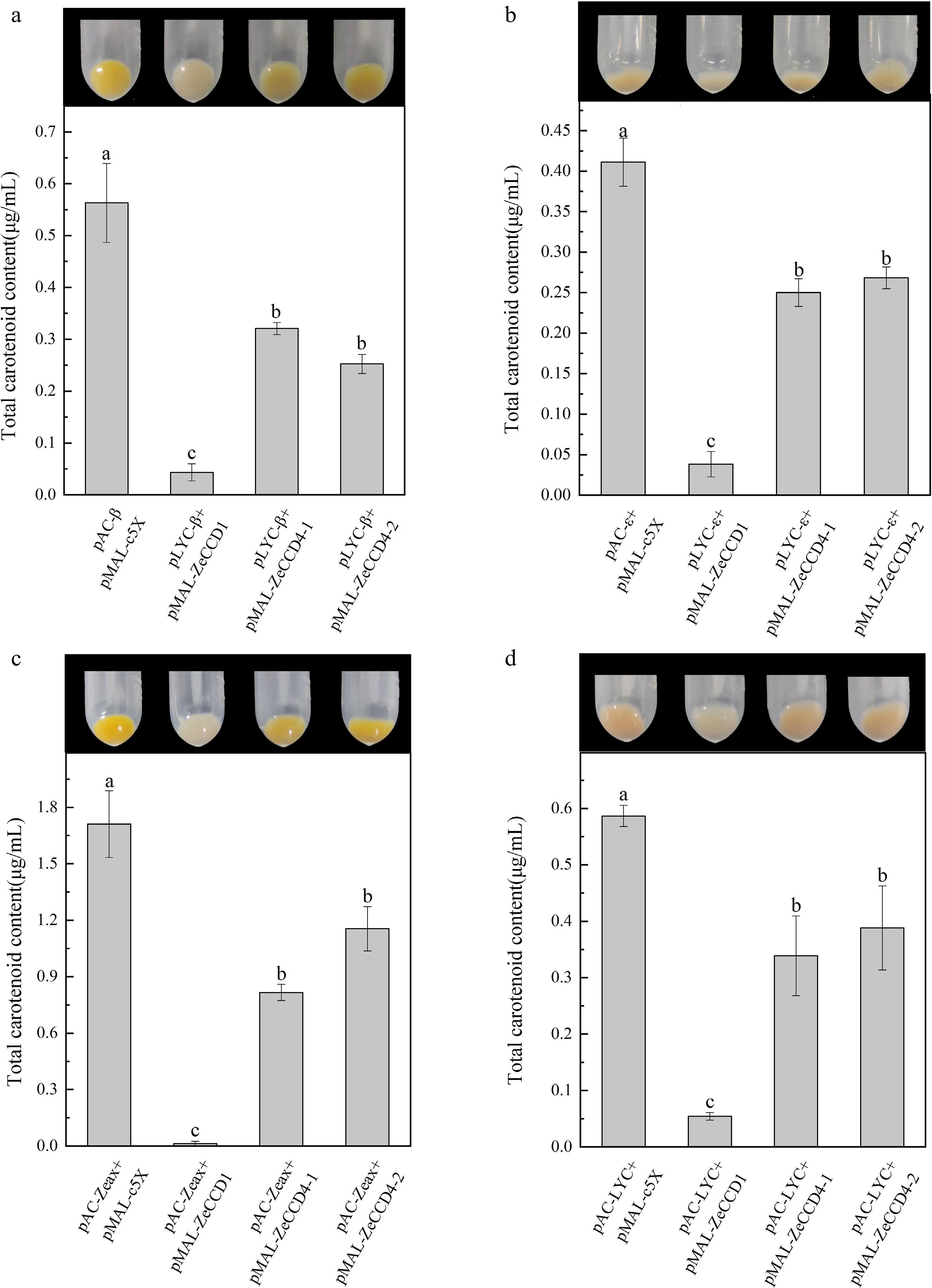
Figure 3.
Functional characterization of ZeCCDs proteins. The pMAL-ZeCCD1, pMAL-ZeCCD4-1, pMAL-ZeCCD4-2 and pMAL-c5X empty vectors were co-transformed into the engineered E. coli strains that were modified to accumulate (a) β-carotene, (b) ε-carotene, (c) zeaxanthin and (d) lycopene. These are mean ± standard errors, three biological replicates. Different letters indicate significant differences in the one-way ANOVA Duncan test, p < 0.05.
Expression patterns of ZeCCDs
-
The tissue specificity and expression patterns during flower development of ZeCCDs were analyzed by qPCR in order to further understand the possible biological functions of ZeCCDs. In DY, ZeCCD1 had the highest expression level in leaves and the lowest expression level in petals, while ZeCCD4-1 had the highest expression level in petals and stems. The expression level of ZeCCD4-2 was the highest in leaves and phyllaries (Fig. 4a). Significantly, the expression levels of ZeCCD1 and ZeCCD4-2 in petals were much lower than that of ZeCCD4-1.
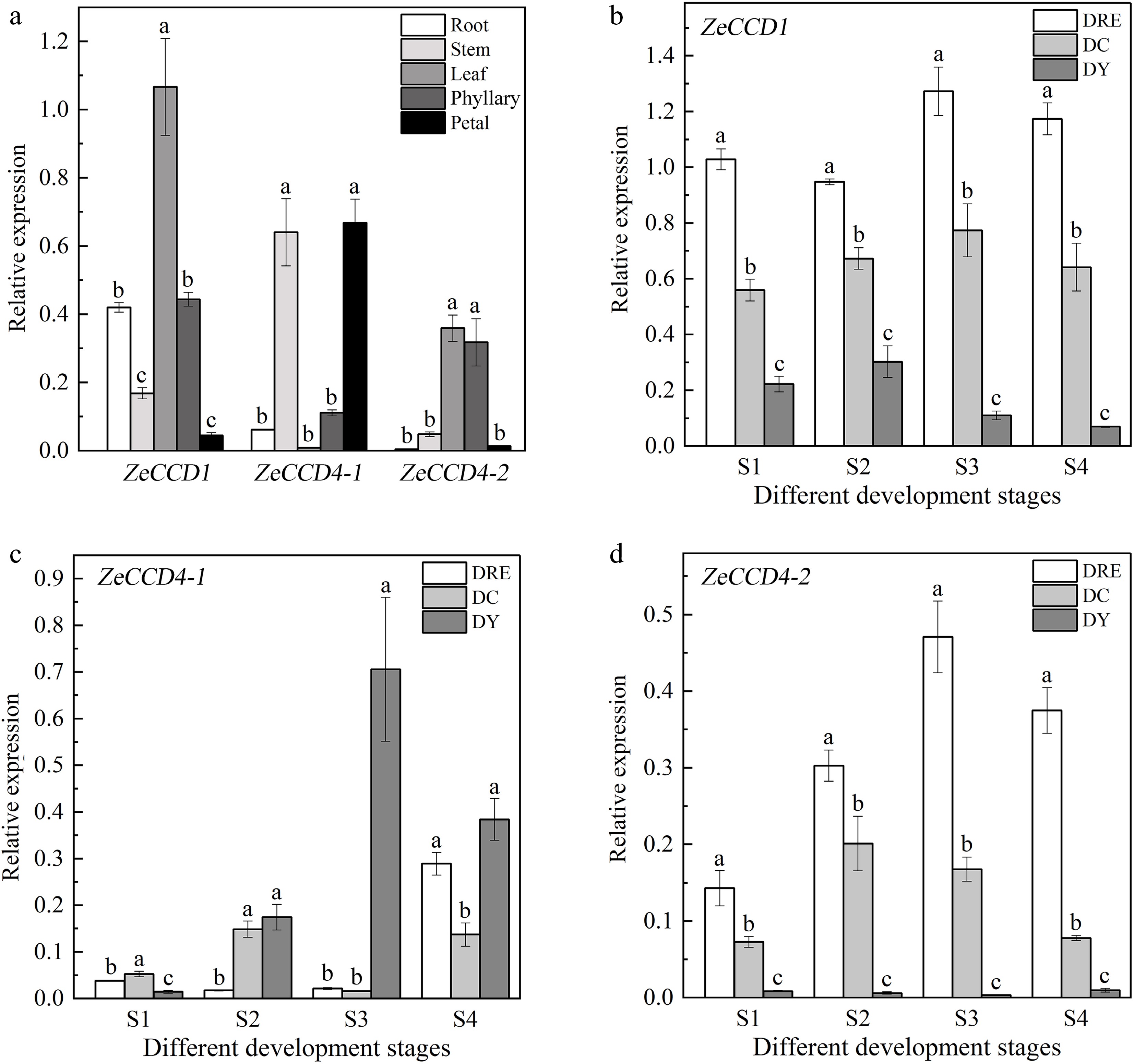
Figure 4.
The expression patterns of ZeCCDs. (a) Expression patterns of three genes in different tissues of DY. The expression patterns of (b) ZeCCD1, (c) ZeCCD4-1 and (d) ZeCCD4-2 in different cultivars and developmental stages. These are mean ± standard errors, three biological replicates. Different letters indicate significant differences between cultivars at the same developmental stage in the one-way ANOVA Duncan test, p < 0.05.
The results shows that ZeCCD1 has the highest expression level in DRE, followed by DC, and the lowest in DY at all four developmental stages (Fig. 4b). Compared with ZeCCD1, ZeCCD4-1 had the highest expression level in S2, S3 and S4 stages of DY. At the S1 stage, the expression level of ZeCCD4-1 in DC was higher than that in other cultivars (Fig. 4c). The expression pattern of ZeCCD4-2 was similar to that of ZeCCD1. During petal development, both ZeCCD4-2 and ZeCCD1 had the highest expression level in DRE and the lowest expression level in DY (Fig. 4d).
To further confirm the relationship between the expression levels of ZeCCDs and total carotenoid content, person correlation analysis showed that the expression levels of ZeCCD1 and ZeCCD4-2 were significantly negatively correlated with the total carotenoid content (Table 2).
Table 2. Correlation analysis of total carotenoid content and expression levels of ZeCCDs in petals from Zinnia elegans.
ZeCCD1 ZeCCD4-1 ZeCCD4-2 Total carotenoid content −0.915** 0.543 −0.757* '*' and '**' indicates significant negative correlation at p < 0.05 and p < 0.01 level between ZeCCDs expression levels and total carotenoid content respectively. Subcellular localization analysis
-
To determine the subcellular localization of ZeCCDs, we constructed a Pro35S::ZeCCDs-eYFP fusion structure and expressed it in epidermal cells of tobacco leaves, which was then visualized by using fluorescence microscopy. The results of transient expression in tobacco leaves demonstrated that the 35S::ZeCCD1-eYFP fusion protein was localized in the cytosol, while control protein (35S::eYFP) was spread throughout the cell. It is worth noting that the 35S::ZeCCD4-1-eYFP and 35S::ZeCCD4-2-eYFP fusion protein was co-localized with chloroplasts (Fig. 5).

Figure 5.
The subcellular localization of ZeCCDs. The fusion constructs (Pro35S::ZeCCDs-eYFP) were transiently transformed into Nicotiana benthamiana leaf epidermal cells, and observed by confocal fluorescence microscope (ZEISS, Germany) with chloroplast autofluorescence as the maker. For each construct, yellow (eYFP), red (chloroplasts), gray (bright-field), and merged are shown. Bars = 50 μm.
-
CCDs can change the type and content of carotenoids in plants by acting on different sites of carotenoids, especially CCD1 and CCD4 which are closely related to plant flower color. In order to explore the metabolic mechanism of carotenoids in the petals of Zinnia elegans, three CCDs, including one CCD1 and two CCD4, were identified and isolated from Zinnia elegans by using homology-based search on petal transcriptome, and named ZeCCD1, ZeCCD4-1, and ZeCCD4-2 (Table 1). This result was similar to that of Crocus sativus, which had two isoforms of CCD4[32]. Noteworthy, four histidine residues essential for carotenoid cleavage activity were conserved in these proteins. Four histidine residues together with a divalent iron center form a 4-His-ligated iron center, which is required for substrate recognition and correct positioning of substrates in the active site[33], suggesting that four histidine residues are necessary for CCD to perform its cleavage function. The specific structure of the 4-His-ligated iron center is conserved in the CCD family, which requires the amino acid sequences of CCDs to be evolutionarily conserved[34].
ZeCCDs can cleave diversiform substrates in E. coli
-
The pigment complementation system in E. coli showed that ZeCCD1, ZeCCD4-1 and ZeCCD4-2 cleaved four carotenoids, including β-carotene (Fig. 3), which is the most abundant carotenoids in the petals of Zinnia elegans[24]. This is similar to the cleavage activity of CCD1 from other plants with extensive affinity for C40 carotenoids. For example, LeCCD1 from tomato can catalyze the cleavage of several carotenoids such as lycopene, β-carotene, ε-carotene and zeaxanthin[10]. DoCCD1 from Dendrobium officinale and RdCCD1 from Rosa damascene also catalyze the cleavage of lycopene, β-carotene and zeaxanthin[35,36]. Similarly, two ZeCCD4 have catalytic activity for all four carotenoids, consistent with the reported cleavage activity of CCD4 from Camellia sinensis in E. coli[37]. In contrast, GmCCD4 from soybeans (Glycine max) can only specifically catalyze the cleavage of β-carotene and has affinity or cleavage activity for lycopene[38]. Although CCD4 enzymes from different plant species exhibit differences in substrate specificity and regioselectivity, it is generally accepted that β-carotene is recognized as the most favorable substrate[5]. Moreover, ZeCCD1 seemed to have higher cleavage activity for diversiform C40 carotenoids, and only a small quantity of carotenoids could be extracted from E. coli co-transfected with different pAC plasmids, compared with ZeCCD4-1 and ZeCCD4-2 (Fig. 3). This could be partly attributed to the differences of cleavage site and substrate specificity. The CCD1 enzymes are characterized by their extensive substrate and double-bond specificity, and they cleave different linear and cyclic C40 carotenoids and apocarotenoid carotenoids at 9, 10 and 9′, 10′ positions[36]. Accordingly, we surmise that ZeCCD1 catalyzes the cleavage of both ends of carotenoids in two steps, from C40 carotenoids via C27 apocarotenoids to C13/C14 apocarotenoid end-products in E. coli, with the second reaction proceeding in a way that may promote the first step. This hypothesis is supported by CCD1 breakdown both C40 carotenoids and C27 apocarotenoids[11]. In contrast, ZeCCD4-1 and ZeCCD4-2 may catalyze single cleavage of C40 carotenoids at 9, 10 (9′, 10′), which is similar to research on CCD4 from potato (Solanum tuberosum)[39], sweet orange (Citrus sinensis)[40] and Satsuma mandarin (Citrus unshiu)[41]. In Citrus species, this cleavage led to the production of C30 apocarotenoid pigments, such as β-citraurinene and β-citraurin, which are essential for peel pigmentation in citrus fruits[42]. Although, it has been reported that CCD4 catalyzes double cleavage of C40 carotenoids at 7, 8 (7′, 8′), they are well correlated with the biosynthesis of some specific substances in plants, such as crocins and bixin[43,44], which contain the conjugated polyene system of carotenoids. Therefore, it is necessary to study CCD, which will provide a better understanding of the original biological functions of these enzymes and further determine their hypothetical roles in material metabolism.
ZeCCD4-2 participates in the cleavage of carotenoids in petals of Zinnia elegans
-
We analyzed the expression patterns of ZeCCDs at four developmental stages in the petals of different Zinnia elegans cultivars (Fig. 4). The expression levels of ZeCCD1 and ZeCCD4-2 were significantly negatively correlated with total carotenoid content in petals of Zinnia elegans (Table 2). In general, the expression levels of CCDs are negatively correlated with carotenoid content due to the cleavage function of CCDs on carotenoids. The higher expression level of SiCCD1 at the grain maturity stage was considered to be the primary reason for the lower carotenoid content in white cultivars of Setaria italica[45]. What’s more, the expression level of CmCCD4a in chrysanthemum, which belongs to Asteraceae family, also showed a significant negative correlation with the total carotenoid content. After the introduction of an RNAi construct of CmCCD4a in the white cultivar, the petals turned yellow[8]. The results of the subcellular localization assay showed that ZeCCD1 was localized in the cytosol while ZeCCD4-1 and ZeCCD4-2 were localized in the chloroplasts (Fig. 5). This is consistent with previous studies in Prunus mume that PmCCD1 and PmCCD1-like-c were localized in the cytoplasm, and PmCCD4 was localized in the plastid[46]. In plants, the enzymes involved in carotenoid biosynthesis are located in the plastids[35]. At the same time, carotenoids, as the hypothetical substrate of CCDs, are mostly stored in plastids due to their hydrophobicity, while CCD1 enzyme, due to the lack of plastid localization peptides, is the only member of the CCDs family located in the cytoplasm. It cannot directly obtain the C40 carotenoid substrate in the plastid, but acts on the C27 apocarotenoid, the cleavage product of the C40 carotenoid that is transported to the cytoplasm[47]. Unlike the CCD1 enzymes located in the cytoplasm, most of the CCD4 enzymes are located in the plastids[13], suggesting that the CCD4 enzymes can contact its carotenoid substrate. For example, in Satsuma mandarin (Citrus unshiu), CitCCD4 is located in the plastids and can cleave the carotenoid substrates β-cryptoxanthin and zeaxanthin stored in the plastids to produce β-citraurin[45]. Therefore, it was presumed that ZeCCD4-2 might be a critical gene affecting carotenoid metabolism in the petals of Zinnia elegans. Our work preliminarily revealed the function of carotenoid cleavage dioxygenase genes in carotenoid accumulation in Zinnia elegans, and laid a foundation for further exploring the molecular mechanism of carotenoid metabolism in Zinnia elegans.
Carotenoid accumulation in plants is not only affected by carotenoid metabolic genes, transcription factors can also be involved in the regulation of carotenoid accumulation. For example, the promoter region of CCD2 in Crocus sativus contained a MYB-binding region, and the expression level of CstCCD4b was significantly increased in transgenic lines for the MYB transcription factors CstMYB1 and CstMYB1R2[48]. Another study showed that in Chrysanthemum morifolium, two MADS box-containing transcription factors, APETALA3 (AP3) and ULT1 Interacting Factor 1 (UIF1), could be connected by PISTILLATA (PI) to form the CmAP3-CmPI-CmUIF1 protein complex, regulated carotenoid metabolism by directly activating the expression of CmCCD4a-2[49]. At the same time, there were many GATA binding sites in the promoter region of CmCCD4a-5, and the GATA family transcription factor CmGATA4 of Chrysanthemum morifolium could bind directly to the promoter of CmCCD4a-5 and inhibit its activity[50]. The WRKY transcription factor OfWRKY3 and the ethylene responsive factor (ERF) transcription factor OfERF61 of Osmanthus fragrans were found to bind directly to the promoter of OfCCD4 and significantly promote the expression of OfCCD4[51]. Transcription factors related to the regulation of CCDs in Zinnia elegans have not been determined, so more studies should be conducted to identify and reveal the function of up-stream potential transcription factors, which could directly regulate the expression of CCDs in Zinnia elegans.
-
Three CCDs were isolated from Zinnia elegans. These three ZeCCDs could cleave β-carotene, ε-carotene, zeaxanthin and lycopene. The expression levels of ZeCCD1 and ZeCCD4-2 were significantly negatively correlated with the total carotenoid content in petals from different cultivars. Subcellular localization analysis showed that ZeCCD1 was localized in the cytosol, while ZeCCD4-1 and ZeCCD4-2 were localized in the chloroplasts. Based on the above results, it is speculated that ZeCCD4-2 plays an more important role in carotenoid cleavage in Zinnia elegans and affects the petal coloration of ray florets.
-
The authors confirm contribution to the paper as follows: literature collation: Qing H; experimental design and implementation: Qing H, Fu J, Zhang C; data analysis and visualization: Qing H, Liu X, Chen J, Li L, Qian J; writing and polishing: Qing H, Liu X; resources and project administration: Fu J, Zhang C; review and editing: Fu J, Zhang C. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
We are grateful to Drs E. Gantt and F.X. Cunningham for the E. coli functional complementation system and Dr. Xing-Qi Huang for the gift of plasmids pAC-LYC, pAC-β, pAC-ε, pAC-Zea and pMAL-C5X. This research was supported by Zhejiang Provincial Natural Science Foundation of China (Grant No. LY24C150001) and College Students Research Training Program of Zhejiang Agriculture and Forestry University (No. 2023KX044).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Hongsheng Qing, Xinyue Liu
- Supplemental Table S1 Primer sequences used in this research.
- Supplemental Table S2 The GenBank accession of carotenoid cleavage dioxygenases sequences from other species.
- Supplemental Table S3 The primer sequences for qPCR used in this study.
- Supplemental Fig. S1 Protein sequence alignment analysis of CCD1, CCD4-1 and CCD4-2 in Zinnia elegans of different cultivars. The black arrow refers to the histidine residue that binds to the Fe2+ cofactor.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Qing H, Liu X, Chen J, Li L, Qian J, et al. 2024. Carotenoid cleavage dioxygenase catalyzes carotenoid degradation and regulates carotenoid accumulation and petal coloration in Zinnia elegans. Ornamental Plant Research 4: e005 doi: 10.48130/opr-0024-0003 

Carotenoid cleavage dioxygenase catalyzes carotenoid degradation and regulates carotenoid accumulation and petal coloration in Zinnia elegans
- Received: 26 September 2023
- Revised: 28 December 2023
- Accepted: 05 January 2024
- Published online: 27 February 2024
Abstract: Carotenoids are one of the important coloring substances in the ray florets of Zinnia elegans. Carotenoid cleavage dioxygenase (CCD) directly affects carotenoid contents by cleaving the double bond of carotenoids, thereby affecting the flower color. However, few reports have elucidated its function in Zinnia elegans. In the present study, through transcriptome homology searches, three ZeCCDs, ZeCCD1, ZeCCD4-1, and ZeCCD4-2 were screened from Zinnia elegans. The function of ZeCCDs was verified by bacterial pigment complementation system, and it revealed that all of the three ZeCCDs had the ability to cleave β-carotene and other carotenoids (ε-carotene, zeaxanthin and lycopene). Meanwhile, the expression levels of ZeCCD1 and ZeCCD4-2 in the petals of different cultivars were significantly negatively correlated with the carotenoid content. Furthermore, compared with the cytoplasmic localization of ZeCCD1, ZeCCD4-2 is localized in the plastid. Our results indicate that ZeCCD4-2 is the key gene responsible for the differential accumulation of carotenoids in the petals of different Zinnia elegans cultivars. This study lays a foundation for further research on the molecular mechanism of petal coloration and carotenoid metabolism of Zinnia elegans.
-
Key words:
- Zinnia elegans /
- Carotenoid /
- Petal color /
- Carotenoid cleavage dioxygenase /
- β-carotene